Introduction
Non-small cell lung cancer (NSCLC) is the leading
cause of lung cancer-associated mortality (1). It has been reported that NSCLC
constitutes nearly 90% of lung cancer (2). Since the majority of patients diagnosed
with NSCLC are in an advanced stage, the therapeutic outcomes for
NSCLC are not optimal (3). The use of
chemotherapy has enhanced the treatment of NSCLC over the past 50
years (4). In particular,
platinum-based chemotherapy, which is associated with improved
survival, is currently the standard therapeutic treatment for
patients with advanced NSCLC (5).
However, a number of patients with NSCLC develop resistance to
platinum-based chemotherapy, resulting in unsatisfactory treatment
outcomes. It has also been reported that a number of molecules have
anti-cancer effects on NSCLC, while few patients have advanced
sites that are accessible via relatively non-invasive biopsy
procedures (6). Therefore, it is
necessary to develop novel chemotherapeutic drugs for the treatment
of NSCLC.
Methylxanthine (MX) is a naturally occurring
compound in animals and plants (7,8). As
reported in previous studies, MX may prevent the proliferation of a
number of tumor cells, including oral cancer (9), breast cancer (10) and lung cancer (11,12). In
addition, it has been demonstrated that caffeine, a class of MX,
blocks the proliferation of hepatocellular carcinoma and pancreatic
cancer cell lines by inhibiting the phosphoinositide 3-kinase
(PI3K)/RAC-α serine/threonine-protein kinase (AKT) pathway
(13). Similarly, theophylline,
another MX derivative, possesses the capacity to inhibit the
proliferation and migration of melanoma cancer cells (14). Diprophylline (DPL), known as a novel
MX derivative with fewer adverse effects, is usually employed for
relieving the clinical symptoms of and preventing bronchial asthma
or other respiratory diseases (15,16). In
previous studies, there have been limited reports on the role of
DPL in the treatment of cancer, particularly NSCLC (17). Therefore, it may be beneficial to
investigate the ability of DPL to inhibit the cell proliferation
and migration of A549 cells.
In the present study, the role of DPL in the growth,
invasion, migration and apoptosis of NSCLC A549 cells was studied.
Additionally, a potential molecular pathway was also detected
following DPL treatment in A549 cells.
Materials and methods
Cell culture
The human lung adenocarcinoma cell line A549 and
normal human bronchial epithelial cells (HBEs) were purchased from
the American Type Culture Collection (Manassas, VA, USA). The cells
were incubated in RPMI-1640 (HyClone; GE Healthcare Life Sciences,
Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS;
Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
penicillin (100 U/ml) and streptomycin (0.1 mg/ml). Cells were
cultured at 37°C in an incubator containing 5% CO2.
Cells in the exponential growth phase (~1×106 cells/ml)
were used for the following experiments.
Cell proliferation assay
Following the manufacturer's instructions, cell
proliferation was determined using the Cell Counting Kit-8 (CCK-8;
Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China). Following culture for 72 h, A549 and HBE cells were treated
with DPL (MedChemExpress USA, Monmouth Junction, NJ, USA) at
varying concentrations (0, 0.1, 1, 10 and 100 µM) with 0 µM DPL as
the control. In detail, ~1×103 cells/well were seeded
into 96-well plates and cultured in 100 µl RPMI-1640 medium
supplemented with 10% FBS overnight. Based on the effects of
different concentrations in the A549 and HBE cells, 10 µM DPL was
chosen as the effective concentration to be used in subsequent
experiments. To exclude the effects of cell death on cell
proliferation, migration, invasion and apoptosis, 1 µM DPL was also
added, in order to perform cell functional analysis, including cell
proliferation, migration, invasion and apoptosis. Cells were
incubated with 1 or 10 µM DPL and 1% DMSO culture medium as the
normal control group (NC). For performing the CCK-8 assay, 10 µl
CCK-8 reagent was added to each well and the plates were incubated
at 37°C for 1.5 h. The cell viability was measured every 24 h, and
the optical density (OD) was measured at 450 nm using a microplate
reader.
Transwell migration and invasion
assay
Cell migratory and invasive abilities were assessed
using 24-well Transwell chambers (EMD Millipore, Billerica, MA,
USA) with membrane pore size of 8.0 µm. In detail, 100 µl
serum-free medium with ~1×105 cells was plated in the
upper chambers of the Transwell plates, while 500 µl medium
containing 10% FBS was added to the lower chamber. Following
incubation at 37°C with 5% CO2 overnight, non-migrating
cells on the top chamber were scraped off with cotton-tipped swabs,
and cells that had migrated through the membrane were fixed with 4%
paraformaldehyde at room temperature for 30 min. Cells were stained
with 0.1% crystal violet at room temperature for 20 min.
