Introduction
Ovarian cancer is the fifth most common cause of
cancer- associated mortality in females worldwide, and 90% of cases
of ovarian cancer are epithelial-derived malignancies (1). Although ovarian cancer has an improved
response to chemotherapy, the prognosis of patients with ovarian
cancer is not ideal clinically, with very low survival rate
(2,3).
Studies have indicated that clinical pathological parameters,
including age, tumor stage, patient's body function and residual
tumor volume, are independent predictors of the prognosis of
ovarian cancer (1–3); however, for patients with similar status
and treatments, their survival rates were not the same (4). Therefore, it is urgent to identify the
factors relating to the occurrence and development of ovarian
cancer to provide a scientific basis for the development of
effective treatment strategies of ovarian cancer and enhance the
survival rate of patients with ovarian cancer.
B-cell lymphoma 2 (Bcl-2) protein family includes a
number of members, including anti-apoptosis factors Bcl2-like
(Bcl-2L), Bcl-extra-large (xL) and Mcl1, and the pro-apoptosis
factor containing the structure of BH3-only (5,6), and
includes Bcl2-2L 11, p53 upregulated modulator of apoptosis (PUMA),
BH3 interacting domain death agonist, Bcl-2 associated agonist of
cell death, Bcl-2 and Bcl-2 modifying factor (BMF) (7,8). Similar
to PUMA, BMF serves a role in initiating cell apoptosis by binding
to the anti-apoptosis factors B-cell lymphoma (Bcl)2, Bcl-xL and
Bcl-w proteins (9). During the
occurrence of a variety of tumor types including lymphoma (10), colorectal cancer (11) and breast cancer (12), BMF serves a role of tumor suppressor
gene.
microRNA (miRNA) is a class of endogenous non-coding
RNAs ~22 nucleotides in size (12).
miRNA can regulate the translation of its target gene mRNA or
degrade mRNA by regulating the post-transcriptional level, which is
involved in the regulation of numerous biological functions,
including cell proliferation, migration, invasion and apoptosis
(13–15). miR-221 is a miRNA that is closely
associated with tumorigenesis and serves a tumor-promoting role in
a variety of tumor types, including breast (16), pancreatic (17), prostate (18,19) and
non-small cell lung cancer (20), and
miR-221 serves a role as a tumor suppressor gene (21). A number of studies have demonstrated
that the expression of miR-221 in tumor tissue or peripheral blood
of patients with ovarian cancer was significantly increased,
indicating that miR-221 may serve a role of a cancer-promoting gene
in the occurrence of ovarian cancer (21,22).
MicroRNA.org online (22) predicted that BMF was the possible
target gene of miR-221. In the present study, the expression levels
of miR-221 and BMF in ovarian cancer tissues were detected by
reverse transcription-quantitative polymerase chain reaction
(RT-qPCR), and transfected the siRNA of anti-miR-221 or BMF into
the ovarian cancer cell line SKOV3 to observe the effect of miR-221
on SKOV3 cell proliferation and apoptosis, and validate the
association between miR-221 and target gene BMF and the regulatory
effect on the proliferation and apoptosis of SKOV3 cells, to
identify a novel treatment target for the clinical treatment of
ovarian cancer.
Materials and methods
Clinical data
A total of 30 patients with ovarian cancer who
received their treatment in The Department of Obstetrics and
Gynecology in The First Affiliated Hospital of Fujian Medical
University from February 2013 to December 2014 were collected. None
of the patients underwent chemotherapy and radiotherapy prior to
surgery. The patients were aged from 33–72 years, with a mean age
of 51.3±21.4 years. The tumor specimens and normal para-carcinoma
tissue specimens were collected by surgery. Tumor tissue specimens
were pathologically confirmed as ovarian cancer, whilst the normal
tissues were confirmed as having no cancerous tissue invasion by
two pathologists from the First Affiliated Hospital of Fujian
Medical University. Specimens were preserved in liquid nitrogen
within 10 min following acquisition. All specimens were collected
with the patient's informed consent and approved by the Ethics
Committee of the First Affiliated Hospital of Fujian Medical
University.
Cells and main reagents
Ovarian cancer cell line SKOV3 cells were purchased
from the Institute of Basic Medical Sciences, Chinese Academy of
Medical Sciences in Beijing, China. Dulbecco's modified Eagle's
medium (DMEM) and fetal bovine serum (FBS) were purchased from
Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Horseradish peroxidase-labeled goat anti-mouse IgG secondary
antibody (catalog no. SA00001) was purchased from OriGene
Technologies, Inc. (Beijing, China). Annexin V/PI staining kits and
caspase-3 enzyme activity test kits were purchased from Beyotime
Institute of Biotechnology (Jiangsu, China). TRIzol® and
transfection reagent Lipofectamine® 2000 were purchased
from Invitrogen (Thermo Fisher Scientific, Inc.). Mimic NC, miR-221
mimic, anti-miR-NC and anti-miR-221 were purchased from Guangzhou
Ribobio Co., Ltd. (Guangzhou, China). Luciferase activity detection
kits were purchased from Promega Corporation (Madison, WI, USA).
Mouse anti-human β-actin (catalog no. MS123A1) and BMF antibodies
(catalog no. A21422), and si-NC (catalog no. A24185) and si-BMF
(catalog no. A36521) were purchased from Santa Cruz Biotechnology,
Inc. (Dallas, TX, USA). The cell proliferation detection kit
(Click-iT EdU Alexa Fluor 488 Flow Cytometry Assay Kit) was
purchased from Molecular Probes (Thermo Fisher Scientific, Inc.).
Overexpression plasmid phosphorylated internal ribosome entry site
2 (pIRES2) was purchased from Beijing BioVector (Beijing, China;
http://www.biovector.net/).
Detection of gene expression by
RT-qPCR
A total of 1 ml TRIzol was added to every 20 mg of
tissue or every 3×106 cells. Following full lysis,
chloroform was added for extraction. Following separation, the RNA
supernatant was transferred to a new Eppendorf (EP)
Tube®, precipitated with isopropanol and washed with 70%
ethanol, and then dissolved in diethyl pyrocarbonate water to
extract RNA. The RNA was reverse transcribed into cDNA using the
PrimeScript™ RT reagent Kit (Perfect Real Time, Takara,
Japan; URL: http://www.takara-bio.com/), then using cDNA as a
template, qPCR was performed for detection of gene expression. The
reverse transcription reaction system included 0.5 µl oligdT Primer
(50 µM), 0.5 µl Random 6-mers (100 µM), 0.5 µl PrimeScript RT
Enzyme Mix, 1.0 µg RNA, 2 µl 5X PrimeScript Buffer, then RNase-free
water was added to a total volume of 10.0 µl. The reverse
transcription reaction condition: 37°C for 15 min; and 85°C for 5
sec. The RT-qPCR reaction system included SYBR Fast qPCR Mix 10.0
µl, 0.8 µl forward Primer (10 µM), 0.8 µl Reverse Primer (10 µM),
2.0 µl cDNA and 6.4 µl RNase Free dH2O. The qPCR
reaction conditions: Initial denaturation 10 min at 95°C;
denaturation 10 sec at 95°C; annealing 20 sec at 60°C; and
extension 15 sec at 72°C; 40 cycles, and finally the gene
expression was detected in ABI ViiA 7 real-time qPCR instrument.
