Introduction
Nasopharyngeal carcinoma (NPC) is an
endothelium-associated malignancy with a number of unique
etiological and biological characteristics (1,2). The
neoplasm in patients with NPC originates from the superior aspect
of the pharynx and extends along the soft palate. Primary
etiological factors include genetics and environment, in addition
to infection with Epstein Barr virus (EBV), which presents in 90%
of cases of NPC (3). There is
evidence to suggest that transfection of the viral latent membrane
protein 1 (LMP1) gene into normal epithelial cells triggers the
immortalization of rodent cells and enhances a number of prominent
characteristics of carcinoma (4,5). Various
signaling cascades, including those of phosphatidylinositol
3-kinase/RAC-α serine/threonine-protein kinase (PI3K/Akt),
extracellular signal-regulated kinase and Wnt/β-catenin are broadly
activated by LMPs, and subsequently impact cell proliferation and
motility (4,6,7). Notably,
the PI3K/Akt signaling pathway may be involved in the inhibition of
apoptosis and the promotion of proliferation (8).
Triptolide, a type of epoxy diterpene monomer, is
the principal bio-active component of Tripterygium
wilfordii, and is widely used in Chinese herbal medicine
(9). Triptolide possesses
anti-inflammatory, immunosuppressive and anti-procreation activity,
and has been widely used in the treatment of autoimmune diseases
and the inhibition of immune rejection in patients with organ
transplants (10,11). In addition, triptolide induces
apoptosis in a number of cancer cell lines and increases the
sensitivity of tumor cells to other anticancer drugs (12–14). Such
broad-spectrum anticancer activity makes triptolide an interesting
research prospect. Therefore, it was hypothesized that triptolide
may represent a promising therapy option for nasopharyngeal cancer
by inducing apoptosis through the PI3K/Akt pathway.
Materials and methods
Reagents
Triptolide (purity, >98%) was purchased from
Guangzhou PI&PI Biotech, Inc. (Guangzhou, China). Rabbit
polyclonal anti-cleaved caspase 3 (cat. no: 9661), anti-apoptosis
regulator BAX (Bax, cat. no: 2774), anti-apoptosis regulator Bcl-2
(Bcl-2, cat. no: 2872) and anti-Akt (cat. no: 9272) and mouse
monoclonal anti-phospho-Akt (cat. no: 12694) antibodies were
obtained from Cell Signaling Technology (Danvers, MA, USA). Rabbit
polyclonal anti-GAPDH (cat. no: A00227-1) antibody was obtained
from Wuhan Boster Biological Technology, Ltd., (Wuhan, China).
Horseradish peroxidase (HRP) -labeled goat anti-mouse (cat.no:
115-035-003) or rabbit (cat. no: 111-035-003) antibodies were
purchased from Jackson ImmunoResearch (Bar Harbor, ME, USA). MTT
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and LY294002
(MedChemExpress, Monmouth Junction, NJ, USA) were dissolved in
dimethyl sulfoxide (DMSO) and stored at −20°C, protected from the
light.
Cell culture
The human EBV-positive NPC cell line C666-1, and the
immortalized normal nasopharyngeal epithelial cell line NP69 were
purchased from The American Type Culture Collection (Manassas, VA,
USA). The C666-1 cells were cultured in RPMI 1640 medium (Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (Gibco) with 1% penicillin and 1%
streptomycin. The NP69 cells were cultured in Keratinocyte-SFM
medium (Gibco; Thermo Fisher Scientific, Inc.) supplemented with
10% fetal bovine serum and 1 ng/ml epidermal growth factor
(PeproTech, Inc., Rocky Hill, NJ, USA). All cells were cultured at
37°C with 5% CO2.
Cell proliferation assays
MTT assays were performed according to the
manufacturer's protocol (Sigma-Aldrich; Merck KGaA). Cells were
plated at a density of 5×103 per well in a 96-well
plate, and incubated in the appropriate culture medium with 12.5,
25, 50 and 100 nM triptolide. Alternatively, the plated cells were
cultured in serum-free medium for 24 h and pretreated with 1, 10 or
50 µM LY294002 for 2 h. Subsequently, the C666-1 cells were
cultured in complete medium with 50 nM triptolide or control
solvent. Tetrazolium salts were added and incubated for the final 4
h of the experiment. A volume of 150 µl DMSO was used to dissolve
the formazan crystals. The absorbances were determined at 570
nm.