Subsequently, the migrated cells were counted under a microscope at
×200 magnification using five randomly selected visual fields. For
cell invasion detection, Transwell membranes were precoated with
Matrigel® (BD Biosciences, San Jose, CA, USA) at 37°C
for 30 min prior to the experiment.
Flow cytometry apoptosis assay
Following the manufacturer's protocol, apoptotic
A549 cells were evaluated using an Annexin V-fluorescein
isothiocyanate (Annexin V-FITC)/propidium iodide (PI) Apoptosis
Detection kit (Beijing 4A Biotech Co., Ltd., Beijing, China). A549
cells were treated with DPL and harvested by trypsinization without
EDTA, washed twice with cold PBS, centrifuged at 560 × g at 4°C for
5 min and the supernatant was discarded. The cell pellet was
resuspended in 1X binding buffer and the cell density adjusted to
1–5×106 cells/ml. A total of 5 µl FITC-conjugated
Annexin V and 10 µl PI were added and incubated for 5 min at room
temperature in the dark. Following the addition of PI (10 µl) and
PBS (400 µl), cells were analyzed using a flow cytometer and the
data analyzed using FlowJo 7.6 software (FlowJo LLC, Ashland, OR,
USA).
Western blot analysis
Following treatment with DPL or vehicle for 24 h,
total protein was extracted from cells using
radioimmunoprecipitation assay buffer (CW Biotech, Co., Ltd.,
Beijing, China). Following lysis for 30 min on ice, the lysates
were centrifuged at 13,400 × g at 4°C for 10 min. The Bicinchoninic
Acid Protein Assay kit (CW Biotech, Co., Ltd.) was used to
determine the protein concentration. A total of 20 µg protein/lane
was separated via SDS-PAGE on an 8–10% Tris-glycine gradient gel.
Subsequently, the proteins were transferred onto a polyvinylidene
diflouride membrane for 2 h and blocked with 5% non-fat milk in TBS
containing 0.1% Tween-20 (pH 7.4) at room temperature for 1 h. The
membranes were incubated with primary antibodies (1:1,000 dilution)
against AKT (ab18785), phosphorylated (p)-AKT (ab38449),
serine/threonine-protein kinase mTOR (mTOR) (ab87540), p-mTOR
(ab131538), Cyclin D1 (ab226977), p-ribosomal protein S6 kinase β-1
(p70S6K) (ab186753), apoptosis regulator Bcl-2 (Bcl-2) (ab196495),
apoptosis regulator BAX (Bax) (ab53154) and active Caspase-3
(ab32042), and against GAPDH (1:5,000 dilution) (ab181603), at 4°C
overnight, and were detected using goat anti-rabbit/mouse
horseradish peroxidase-conjugated secondary antibodies
(ab6721/ab6789, 1:5,000 dilution). Primary and secondary antibodies
were from Abcam (Cambridge, MA, USA). Western blot bands were
visualized using an enhanced chemiluminescence reagent system (PTG
Company; USA) and were quantified using Quantity One 4.6.7 software
(Bio-Rad Laboratories, Hercules, CA, USA).
Statistical analysis
Statistical analysis was performed using SPSS 18.0
(SPSS, Inc., Chicago, IL, USA) and GraphPad Prism 5 software
(GraphPad Software, Inc., La Jolla, CA, USA). Measurement data are
expressed as the mean ± standard deviation. Means were compared
using the Student's t-test and one-way Analysis of Variance
(ANOVA). For one-way ANOVA, post hoc testing was performed using
Dunnett's test. P<0.05 was considered to indicate a
statistically significant difference.