The 2−∆∆Cq method was used to compare gene expression
between clinical samples (23).
The primer sequence used were: miR-221P forward,
5′-GTTGGTGGGAGCTACATTGTCTGC-3′, and reverse,
5′-GTGTCGTGGACTCGGCAATTC-3′; U6P forward,
5′-ATTGGAACGATACAGAGAAGATT-3′, and reverse,
5′-GGAACGCTTCACGAATTTG-3′; BMFP forward,
5′-TTTATGGCAATGCTGGCTATCG-3′, and reverse,
5′-GCAATCTGTACCTCTGCTTGATG-3′; and β-actin forward,
5′-GAACCCTAAGGCCAAC-3′, and reverse, 5′-TGTCACGCACGATTTCC-3′.
Construction of BMF overexpression
plasmids
The fragments of the coding region of the BMF gene
were amplified and the size of target fragments was identified by
gel electrophoresis (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany). SDS PAGE gel (9%) was prepared as follows: 50 g of
protein sample was loaded for 3 h and then placed into the electric
transducers, in which the buffer was added to the transmembrane for
1.5 h and the target protein was transferred onto nitrocellulose
membranes. PCR product was recovered from gel, and following
dual-enzyme digestion they were ligated into pIRES2 plasmids, then
following transformation the positive strains were selected, and
strains were amplified and cultured to extract the recombinant
plasmid containing the target fragments. Sequencing was performed
to confirm that the target fragment of BMF gene was inserted
correctly, and termed as pIRES2-BMF; in addition, the blank vector
pIRES2-blank was used as control.
SKOV3 cell culture and
transfection
SKOV3 cells were cultured in DMEM containing 10% FBS
and 1% streptomycin, and cultured in a cell incubator at 37°C and
an atmosphere containing 5% CO2. Medium was replaced
once every two days. Cells were subcultured according to the ratio
of 1:4; cells in good growth condition at the logarithmic period
were taken to carry out experiments. One day prior to transfection,
cells were inoculated in a 10 cm culture dish, to ensure that the
cell density could be ≤50–60% on the day of transfection. A total
of 2 µl Lipofectamine 2000, 30 nmol anti-miR-221p, 20 nmol
pIRES2-BMF, and 20 nmol si-BMF were diluted into 50 ml Opti-MEM
(Invitrogen; Thermo Fisher Scientific, Inc.) and added to the
cells. Following incubation at room temperature for 5 min, the
diluted Lipofectamine 2000 and all transfectants were gently mixed
well and placed at room temperature for 20 min, then according to
difference of the grouping, the transfected mixture was added to
cells free of serum and double antibody culture medium (Invitrogen;
Thermo Fisher Scientific, Inc.), incubated in 37°C incubator for 6
h, and then the medium was changed to fresh complete DMEM medium
containing serum (Invitrogen; Thermo Fisher Scientific, Inc.) and
double antibody, to continue to culture 48 h, and cells were
collected for detection. The transfected cells were divided into
four groups: Anti-miR-NC, anti-miR-221, pIRES2-blank and
pIRES2-BMF.
Cell proliferation activity detected
by MTT assay
The SKOV3 cells transfected with anti-miR-NC,
anti-miR-221, pIRES2-blank and pIRES2-BMF were inoculated into
96-well culture plates. Following incubation for 24, 48 and 72 h at
37°C following transfection, 10 µl MTT solution (Qiagen GmbH,
Hilden, Germany) was added to each well for 4 h at 37°C, the
supernatant was removed, and 150 µl dimethyl sulfoxide solution was
added, following vibration for 10 min at low speed, the absorbance
A450 at the wavelength of 450 nm was measured.
Detection of cell proliferation by EdU
staining flow cytometry
Cells in all transfection groups were re-suspended
using the complete medium (Qiagen GmbH). The kit used was the
Click-iT® EdU flow cytometry cell proliferation assay
(Qiagen GmbH, Hilden, Germany). Following incubation for 120 min
with 10 µM EdU at 37°C, cells were cultured for 48 h at 37°C.
Following digestion with trypsin, cells were centrifuged at 1,000 ×
g for 5 min at 37°C and washed once with 1% bovine serum albumin
(BSA) in PBS. The cells were fixed with 100 µl Click-iT fixative
for 15 min at room temperature, centrifuged at 1,000 × g for 5 min
at 37°C and washed once with 1% BSA in PBS. A total of 100 µl
permeabilization and washing reagent (0.2%) was added for
permeabilization 15 min at room temperature, then 500 µl of
reaction solution containing PBS (496 ml), CuSO4 (1 mM),
Alexa Fluor 488 (10 µM), buffer additive (4 ml, component F; Qiagen
GmbH, Hilden, Germany) was added to incubate for 30 min in the
darkness at room temperature. Following this, 3 ml permeabilization
and wash reagent was added and the mixture was centrifuged at 1,000
× g for 5 min at 37°C and washed with PBS once. The cells were then
re-suspended with 500 µl permeabilization and wash reagent, and
cell proliferation was detected by the Beckman Coulter FC 500
MCL/MPL flow cytometer.
Detection of apoptosis
Cells in all transfection groups were digested and
harvested by trypsin, centrifuged for 5 min at 1,000 × g 37°C and
washed once using 1% BSA PBS. Cells were re-suspended in 100 µl
binding buffer (part of the apoptosis detection kit), then cells
were incubated with a cell apoptosis detection kit [10 µl (15 M)
Annexin V dye (FITC) and 5 µl (15 M) PI dye; Beyotime Institute of
Biotechnology, Haimen, China] in the dark for 15 min at 37°C. Cell
apoptosis was detected with a Beckman Coulter FC 500 MCL/MPL flow
cytometer and analyzed using FCS Express 3.0 software (DeNovo
software, Glendale, CA, USA).
Detection of Caspase-3 activity
The pNA standard was prepared according to the
protocols of the Caspase-3 activity assay kit, and A405 was
measured and a standard curve was plotted. Cells in all treatment
groups were digested by lysis solution on ice (100 µl lysate for
5×106 cells), then the supernatant was transferred to a
new pre-cooled EP Tube for standby. The buffer, test article,
Ac-DEVD-pNA (Qiagen GmbH, Hilden, Germany) were added to the
96-well plate successively, and incubated at 37°C for 120 min. When
an evident color change was determined, the absorbance value A405
was measured immediately using a microplate reader. The absorbance
value A405 reflected the activity of Caspase-3 of the test
article.
