Introduction
Breast cancer is a malignant cancer that has the
highest cancer-associated morbidity rate worldwide, with an
increasing age of onset (1). Each
year, ~1.2 million women are diagnosed with breast cancer globally,
among which the majority succumb at 40–45 years of age (2). Over 33% of patients with breast cancer
eventually succumb to the disease according to epidemiological
investigation at 2013 (3). The
association between the abnormity of cancer suppressor genes,
particularly tumor protein p53, which acts independently, and
breast cancer occurrence and progression, has become the focus of
an increasing number of studies (4–6).
Therefore, by investigating the p53 pathway, novel therapeutic
methods, or antitumor discoveries for breast cancer, may be
identified.
As chemotherapy and/or radiotherapy are associated
with side effects, and because of high mortality rates, patients
with cancer often use alternative therapies, including natural or
herbal medicines (7). This has
resulted in increased interest in and an active search for novel
anticancer agents from natural products (8). In traditional Chinese medicine (TCM),
medicinal plants and their extracts are used to treat various
diseases. Accumulative data concerning TCM have shown marked
activity with regard to the effects on the tumor cell death
pathway, which can guide tumor treatment decisions and clinical
management (9). Currently, compounds
with unique chemical entities that are derivatives of natural
medicines are a considerable resource for the development of novel
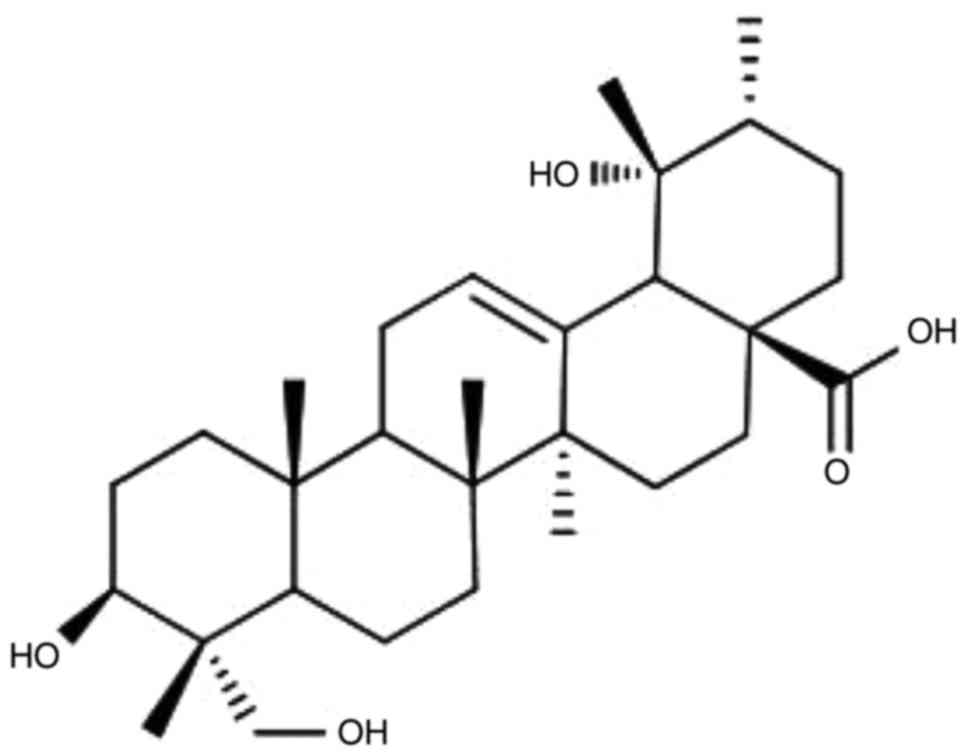
medicines (10). Rotundic acid (RA;
Fig. 1) is a member of the
pentacyclic triterpenoid family and is principally located in
Ilex rotunda, Ilex purpurea, Ilex integra and other
Aquifoliaceae plants, which can be located throughout China
(11). RA can also be isolated from
Mussaenda pubescens and Guettarda platypoda of the
Rubiaceae family (12,13). Olea europaea and
Planchonella duclitan, which are members of the Oleaceae and
Sapotaceae families, respectively, also contain RA (14,15).
The anticancer activity of the majority of currently
used clinical cytotoxic therapies, including radiotherapy, for
patients with cancer, is based on the ability of these therapies to
induce cell death programs, such as apoptosis (16–18). As an
evolutionarily conserved process, apoptosis is characterized by a
number of morphological and biochemical changes, including cell
shrinkage, nuclear DNA fragmentation and membrane blebbing
(19). p53 is an important cancer
suppressor gene in tumors. Upon DNA damage or activation of other
stress signals, p53 is activated and a range of biological
responses occurs, leading to cell cycle arrest, DNA repair and
apoptosis in order to prevent the occurrence and progression of
tumors (20). Furthermore, Xu et
al (21) demonstrated that RA, as
one of a number of isolated compounds, exhibited anticancer
activity. Lee et al (15) also
reported that RA exhibited cytotoxicity, with a half-maximal
inhibitory concentration (IC50) value of 9.5 µM when
applied to the MCF-7 cell line; however, its exact mechanism of
action has not been completely elucidated. The aim of the present
study was to investigate the anticancer effect of RA on the cell
growth and apoptosis of human breast cancer cells, and to evaluate
whether RA may be a potential antitumor drug for the treatment of
the disease.
Materials and methods
Reagents
RA was isolated and purified from I. rotunda,
as reported in our previous study (22). The purity was ≥98% [(as determined by
a high-performance liquid chromatography assay in our previous
study (22)] and the extraction
productivity of RA was much higher than previous methods, up to 100
mg/g. MTT and RNase were purchased from Sigma-Aldrich; Merck KGaA
(Darmstadt, Germany). Propidium iodide and the Annexin
V-Fluorescein Isothiocyanate (FITC) Apoptosis Detection kit were
purchased from BD Biosciences (Franklin Lakes, NJ, USA). Caspase-3
plasmid and the p53 small interfering RNA (siRNA) reagent kit were
purchased from Cell Signaling Technology, Inc (catalog no. 6231,
Danvers, MA, USA). Dulbecco's modified Eagle's medium (DMEM),
trypsin, fetal bovine serum (FBS), PBS, penicillin and streptomycin
were obtained from Gibco; Thermo Fisher Scientific, Inc. (Waltham,
MA, USA).
Preparation of RA
RA was dissolved in PBS (pH, 7.2) to prepare a stock
solution at a concentration of 1.0 mM, which was stored at −20°C.