Results
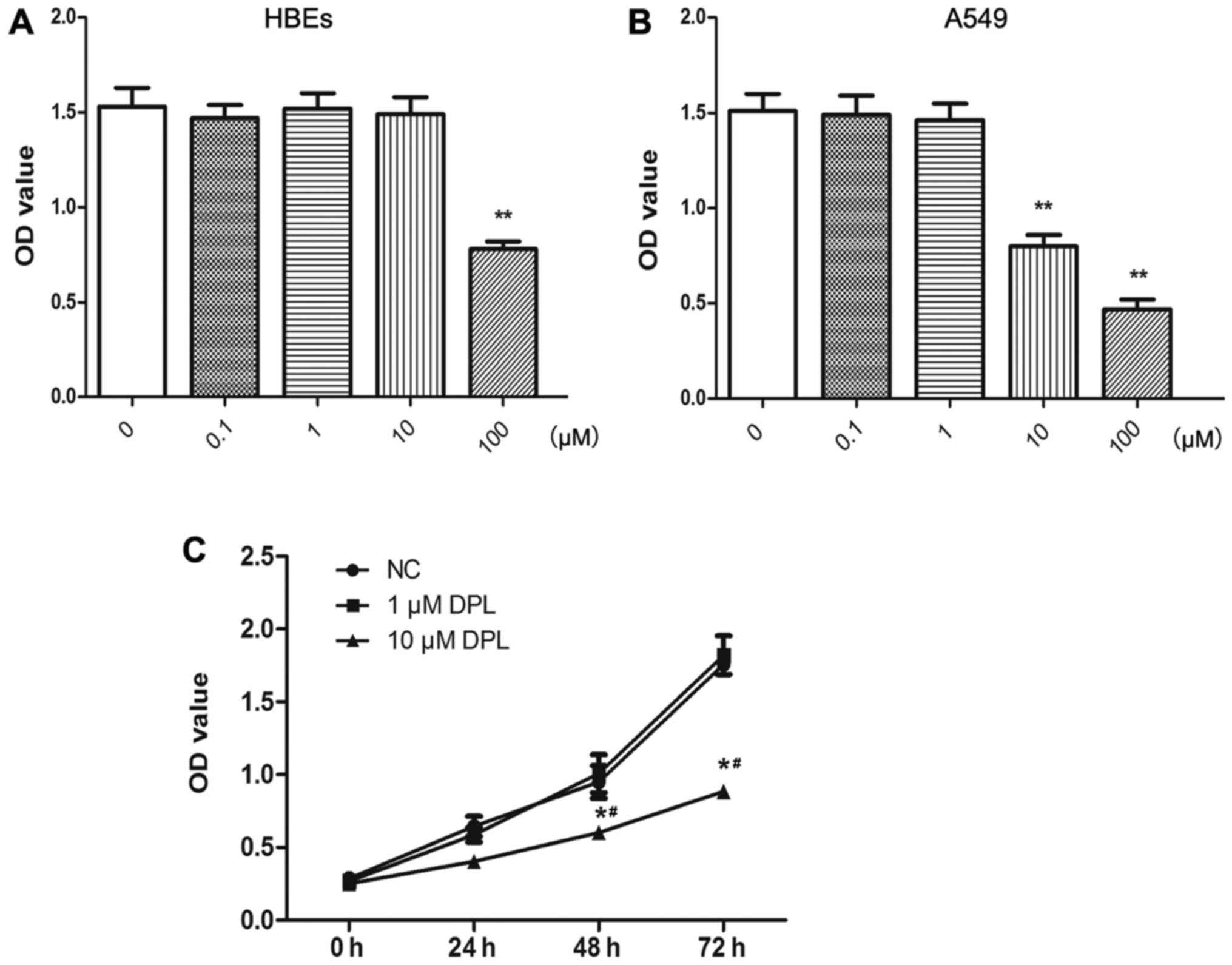
DPL inhibits the proliferation of
NSCLC A549 cells
The effect of DPL on the proliferation of A549 cells
was analyzed using the CCK-8 assay. At concentrations of DPL >10
µM, the viability of HBEs was inhibited (Fig. 1A). The viability of A549 cells was
decreased in a dose-dependent manner at DPL >1 µM (Fig. 1B). Therefore, 10 µM DPL was selected
as the concentration to be used in subsequent experiments. To
exclude the effect of cell death on cell proliferation, migration,
invasion and apoptosis, experiments using 1 µM DPL were also
performed for cell functional analysis, including cell
proliferation, migration, invasion and apoptosis. As presented in
Fig. 1C, the proliferation of A549
cells treated with DPL decreased in a time-dependent manner,
compared with the control group, and the OD values were
significantly decreased at 48 and 72 h, compared with the control
(P<0.05). These results demonstrated that the proliferative
abilities of A549 cells decreased notably following treatment with
DPL.