Dual-luciferase reporter gene
assay
MicroRNA.org online prediction
indicated that the sites 3673–3690 on the 3′-untranslated region
(UTR) of BMF mRNA were possible binding sites for miR-221. The DNA
fragments containing the sites and containing the site mutant were
synthesized in vitro, then cloned to dual luciferase gene
reporter vector pMIR (Thermo Fisher Scientific, Inc.). The pMIR
vector and miR-221 mimic (mimic NC, anti-miR-NC and anti-miR-221)
were co-transfected into SKOV3 cells using
Lipofectamine® 2000 transfection reagent (Thermo Fisher
Scientific, Inc.). Following 48 h of culture at 37°C, the
luciferase activity was detected by dual luciferase assay kit. The
red firefly luciferase signal was utilized as a normalization
control.
Detection of protein expression by
western blot analysis
Tissues or cells were lysed by
radioimmunoprecipitation assay lysate (Thermo Fisher Scientific,
Inc.). Supernatant was removed and the protein concentration was
determined with a bicinchoninic acid assay reagent (Pierce; Thermo
Fisher Scientific, Inc.). A total of 40 µg solution was loaded onto
SDS-PAGE for electrophoresis separation (10% separation gel and 5%
spacer gel), transferred to polyvinylidene fluoride (PVDF)
membranes. The membranes were blocked with PBS with Tween 20 (PBST,
Sigma-Aldrich; Merck KGaA) containing 5% BSA for 1 h at 37°C,
followed by incubation with primary antibody overnight at 4°C (the
dilution ratios of BMF and β-actin were 1:2,000 and 1:4,000,
respectively). The next day, the membranes were washed three times
using 0.1% tris-buffered saline and Tween 20 (TBST), 5 min each
time, then HRP-labeled secondary antibody (dilution ratio of
1:10,000) was added and incubated for 1 h at room temperature, then
washed by 0.1% TBST ×3. The bands of PVDF membrane were developed
by Supersignal West Femto HRP sensitive chemiluminescent substrate
(Sigma-Aldrich; Merck KGaA), using β-actin as the internal
reference substance.
Statistical analysis
Statistical analysis was performed by SPSS 18.0
(SPSS, Inc., Chicago, IL, USA). Measurement data are expressed as
mean ± standard deviation, the measurement data between the two
groups were compared using Student's t-test; the measurement data
between the multiple groups were compared first using one-way
analysis of variance, then using the Bonferroni method for
comparison. The expression levels of miR-221 and BMF mRNA in
tissues were compared by Mann-Whitney U test. P<0.05 was
considered to indicate a statistically significant difference.
Results
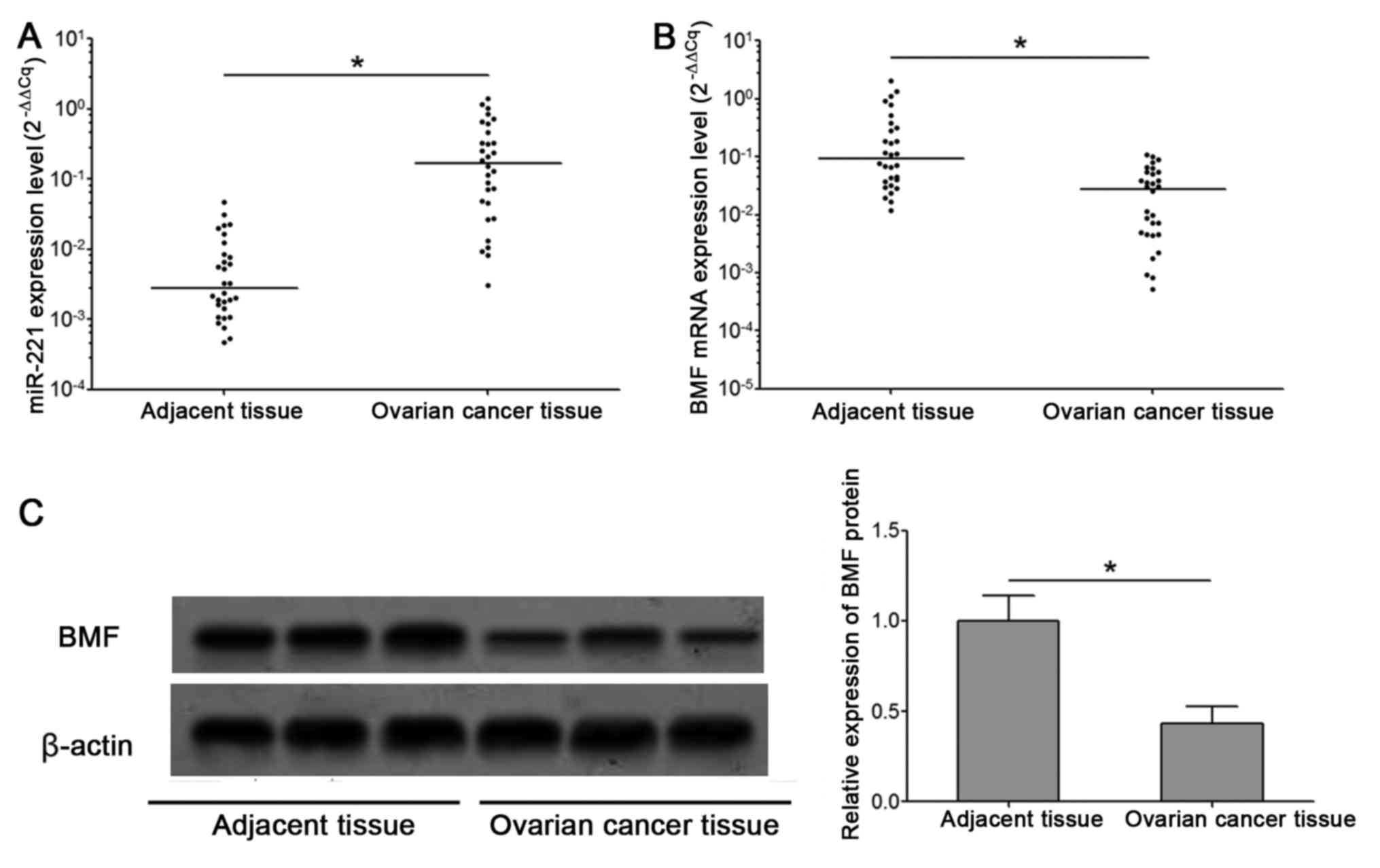
Abnormal change in the expression
levels of miR-221 and BMF in ovarian cancer tissues
The results of RT-qPCR demonstrated that, compared
with the para-cancer tissues, the expression of miR-221 in tumor
tissues was significantly increased (Mann-Whitney U=48, P<0.001;
Fig. 1A), and the expression of BMF
mRNA was significantly decreased in patients with ovarian cancer
(Mann-Whitney U=162, P<0.001; Fig.
