Introduction
Endometrial cancer is a common malignant cancer that
is diagnosed in premenopausal and postmenopausal females, which
occurs in the endometrial epithelium of the reproductive tract
(1). In the last five years, studies
have reported that endometrial cancer accounts for 20–30% of
malignant tumors of the female reproductive tract and that the
incidence of endometrial cancer is increasing (2,3). Although
a number of anti-cancer treatment strategies are available for
endometrial cancer, increasing the level of apoptosis in tumor
cells has become an important method for the clinical treatment of
endometrial cancer (4–6).
Sirtuin-7 is a member of the sirtuin family, which
is a family of evolutionarily conserved NAD-dependent deacetylases
(7). Studies have indicated that
sirtuin-7 serves an important role in carcinogenesis, and the
downregulation of sirtuin-7 may be a potential pharmacological
strategy to increase chemoradiation-induced apoptosis in cancer
cells in patients with cancer (8). A
previous study has demonstrated that sirtuin-7 maintains critical
features that define cancer cells by removing the acetylation mark
on lysine 18 of histone H3, which has been described as a general
marker of tumor prognosis and oncological treatments (9). However, the role of sirtuin-7 in
endometrial cancer has not been the subject of in-depth
investigations.
The inhibition of nuclear factor (NF)-κB activation
can lead to the inhibition of cell growth and invasion in various
types of cancer cells (10–12). A previous study indicated that the
anti-malarial drug dihydroartemisinin significantly suppressed the
metastasis of non-small-cell lung cancer via inhibiting NF-κB
activity (13). Additionally, NF-κB
has been shown to suppress apoptosis and promote the proliferation
of bladder cancer cells by upregulating survivin expression in
vitro and in vivo (14).
Furthermore, Yi et al (15)
reported that the NF-κB p53 apoptosis signaling pathway is
associated with poor prognosis in colorectal cancer via regulation
of cell proliferation. Furthermore, the results have concluded that
NF-κB expression is overexpressed in nonmalignant endometrial
tumors when compared with normal endometrial tissues (16). However, the associations between
sirtuin-7 and NF-κB have not been clearly investigated in
endometrial cancer cells. Therefore, in the present study, the role
of sirtuin-7 was investigated, and its possible apoptotic mechanism
in endometrial cancer was analyzed.
Materials and methods
Patients
A total of 5 female endometrial cancer patients were
recruited at the Harbin Second Hospital (Harbin, China) between
June 2014 and January 2017. The mean age of patients was 42.2 years
(range, 34.2–48.5 years). All patients provided written informed
consent. The Ethics Committee of Harbin Second Hospital approved
the present study.
Cell lines and culture
The endometrial cancer cell lines (KLE, RL95-2,
AN3CA and Ishikawa) were provided by Randy Kramer (University of
California, San Francisco, CA, USA). Normal primary endometrial
cells (ESCs) were derived from normal endometrial tissues that were
discarded from periodontal surgical procedures, and this was
approved by the Harbin Second Hospital. The 1–5th passages of ESC
cells were cultured in Dulbecco's modified Eagle medium (DMEM;
Thermo Fisher Scientific, Inc., Waltham, MA, USA) that was
supplemented with 15% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.). The 6th passages of ESC were authenticated by
Suzhou Microread Genetics Co., Ltd (Suzhou, China) and used for
analysis of protein expression. KLE, RL95-2, AN3CA and Ishikawa
cells were cultured in DMEM that was supplemented with 10% FBS at
37°C in a humidified incubator at 5% CO2.
Small interfering RNA (siRNA)
Ishikawa cells were harvested when 85–95% confluence
was attained. The cells were transfected the following siRNAs to
silence sirtuin-7 target gene using Lipofectamine® 2000
(Invitrogen, Thermo Fisher Scientific, Inc.) according to the
manufacturer's instructions. The sequences are as follows:
siRNA-vector, 5′-UUCUCCGAACGUGUCACGU-3′ and siRNA-sirtuin-7 siRNA,
5′-GCACCGTCCATCGTGTTTATT-3′. All siRNAs were purchased from
GeneChem Inc. (Shanghai, China). Further analysis was performed 72
h after transfection.
NF-κB1 overexpression
Ishikawa cells were harvested after reaching 85–95%
confluence. All the DMEM media (Thermo Fisher Scientific, Inc.) was
then removed. Ishikawa cells were transfected by pCMVp-NEO-NF-κB1
(100 pmol, Invitrogen; Thermo Fisher Scientific, Inc.) or pCMVp-NEO
vector (100 pmol, Invitrogen; Thermo Fisher Scientific, Inc.) using
Lipofectamine® 2000 according to the manufacturer's
instructions. Cells were used for further analysis 72 h after
transfection.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from tumor cells using
RNeasy Mini kit (Qiagen Sciences, Inc., Gaithersburg, MD, USA) in a
20 µl volume with 5 pmol of each primer and 1 µl of RNA, 0.5 µl
reverse transcriptase, 2 µl buffer, 2 µl dNTPs (both Takara
Biotechnology Co., Ltd., Dalian, China) and 18 µl deionized water
for 2 h at 37°C according to manufacturer's protocol. The
expressions levels of sirtuin-7 in cells were analyzed by RT-qPCR
with β-actin as an endogenous control (17). All forward and reverse primers were
synthesized by Invitrogen (Thermo Fisher Scientific, Inc.). The
primer sequences are as follows: Sirtuin-7 forward,
5′-AGAAGCGTTAGTGCTGCCG-3′ and reverse, 5′-GAGCCCGTCACAGTTCTGAG-3′;
β-actin forward, 5′-CGGAGTCAACGGATTTGGTC-3′ and reverse,
5′-AGCCTTCTCCATGGTCGTGA-3′. PCR amplification followed preliminary
denaturation at 94°C for 2 min, followed by 45 cycles of 95°C for
30 sec, annealing temperature reduced to 56°C for 30 sec and 72°C
for 10 min. The reaction was performed in a total volume of 20 µl
that contains 50 ng genomic DNA, 200 µM dNTP, 2.5 U Taq DNA
polymerase and 200 µM of each primer. The relative mRNA expression
changes were calculated by 2−ΔΔCq (18). The results are expressed as the n-fold
way compared with β-actin.