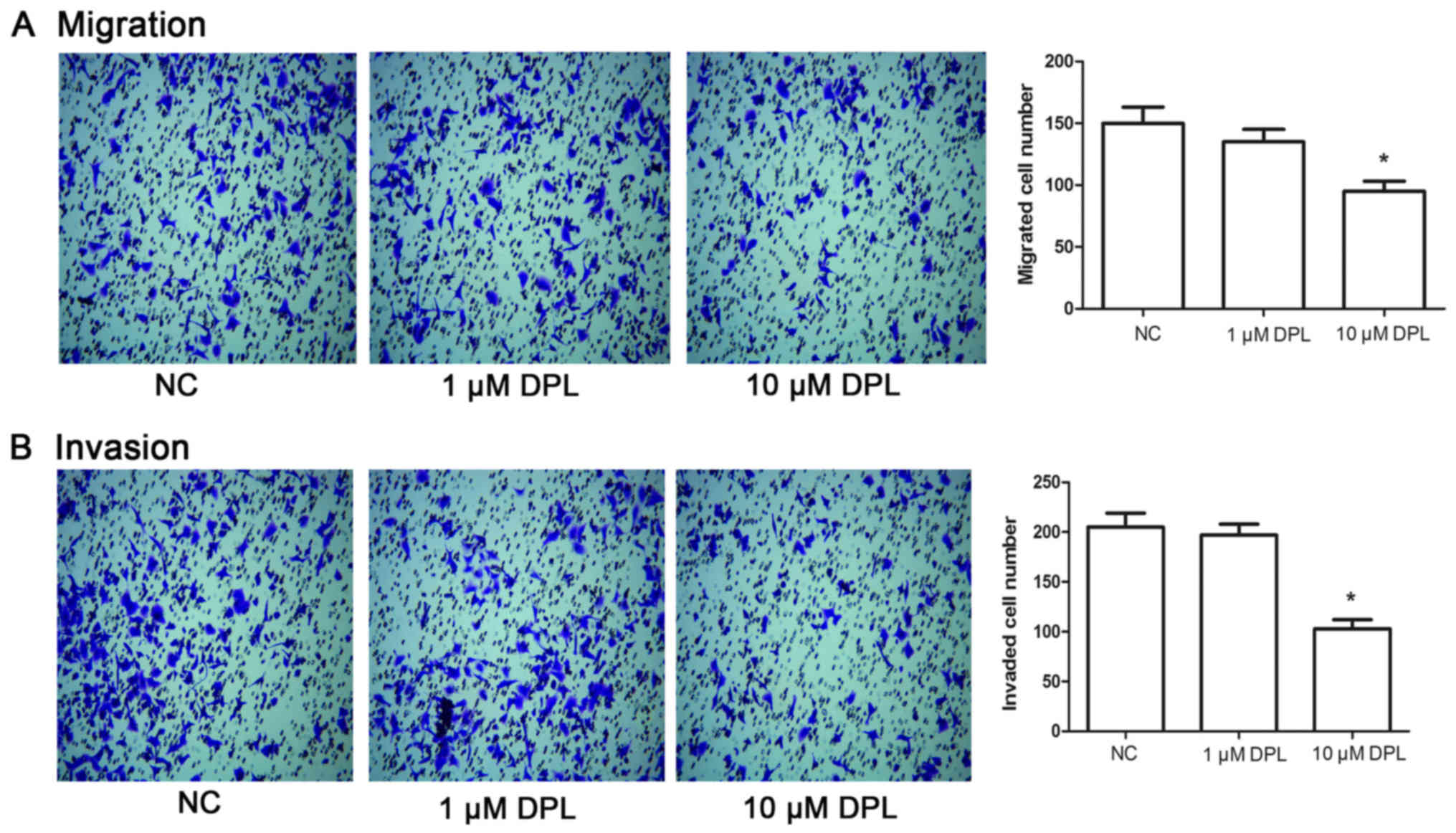
DPL inhibits the migration and
invasion of NSCLC A549 cells
Following 24 h treatment with DPL, Transwell assays
were performed to determine the potential effect of DPL on cell
migration and invasion in A549 cells. As presented in Fig. 2A, following treatment with 10 µM DPL,
the number of migrated cells was markedly decreased compared with
the NC groups (P<0.05), while there were no detectable
alterations in migrated cell numbers between the NC and 1 µM DPL
groups. Likewise, the invaded cell number in the group treated with
10 µM DPL was significantly decreased compared with the NC group
(Fig. 2B; P<0.05), while cells
treated with 1 µM DPL displayed no significant difference compared
with the NC group. These results indicated that DPL may
significantly inhibit migration and invasion in A549 cells.