1B). Western blot analysis indicated that the expression of BMF
proteins in ovarian cancer tissue was significantly lower than that
in para-cancer tissues (P<0.001). The representative test
results were depicted in Fig. 1C.
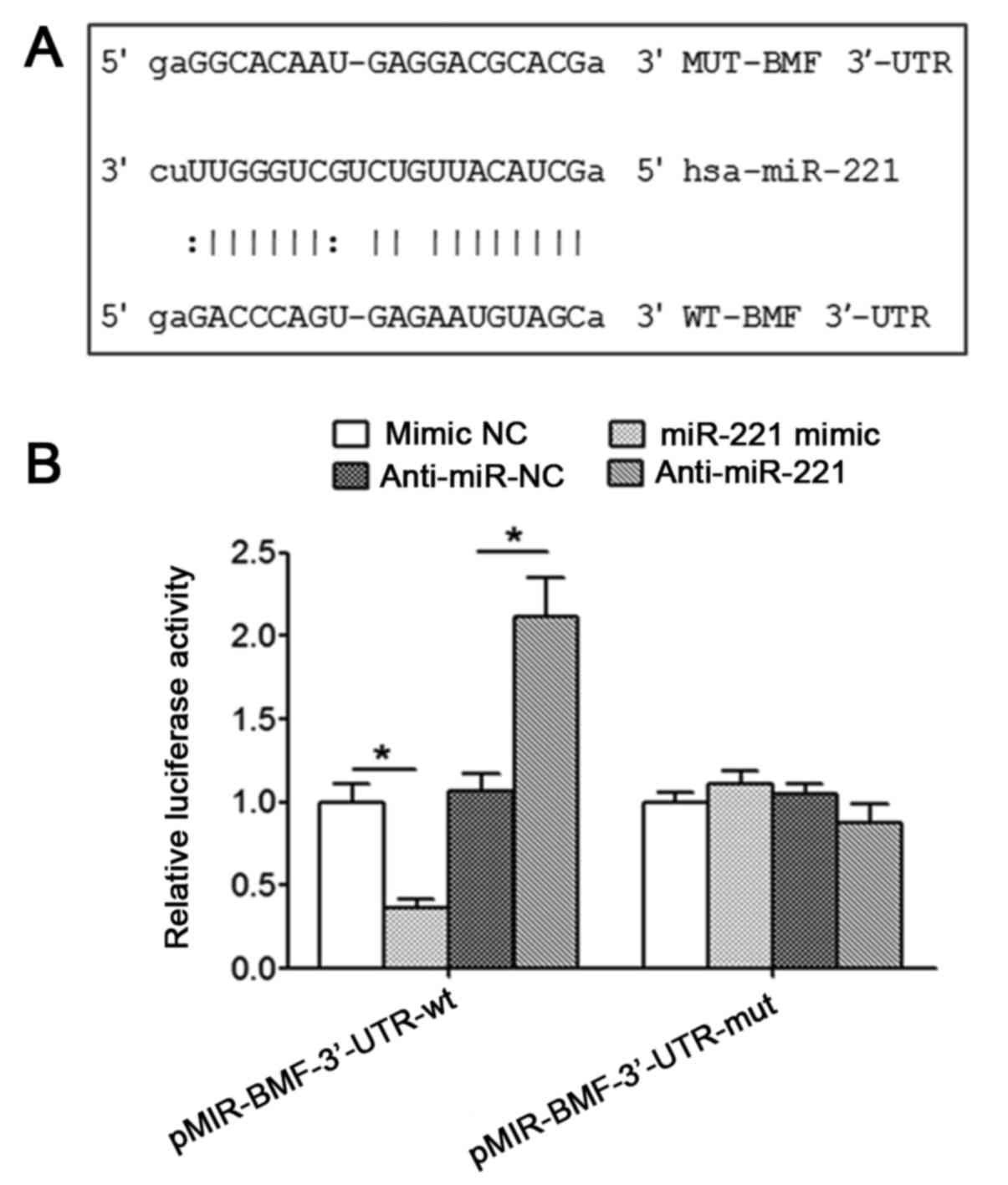
Targeted regulatory effect between
miR-221 and BMF
The microRNA.org
online prediction indicated that BMF may be the target gene for
miR-221. The predicted BMF 3′-UTR target site of miR-221 was
depicted in Fig. 2A. The results of
the luciferase reporter assay demonstrated that the co-transfection
of the wild-type pMIR-BMF-3′-UTR-wt vector and miR-221 mimic
significantly decreased the activity of luciferase (P<0.05), and
the co-transfection of wild-type pMIR-BMF-3′-UTR-wt vector and
anti-miR-221 significantly increased the activity of luciferase
(P<0.05); however, the co-transfection of mutant
pMIR-BMF-3′-UTR-mut vector and miR-221 mimic or anti-miR-221 did
not significantly change the activity of luciferase, indicating a
targeted regulatory association between miR-221 and BMF mRNA
(Fig. 2B).