Cell cycle determination
Cells with a density of 1×105 cell/well
were plated in 6-well plates. The cells were incubated with 12.5,
25, 50 and 100 nM triptolide. After 24 h of incubation at 37°C,
cell cycle analysis was performed. Cells were fixed with cold
ethanol (70%) at 4°C for 12 h, incubated with 100 µl RNase A
solution (Nanjing Jiancheng Bioengineering Institute, Nanjing,
China) for 30 min at 37°C, and then mixed with 400 µl propidium
iodide (PI) solution (Nanjing Jiancheng Bioengineering Institute)
for 30 min at 4°C, protected from light. The labeled cells were
analyzed with a FACSCalibur flow cytometer (BD Biosciences, San
Jose, CA, USA) and the data were processed with FlowJo software VX
(Tree Star, Inc., Ashland, OR, USA).
Apoptosis assay
Apoptosis assays were performed using an Apoptosis
Detection kit (Vazyme, Piscataway, NJ, USA) according to the
manufacturer's protocol. The cells treated with triptolide were
collected, resuspended and incubated in 500 µl binding buffer with
5 µl Annexin V and 5 µl PI. After 15 min of incubation at room
temperature, the labeled cells were analyzed using a FACSCalibur
flow cytometer and the data were processed using FlowJo
software.
Western blot analysis
Cells were lysed in radioimmunoprecipitation assay
buffer containing phenylmethylsulfonyl fluoride and soybean
protease inhibitor (Beyotime Institute of Biotechnology, Haimen,
China). Following 13,000 × g centrifugation at 4°C for 15 min, the
supernatants were harvested for protein quantification using a
bicinchoninic acid assay kit (Beyotime Institute of Biotechnology).
A total of 40 µg protein/lane was loaded, and separated using a 10
or 12% polyacrylamide gel. The proteins were transferred to a PVDF
membrane (EMD Millipore, Billerica, MA, USA). Following washing
three times in deionized water, the membranes were blocked with 5%
skim milk for 120 min at room temperature, incubated with the
primary rabbit anti-cleaved caspase 3 antibody, anti-Bax antibody,
anti-Bcl-2 antibody, anti-Akt antibody (all Cell Signaling
Technology, Inc., Danvers, MA, USA) anti-GAPDH antibody (Wuhan
Boster Biological Technology, Ltd.) or mouse monoclonal
anti-phospho-Akt antibody (Cell Signaling Technology, Inc.)
overnight at 4°C (dilution, 1:1,000). Following washed three times
with Tris buffered saline/0.5% Tween-20, the members were
subsequently incubated with HRP-conjugated goat anti-rabbit or
anti-mouse antibodies (dilution, 1:1,000 or 1:5,000 respectively)
at room temperature for 60 min. The membranes were visualized with
an enhanced chemiluminescence kit (EMD Millipore), with GAPDH as a
control. All the images were quantified by Image J 1.1 software
(National Institutes of Health, Bethesda, MD, USA).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The RNA was reverse transcribed using Moloney Murine
Leukemia Virus RT (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocol. RT-qPCR was performed
with the SYBR qPCR kit (Vazyme) using a Stratagene Mx3000
Quantitative RT-PCR System. Amplification procedures were as
follows: 95°C for 5 min, 30 cycles of 95°C for 20 sec, 62°C for 30
sec and ultimately 72°C for 10 min. The primer pairs for the human
Akt1 gene were as follows: Akt1 forward 5′-GCAGCACGTGTACGAGAAGA-3′;
Akt1 reverse 5′-GGTGTCAGTCTCCGACGTG-3′; GAPDH forward
5′-ATCAGCAATGCCTCCTGCAC-3′; GAPDH REVERSE
5′-TGGCATGGACTGTGGTCATG-3′. The relative Akt1 mRNA level was
normalized to GAPDH. Relative quantification was performed using
the 2−ΔΔCq method (15).
Statistical analysis
All graphs were processed by GraphPad Prism version
5 (GraphPad Software, Inc., La Jolla, CA, USA). The data are
displayed as the mean ± standard deviation, and were analyzed using
the SPSS 16.0 statistical package (SPSS, Inc., Chicago, IL, USA).
Statistical significance among the indicated groups was determined
by ANOVA following Tukey's test or unpaired Student's t-test.
P<0.05 was considered to indicate a statistically significance
difference.