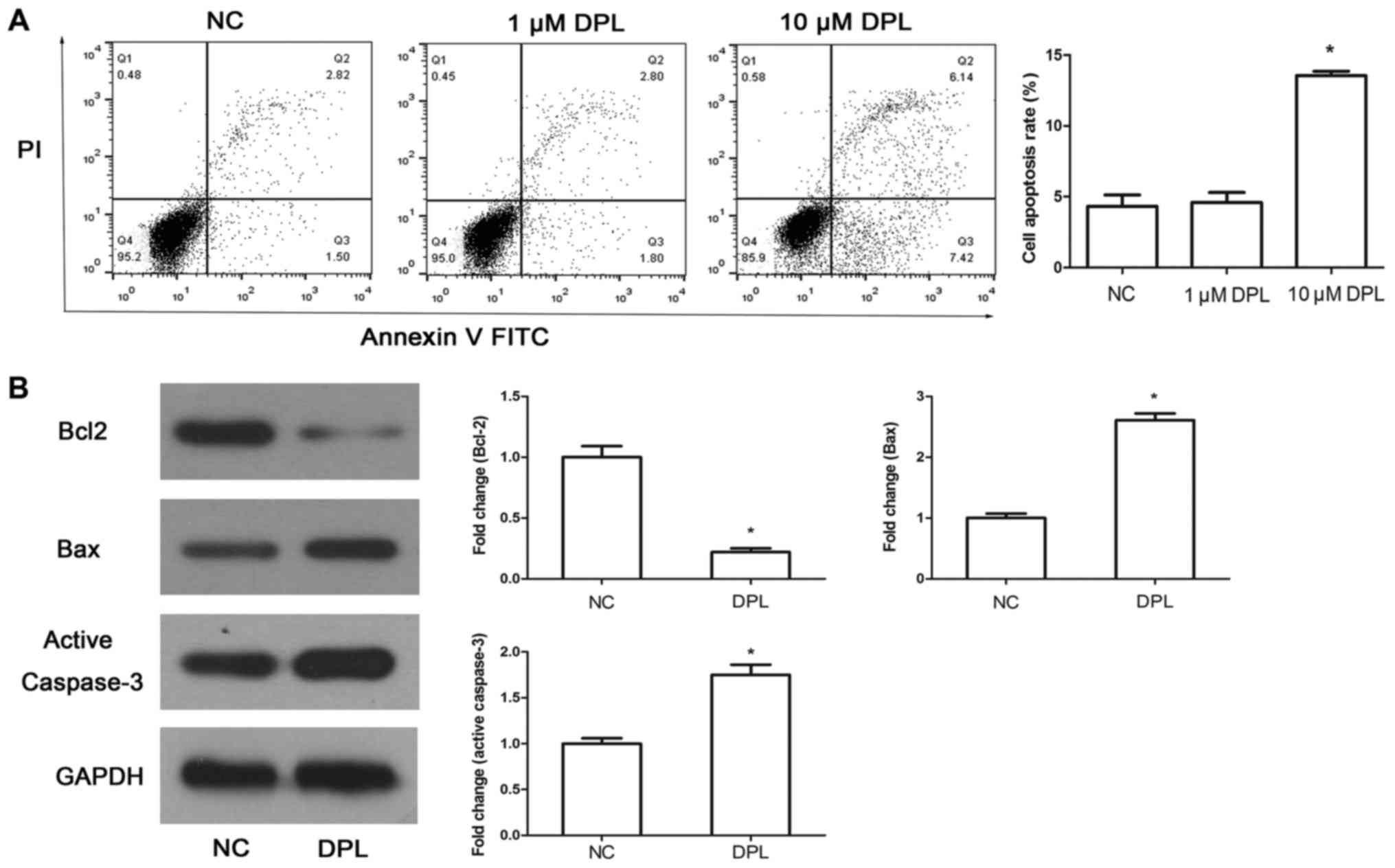
DPL promotes the apoptosis of NSCLC
A549 cells
To determine whether DPL could induce A549 cell
apoptosis, Annexin-V-FITC/PI was used following 24 h treatment with
DPL. The results demonstrated that the apoptosis rate in cells
treated with 10 µM DPL was significantly increased compared with
the NC group (Fig. 3A; P<0.05)
while there were no detectable alterations between cells treated
with 1 µM DPL and the NC group (Fig.
3A; P>0.05). Subsequently, to study the mechanism mediating
DPL-induced apoptosis, a western blot assay was used to determine
the expression of Bcl-2, Bax and active caspase-3. The results
demonstrated that following treatment with DPL, the expression
level of the anti-apoptotic protein Bcl-2 was reduced, while the
expression levels of the pro-apoptotic proteins Bax and active
caspase-3 were increased significantly (Fig. 3B; P<0.05). The results confirmed
that DPL may promote the apoptosis of A549 cells.
DPL inhibits the PI3K signaling
pathway in NSCLC A549 cells
It has previously been demonstrated that the PI3K
signaling pathway is a crucial in a variety of tumors and its key
proteins, including AKT and mTOR, serve important roles in the
proliferation and migration of tumors (18,19).
Therefore, a series of specific key proteins were detected using
western blot analysis, to determine the effect of DPL on the PI3K
signaling pathway. As presented in Fig.
4, treatment of A549 cells with DPL led to a decrease in the
phosphorylation levels of AKT and mTOR, yet there were no
significant alterations in the total AKT and mTOR levels. The
activity and expression levels of PI3K downstream molecules,
including p-p70S6K and Cyclin D1, were reduced following treatment
with DPL. Therefore, DPL may have the ability to downregulate the
PI3K signaling pathway in A549 cells.