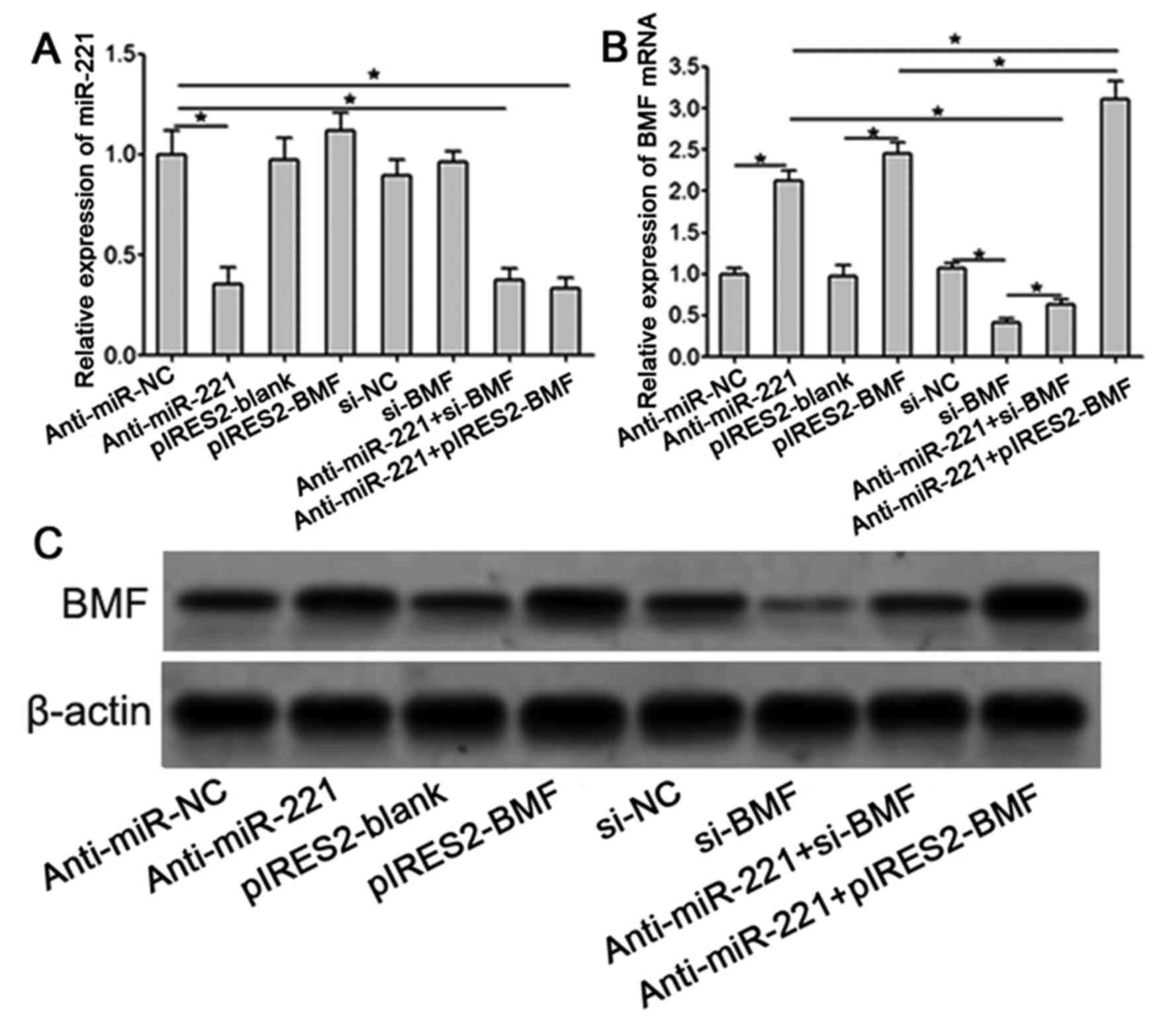
Transfection of anti-miR-221 or
pIRES2-BMF significantly upregulates the expression of BMF in
ovarian cancer SKOV3 cells
The RT-qPCR test results demonstrated that,
following transfection with anti-miR-221, the expression of miR-221
in SKOV3 cells was significantly decreased (P<0.001). The
transfection of si-BMF or pIRES2-BMF had no significant effect on
the expression of miR-221 in SKOV3 cells (Fig. 3A). Compared with anti-miR-NC group,
the expression of BMF mRNA in SKOV3 cells was increased in the
anti-miR-221 transfection group. Compared with the pIRES2-blank
group, the expression of BMF mRNA SKOV3 cells was significantly
increased in the pIRES2-BMF transfection group (P<0.001), and
the transfection of si-BMF downregulated the expression of BMF mRNA
and attenuated the upregulation of anti-miR-221 on the expression
of BMF proteins (Fig. 3B). Western
blot analysis indicated that the transfection of anti-miR-221 or
pIRES2-BMF significantly upregulated the expression of BMF protein
in SKOV3 cells (P<0.001), and the expression level of BMF
protein was highest in the anti-miR-221 and pIRES2-BMF combined
transfection group. The transfection of si-BMF significantly
inhibited the expression of BMF protein and reduced the
upregulation of anti-miR-221 on the expression of BMF protein
(Fig. 3C).
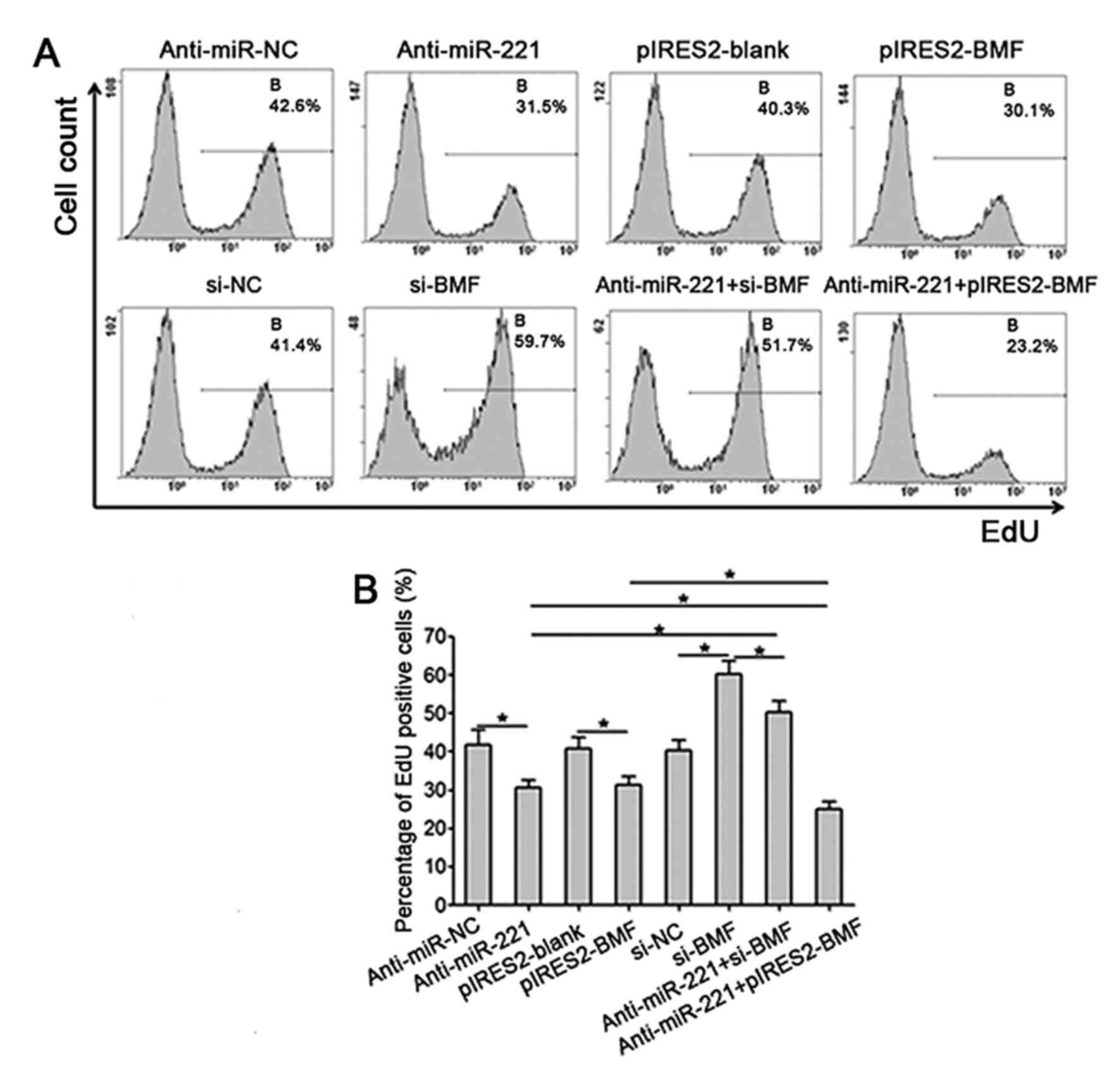
Transfection of anti-miR-221 or
pIRES2-BMF significantly attenuates the proliferation of SKOV3
cells
The EdU staining flow cytometry demonstrated that
the proliferation of SKOV3 cells in the anti-miR-221 transfection
group was significantly lower, compared with the anti-miR-NC group
(P<0.001). Compared with the pIRES2-blank group, the
proliferation of SKOV3 cells in the pIRES2-BMF transfection group
was reduced significantly (P<0.001), and the transfection of
pIRES2-BMF enhanced the inhibitory effect of anti-miR-221 on the
proliferation of SKOV3 cells (Fig. 4A and
B). The transfection of si-BMF significantly promoted cell
proliferation and attenuated the inhibitory effect of anti-miR-221
on the cell proliferation (Fig. 4A and
B).