MTT cytotoxicity assays
Ishikawa cells were incubated with melittin (2.0
mg/ml) in 96-well plates for 24, 48 and 72 h in triplicate for each
condition at 37°C. For the control, PBS was added instead of
melittin. At each time point, 20 µl MTT (5 mg/ml) in PBS solution
was added to each well and the plate was then further incubated for
4 h at 37°C. The medium was removed, and 100 µl dimethyl sulfoxide
(DMSO) was added into the wells to solubilize the crystals. The
optical density was analyzed with a BIO-RAD (ELISA) reader (Bio-Rad
Laboratories, Inc.) at a wavelength of 490 nm.
Cell invasion and migration
assays
Ishikawa cells were treated with cisplatin (5.0
mg/ml) for 24 h and non-treated cells were used as control.
Migration and invasion assays of Ishikawa cells were conducted in a
6-well culture plate with chamber inserts (BD Biosciences Franklin
Lakes, NJ, USA). For migration assays, Ishikawa cells
(1×104/well) were placed into the upper chamber with the
non-coated membrane (Transwell inserts; BD Biosciences) and DMEM
supplemented with 5% PBS was plated in the lower chamber for 48 h
at 37°C. For invasion assays, the cells (1×104/well)
were placed into the upper chamber with the Matrigel inserts
membrane for 48 h at 37°C. For migration and invasion assays,
Ishikawa cells were counted in at least three randomly
stained-fields using a light microscope (Olympus Corporation,
Tokyo, Japan) at magnification, ×400.
Proliferation assays
Ishikawa cells (0.5×103/well) were seeded
in a six-well plate and cultured for 10 days at 37°C in a
humidified incubator at 5% CO2. The cell colonies were
fixed with 10% formaldehyde for 10 min at 37°C and stained for 5
min with 0.5% crystal violet for 10 min at 37°C. The number of cell
colonies was then calculated using ImageJ software 3.0 (National
Institutes of Health, Bethesda, MD, USA). The images were taken
under a light microscope (Olympus Corporation).
Apoptosis assays
Ishikawa cells (1×106/well) were seeded
in 6-well plates for 12 h at 37°C in a humidified incubator at 5%
CO2. The cells, siRNA-sirtuin-7 or siRNA-vector were
then incubated with cisplatin (Control, 5.0 mg/ml) or PBS (Mock)
for 24 h at 37°C. The cells were removed, collected and washed with
phosphate buffer solution (PBS) three times. Subsequently, Ishikawa
cells were incubated with fluorescein isothiocyanate
(FITC)-conjugated Annexin V and propidium iodide (PI) for 2 h at
room temperature, using an Annexin V-FITC Apoptosis Detection kit
(BD Biosciences) and according to the manufacturer's protocol. Flow
cytometry was used to analyze the percentage of apoptotic Ishikawa
cells by using FCS Express™ 4 IVD (De Novo Software,
Glendale, CA, USA).
Tumor cell adhesion
Ishikawa cells (1×106/well) were cultured
in six-well plate for 1.5 h at 37°C. Cells were stained with 1%
Methylene blue (Sigma-Aldrich, Merck KGaA, Darmstadt, Germany) for
30 min at room temperature. The number of Ishikawa cells that
adhered on the monolayer was recorded using an Olympus microscope
in at least three field of view (Olympus Corporation).
Western blot analysis
Western blotting was used to evaluate protein levels
according to protocols as previously described by Kurien et
al (19). Briefly, the cells
(1×107/well) were lysed in a lysis buffer that contains
1% phenylmethanesulfonyl fluoride (PMSF) for three times and
centrifuged at 8,000 × g for 10 min at 4°C. Protein concentration
was measured by a BCA protein assay kit (Thermo Fisher Scientific,
Inc.). The proteins (60 µg) were separated using 10% SDS-PAGE.
Then, the proteins were transferred onto nitrocellulose membranes.
The nitrocellulose membranes were blocked with 5% (w/v) non-fat
milk powder, which was dissolved in tris-buffered saline plus
Tween-20 (TBST) solution, for 2 h at 37°C. This was followed by
incubation with primary rabbit anti-human antibodies: Anti-SIRT7
(dilution, 1:1,000; cat. no. ab135055), Bcl-2 (dilution, 1:1,000;
cat. no. ab32124), Bcl-xl (dilution, 1:1,000; cat. no. ab32370),
Mcl-1 (cat. no. ab32087), caspase-3 (cat. no. ab2171), Bad (cat.
no. ab32445), Bax (cat. no. ab77566) and β-actin (dilution 1:500;
cat. no. ab8226; all Abcam, Shanghai, China) for 12 h at 4°C. All
antibodies were diluted at 1:1,000. The secondary goat anti-rabbit
antibodies (dilution 1:2,000, cat. no. PV-6001; OriGene
Technologies, Inc., Beijing, China) were incubated and the
membranes were washed with PBS buffer and visualized with ECL
Western blotting detection reagents (GE Healthcare, Chicago, IL,
USA). The density of the bands was analyzed by Quantity One
(version 4.62; Bio-Rad Laboratories, Inc.).