Results
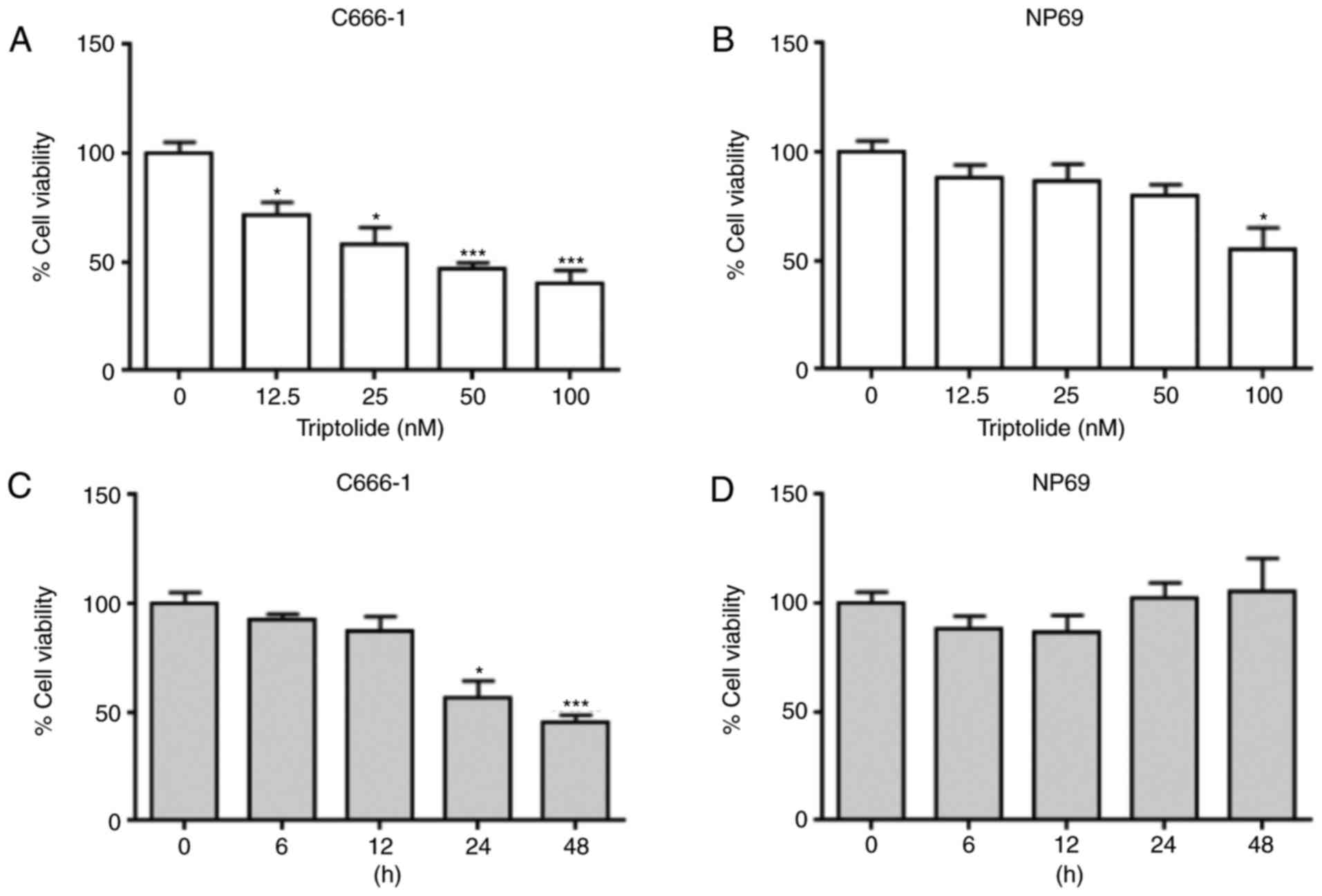
Triptolide reduces the viability of
C666-1 and NP69 cells
To investigate the cytotoxic properties of
triptolide in the C666-1 and NP69 cell lines, cells were incubated
with serial concentrations of triptolide (12.5, 25, 50 and 100 nM;
Fig. 1A and B) for 24 h. Significant
differences were observed at ≥12.5 nM in C666-1 cells, and at 100
nM in NP69 cells. With increasing concentrations of triptolide, the
viability of the C666-1 cells decreased in a dose-dependent manner.
To study the association between cell viability and time, the
C666-1 and NP69 cells were incubated with 50 nM of triptolide and
analyzed at 6, 12, 24 and 48 h (Fig. 1C
and D). The viability of the C666-1 cells was significantly
reduced at >12 h, whilst no significant difference in viability
was observed in NP69 cells at any of the time points tested. These
results indicated that the cytotoxic effects of triptolide were
more potent in C666-1 cells compared with NP69 cells.