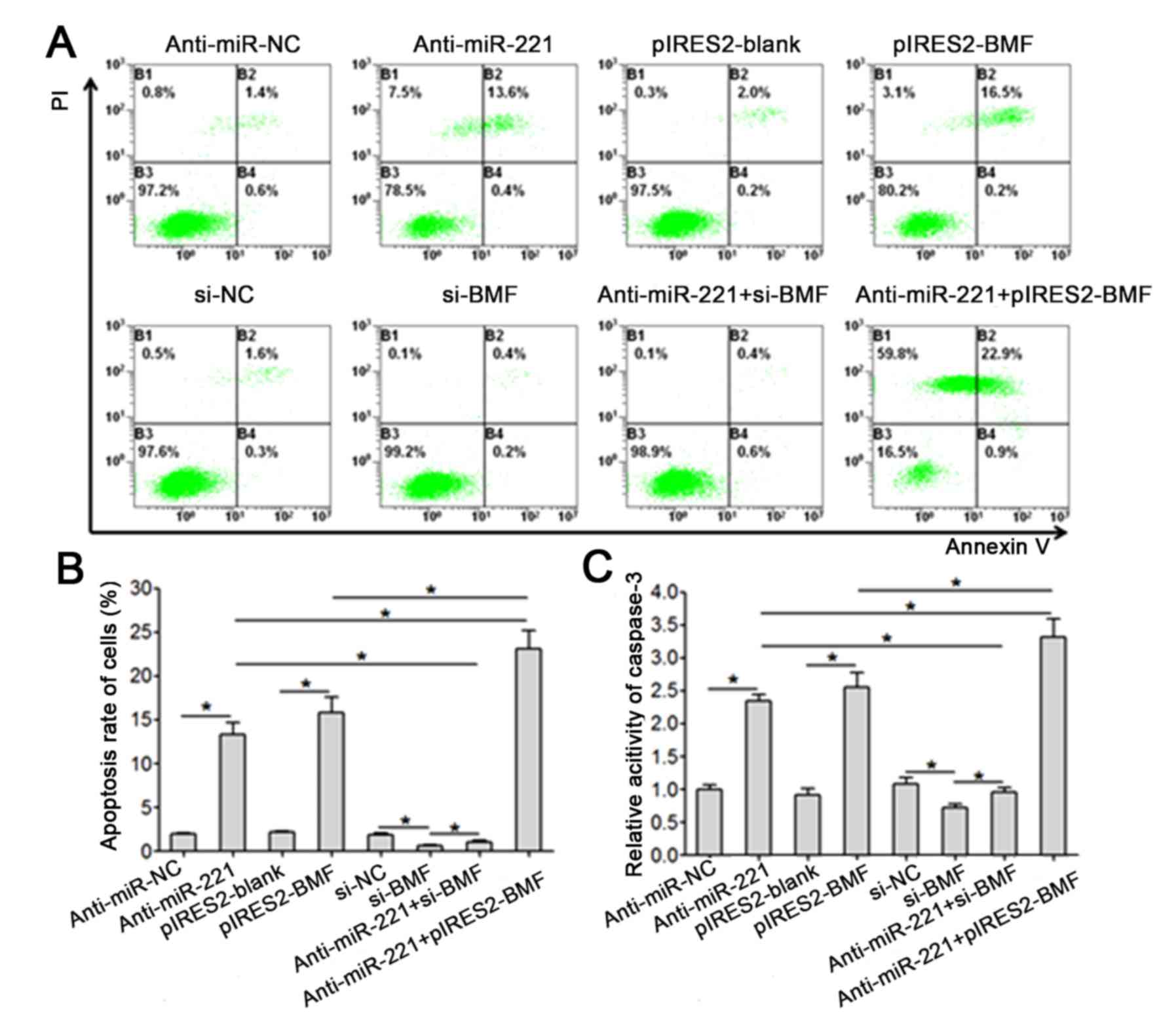
Transfection of anti-miR-221 or
pIRES2-BMF significantly promotes the apoptosis of SKOV3 cells
Flow cytometry indicated that the apoptosis rate of
SKOV3 cells in the anti-miR-221 transfection group was
significantly higher, compared with the anti-miR-NC group
(P<0.001). Similar to the transfection of anti-miR-221, the
transfection of pIRES2-BMF significantly promoted the apoptosis of
SKOV3 cells (P<0.001). The transfection of pIRES2-BMF enhanced
the anti-apoptotic effect of anti-miR-221 on SKOV3 cells (Fig. 5A and B). The transfection of si-BMF
significantly reduced apoptosis and attenuated the proapoptotic
effect of anti-miR-221 on SKOV3 cells (Fig. 5A and B). The results of
spectrophotometry demonstrated that the transfection of
anti-miR-221 or pIRES2-BMF significantly upregulated the activity
of caspase-3 in SKOV3 cells (P<0.001), whilst the transfection
of si-BMF significantly downregulated the activity of caspase-3 in
SKOV3 cells (P<0.001; Fig.
5C).