Immunohistochemistry
Endometrial tumors were fixed using 10% formaldehyde
followed by embedding in paraffin wax. The tissue blocks were cut
into serial sections with a thickness of 4 µm. The tumor sections
were incubated with rabbit anti-human anti-SIRT7 (dilution,
1:1,000; cat. no. ab135055; Abcam) for 12 h at 4°C. Then, the tumor
sections were treated with Goat Anti-Rabbit IgG (Alexa
Fluor® 488; dilution 1:1,000; cat. no. ab150077; Abcam)
for 12 h at 4°C. The sections were examined under a light
microscope at a magnification of ×400.
Statistical analysis
All data are expressed the mean ± standard
deviation, and the experiments were performed three times.
Statistical analyses were performed using one-way analysis of
variance followed by Tukey's multiple comparison post-hoc tests
using Graph Pad Prism software (version 5.0; GraphPad Software,
Inc., La Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
Sirtuin-7 is overexpressed in
endometrial cancer tissues
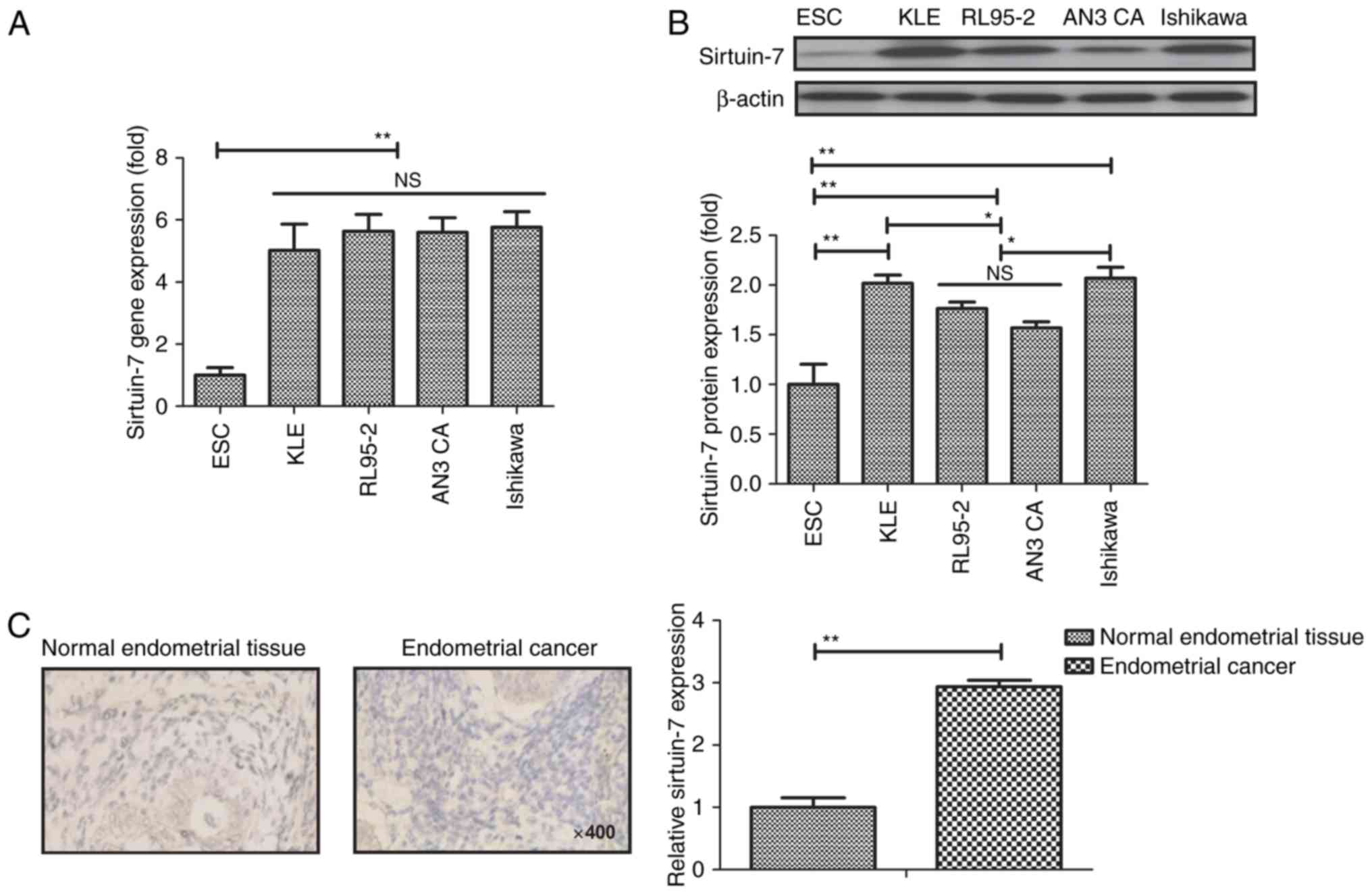
To determine the role of sirtuin-7 in endometrial
cancer, the mRNA and protein levels of all sirtuin-7 were examined
in endometrial cancer cell lines (KLE, RL95-2, AN3 CA and Ishikawa)
and compared with ESCs. Results revealed that sirtuin-7 gene was
higher in endometrial cancer cell lines compared with normal
primary endometrial cells (P<0.01; Fig. 1A). Western blot analysis also
demonstrated that sirtuin-7 protein expression was higher in
endometrial cancer cell lines compared with normal primary
endometrial cells (P<0.01; Fig.