Complete DMEM [DMEM supplemented with 10% fetal bovine serum (FBS)
and 1% aforementioned antibiotics] was added to dilute the RA to
the appropriate concentrations prior to use.
MCF-7 cell culture and treatment
For transient transfection assays, the MCF-7 cells
(American Type Culture Collection, Manassas, VA, USA) at 80%
confluence were transfected with the caspase-3 expression plasmid
(with pcDNA as control plasmid, 2.0 µg/well, DsRed2,BioVector NTCC
Inc. Beijing China) using Lipofectamine® PLUS reagent
(Thermo Fisher Scientific, Inc.) according to the manufacturer's
protocol. Cells were harvested for RNA extraction at 48 h and
protein extraction at 72 h following transfection. The cells with
optimal overexpression of caspase-3 (Cas3-MCF-7 cells) were
confirmed by western blotting and subsequently used for further
experiments. MCF-7 cells or Cas3-MCF-7 cells were routinely
cultured in complete DMEM, which contained 50 U/ml antibiotics
(penicillin-streptomycin) and 10% FBS, under the conditions of 5%
CO2 at 37°C in a cell incubator (HERAcell 150i; Thermo
Fisher Scientific, Inc.). Following trypsinization by 3 ml trypsin
for 3 min and terminate digestion with 3 ml complete DMEM medium,
to passage the cells in a 75T flask for 3–5 days, the cells were
counted and planted in a 96-well plate in complete DMEM with or
without RA for the MTT array or apoptosis detection. All
experiments were performed three times and examined at 12, 24 and
48 h.
Effect of RA on Cas3-MCF-7 and control
MCF-7 cell viability
The effects of RA on the viability of MCF-7 cells
treated with control plasmid and Cas3-MCF-7 cells were detected
using MTT assays. Cells were seeded in 96-well plates at a density
of 1.0×104 cells/ml at a volume of 200 µl/well. All
groups without or with RA (0, 2.0, 5.0 and 12.5 µM) were incubated
at 37°C for 12, 24 and 48 h. MTT (1.0 mg/ml) was added to each
well, and the cells were incubated at 37°C for 4 h. The solution
was then aspirated, and 100 µl dimethylsulfoxide was added. The
absorbance at 540 nm of the 96-well plates was determined using a
microplate spectrophotometer (Synergy H1, BioTek Instruments, Inc.,
Winooski, VT, USA). The inhibition percentage was calculated as
follows: Inhibition percentage=(1-the value in experimental
group/the value in the control group) ×100.
Flow cytometry analysis of Cas3-MCF-7
cell apoptosis
Annexin V-FITC and PI double-staining flow cytometry
analyses were performed. The Cas3-MCF-7 cells were plated in
96-well plates containing 200 µl complete DMEM at a density of
5×104 cells/well. The induction of apoptosis in the
Cas3-MCF-7 cells was examined with or without RA (2.0, 5.0 and 12.5
µM). Cas3-MCF-7 cells in centrifuge tubes were then resuspended in
a binding buffer. Next, 5 µl annexin V-FITC was added to the tubes,
which were incubated at 4°C for 10 min, followed by the addition of
5 µl PI. The samples were then incubated for another 15 min and
immediately analyzed using a flow cytometer (FACScan; BD
Biosciences) with FlowJo 7.6 FACS analysis software (FlowJo LLC,
Ashland, OR, USA). The cells in the different portions represented
the different cell states as follows: The late-apoptotic cells were
present in the upper right portion, the viable cells were present
in the lower left portion and the early-apoptotic cells were
present in the lower right portion.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was isolated from the Cas3-MCF-7 cells
using a RNeasy kit (Qiagen, Inc., Valencia, CA, USA), 5 µg of each
sample was reverse-transcribed using the M-MLV first-stand
synthesis system (Invitrogen; Thermo Fisher Scientific, Inc., cDNAs
were analyzed in triplicate with the MJ Real-Time PCR System
(Bio-Rad Laboratories, Inc., Hercules, CA, USA) according to the
manufacturer's protocol. The primers for p53 and GAPDH were
designed and synthesized by Shanghai Sangon Biological Engineering
Co. Ltd. (Shanghai, China). The primer sequences for p53 were as
follows: Sense, 5′-TTCCCACTGAGGAGTCCAAC-3′; and antisense,
5′-TTGTTCCCGAAACGCTGAG-3′. The GAPDH primers were as follows:
Sense, 5′-CCAGGTGGTCTCCTCTGACTT-3′ and antisense,
5′-GTTGCTGTAGCCAAATTCGTTGT-3′. Amplification was performed for 40
cycles with a denaturation temperature of 94°C, annealing
temperature of 58°C and extension temperature of 74°C in a thermal
cycler (Veriti; Thermo Fisher Scientific, Inc.). The PCR products
were 200 bp in length. RT-qPCR was performed using the Power SYBR
Green Master Mix (Takara Bio, Inc., Otsu, Japan) and an ABI 7300
real-time PCR detection system (Applied Biosystems, Thermo Fisher
Scientific, Inc.). All primers were synthesized by Invitrogen
(Thermo Fisher Scientific, Inc.). Fold changes in expression of
each gene were calculated by a comparative threshold cycle (Ct)
method using the formula 2−ΔΔCq (23).
Western blotting
Cas3-MCF-7 cell lysates were extracted using
radioimmunoprecipitation buffer (Beyotime Institute of
Biotechnology, Shanghai, China) supplemented with a cocktail
protease inhibitor (Roche Molecular Diagnostics, Pleasanton, CA,
USA), and the protein concentration was determined using a BCA
protein assay kit (Beyotime Institute of Biotechnology) according
to the manufacturer's protocol. A total of 5–40 µg cell total
protein was separated by 10% SDS-PAGE. The proteins were then
transferred onto polyvinylidene difluoride membranes (GE
Healthcare, Chicago, IL, USA) by electroblotting. The membranes
were blocked at 37°C for 1 h with 5% skim milk in Tris-buffered
saline (TBS) with Tween-20 (0.1%) and then incubated with the
anti-caspase-3 (cat no. 9662; 1:1,000), anti-cleaved caspase-3 (cat
no. 9661; 1:1,000), anti-p53 (cat no. 9282; 1:1,000) and
anti-β-actin (cat no. 4967; 1:1,000) antibodies for 1.5 h at room
temperature. The membranes were then washed with Tris-buffered
saline washing buffer six times for 5 min each at room temperature
and incubated with horseradish peroxidase-conjugated goat
anti-mouse (cat no. TA130001) or goat anti-rabbit (cat no.