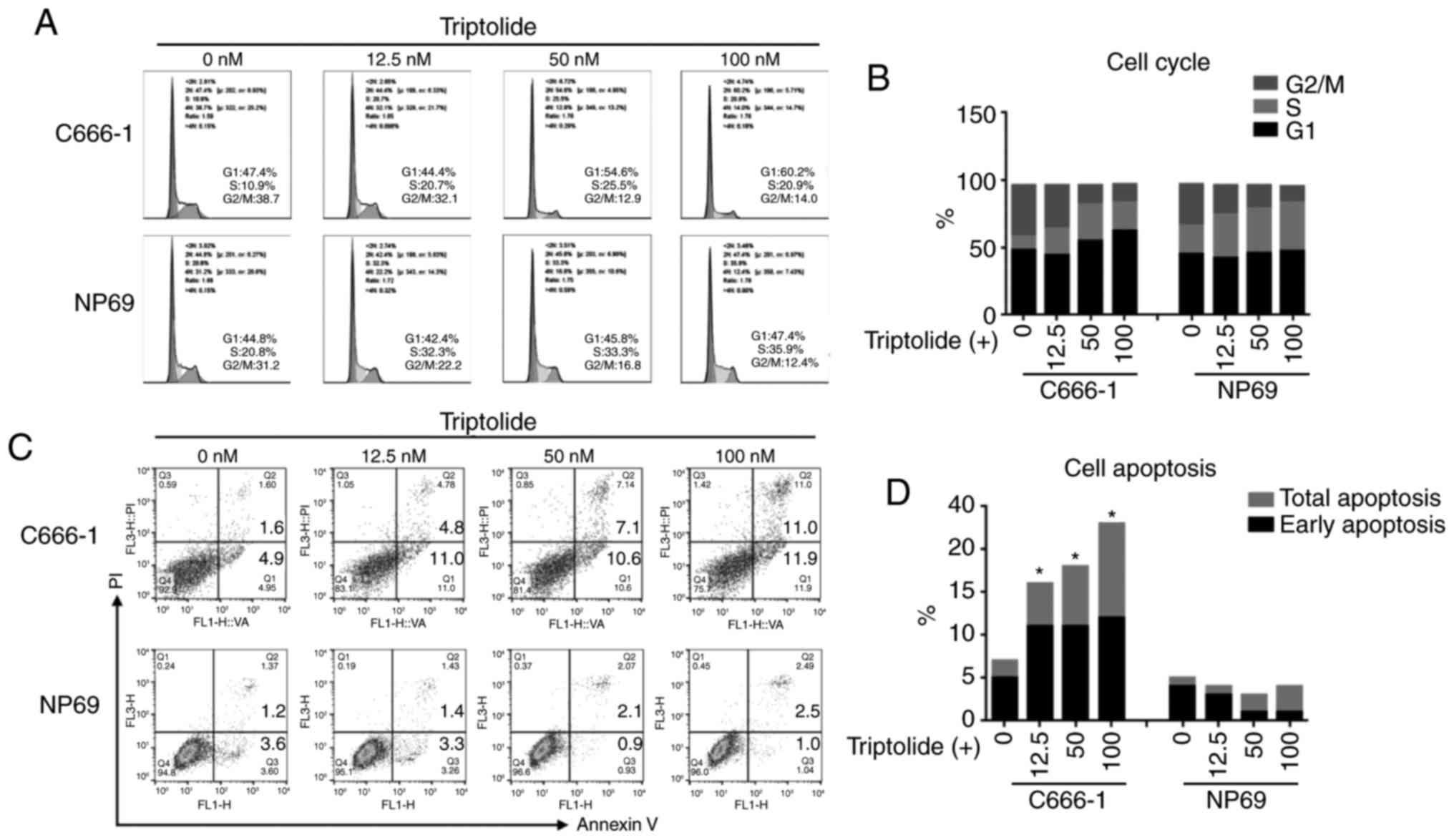
Triptolide induces cell cycle arrest
and promotes apoptosis
To investigate the role of triptolide-induced
cytotoxicity on the cell cycle and apoptosis, cells were incubated
at three concentrations of triptolide for 48 h. The cell cycle
assay suggested that treating C666-1 cells with 50 or 100 nM
triptolide caused an increase in the number of cells in the G0/G1
phase, and a decrease in the number of cells in the G2/M phase,
implying a G0/G1 arrest (Fig. 2A and
B). Notably, although a reduction of the cell population in the
G2/M phase was also observed in NP69 cells, a greater number of
cells were in the S phase rather than the G0/G1 phase. The flow
cytometry assay demonstrated that cell apoptosis rates were
increased in C666-1 cells in a dose-dependent manner, at
concentrations of 12.5, 50 and 100 nM triptolide. The NP69 cells
displayed low levels of apoptosis (Fig.
2C and D).