1B). The expression of sirtuin-7 was significantly higher in
endometrial cancer tissues compared with normal tissues (P<0.01;
Fig. 1C). These results suggest that
sirtuin-7 may be a potential target for the treatment of
endometrial cancer.
Knockdown of sirtuin-7 inhibits the
growth, proliferation and invasiveness of endometrial cancer
cells
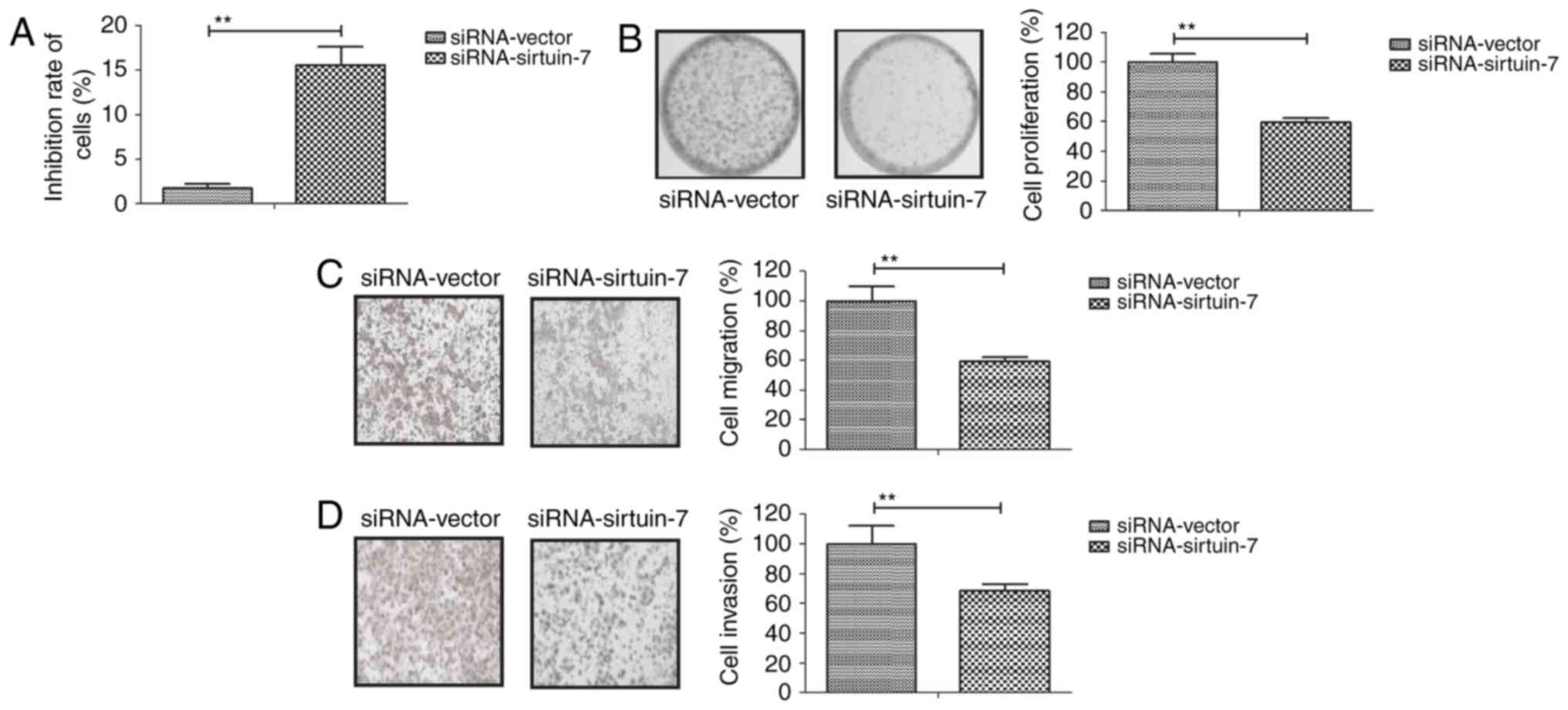
To gain further insights into the role of sirtuin-7
in endometrial cancer cells, sirtuin-7 expression was downregulated
in Ishikawa cells. As indicated in Fig.
2A, siRNA-induced knockdown of sirtuin-7 was able to
significantly inhibit the growth of Ishikawa cells compared with
the control. The downregulation of sirtuin-7 was also able to
inhibit the proliferation of Ishikawa cells (Fig. 2B). Furthermore, the knockdown of
sirtuin-7 was able to suppress the migration and invasion of
Ishikawa cells (Fig. 2C and D). These
results suggest that the downregulation of sirtuin-7 is able to
inhibit the growth and metastasis of endometrial cancer cells.