TA130015) second antibodies (1:2,000; OriGene Technologies, Inc.,
Beijing, China) at 37°C for 1 h. Following washing by TBS buffer,
protein bands were visualized using an enhanced chemiluminescence
system (Thermo Fisher Scientific, Inc.). The primary antibodies
used were all obtained from Cell Signaling Technology Inc. Protein
expression levels were determined semi-quantitatively by
densitometric analysis with the Quantity One software (V4.62,
Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Statistical analysis
All data and results were calculated from at least
three replicate measurements and are presented as the mean ±
standard deviation. The significance between the experimental
groups and the control group were determined using a two-way
analysis of variance (ANOVA) followed by a Dunnett's t-test using
SPSS version 20.0 (IBM Corp., Armonk, NY, USA). P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of RA on MCF-7 and Cas3-MCF-7
cell viability
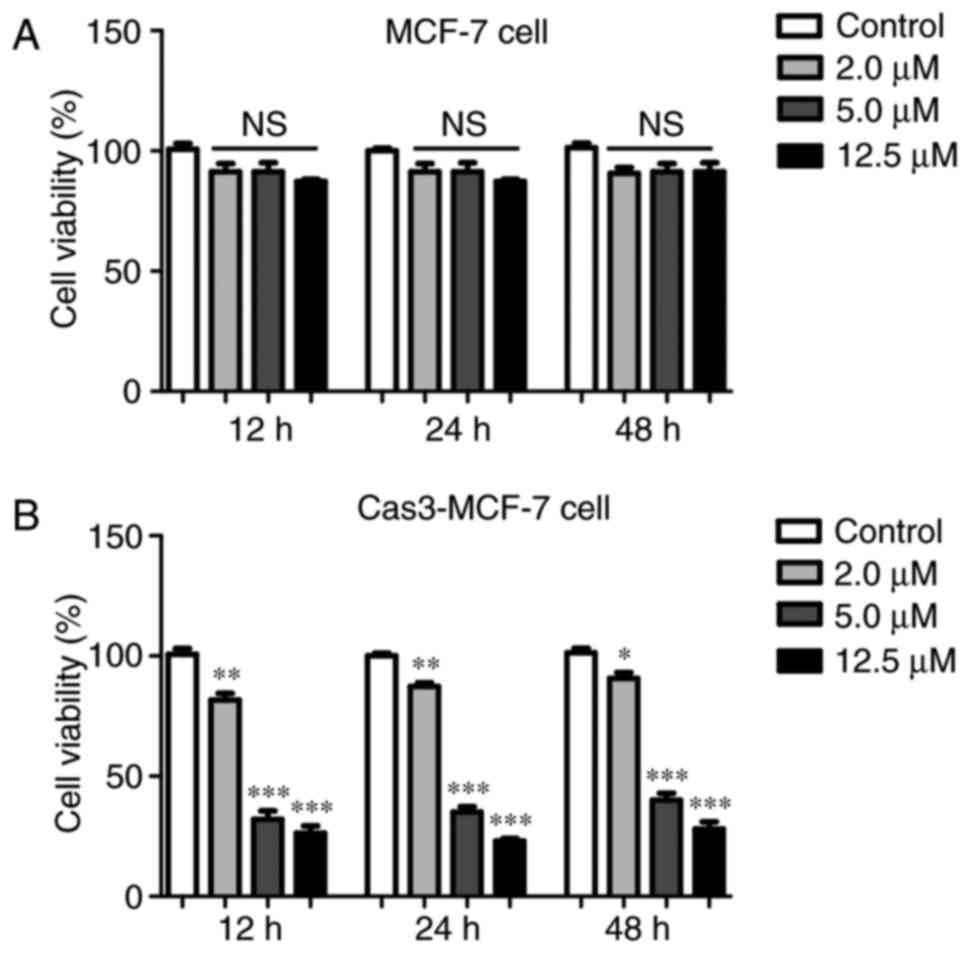
The effect of RA on the viability of MCF-7 cells was
detected using an MTT assay. The structure of RA is illustrated in
Fig. 1. The MTT assay revealed that
RA had limited function with regard to the viability of the MCF-7
cells treated with control plasmid, as illustrated in Fig. 2A. This phenomenon may be due to the
lack of caspase-3 in the MCF-7 cells. Therefore, caspase-3 was
transfected into MCF-7 cells (Cas3-MCF-7 cells) for subsequent
experiments. The inhibitory effects of RA on the Cas3-MCF-7 cells
were dose-dependent within the range of 2–12.5 µmol/l (Fig. 2B). Treatment with RA at 12.5 µmol/l
elicited the greatest inhibitory effect, with a cell viability at
32.3%, compared with that of the blank control group (P<0.001).
There were no differences in inhibition between the different time
points of 12, 24 and 48 h, which revealed similar inhibitory
effects.