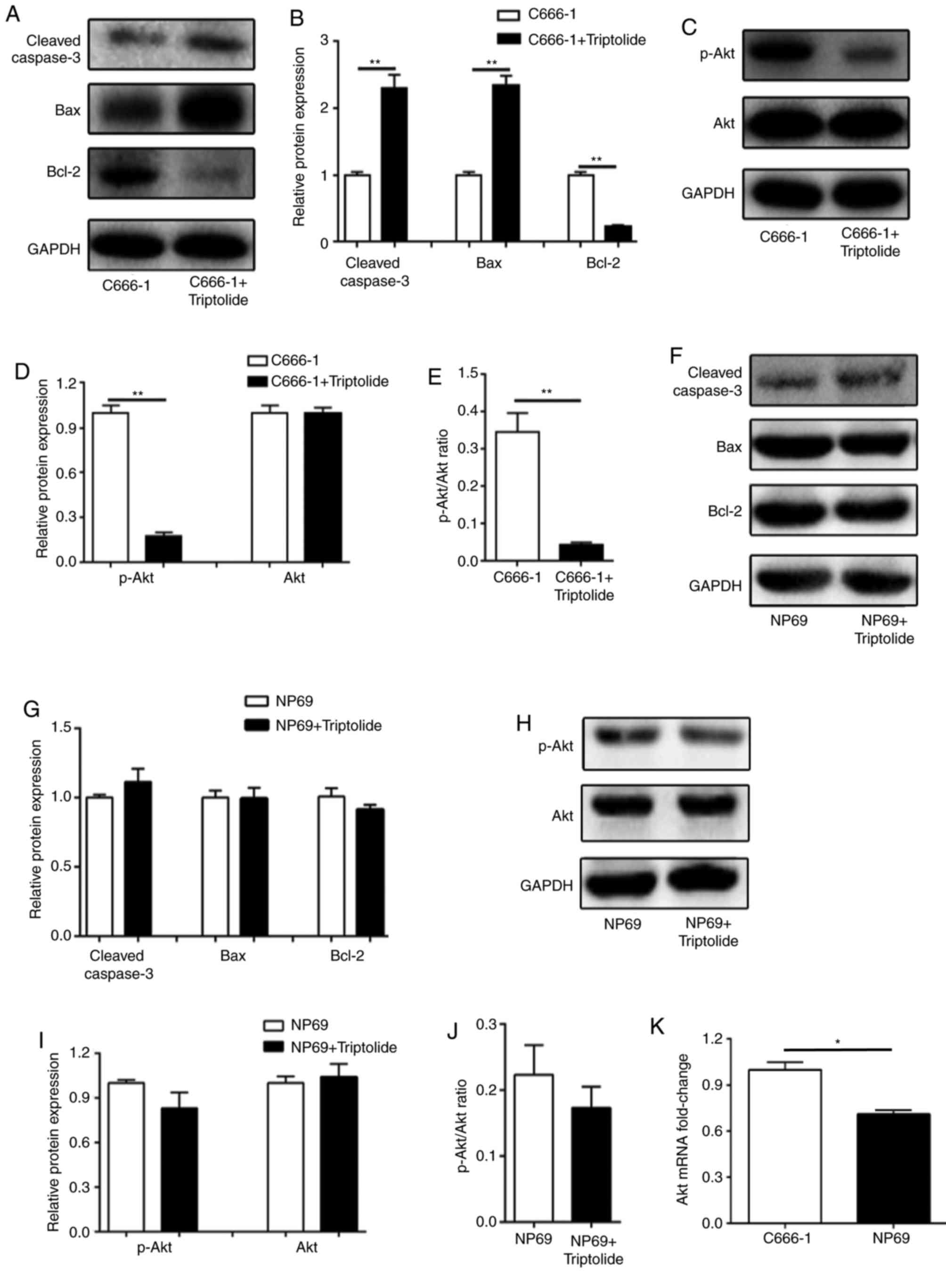
Triptolide modulates pro-apoptotic
protein expression and inhibits the PI3K/Akt signaling pathway
Given that triptolide displayed apoptotic activity
in the C666-1 cells, the expression of cleaved caspase-3, Bax and
Bcl-2 in cells treated with 50 nM triptolide were measured by
western blotting. Upregulated cleaved caspase-3 and Bax expression,
and downregulated Bcl-2 expression, was observed in the C666-1
cells (Fig. 3A and B). Previous
studies have indicated that PI3K/Akt is involved in the
anti-cancerous activity of triptolide, in addition to EBV-mediated
carcinogenesis. The results of the western blotting confirmed that
phosphorylation of Akt was significantly inhibited by triptolide in
the C666-1 cell line (Fig. 3C-E). By
contrast, no significant difference was observed in the expression
levels of cleaved caspase-3, Bax, Bcl-2, p-Akt and Akt in NP69
cells following treatment with 50 nM triptolide (Fig. 3F-J). In addition, the Akt1 mRNA
expression level was significantly higher in C666-1 cells, which
may indicate why C666-1 cells were more sensitive to triptolide
compared with NP69 cells (Fig. 3K).