Knockdown of sirtuin-7 promotes
cisplatin-induced apoptosis of endometrial cancer cells
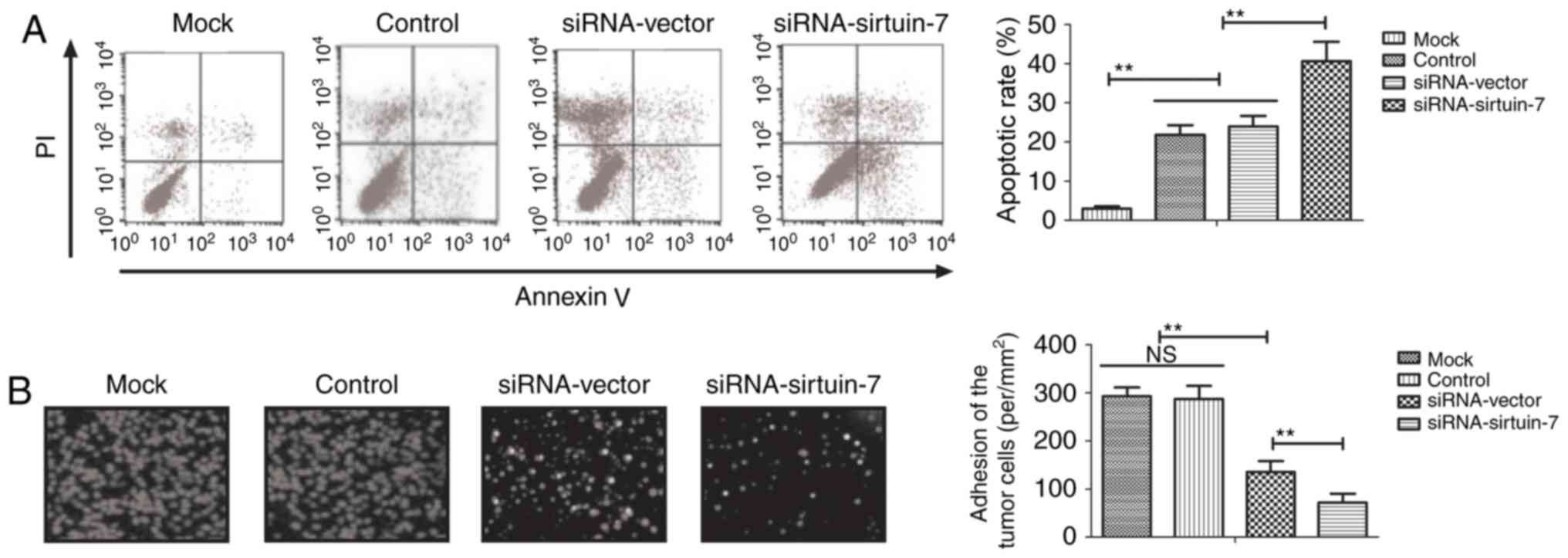
Cisplatin is the most commonly used chemotherapeutic
drug for the treatment of endometrial cancer (20). Therefore, the present study
investigated whether sirtuin-7 knockdown is able to enhance the
apoptotic sensitivity of endometrial cancer cells to cisplatin. It
was indicated that the knockdown of sirtuin-7 was able to
significantly promote cisplatin-induced apoptosis of Ishikawa cells
(Fig. 3A). The results indicated that
the downregulation of sirtuin-7 was able to increase
cisplatin-induced inhibition on the adhesion of Ishikawa cells
compared with untreated controls (Fig.
3B). These results suggest that sirtuin-7 may be involved in
resistance to apoptosis in endometrial cancer cells.
Sirtuin-7 knockdown regulates the
apoptosis of endometrial cancer cells via the NF-κB signaling
pathway
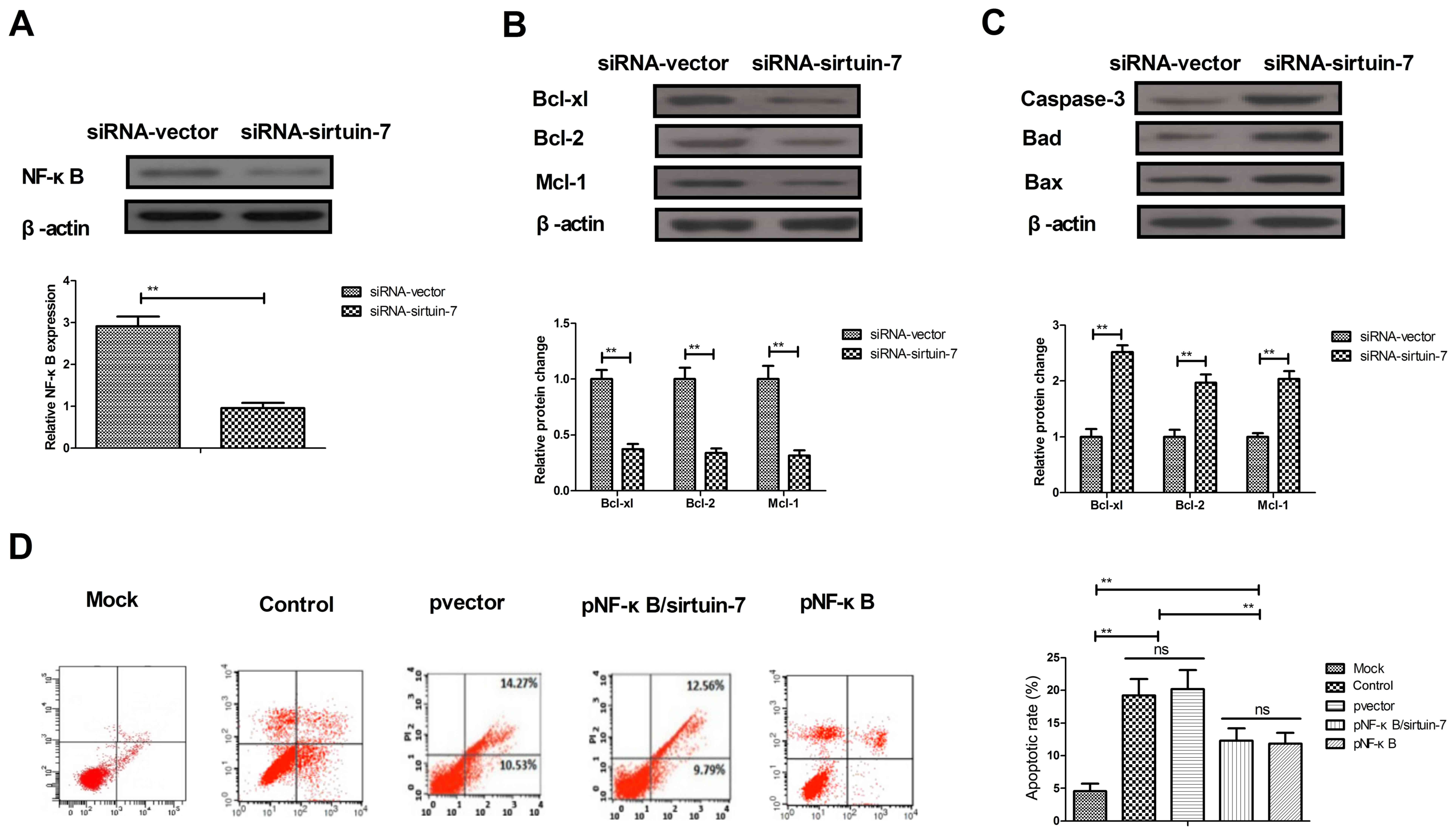
Due to the involvement of NF-κB in apoptosis of
endometrial cancer cells (19), the
potential mechanism of sirtuin-7 in apoptosis was further analyzed.
The downregulation of sirtuin-7 was able to significantly
downregulate NF-κB1 expression in Ishikawa cells (Fig. 4A). The knockdown of sirtuin-7 resulted
in the downregulation of NF-κB1 target proteins (Bcl-xl, Bcl-2, and
Mcl-1), which are anti-apoptotic. The knockdown of sirtuin-7 also
led to the upregulation of NF-κB1 target proteins (Bad and Bax),
which are pro-apoptotic (Fig. 4B and
C). Furthermore, the results indicated that the overexpression
of NF-κB1 inhibited sirtuin-7 knockdown and increased
(pNF-κB/sirtuin-7) apoptosis of endometrial cancer cells induced by
cisplatin (Fig. 4D). These results
suggest that the knockdown of sirtuin-7 regulates the apoptosis of
endometrial cancer cells via the NF-κB signaling pathway.
Discussion
Endometrial cancer is one of the most common female
reproductive tract tumors worldwide (21). Although surgery, chemotherapy,
radiotherapy and endocrine therapies are becoming increasingly
important for the clinical treatment of endometrial cancer, the
prognosis is poor for patients with recurrent or advanced stages of
disease (22). Studies have suggested