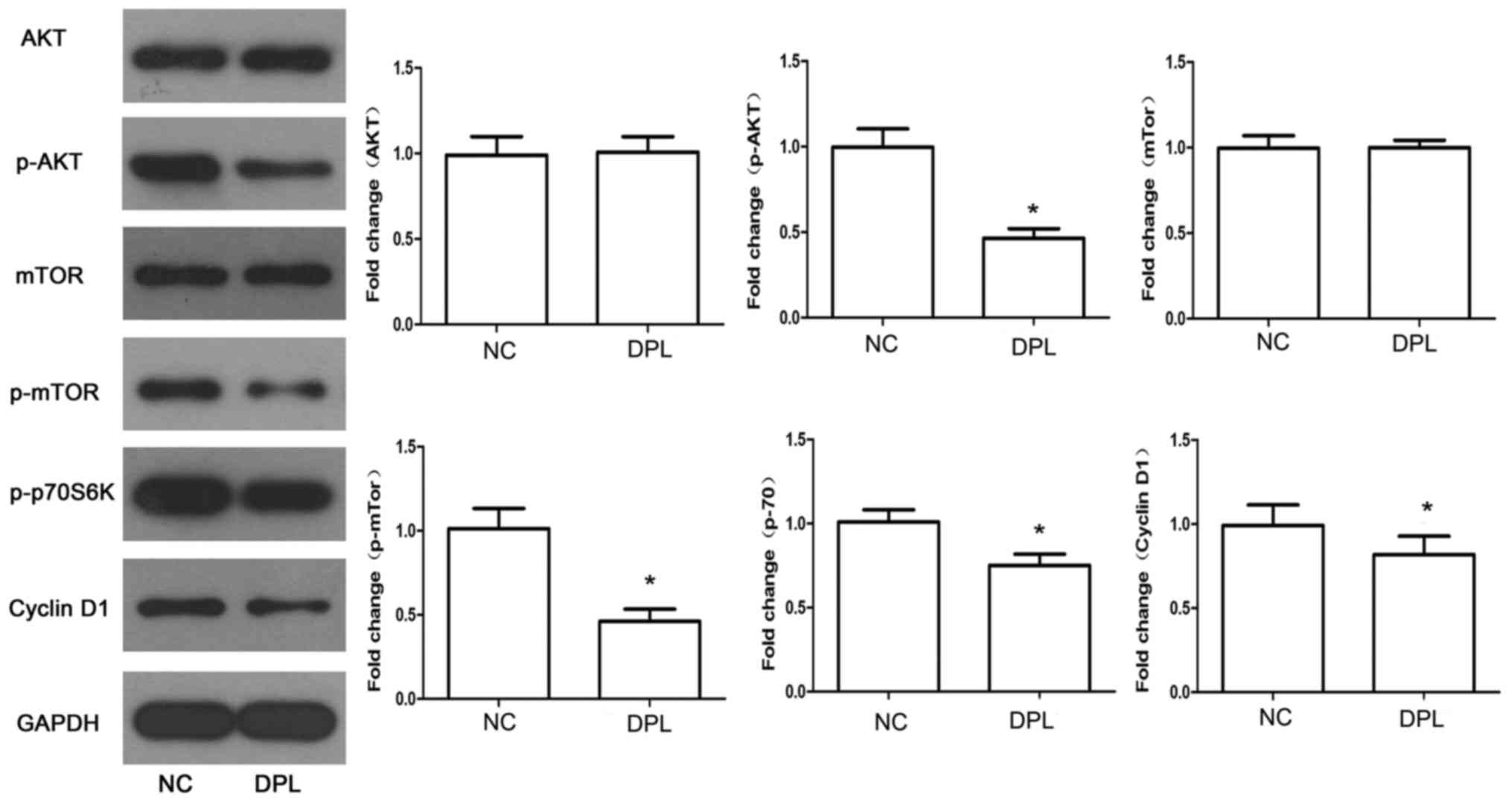 | Figure 4.DPL downregulates the PI3K signaling
pathway. The expression levels of key proteins associated with PI3K
signaling, including AKT, p-AKT, mTOR, p-mTOR, p-p70S6K and Cyclin
D1, were detected by western blot analysis. Data are presented as
the mean ± standard deviation. *P<0.05 vs. NC. DPL,
diprophylline; NC, negative control; p, phosphorylated; AKT, RAC-α
serine/threonine-protein kinase; mTOR, serine/threonine-protein
kinase mTOR; p70S6K, ribosomal protein S6 kinase β-1. |
Discussion
In the present study, the potential therapeutic
effect of DPL in NSCLC was studied. It was reported that DPL
significantly inhibited A549 proliferation, migration and invasion,
and induced apoptosis. In addition, it was revealed that the
anti-tumor effect of DPL may be regulated through inhibition of the
PI3K signaling pathway.
The ability of DPL to inhibit NSCLC A549 cell
proliferation and metastasis is due to numerous mechanisms.