These data implied that the pro-apoptotic effect and inhibition of
proliferation induced by triptolide may be associated with the
inhibition of the PI3K/Akt signaling pathway.
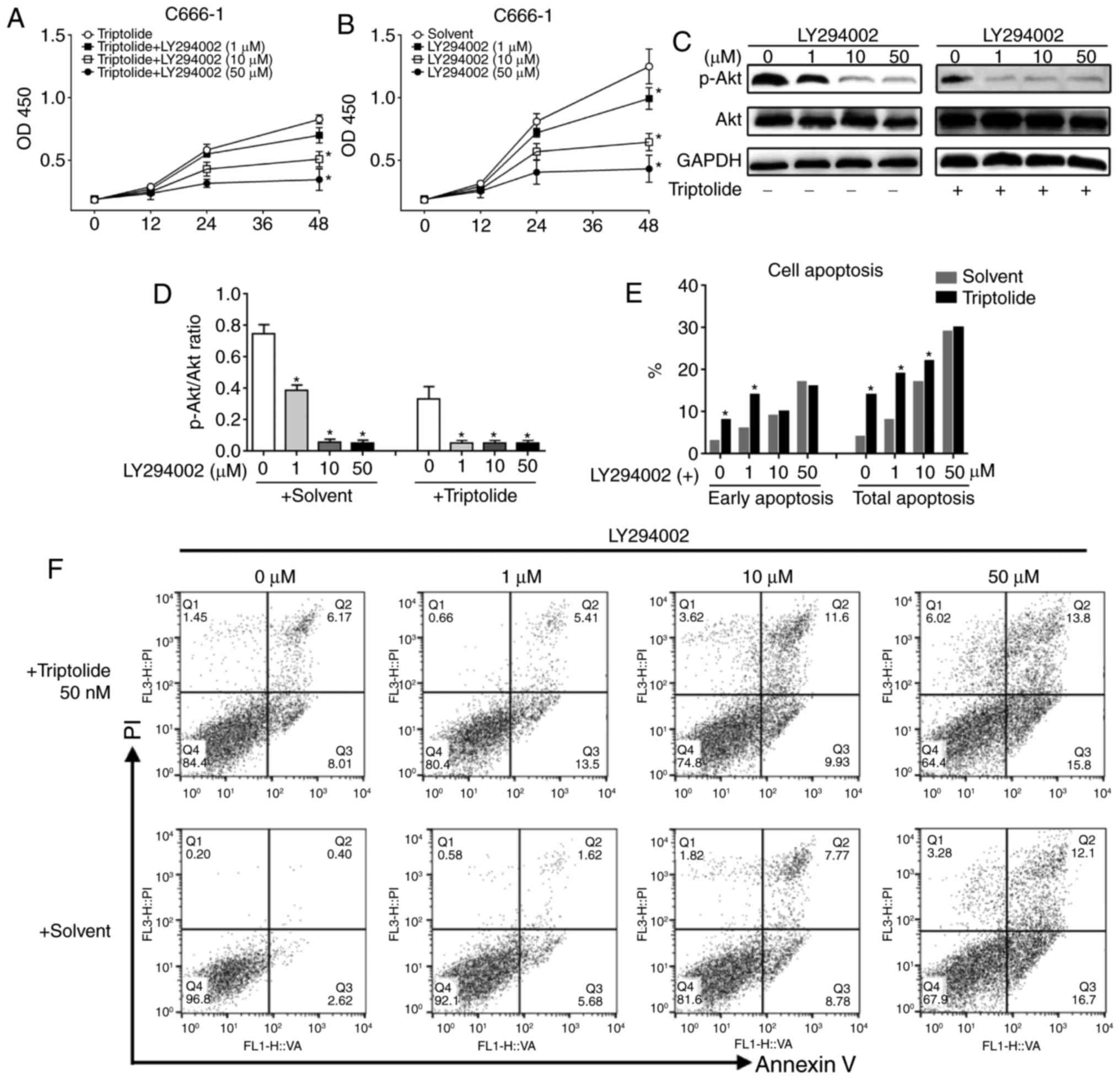
C666-1 cells pretreated with a
PI3K/Akt inhibitor display impaired triptolide-induced apoptosis
and inhibition of proliferation
To confirm the role of PI3K/Akt signaling in
triptolide-induced cytotoxicity, C666-1 cells were pre-incubated
with 1, 10 and 50 µM of the PI3K/Akt inhibitor LY294002 to block
PI3K/Akt signaling. The C666-1 cells were subsequently incubated
with 50 nM triptolide; samples treated with LY294002 only served as
a control. The results revealed that LY294002 inhibited the
proliferation of the C666-1 cells in a dose-dependent manner, and
with increasing concentrations of LY294002, the triptolide-induced
inhibition of cell proliferation was further decreased (Fig. 4A and B). The expression levels of
p-Akt and the ratios of p-Akt/Akt were significantly reduced when
treated with either LY294002 or triptolide (Fig. 4C and D). At a low concentration of
LY294002, the addition of triptolide had a further inhibitory
effect on the phosphorylation of Akt. FACS apoptosis assays were
performed at 48 h post-addition of triptolide. The results
indicated that the pro-apoptotic effect of LY294002 was
dose-dependent, and a further increase in the number of apoptotic
cells was observed with the addition of triptolide (Fig. 4E). The ratios of apoptotic cells are
displayed in Fig. 4F. In summary,
these results suggested that triptolide induces apoptosis and
inhibits cell proliferation in a PI3K/Akt-dependent manner.