that sirtuin-7 is a potential therapeutic target for human cancer
(23,24). In the present study, the role of
sirtuin-7 in the progression of endometrial cancer and its
potential molecular mechanisms were investigated. The present study
reports that sirtuin-7 knockdown was able to significantly inhibit
the growth, proliferation, migration and invasion of endometrial
cancer cells. Additionally, sirtuin-7 knockdown was able to promote
the apoptosis of endometrial cancer cells that was induced by
cisplatin via downregulation of the NF-κB signaling pathway.
Although a previous study has suggested that
sirtuin-7 mRNA expression is upregulated in patients with breast
cancer (25), the expression levels
of sirtuin-7 in endometrial cancer cells had not yet been
investigated. In the present study, it was demonstrated that
sirtuin-7 mRNA and protein expression levels are upregulated in
endometrial cancer cell lines and cancer tissues compared with
normal endometrial cell lines and tissues. Wang et al
(8) indicated that sirtuin-7
exhibited oncogenic potential in human ovarian cancer cells, which
may serve a role as an oncogene in ovarian malignancies and may be
a potential therapeutic target. Additionally, a high expression of
sirtuin-7 has served as a predictor of adverse outcomes in patients
with breast cancer (26). In the
present study, it was demonstrated that the knockdown of sirtuin-7
was able to significantly inhibit the growth and metastasis of
endometrial cancer cells. In a study on pancreatic cancer, McGlynn
et al (27) indicated that
sirtuin-7 may be a potential novel biomarker for determining
patient outcome. The findings of the present study indicate that
sirtuin-7 may be a biomarker for patients with endometrial
cancer.
A number of studies have suggested proapoptotic and
prosurvival roles of sirtuin-7 in human cancer cells (28,29), with
reports indicating that sirtuin inhibitors, sirtinol and
nicotinamide, have been used to inhibit cell growth in several
types of cancer, including breast and lung cancer, and oral
squamous cell carcinoma (30). A
study has suggested that 5-FU induces radiosensitivity via
degradation of sirtuin-7 to favor the cell death pathway in
targeted cancer cells (31).