Firstly, as a derivative of MX, DPL possesses a variety of
pharmacological effects (20,21). Additionally, DPL is a nonselective
phosphodiesterase (PDE) inhibitor. PDE inhibitors have been
reported to be used to treat tumor growth and migration, for
example in NIH3T3 fibroblasts (22)
and hepatocellular carcinoma cells (23). In the present study, it was
demonstrated that DPL had anti-proliferative and anti-migratory
effects in A549 cells. However, whether this effect is mediated via
the MX pathway or the pathway of PDE inhibition requires further
investigation. According to previous studies relating to DPL
utilization in the clinic, DPL (20 µM) exerts stimulatory effects
on the contractility of jejunal segments in low-contractile states,
and suppressive roles in high-contractile states (24). Additionally, according to the results
of the present study, 10 µM DPL was selected as the effective
concentration for the majority of the experiments.
Apoptosis inhibits cancer cell survival. In the
present study, it was reported that the expression levels of the
anti-apoptotic protein Bcl-2 were reduced, while the expression
levels of the pro-apoptotic proteins Bax and active caspase-3 were
increased significantly, in cells treated with DPL. The results
suggested that DPL may be involved in the regulation of apoptosis.
It has been reported in recent years that PDE inhibitor regulation
is one of a series of newly developed strategies in to explore
targeted therapies in different types of cancer (25). In addition, Domvri et al
(26) identified that the PDE
inhibitors roflumilast and theophylline enhance apoptosis, thereby
inhibiting tumor cell proliferation.
In the present study, the phosphorylation levels of
AKT and mTOR were decreased and the activity and expression levels
of the PI3K downstream molecules p-P70S6K and Cyclin D1 were
reduced following treatment with DPL. It is widely reported that
the deregulation of the PI3K signaling pathway is involved in the
development of lung tumors and is associated with high-grade tumors
and advanced disease (27). The
regulation of PI3K signaling has been proposed as a potential
therapeutic target in NSCLC. Mutations of numerous key factors in
this pathway are associated with NSCLC (28) and four of the tested factors in this
pathway, p-AKT, p-mTOR, p-p70S6K and Cyclin D1, may respond to the
overexpression and knockdown of DPL. It may be that DPL is an
inhibitor of the PI3K pathway, mediating cell invasion and
apoptosis by regulating a number of the key factors in the PI3K
pathway.
In summary, the present study elucidated that DPL
may inhibit A549 cell proliferation, migration and invasion, in
addition to promoting apoptosis. DPL may regulate the expression
levels of p-AKT, p-mTOR, p-p70S6K and Cyclin D1, and act as an
inhibitor of the PI3K pathway. Therefore, DPL may be a promising
novel drug for NSCLC treatment. However, there are limitations to
this preliminary study; for example, further investigations into
the underlying mechanisms of DPL may be beneficial, and the use of
animal models may also be advantageous. Thus, further studies are
required in the future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during the present
study are included in this published article.
Authors' contributions
XBR designed and performed the study and revised the
manuscript. HYZ performed experiments and wrote the manuscript. YHR
and YW performed experiments and analyzed the data. All authors
have read and agreed the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lang Y, Kong X, He C, Wang F, Liu B, Zhang
S, Ning J, Zhu K and Xu S: Musashi1 promotes non-small cell lung
carcinoma malignancy and chemoresistance via activating the Akt
signaling pathway. Cell Physiol Biochem. 44:455–466. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Molina JR, Yang P, Cassivi SD, Schild SE
and Adjei AA: Non-small cell lung cancer: Epidemiology, risk
factors, treatment, and survivorship. Mayo Clinic Proc. 83:584–594.
2008. View
Article : Google Scholar
|
|
3
|
Sakashita S, Sakashita M and Tsao Sound M:
Genes and pathology of non-small cell lung carcinoma. Semin Oncol.
41:28–39. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Wakelee H, Kelly K and Edelman MJ: 50
years of progress in the systemic therapy of non-small cell lung
cancer. Am Soc Clin Oncol Educ Book. 34:177–189. 2014. View Article : Google Scholar
|
|
5
|
Du L and Morgensztern D: Chemotherapy for
advanced-stage non-small cell lung cancer. Cancer J. 21:366–370.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hasegawa T, Sawa T, Futamura Y, Horiba A,
Ishiguro T, Marui T and Yoshida T: Feasibility of rebiopsy in
non-small cell lung cancer treated with epidermal growth factor
receptor-tyrosine kinase inhibitors. Intern Med. 54:1977–1980.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Franco R, Oñatibia-Astibia A and
Martínez-Pinilla E: Health benefits of methylxanthines in cacao and
chocolate. Nutrients. 5:4159–4173. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Boison D: Methylxanthines, seizures, and
excitotoxicity. Handb Exp Pharmacol. 251–266. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Zubairy YF, Patil VW, Benjamin T, Jangam
D, Bijle MN and Patil S: Effect of methylxanthines (coffee/tea
consumers) on oral precancer and oral cancer patients with smoking
and smokeless tobacco habits. J Contemp Dent Pract. 13:745–758.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Goel PN and Gude RP: Unravelling the
antimetastatic potential of pentoxifylline, a methylxanthine
derivative in human MDA-MB-231 breast cancer cells. Mol Cell
Biochem. 358:141–151. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lange A, Gustke H, Glassmeier G, Heine M,
Zangemeister-Wittke U, Schwarz JR, Schumacher U and Lange T:
Neuronal differentiation by indomethacin and IBMX inhibits
proliferation of small cell lung cancer cells in vitro. Lung
Cancer. 74:178–187. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Malki AM, Gentry J and Evans SC:
Differential effect of selected methylxanthine derivatives on
radiosensitization of lung carcinoma cells. Exp Oncol. 28:16–24.