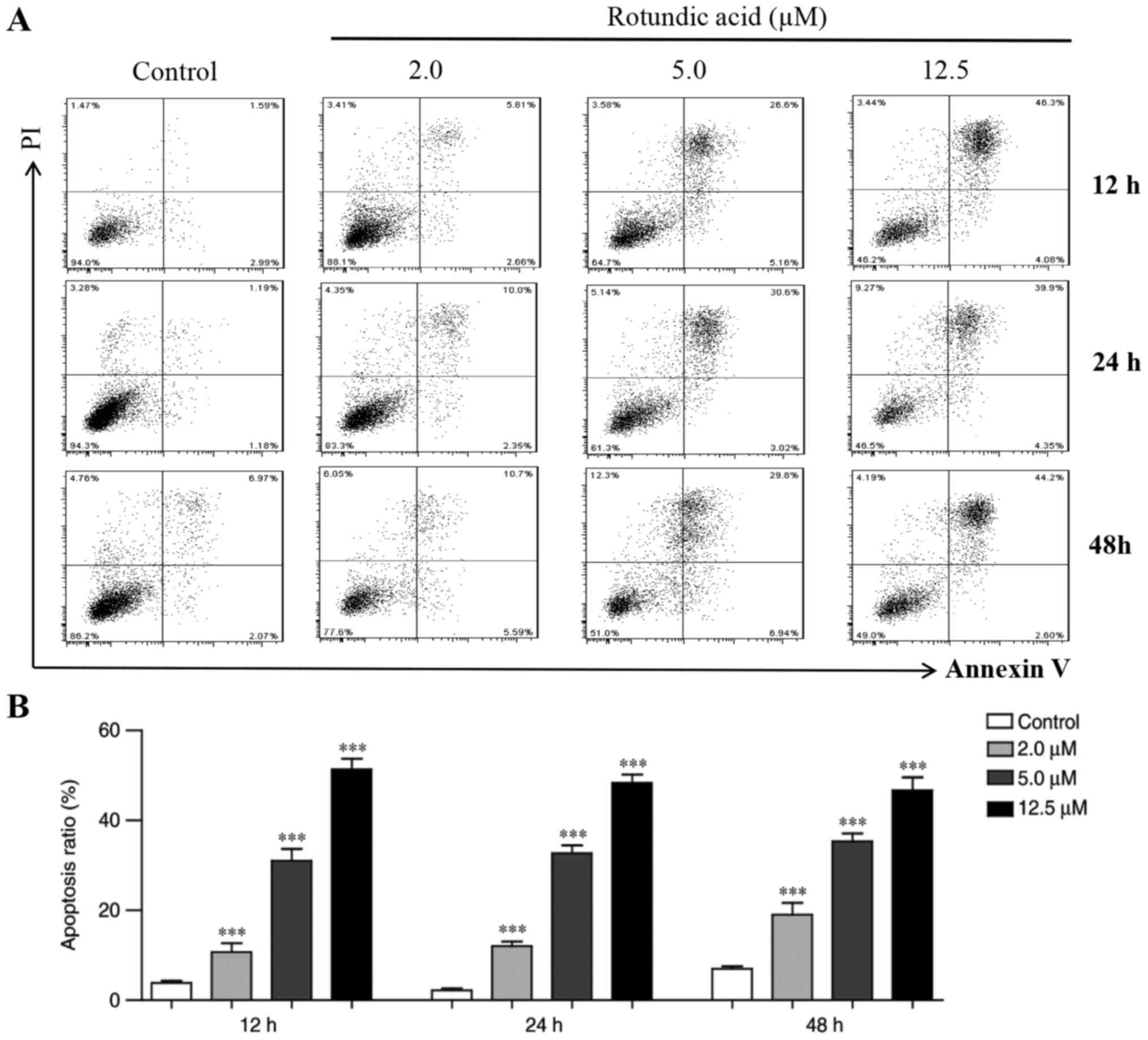
Annexin V-FITC and PI double-staining
assay
To investigate whether RA also induced apoptosis in
Cas3-MCF-7 cells, an annexin V-FITC and PI double-staining assay
was performed. Cas3-MCF-7 cells were treated with RA at
concentrations of 2.0, 5.0 and 12.5 µmol/l for 12, 24 and 48 h, and
were then analyzed by flow cytometry. Fig. 3A indicates that, compared with that in
the control group, the number of the early- and late-apoptotic
cells increased significantly in the RA-treated groups. The
inhibitory effects of RA on the Cas3-MCF-7 cells were
dose-dependent within the range of 2–12.5 µmol/l. Treatment with RA
at 12.5 µmol/l elicited the greatest apoptosis capacity, which
reached 52.8% at 12 h, 50.2% at 24 h and 49.6% at 48 h (Fig. 3B). Similar to the MTT assay results,
there were no significant differences in the proportion of
apoptotic cells between the different time points at 12 to 48 h,
which revealed a similar inhibitory effect at all three doses.
These results indicate that RA significantly induced Cas3-MCF-7
cell apoptosis.
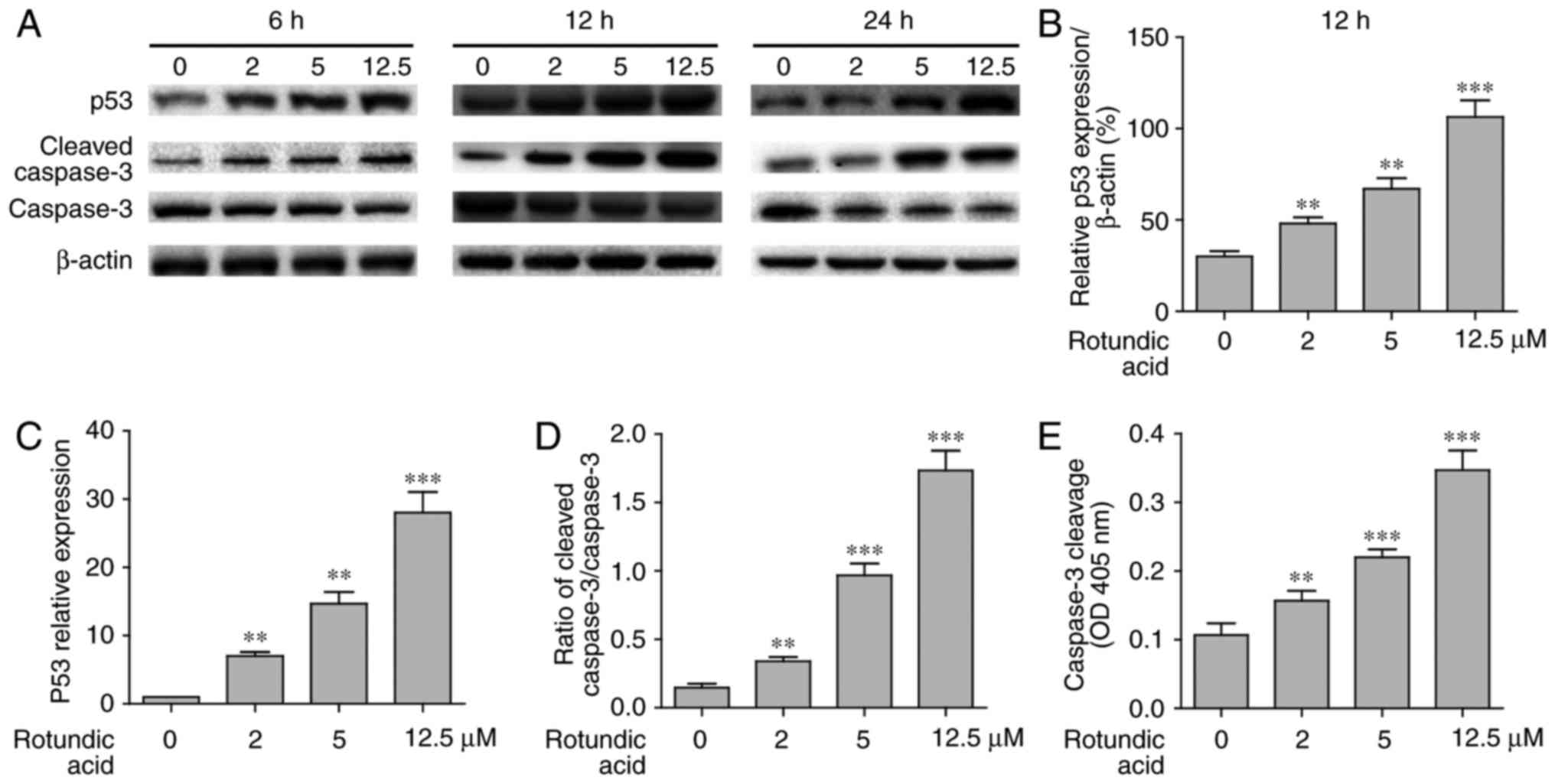
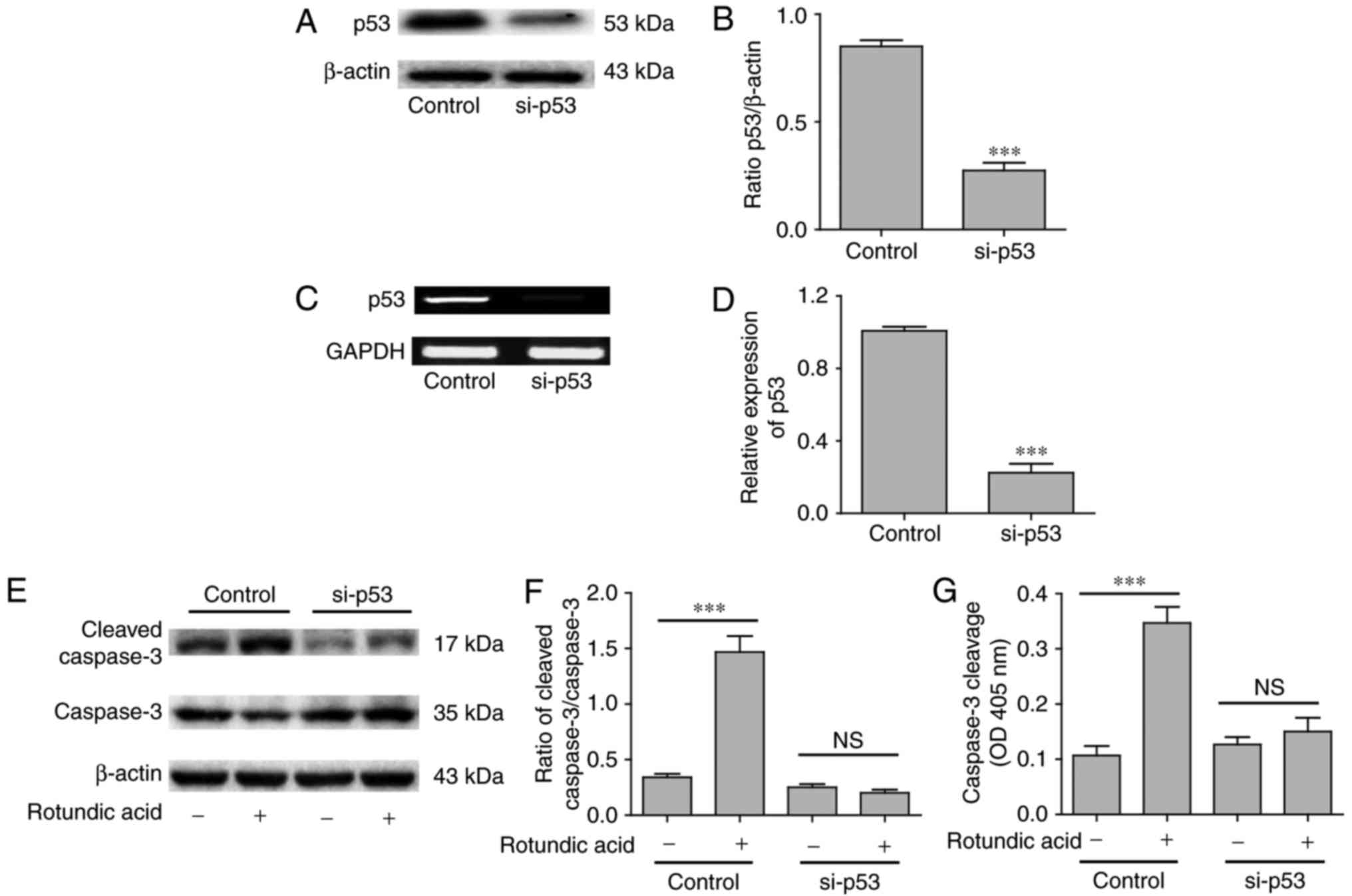
RA induces Cas3-MCF-7 cell apoptosis
via the p53 signaling pathway
Cas3-MCF-7 cells were treated with RA at a
concentration of 2–12.5 µmol/l for 6, 12 and 24 h. p53, cleaved
caspase-3 and total caspase-3 levels were detected by western