In the present study, it was demonstrated that
sirtuin-7 knockdown was able to enhance the apoptosis of
endometrial cancer cells, which was induced by cisplatin. Previous
studies have also suggested that the inhibition of NF-κB activation
suppressed the production of angiogenic factors, which are involved
in the apoptosis of endometrial cancer cells (32,33). The
results of the present study revealed that the knockdown of
sirtuin-7 was able to decrease NF-κB expression, and the apoptosis
of endometrial cancer cells was promoted. However, the
overexpression of NF-κB inhibited sirtuin-7 knockdown and abolished
sirtuin-7 knockdown and increased apoptosis induced by cisplatin.
Yang et al (34) reported that
NF-κB is able to regulate the expression of Bcl-2 and caspase-3 in
gastric cancer cells, which was induced by tumor necrosis
factor-related apoptosis inducing ligand. The present study
indicated that the knockdown of sirtuin-7 resulted in the
downregulation of the NF-κB-targeted anti-apoptotic proteins
(Bcl-xl, Bcl-2 and Mcl-1) and the upregulation of the
NF-κB-targeted pro-apoptotic proteins (caspase-3, Bad and Bax).
In conclusion, the findings of the present study
indicate a role for sirtuin-7 in the apoptosis of endometrial
cancer cells, and this may be mediated via the NF-κB signaling
pathway. The findings suggest that sirtuin-7 may be a potential
therapeutic target for endometrial cancer therapy, however further
study is required.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The analyzed data sets generated during the study
are available from the corresponding author upon reasonable
request.
Authors' contributions
SM conducted the experiments. JM analyzed the data
in the present study and HY designed the experiments.
Ethics approval and consent to
participate
All patients provided written informed consent. The
Ethics Committee of Harbin Second Hospital (Harbin, China) approved
the present study.
Consent for publication
All identifying patient information has been removed
and written, informed consent for the publication of the present
study was provided by all patients.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bourgin C, Saidani M, Poupon C, Cauchois
A, Foucher F, Leveque J and Lavoue V: Endometrial cancer in elderly
women: Which disease, which surgical management? A systematic
review of the literature. Eur J Surg Oncol. 42:166–175. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee B, Suh DH, Kim K, No JH and Kim YB:
Influence of positive peritoneal cytology on prognostic factors and
survival in early-stage endometrial cancer: A systematic review and
meta-analysis. Jpn J Clin Oncol. 46:711–717. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lheureux S and Oza AM: Endometrial
cancer-targeted therapies myth or reality? Review of current
targeted treatments. Eur J Cancer. 59:99–108. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Guo D, Li L, Zhang H and Zheng W:
Pathologic assessment and clinical impacts for endometrial cancer
and precursors after progestin treatment. Zhonghua Bing Li Xue Za
Zhi. 44:216–220. 2015.(In Chinese). PubMed/NCBI
|
|
5
|
Turkevich VG: Clinical evaluation of the
effectiveness of radiation treatment for endometrial cancer. Vopr
Onkol. 60:371–374. 2014.(In Russian). PubMed/NCBI
|
|
6
|
Karnjus-Begonja R, Vrdoljak E, Corusić A,
Haller H, Jelavic TB, Matković V, Strinić T, Barisić D, Tomic S,
Kukura V, et al: Clinical recommendations for diagnosing, treatment
and monitoring of patients with endometrial cancer-Croatian
Oncology Society and Croatian Society for Gynecology and Obstetrics
as Croatian Medical Association units and Croatian Society of
Gynecological Oncology. Lijec Vjesn. 135:230–234. 2013.(In
Croatian). PubMed/NCBI
|
|
7
|
Jung-Hynes B, Nihal M, Zhong W and Ahmad
N: Role of sirtuin histone deacetylase SIRT1 in prostate cancer. A
target for prostate cancer management via its inhibition? J Biol
Chem. 284:3823–3832. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wang HL, Lu RQ, Xie SH, Zheng H, Wen XM,
Gao X and Guo L: SIRT7 exhibits oncogenic potential in human
ovarian cancer cells. Asian Pac J Cancer Prev. 16:3573–3577. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Martinez-Redondo P, Santos-Barriopedro I
and Vaquero A: A big step for SIRT7, one giant leap for Sirtuins in
cancer. Cancer Cell. 21:719–721. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Huang X, Teng Y, Yang H and Ma J: Propofol
inhibits invasion and growth of ovarian cancer cells via regulating
miR-9/NF-κB signal. Braz J Med Biol Res. 49:e57172016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Ji BL, Xia LP, Zhou FX, Mao GZ and Xu LX:
Aconitine induces cell apoptosis in human pancreatic cancer via
NF-kappaB signaling pathway. Eur Rev Med Pharmacol Sci.
20:4955–4964. 2016.PubMed/NCBI
|
|
12
|
Chen H, Huang Y, Huang J, Lin L and Wei G:
Gigantol attenuates the proliferation of human liver cancer HepG2
cells through the PI3K/Akt/NF-kappaB signaling pathway. Oncol Rep.
37:865–870. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Jiang J, Geng G, Yu X, Liu H, Gao J, An H,
Cai C, Li N, Shen D, Wu X, et al: Repurposing the anti-malarial
drug dihydroartemisinin suppresses metastasis of non-small-cell
lung cancer via inhibiting NF-κB/GLUT1 axis. Oncotarget.