Discussion
The anti-cancer effects of triptolide have been
discussed with respect to multiple types of cancer. However, the
underlying mechanism and its application in NPC was yet to be
explored. The results of the present study indicated that
triptolide exerted greater toxicity in NPC C666-1 cells when
compared with normal nasal epithelial NP69 cells. Triptolide
inhibited proliferation and cell cycle progression, and induced
apoptosis in C666-1 cells. The mechanism was associated with the
activation of caspase-3, Bax, and the inhibition of Bcl2, in
addition to Akt phosphorylation. The PI3K/Akt signaling pathway
served an important role in the apoptosis and the inhibition of
proliferation induced by triptolide.
Cell cycle arrest contributes to the inhibition of
cancer cell proliferation induced by various cytotoxic anticancer
agents. Triptolide may induce different types of cell cycle arrest,
depending on the characteristics of the tumor cells concerned. A
previous study indicated that triptolide results in the
accumulation of certain cells in the S phase, including MDA-MB-231,
HT-1080 and OS-RC-2 cells (15). In
the present study, analysis of the cell cycle suggested that
triptolide induced a potent G0/G1 phase arrest in NPC C666-1 cells
and to some extent, influenced the accumulation of NP69 cells in
the S phase.
Triptolide possesses potent anticancer activity with
a half-maximal inhibitory concentration (IC50) ranging
between 4 and 50 nM (16). Previous
studies have identified that triptolide-associated apoptosis is the
central cause of its cytotoxicity (17,18). The
underlying mechanism of apoptosis induced by triptolide is
associated with the activation of caspase-3, 8 and 9, the cleavage
of poly ADP ribose polymerase to release cytochrome C from
mitochondria, and the initiation of cellular tumor antigen
p53-dependent pathways (16). Of
note, the Bcl-2 family dominates the intrinsic apoptosis pathways.
The results of the present study suggested that triptolide promotes
the activation of caspase-3, upregulates the pro-apoptotic protein
Bax, and downregulates the anti-apoptotic protein Bcl-2 to activate
the mitochondrial apoptosis pathway.
Given that the PI3K/Akt signaling pathway is
highlighted in EBV-positive NPC, the present study investigated any
alterations in Akt signaling in C666-1 cells. The results indicated
that Akt phosphorylation was significantly inhibited by triptolide.
As Akt is identified as a key modulator of apoptosis and cell
proliferation, C666-1 cells were pretreated with the PI3K/Akt
inhibitor LY294002 prior to the assessment of proliferation and
apoptosis. Increasing concentrations of LY294002 further induced
triptolide-associated apoptosis and inhibition of cell
proliferation in the C666-1 cells. In addition, studies have also
demonstrated that the inactivation of Akt is closely associated
with the triptolide-induced inhibition of N-terminally-truncated
retinoid X receptor-α in various cancer cells, and E3
ubiquitin-protein ligase Mdm2 in human breast cancer (15,19,20).
In conclusion, the present study demonstrated that
triptolide reduced the viability of C666-1 cells. The NPC C666-1
cells were highly susceptible to triptolide and exhibited potent
apoptosis following incubation in a PI3K/Akt-dependent manner.
Acknowledgements
The authors would like to acknowledge Mr. Shuihong
Zhou, Dr Zhili Zhang and Mr. Libo Dai for their help with the
design of the experiment and data analysis.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
MW and BC performed the experiments, analyzed the
data and wrote the manuscript. LC, designed the experiment, drafted
the manuscript, provided materials and supervised the
experiments.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lo KW, Chung GT and To KF: Deciphering the
molecular genetic basis of NPC through molecular, cytogenetic, and
epigenetic approaches. Semin Cancer Biol. 22:79–86. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Maxwell JH, Kumar B, Feng FY, McHugh JB,
Cordell KG, Eisbruch A, Worden FP, Wolf GT, Prince ME, Moyer JS, et
al: HPV-positive/p16-positive/EBV-negative nasopharyngeal carcinoma
in white North Americans. Head Neck. 32:562–567. 2010.PubMed/NCBI
|
|
4
|
Dawson CW, Port RJ and Young LS: The role
of the EBV-encoded latent membrane proteins LMP1 and LMP2 in the
pathogenesis of nasopharyngeal carcinoma (NPC). Semin Cancer Biol.