2006.PubMed/NCBI
|
|
13
|
Edling CE, Selvaggi F, Ghonaim R, Maffucci
T and Falasca M: Caffeine and the analog CGS 15943 inhibit cancer
cell growth by targeting the phosphoinositide 3-kinase/Akt pathway.
Cancer Biol Ther. 15:524–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lentini A, Kleinman HK, Mattioli P,
Autuori-Pezzoli V, Nicolini L, Pietrini A, Abbruzzese A, Cardinali
M and Beninati S: Inhibition of melanoma pulmonary metastasis by
methylxanthines due to decreased invasion and proliferation.
Melanoma Res. 8:131–137. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Morais JA, Neves MC, Silva JR and Avila R:
Absorption of intra-bronchial diprophylline-methodology and
preliminary results. Eur J Drug Metab Pharmacokinet. 17:187–193.
1992. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Basnet RM, Zizioli D, Guarienti M, Finazzi
D and Memo M: Methylxanthines induce structural and functional
alterations of the cardiac system in zebrafish embryos. BMC
Pharmacol Toxicol. 18:722017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Levi-Schaffer F and Touitou E: Xanthines
inhibit 3T3 fibroblast proliferation. Skin Pharmacol. 4:286–290.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Polivka J Jr and Janku F: Molecular
targets for cancer therapy in the PI3K/AKT/mTOR pathway. Pharmacol
Ther. 142:164–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Martini M, De Santis MC, Braccini L,
Gulluni F and Hirsch E: PI3K/AKT signaling pathway and cancer: An
updated review. Ann Med. 46:372–383. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee IA, Kamba A, Low D and Mizoguchi E:
Novel methylxanthine derivative-mediated anti-inflammatory effects
in inflammatory bowel disease. World J Gastroenterol. 20:1127–1138.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Schoen K, Yu T, Stockmann C, Spigarelli MG
and Sherwin CM: Use of methylxanthine therapies for the treatment
and prevention of apnea of prematurity. Paediatr Drugs. 16:169–177.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stork PJ and Schmitt JM: Crosstalk between
cAMP and MAP kinase signaling in the regulation of cell
proliferation. Trends Cell Biol. 12:258–266. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ionta M, Rosa MC, Almeida RB, Freitas VM,
Rezende-Teixeira P and Machado-Santelli GM: Retinoic acid and cAMP
inhibit rat hepatocellular carcinoma cell proliferation and enhance
cell differentiation. Braz J Med Biol Res. 45:721–729. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu FF, Chen DP, Xiong YJ, Lv BC and Lin
Y: Characteristics of diprophylline-induced bidirectional
modulation on rat jejunal contractility. Korean J Physiol
Pharmacol. 18:47–53. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Koiri RK, Mehrotra A and Trigun SK:
Targetting cancer with Ru(III/II)-phosphodiesterase inhibitor
adducts: A novel approach in the treatment of cancer. Med
Hypotheses. 80:841–846. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Domvri K, Zarogoulidis K, Ziogas N,
Zarogoulidis P, Petanidis S, Porpodis K, Kioseoglou E and
Hohenforst-Schmidt W: Potential synergistic effect of
phosphodiesterase inhibitors with chemotherapy in lung cancer. J
Cancer. 8:3648–3656. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Scrima M, De Marco C, Fabiani F, Franco R,
Pirozzi G, Rocco G, Ravo M, Weisz A, Zoppoli P, Ceccarelli M, et
al: Signaling networks associated with AKT activation in non-small
cell lung cancer (NSCLC): New insights on the role of
phosphatydil-inositol-3 kinase. PLoS One. 7:e304272012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Ding L, Getz G, Wheeler DA, Mardis ER,
McLellan MD, Cibulskis K, Sougnez C, Greulich H, Muzny DM, Morgan
MB, et al: Somatic mutations affect key pathways in lung
adenocarcinoma. Nature. 455:1069–1075. 2008. View Article : Google Scholar : PubMed/NCBI
|