Discussion
In the present study, the expression levels of
miR-221 and BMF mRNA in ovarian cancer tissues were detected by
RT-qPCR. Results indicated that the expression level of miR-221 in
ovarian cancer tissues was significantly higher, compared with the
para-carcinoma tissues, whilst the expression of BMF mRNA in
ovarian cancer tissues was lower, compared with the para-carcinoma
tissues, indicating that miR-221 serves as a tumor-promoting gene
and BMF as a tumor suppressor gene in the ovarian cancer. In the
previous studies regarding the association between miR-221 and
ovarian cancer, Dahiya et al (24) conducted detection of miR-221 via a
gene microarray and demonstrated that the expression levels of
miR-221 in tumor tissues of patients with ovarian cancer and
ovarian cancer cell lines was significantly increased. The
detection by Gadducci et al (25) indicated that, compared with normal
ovarian tissue, the expression of miR-221 in patients with ovarian
cancer was increased abnormally. Hong et al (22) conducted RT-qPCR detection and
demonstrated that, compared with the healthy control populations,
the expression of miR-221 in the peripheral blood of patients with
ovarian cancer was abnormally increased, and its expression level
was correlated with the disease stage of International Federation
of Gynecology and Obstetrics and histopathological grading, with
the higher the expression of miR-221, the lower the survival rate
and prognosis of patients. Li et al (21) determined that the expression levels of
miR-221 were significantly increased in ovarian cancer cells A2780,
OVCAR3, SKOV3 and 3AO, compared with the immortalized ovarian
epithelial IOSE25 cells; and compared with para-cancer tissues, the
expression of miR-221 in the tumor tissues of patients with ovarian
cancer was also increased abnormally, with the overall survival
rate and progression-free survival rate of patients with higher
expression of miR-221 being significantly lower than those in
patients with low expression of miR-221. In the present study, the
expression of miR-221 in tumor tissue of patients with ovarian
cancer was significantly higher, compared with the para-cancer
tissues, which may be carcinogenic factor in the occurrence of
ovarian cancer, similar to the results obtained by Gadducci et
al (25), Hong et al
(22) and Li et al (21).
miRNAs serve biological roles mainly by regulating
the downstream target genes. Different functions of downstream
target genes can cause different functions of the same miRNAs in
different tumor tissues (24,25). The present study confirmed that
miR-221 is predominantly tumor-promoting in ovarian cancer tissues,
whilst BMF, which has an important function in promoting cell
apoptosis and tumor inhibition according to the microRNA.org online prediction, may be a target gene
of miR-221; therefore, BMF was the target gene of interest for the
present study. The results of the dual luciferase reporter assay
demonstrated that the co-transfection of wild-type pMIR-BMF-3′-UTR
vector and miR-221 mimic significantly decreased the activity of
luciferase, and the co-transfection of wild-type pMIR-BMF-3′-UTR
vector and anti-miR-221 significantly increased the activity of
luciferase; whilst the co-transfection of mutant pMIR-BMF-3′-UTR
vector and miR-221 mimic or anti-miR-221 did not significantly
change the activity of luciferase. This indicated that there was a
direct interaction between miR-221 and BMF mRNA, and BMF was the
downstream target gene of miR-221.
To investigate whether miR-221 regulating BMF would
influence the biological effects of ovarian cancer SKOV3 cells,
including proliferation and apoptosis, the changes in proliferation
and apoptosis of SKOV3 cells following transfected with
anti-miR-221or pIRES2-BMF in the SKOV3 cells were detected. The
detection results indicated that the transfection of anti-miR-221
significantly inhibited the proliferation of SKOV3 cells, and the
transfection of pIRES2-BMF significantly attenuated the
proliferation of SKOV3 cells. The transfection of anti-miR-221
significantly promoted the apoptosis of SKOV3 cells, and the
transfection of pIRES2-BMF also resulted in a significant increase
in the number of apoptotic SKOV3 cells, similar to that of
anti-miR-221. The transfection of pIRES2-BMF or si-BMF enhanced or
attenuated the proapoptotic and proliferation-inhibition effect of
anti-miR-221 on SKOV3 cells, respectively. The results indicated
that miR-221 reduced the apoptosis level and enhanced the
proliferation ability of ovarian cancer cells, which developed the
ovarian cancer by inhibiting the expression of BMF. In the present
study, it was firstly confirmed that the miR-221 expression in
ovarian cancer tissues was significantly higher, compared with the
para-cancer tissues, through the RT-qPCR method, and it was
hypothesized that the increase of miR-221 in ovarian cancer tissues
decreased the expression of pro-apoptotic factor BMF, enhanced the
proliferation and reduced the apoptosis of ovarian cancer cells,
which promoted the occurrence and development of ovarian cancer. Li
et al (21) demonstrated that
the transfection of miR-221 inhibitor significantly upregulated the
expression of target gene apoptotic peptidase activating factor 1
and attenuated the proliferation and migration of ovarian cancer
A2780 and SKOV3, whilst the cell apoptosis was significantly
increased, confirming that miR-221 served a tumor-promoting role in
promoting the proliferation of ovarian cancer cells and reducing
cell apoptosis, which confirmed and supported the results in the
present study. He et al (26)
indicated that miR-221 promoted the proliferation of hepatocellular
carcinoma cells by targeting the inhibition of BMF expression,
whilst downregulating the expression of miR-221 inhibited the
proliferation, migration and invasion of hepatocellular carcinoma
cells and promoted the apoptosis of hepatoma cells. This indicated
that the increased expression of miR-221 in liver cancer serves a
role in promoting cancer. The results of Gramantieri et al
(27) demonstrated that abnormally
elevated miR-221 expression also serves a regulatory role in
targeting the inhibition of BMF expression and promoting liver
cancer. In the present study, the dual-luciferase gene reporter
assay confirmed the existence of targeted regulation between
miR-221 and BMF, which further corroborates the results of He et
al (26) and Gramantieri et
al (27). At present, the
association between miR-221 and ovarian cancer is rarely reported,
and the majority of studies focus on the abnormal changes in the
expression of miR-221, without studying its effect in regulating
the biological process of ovarian cancer cells. In the present
study, the abnormal expression of miR-221 in the ovarian cancer
cells were detected, and its biological process in influencing the
proliferation and apoptosis of ovarian cancer cells by regulating
the expression of target gene BMF was investigated, revealing the
tumor-promoting effect of miR-221; however, the process of miR-221
regulation of BMF in ovarian cancer cells in vivo process is
not clear, and requires further animal studies, which is also a
limitation of the present study.
In summary, the expression of miR-221 was
significantly increased and the expression of BMF was significantly
decreased in ovarian cancer tissues. The miR-221 with abnormally
elevated expression antagonized the apoptosis of ovarian cancer
SKOV3 cell and promoted the cell proliferation by targeted
inhibition of the expression of pro-apoptosis factor BMF, which may
serve a role in the pathogenesis of ovarian cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
Guarantor of integrity of the entire study: LC.
Study concepts: YH. Study design: XX. Definition of intellectual
content: YH. Literature research: XX and JW. Clinical studies: XX
and YH. Experimental studies: XX. Data acquisition: JW. Data
analysis: XX and YH. Statistical analysis: XX and LC. Manuscript
preparation: JW. Manuscript editing: XX. Manuscript review: YH and
JC.
Ethics approval and consent to
participate
All specimens were collected with the patient's
written informed consent and the present study was approved by the
Ethics Committee of the First Affiliated Hospital of Fujian Medical
University.