blotting, RT-qPCR or ELISA. As presented in Fig. 4A, the expression of p53 induced by RA
in the Cas3-MCF-7 cells was dose-dependent within the range of
2–12.5 µmol/l at 6, 12 and 24 h. However, only the expression of
p53 at 24 h demonstrated the peak value. Quantification of p53
expression at 12 h is presented in Fig.
4B. The RT-qPCR analysis confirmed that p53 gene expression was
also induced by RA in a dose-dependent manner (Fig. 4C). The relative expression of the
cleaved caspase-3/caspase-3 may reflect the molecular mechanism
underlying apoptosis. The caspase-3 cell apoptosis signal that was
activated by RA in the Cas3-MCF-7 cells also exhibited a
dose-dependent effect within the range of 2–12.5 µmol/l (Fig. 4D). The caspase-3 activity was also
confirmed by ELISA (Fig. 4E). To
further clarify the association between p53 and the caspase-3
activation in Cas3-MCF-7 cell apoptosis, RNA interference (RNAi)
technology was applied to inhibit p53 expression in Cas3-MCF-7
cells. p53 expression in the Cas3-MCF-7 cells was significantly
decreased following p53 gene silencing according to western blot
analysis (P<0.001; Fig. 5A and B)
and RT-qPCR (P<0.001; Fig. 5C and
D) analyses. When the p53 gene was silenced, RA could not
induce Cas3-MCF-7 cell caspase-3 activity (P>0.05; Fig. 5E-G), which was confirmed by western
blotting and ELISA. These results indicate that RA induced
Cas3-MCF-7 cell apoptosis via the p53 signaling pathway.