22:144–153. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Shah KM, Stewart SE, Wei W, Woodman CB,
O'Neil JD, Dawson CW and Young LS: The EBV-encoded latent membrane
proteins, LMP2A and LMP2B, limit the actions of interferon by
targeting interferon receptors for degradation. Oncogene.
28:3903–3914. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dawson CW, Rickinson AB and Young LS:
Epstein-barr virus latent membrane protein inhibits human
epithelial cell differentiation. Nature. 344:777–780. 1990.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Allen MD, Young LS and Dawson CW: The
Epstein-barr virus-encoded LMP2A and LMP2B proteins promote
epithelial cell spreading and motility. J Virol. 79:1789–1802.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oda K, Okada J, Timmerman L,
Rodriguez-Viciana P, Stokoe D, Shoji K, Taketani Y, Kuramoto H,
Knight ZA, Shokat KM and McCormick F: PIK3CA cooperates with other
phosphatidylinositol 3′-kinase pathway mutations to effect
oncogenic transformation. Cancer Res. 68:8127–8136. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Corson TW and Crews CM: Molecular
understanding and modern application of traditional medicines:
Triumphs and trials. Cell. 130:769–774. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ma J, Dey M, Yang H, Poulev A, Pouleva R,
Dorn R, Lipsky PE, Kennelly EJ and Raskin I: Anti-inflammatory and
immunosuppressive compounds from Tripterygium wilfordii.
Phytochemistry. 68:1172–1178. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yang SX, Gao HL, Xie SS, Zhang WR and Long
ZZ: Immunosuppression of triptolide and its effect on skin
allograft survival. Int J Immunopharmacol. 14:963–969. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Banerjee S, Sangwan V, McGinn O, Chugh R,
Dudeja V, Vickers SM and Saluja AK: Triptolide-induced cell death
in pancreatic cancer is mediated by O-GlcNAc modification of
transcription factor Sp1. J Biol Chem. 288:33927–33938. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu PP, Liu KC, Huang WW, Ma CY, Lin H,
Yang JS and Chung JG: Triptolide induces apoptosis in human adrenal
cancer NCI-H295 cells through a mitochondrial-dependent pathway.
Oncol Rep. 25:551–557. 2011.PubMed/NCBI
|
|
14
|
Yang S, Chen J, Guo Z, Xu XM, Wang L, Pei
XF, Yang J, Underhill CB and Zhang L: Triptolide inhibits the
growth and metastasis of solid tumors. Mol Cancer Ther. 2:65–72.
2003.PubMed/NCBI
|
|
15
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xiong J, Su T, Qu Z, Yang Q, Wang Y, Li J
and Zhou S: Triptolide has anticancer and chemosensitization
effects by down-regulating Akt activation through the MDM2/REST
pathway in human breast cancer. Oncotarget. 7:23933–23946. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Q, Lu Z, Jin Y, Wu Y and Pan J:
Triptolide inhibits Jak2 transcription and induces apoptosis in
human myeloproliferative disorder cells bearing Jak2V617F through
caspase-3-mediated cleavage of Mcl-1. Cancer Lett. 291:246–255.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Carter BZ, Mak DH, Schober WD, Dietrich
MF, Pinilla C, Vassilev LT, Reed JC and Andreeff M: Triptolide
sensitizes AML cells to TRAIL-induced apoptosis via decrease of
XIAP and p53-mediated increase of DR5. Blood. 111:3742–3750. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Liu L, Li G, Li Q, Jin Z, Zhang L, Zhou J,
Hu X, Zhou T, Chen J and Gao N: Triptolide induces apoptosis in
human leukemia cells through caspase-3-mediated ROCK1 activation
and MLC phosphorylation. Cell Death Dis. 4:e9412013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang PY, Zeng WJ, Liu J, Wu YL, Ma Y, Zeng
Z, Pang JY, Zhang XK, Yan X, Wong AST and Zeng JZ: TRC4, an
improved triptolide derivative, specifically targets to truncated
form of retinoid X receptor-alpha in cancer cells. Biochemical
pharmacology. 124:19–28. 2017. View Article : Google Scholar : PubMed/NCBI
|