Patient consent for publication
Witten informed consent was obtained from the
patients for the publication of any associated data.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Fedewa SA, Miller KD,
Goding-Sauer A, Pinheiro PS, Martinez-Tyson D and Jemal A: Cancer
statistics for Hispanics/Latinos, 2015. CA Cancer J Clin.
65:457–480. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Power L, Lefas G, Lambert P, Kim D,
Evaniuk D, Lotocki R, Dean E, Nachtigal MW and Altman AD: Hormone
use after nonserous epithelial ovarian cancer: Overall and
disease-free survival. Obstet Gynecol. 127:837–847. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pradjatmo H and Pradjatmo H: Impact of
preoperative serum levels of CA 125 on epithelial ovarian cancer
survival. Asian Pac J Cancer Prev. 17:1881–1886. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liu CF, Liu SY, Min XY, Ji YY, Wang N, Liu
D, Ma N, Li ZF and Li K: The prognostic value of CXCR4 in ovarian
cancer: A meta-analysis. PLoS One. 9:e926292014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siddiqui WA, Ahad A and Ahsan H: The
mystery of BCL2 family: Bcl-2 proteins and apoptosis: An update.
Arch Toxicol. 89:289–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kontos CK, Christodoulou MI and Scorilas
A: Apoptosis-related BCL2-family members: Key players in
chemotherapy. Anticancer Agents Med Chem. 14:353–374. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shamas-Din A, Brahmbhatt H, Leber B and
Andrews DW: BH3-only proteins: Orchestrators of apoptosis. Biochim
Biophys Acta. 1813:508–520. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moldoveanu T, Follis AV, Kriwacki RW and
Green DR: Many players in BCL-2 family affairs. Trends Biochem Sci.
39:101–111. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Thomadaki H and Scorilas A: BCL2 family of
apoptosis-related genes: Functions and clinical implications in
cancer. Crit Rev Clin Lab Sci. 43:1–67. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Frenzel A, Labi V, Chmelewskij W, Ploner
C, Geley S, Fiegl H, Tzankov A and Villunger A: Suppression of
B-cell lymphomagenesis by the BH3-only proteins Bmf and Bad. Blood.
115:995–1005. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang R, Kang Y, Löhr CV, Fischer KA,
Bradford CS, Johnson G, Dashwood WM, Williams DE, Ho E and Dashwood
RH: Reciprocal regulation of BMF and BIRC5 (Survivin) linked to
Eomes overexpression in colorectal cancer. Cancer Lett.
381:341–348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kutuk O and Letai A: Displacement of Bim
by Bmf and Puma rather than increase in Bim level mediates
paclitaxel-induced apoptosis in breast cancer cells. Cell Death
Differ. 17:1624–1635. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li D, Xu D, Xu Y, Chen L, Li C, Dai X,
Zhang L and Zheng L: MicroRNA-141-3p targets DAPK1 and inhibits
apoptosis in rat ovarian granulosa cells. Cell Biochem Funct.
35:197–201. 2017. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu H, Pan Y, Han X, Liu J and Li R:
MicroRNA-216a promotes the metastasis and epithelial-mesenchymal
transition of ovarian cancer by suppressing the PTEN/AKT pathway.
Onco Targets Ther. 10:2701–2709. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Hu K and Liang M: Upregulated microRNA-224
promotes ovarian cancer cell proliferation by targeting KLLN. In
Vitro Cell Dev Biol Anim. 53:149–156. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
PLOS ONE Editors: Retraction: miR-221
promotes tumorigenesis in human triple negative breast cancer
cells. PLoS One. 12:e01758692017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu Q, Li P, Chen X, Zong L, Jiang Z, Nan
L, Lei J, Duan W, Zhang D, Li X, et al: miR-221/222 induces
pancreatic cancer progression through the regulation of matrix
metalloproteinases. Oncotarget. 6:14153–14164. 2015.PubMed/NCBI
|
|
18
|
Gordanpour A, Stanimirovic A, Nam RK,
Moreno CS, Sherman C, Sugar L and Seth A: miR-221 Is down-regulated
in TMPRSS2: ERG fusion-positive prostate cancer. Anticancer Res.
31:403–410. 2011.PubMed/NCBI
|
|
19
|
Yang X, Yang Y, Gan R, Zhao L, Li W, Zhou
H, Wang X, Lu J and Meng QH: Down-regulation of mir-221 and mir-222
restrain prostate cancer cell proliferation and migration that is
partly mediated by activation of SIRT1. PLoS One. 9:e988332014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yamashita R, Sato M, Kakumu T, Hase T,
Yogo N, Maruyama E, Sekido Y, Kondo M and Hasegawa Y: Growth
inhibitory effects of miR-221 and miR-222 in non-small cell lung
cancer cells. Cancer Med. 4:551–564. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li J, Li Q, Huang H, Li Y, Li L, Hou W and
You Z: Overexpression of miRNA-221 promotes cell proliferation by
targeting the apoptotic protease activating factor-1 and indicates
a poor prognosis in ovarian cancer. Int J Oncol. 2017.
|
|
22
|
Hong F, Li Y, Xu Y and Zhu L: Prognostic
significance of serum microRNA-221 expression in human epithelial
ovarian cancer. J Int Med Res. 41:64–71. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bustin SA, Benes V, Garson JA, Hellemans
J, Huggett J, Kubista M, Mueller R, Nolan T, Pfaffl MW, Shipley GL,
et al: The miqe guidelines: Minimum information for publication of
quantitative real-time pcr experiments. Clin Chem. 55:611–622.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dahiya N, Sherman-Baust CA, Wang TL,
Davidson B, Shih Ie M, Zhang Y, Wood W III, Becker KG and Morin PJ:
MicroRNA expression and identification of putative miRNA targets in
ovarian cancer. PLoS One. 3:e24362008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Gadducci A, Sergiampietri C, Lanfredini N
and Guiggi I: Micro-RNAs and ovarian cancer: The state of art and
perspectives of clinical research. Gynecol Endocrinol. 30:266–271.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He XX, Guo AY, Xu CR, Chang Y, Xiang GY,
Gong J, Dan ZL, Tian DA, Liao JZ and Lin JS: Bioinformatics
analysis identifies miR-221 as a core regulator in hepatocellular
carcinoma and its silencing suppresses tumor properties. Oncology
Rep. 32:1200–1210. 2014. View Article : Google Scholar
|
|
27
|
Gramantieri L, Fornari F, Ferracin M,
Veronese A, Sabbioni S, Calin GA, Grazi GL, Croce CM, Bolondi L and
Negrini M: MicroRNA-221 targets Bmf in hepatocellular carcinoma and
correlates with tumor multifocality. Clin Cancer Res. 15:5073–5081.
2009. View Article : Google Scholar : PubMed/NCBI
|