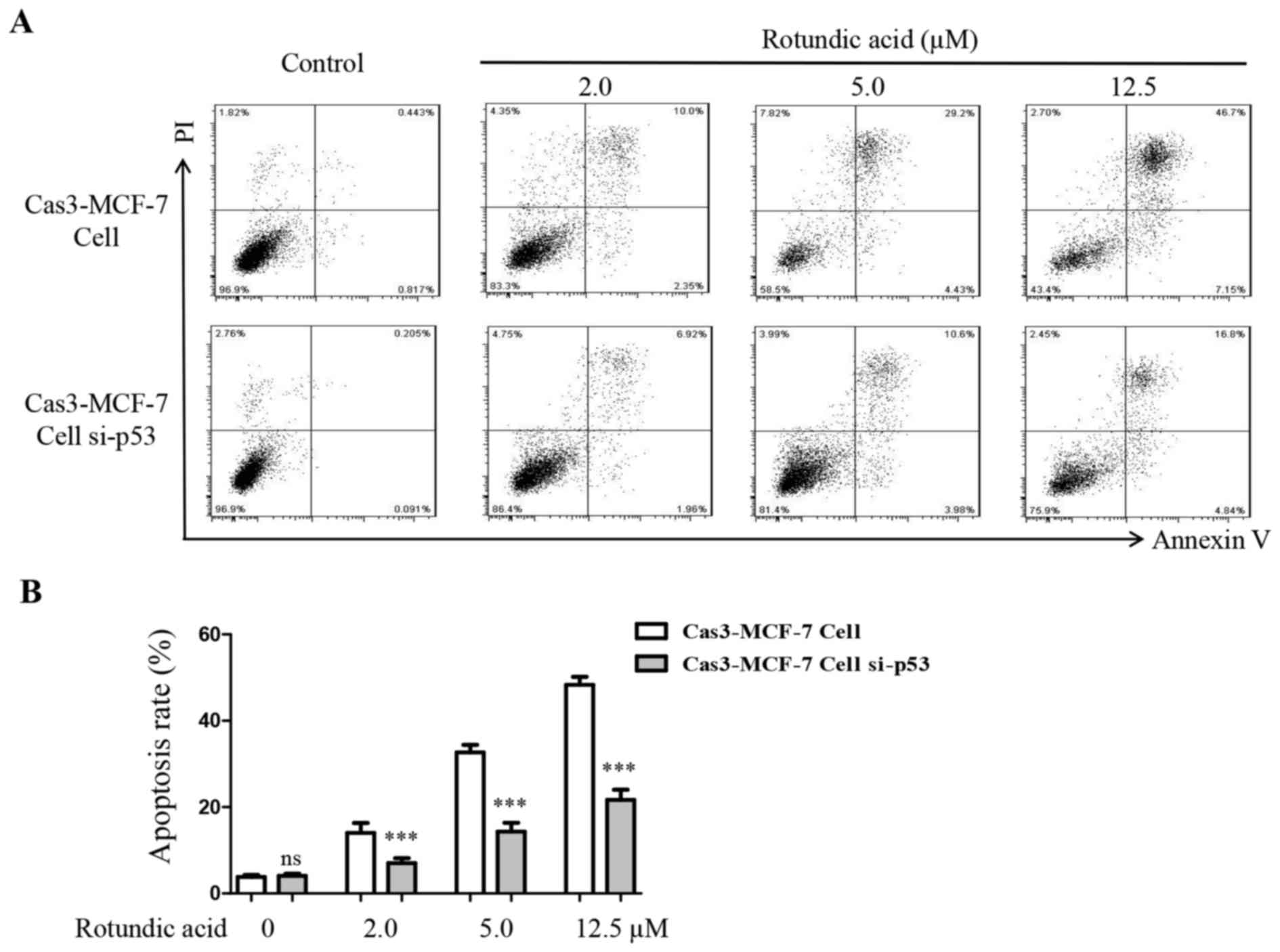
Silencing of p53 decreases RA-induced
Cas3-MCF-7 cell apoptosis
An annexin V-FITC and PI double-staining assay was
also performed to investigate the outcome when the p53 gene was
silenced in Cas3-MCF-7 cells following treatment with RA. The
results demonstrated that the numbers of early- and late-apoptotic
cells decreased significantly on the p53-silenced Cas3-MCF-7 cells
comparing with untreated Cas3-MCF-7 cells when treated with RA at
all three doses (P<0.001; Fig. 6A and
B). These results indicated that p53 serves an important
function in RA-induced Cas3-MCF-7 cell apoptosis.
Discussion
Breast cancer is a common malignant cancer that has
threatened the health of women worldwide over the last three
decades (3). There are numerous
factors that affect the biology of tumor cells during the
occurrence and progression of breast cancer (24). Oncogenes have become increasingly
important in the development of cancer. Caspase-3 deficiency may
contribute to the chemotherapy-resistance of breast cancers.
Reconstitution of caspase-3 sensitizes MCF-7 breast cancer cells to
chemotherapy. Breast cancer metastasis and prognosis are associated
with oncogene mutations and abnormal expression (25). p53 is an important tumor suppressor
gene that resides in the majority of normal cells; however, it is
often mutated in cancer cells (26,27).
Previous studies have demonstrated that p53 gene mutations are
associated with the resistance of breast cancer (28,29). Lee
et al (15) also reported that
RA demonstrated cytotoxicity, with an IC50 value of 9.5
µM when applied to the MCF-7 cell lines; however, the precise
mechanism has not been thoroughly studied and elucidated. The
results of the present study are inconsistent with those of the
study by Lee et al (15), as
it was revealed that RA had limited function on the inhibition of
the proliferation of MCF-7 cells treated with control plasmid, but
the inhibitory effects of RA on the Cas3-MCF-7 cells were
dose-dependent within the range of 2–12.5 µmol/l. This may be due
to the transfection treatment of MCF-7 cells, or the different
sources and the purity of the RA. However, this phenomenon
indicates that caspase-3 serves a critical function in rotundic
acid-induced apoptosis, and suggests that caspase-3 deficiency may
contribute to the chemotherapy resistance of breast cancer
(30).