7:87271–87283. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cui X, Shen D, Kong C, Zhang Z, Zeng Y,
Lin X and Liu X: NF-kappaB suppresses apoptosis and promotes
bladder cancer cell proliferation by upregulating survivin
expression in vitro and in vivo. Sci Rep. 7:407232017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yi W, Xiao E, Ding R, Luo P and Yang Y:
High expression of fibronectin is associated with poor prognosis,
cell proliferation and malignancy via the NF-κB/p53-apoptosis
signaling pathway in colorectal cancer. Oncol Rep. 36:3145–3153.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Faloppa CC, Baiocchi G, Cunha IW, Fregnani
JH, Osorio CA, Fukazawa EM, Kumagai LY, Badiglian-Filho L, Pinto GL
and Soares FA: NF-κB and COX-2 expression in nonmalignant
endometrial lesions and cancer. Am J Clin Pathol. 141:196–203.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xiao S, Wang J and Xiao N: MicroRNAs as
noninvasive biomarkers in bladder cancer detection: a diagnostic
meta-analysis based on qRT-PCR data. Int J Biol Markers.
31:e276–e285. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kurien BT and Scofield RH: Validating
antibody specificities for immunohistochemistry by protein blotting
methods. Methods Mol Biol. 1554:61–73. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang X, Kyo S, Nakamura M, Mizumoto Y,
Maida Y, Bono Y, Takakura M and Fujiwara H: Imatinib sensitizes
endometrial cancer cells to cisplatin by targeting CD117-positive
growth-competent cells. Cancer Lett. 345:106–114. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
van der Steen-Banasik E, Christiaens M,
Shash E, Coens C, Casado A, Herrera FG and Ottevanger PB: European
Organisation for Research and Treatment of Cancer, Gynaecological
Cancer Group (EORTC-GCG): Systemic review: Radiation therapy alone
in medical non-operable endometrial carcinoma. Eur J Cancer.
65:172–181. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mouka V, Tsili AC, Messinis T,
Papoudou-Bai A, Kamina S and Argyropoulou MI: Solitary adrenal
metastasis from early-stage dedifferentiated endometrial carcinoma:
CT findings and review of the literature. J Obstet Gynaecol.
36:881–882. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsai YC, Greco TM and Cristea IM: Sirtuin
7 plays a role in ribosome biogenesis and protein synthesis. Mol
Cell Proteomics. 13:73–83. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hubbi ME, Hu H, Kshitiz Gilkes DM and
Semenza GL: Sirtuin-7 inhibits the activity of hypoxia-inducible
factors. J Biol Chem. 288:20768–20775. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Aljada A, Saleh AM, Alkathiri M, Shamsa
HB, Al-Bawab A and Nasr A: Altered Sirtuin 7 expression is
associated with early stage breast cancer. Breast Cancer (Auckl).
9:3–8. 2015.PubMed/NCBI
|
|
26
|
Geng Q, Peng H, Chen F, Luo R and Li R:
High expression of Sirt7 served as a predictor of adverse outcome
in breast cancer. Int J Clin Exp Pathol. 8:1938–1945.
2015.PubMed/NCBI
|
|
27
|
McGlynn LM, McCluney S, Jamieson NB,
Thomson J, MacDonald AI, Oien K, Dickson EJ, Carter CR, McKay CJ
and Shiels PG: SIRT3 & SIRT7: Potential novel biomarkers for
determining outcome in pancreatic cancer patients. PLoS one.
10:e01313442015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Paredes S, Villanova L and Chua KF:
Molecular pathways: Emerging roles of mammalian Sirtuin SIRT7 in
cancer. Clin Cancer Res. 20:1741–1746. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Han Y, Liu Y, Zhang H, Wang T, Diao R,
Jiang Z, Gui Y and Cai Z: Hsa-miR-125b suppresses bladder cancer
development by down-regulating oncogene SIRT7 and oncogenic long
non-coding RNA MALAT1. FEBS Lett. 587:3875–3882. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Alhazzazi TY, Kamarajan P, Joo N, Huang
JY, Verdin E, D'Silva NJ and Kapila YL: Sirtuin-3 (SIRT3), a novel
potential therapeutic target for oral cancer. Cancer.
117:1670–1678. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Tang M, Lu X, Zhang C, Du C, Cao L, Hou T,
Li Z, Tu B, Cao Z, Li Y, et al: Downregulation of SIRT7 by
5-fluorouracil induces radiosensitivity in human colorectal cancer.
Theranostics. 7:1346–1359. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang J, Song H, Lu Y, Chen H, Jiang S and
Li L: Effects of estradiol on VEGF and bFGF by Akt in endometrial
cancer cells are mediated through the NF-kappaB pathway. Oncology
reports. 36:705–714. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Davies S, Dai D, Feldman I, Pickett G and
Leslie KK: Identification of a novel mechanism of NF-κB
inactivation by progesterone through progesterone receptors in
Hec50co poorly differentiated endometrial cancer cells: Induction
of A20 and ABIN-2. Oncol Rep. 94:463–470. 2004.
|
|
34
|
Yang LQ, Fang DC, Wang RQ and Yang SM:
Effect of NF-kappaB, survivin, Bcl-2 and Caspase3 on apoptosis of
gastric cancer cells induced by tumor necrosis factor related
apoptosis inducing ligand. World J Gastroenterol. 10:22–25. 2004.
View Article : Google Scholar : PubMed/NCBI
|