Although there are numerous studies pertaining to
the extraction of RA in China, there have been few studies on its
bioactivity due to a lack of interest from pharmacological
researchers (15,22). In our open patent, a considerable
quantity of RA was isolated and purified from I. rotunda
(31). As RA has the potential to be
a native anticancer drug with sufficient sources, our research
group has investigated and applied for a series of patents
regarding RA and its derivatives over the last few years to
investigate and use (32–36). The aim of the present study was to
investigate the anticancer effect of RA on cell viability and
apoptosis in the human breast cancer Cas3-MCF-7 cell line. RA was
able to inhibit the viability of Cas3-MCF-7 cells at 2.0–12.5 µM
between 12 and 48 h. RA enhanced the apoptotic effector of
caspase-3 activity and induced cell death. The RT-qPCR analysis and
western blot assay confirmed that RA induced p53 gene and protein
expression in a dose-dependent manner, which was consistent with
caspase-3 activity. Caspase-3 activation could be initiated by a
number of upstream signal-regulated molecules (37–39). The
increase in p53 gene expression induced cell apoptosis via caspase
activity. In the present study, p53 was demonstrated to be
increased in Cas3-MCF-7 cells treated with RA, indicating that RA
induced p53 and caspase-3 activity in Cas3-MCF-7 cells, which then
caused cell death.
To confirm the importance of the p53 pathway in the
caspase-3 activation and RA-induced Cas3-MCF-7 cell apoptosis, RNAi
technology was applied to inhibit p53 expression in MCF-7 cells.
Subsequent to using a p53 siRNA kit to treat Cas3-MCF-7 cells for
24 h, the expression levels of the p53 protein and gene were
significantly decreased, which were then detected by western blot
analysis and RT-qPCR assays. Flow cytometric analysis revealed that
the numbers of early and late apoptotic cells decreased
significantly compared with those of normal MCF-7 cells at 2.0, 5.0
and 15.5 µM RA. On the basis of these results, it was concluded
that RA exerted antitumor functions by inducing p53/caspase-3
activation in target cells. In summary, the results of the present
study identified that RA functioned to inhibit Cas3-MCF-7 cell
viability by inducing apoptosis. To the best of our knowledge, the
present study is the first to reveal the exact mechanism of RA in
the induction of Cas3-MCF-7 cell apoptosis. Caspase-3 serves an
important function in the execution of apoptotic. Caspase-3
deficiency or downregulation has been reported in breast and other
types of cancer. Although caspase-3 is deficient in MCF-7 cells, it
has been demonstrated that caspase-3 reconstitution significantly
enhanced radiation-induced apoptosis, with a decrease in the
survival fraction, an increase in caspase activation, cleavage of
cellular death substrates and mitochondrial depolarization
(40). Caspase-3 reconstitution may
be used as a gene targeting therapy combined with chemotherapy or
radiotherapy for the treatment of caspase-3-deficient cancer.
Collectively, the results of the present study indicate that
caspase-3 serves a critical function in RA-induced apoptosis, and
suggest that caspase-3 deficiency may contribute to the
chemotherapy-resistance of breast cancers. Reconstitution of
caspase-3 sensitizes MCF-7 breast cancer cells to chemotherapy. RA
has the potential for development as a new drug in combination with
gene therapy for the treatment of human breast cancer with
caspase-3 deficiency.
Acknowledgements
Not applicable.
Funding
The present study was supported in part by the Jilin
Provincial Natural Science Foundation of China (grant nos.
20140520014JH and 20180101135JC), the Interdisciplinary Chemistry
and Medicine Foundation of Jilin University (grant no. JDYYJCHX004)
and the National Natural Science Foundation of China (grant no.
31470418).
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
MN, XW and HL planned and performed the experiments,
analyzed data and wrote the manuscript. DY, WS and HX performed the
experiments. YH and QZ designed, interpreted and funded the study,
and wrote the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Metzger-Filho O, de Azambuja E, Bradbury
I, Saini KS, Bines J, Simon SD, Dooren VV, Aktan G, Pritchard KI,
Wolff AC, et al: Analysis of regional timelines to set up a global
phase III clinical trial in breast cancer: The adjuvant lapatinib
and/or trastuzumab treatment optimization experience. Oncologist.
18:134–140. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bower JE, Greendale G, Crosswell AD, Garet
D, Sternlieb B, Ganz PA, Irwin MR, Olmstead R, Arevalo J and Cole
SW: Yoga reduces inflammatory signaling in fatigued breast cancer
survivors: A randomized controlled trial. Psychoneuroendocrinology.
43:20–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pu Z, Zhang X, Chen Q, Yuan X and Xie H:
Establishment of an expression platform of OATP1B1 388GG and 521CC
genetic polymorphism and the therapeutic effect of tamoxifen in
MCF-7 cells. Oncol Rep. 33:2420–2428. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hu D, Su C, Jiang M, Shen Y, Shi A, Zhao
F, Chen R, Shen Z, Bao J and Tang W: Fenofibrate inhibited
pancreatic cancer cells proliferation via activation of p53
mediated by upregulation of LncRNA MEG3. Biochem Biophys Res
Commun. 471:290–295. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bertheau P, Lehmann-Che J, Varna M, Dumay
A, Poirot B, Porcher R, Turpin E, Plassa LF, de Roquancourt A,
Bourstyn E, et al: p53 in breast cancer subtypes and new insights
into response to chemotherapy. Breast. 2 Suppl 22:S27–S29. 2013.
View Article : Google Scholar
|
|
6
|
Sana M and Malik HJ: Current and emerging
breast cancer biomarkers. J Cancer Res Ther. 11:508–513. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Shah U, Shah R, Acharya S and Acharya N:
Novel anticancer agents from plant sources. Chin J Nat Med.
11:16–23. 2013.(In Chinese). View Article : Google Scholar
|
|
8
|
Khan F, Ahmed F, Pushparaj PN, Abuzenadah
A, Kumosani T, Barbour E, AlQahtani M and Gauthaman K: Ajwa date
(Phoenix dactylifera L.) extract inhibits human breast
adenocarcinoma (MCF7) cells in vitro by inducing apoptosis and cell
cycle arrest. PLoS One. 11:e01589632016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gerber DE: Targeted therapies: A new
generation of cancer treatments. Am Fam Physician. 77:311–319.
2008.PubMed/NCBI
|
|
10
|
He YF, Nan ML, Zhao YW, Sun WY, Li W and
Zhao QC: Design, synthesis and evaluation of antitumor activity of
new rotundic acid acylhydrazone derivatives. Z Naturforsch C.
71:95–103. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Haraguchi H, Kataoka S, Okamoto S, Hanafi
M and Shibata K: Antimicrobial triterpenes from Ilex integraand the
mechanism of antifungal action. Phytother Res. 13:151–156. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhao WM, Wolfender JL, Hostettmann K,
Cheng KF, Xu RS and Qin GW: Triterpenes and triterpenoid saponins
from mussaenda pubescens. Phytochemistry. 45:1073–1078. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bhattacharyya J and Almeida MD: Isolation
of the constituents of the root-bark of guettarda platypoda. J Nat
Prod. 48:1481985. View Article : Google Scholar
|
|
14
|
Saimaru H, Orihara Y, Tahsakul P, Kang YH,
Shibuya M and Ebizuka Y: Production of triterpene acids by cell
suspension cultures of Olea europaea. Chem Pharm Bull (Tokyo).
55:784–788. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee TH, Juang SH, Hsu FL and Wu CY:
Triterpene acids from the leaves of planchonella duclitan (Blanco)
bakhuizan. J Chin Chem Soc. 52:1275–1280. 2005. View Article : Google Scholar
|
|
16
|
Fulda S: Inhibitor of apoptosis (IAP)
proteins as therapeutic targets for radiosensitization of human
cancers. Cancer Treat Rev. 38:760–766. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yin J, Wang F, Kong Y, Wu R, Zhang G, Wang
N, Wang L, Lu Z and Liang M: Antithrombin III prevents progression
of chronic kidney disease following experimental
ischaemic-reperfusion injury. J Cell Mol Med. 21:3506–3514. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu Z, Cheng D, Yin J, Wu R, Zhang G, Zhao
Q, Wang N, Wang F and Liang M: Antithrombin III protects against
contrast-induced nephropathy. EBioMedicine. 17:101–107. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hengartner MO: The biochemistry of
apoptosis. Nature. 407:770–776. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Antony ML, Kim SH and Singh SV: Critical
role of p53 upregulated modulator of apoptosis in benzyl
isothiocyanate-induced apoptotic cell death. PLoS One.
7:e322672012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xu R: Studies on the chemical components
and antitumor activity of ilex rotunda thunb. Ph.D. Thesis.
Guangzhou University of Chinese Medicine; 2009, (In Chinese).
|
|
22
|
He YF, Nan ML, Sun JM, Meng ZJ, Yue FG,
Zhao QC, Yang XH and Wang H: Synthesis, characterization and
cytotoxicity of new rotundic acid derivatives. Molecules.
17:1278–1291. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mrózek E, Layman R, Ramaswamy B, Lustberg
M, Vecchione A, Knopp MV and Shapiro CL: Phase II trial of
neoadjuvant weekly nanoparticle albumin-bound paclitaxel,
carboplatin, and biweekly bevacizumab therapy in women with
clinical stage II or III HER2-negative breast cancer. Clin Breast
Cancer. 14:228–234. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zuo S, Liu C, Wang J, Wang F, Xu W, Cui S,
Yuan L, Chen X, Fan W, Cui M and Song G: IGFBP-rP1 induces p21
expression through a p53-independent pathway, leading to cellular
senescence of MCF-7 breast cancer cells. J Cancer Res Clin Oncol.
138:1045–1055. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Steigerwald C, Rasenberger B, Christmann M
and Tomicic MT: Sensitization of colorectal cancer cells to
irinotecan by the survivin inhibitor LLP3 depends on XAF1
proficiency in the context of mutated p53. Arch Toxicol.
92:2645–2648. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Meng X, Bi J, Li Y, Yang S, Zhang Y, Li M,
Liu H, Li Y, Mcdonald ME, Thiel KW, et al: AZD1775 increases
sensitivity to olaparib and gemcitabine in cancer cells with p53
mutations. Cancers (Basel). 10:E1492018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shokouh TZ, Ezatollah A and Barand P:
Interrelationships Between Ki67, HER2/neu, p53, ER, and PR status
and their associations with tumor grade and lymph node involvement
in breast carcinoma subtypes: Retrospective-observational
analytical study. Medicine (Baltimore). 94:e13592015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Verma S and Rao BJ: p53 suppresses
BRCA2-stimulated ATPase and strand exchange functions of human
RAD51. J Biochem. 154:237–248. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wesierska-Gądek J, Hackl S, Zulehner N,
Maurer M and Komina O: Reconstitution of human MCF-7 breast cancer
cells with caspase-3 does not sensitize them to action of CDK
inhibitors. J Cell Biochem. 112:273–288. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhao QC, Nan ML, He YF and Chen SW:
Application of rotundic acid in the cardiovascular disease
prevention. Chn 201010204596.9. 2010.(In Chinese).
|
|
32
|
Zhao QC, Nan ML, He YF and Chen SW:
Application of rotundic acid in the preparation of lipid-lowering
drugs. CHN 201010204607. 3:2010.(In Chinese).
|
|
33
|
He YF, Zhao QC, Nan ML, Wang HL, Ma JS,
Zhao YW, et al: Application of rotundic acid and its derivatives in
the preparation of anticancer drugs. CHN 201010607515. x:2010.(In
Chinese).
|
|
34
|
Nan ML, Zhao QC, He YF, Chen SW, Zhao YW
and Wang LP: Pharmaceutical compositions from ilex rotunda thunb.
and Its Application. CHN 201010607550. 1:2010.(In Chinese).
|
|
35
|
Zhao QC, He YF, Nan ML, Chen SW, Zhao YW
and Wang LP: Synthesis method of rotundic acid derivatives and
their application in the preparation of cardiovascular disease
prevention drugs. CHN 20110030007. 4:2011.(In Chinese).
|
|
36
|
He YF, Nan ML, Zhao QC, Zhao YW and Yue
FG: Application of amino acid modified rotundic acid derivatives in
the preparation of anticancer drugs. CHN 201110351365.5. 2011.(In
Chinese).
|
|
37
|
Zhang M, Yan H, Li S and Yang J:
Rosmarinic acid protects rat hippocampal neurons from cerebral
ischemia/reperfusion injury via the Akt/JNK3/caspase-3 signaling
pathway. Brain Res. 1657:9–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Venkatesan RS and Sadiq AM: Effect of
morin-5′-sulfonic acid sodium salt on the expression of apoptosis
related proteins caspase 3, Bax and Bcl 2 due to the mercury
induced oxidative stress in albino rats. Biomed Pharmacother.
85:202–208. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mondal A and Bennett LL: Resveratrol
enhances the efficacy of sorafenib mediated apoptosis in human
breast cancer MCF7 cells through ROS, cell cycle inhibition,
caspase 3 and PARP cleavage. Biomed Pharmacother. 84:1906–1914.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang XH, Edgerton S and Thor AD:
Reconstitution of caspase-3 sensitizes MCF-7 breast cancer cells to
radiation therapy. Int J Oncol. 26:1675–1680. 2005.PubMed/NCBI
|