Introduction
The increase in early diagnosis rate, in addition
with emerging treatments and targeting medicine, have decreased the
mortality rate of patients with breast cancer in a number of
developed countries (1,2). However, the incidence of breast cancer
is increasing in the majority of countries, including China
(3). The primary causes of mortality
in breast cancer are recurrence and metastasis (4), whereas angiogenesis serves an important
function in the process of breast cancer disease occurrence,
recurrence and metastasis (5,6).
Deleted in liver cancer 2 (DLC2), also named
StAR-related lipid transfer domain 13, was successfully cloned by
Ching et al in 2003 (7). As an
adhesion molecule, DLC2 lies on the adhesion plaque of the cell
membrane and is involved in cell adhesion (7), mitosis (8)
and sensory nerve conduct (9) under
physiological conditions. DLC2 is poorly expressed in a variety of
tumors, including liver cancer, lung cancer, colon cancer and
rectal cancer (10), and is
considered to be a potential tumor suppressor gene (7,10). In a
previous study, a DLC2-knockout mouse model was established and it
was indicated that DLC2-knockout mice exhibited more marked
angiogenesis ability compared with that of wild-type mice (11). In addition, it was indicated that DLC2
was downregulated in ~50% of cases of breast cancer at the mRNA
level (data not shown). This suggests that, as an
adhesion-associated molecule, DLC2 may be involved in tumor
formation and metastasis in breast cancer.
In the present study, the expression of DLC2 in
breast cancer was investigated, and its association among
clinicopathological parameters and the survival of patients was
examined. The effect of DLC2 on proliferation, migration and
invasion of breast cancer cells was additionally observed, in an
attempt to determine the function of DLC2 in the carcinogenesis and
development of breast cancer.
Materials and methods
Patients and tissue specimens
A total of 131 paraffin-embedded surgical specimens
of breast cancer were collected at The First Affiliated Hospital of
Sun Yat-sen University (Guangzhou, China) between January 2007 and
December 2007 for immunohistochemical assay. All specimens were
from primary tumors. All patients were female and the median age of
the patients was 43 years (range, 33–79 years). The ethics
committee at The First Affiliated Hospital of Sun Yat-sen
University (Guangzhou, China) approved the present study. The
clinicopathological data are presented in Table I. All patients were diagnosed with
primary breast cancer, and none of the patients received any form
of medical treatment prior to surgery. All patients provided
written informed consent prior to participation in the present
study.
 | Table I.Association between DLC2 and
clinicopathological parameters. |
Table I.
Association between DLC2 and
clinicopathological parameters.
|
|
| DLC2, no. of
patients |
|---|
|
|
|
|
|---|
| Clinicopathological
parameter | No. of patients
(%) | − | + | P-value |
|---|
| Median age,
years |
|
|
| 0.762 |
|
≤43 | 78 (59.5) | 43 | 44 |
|
|
>43 | 53 (40.5) |
|
|
|
| Tumor size, cm |
|
|
| 0.417 |
| T1
(≤2) | 66 (50.4) | 33 | 33 |
|
| T2
(2–5) | 42 (32.1) | 27 | 15 |
|
| T3
(>5) | 23 (17.5) | 15 | 8 |
|
| Lymphatic
metastasis |
|
|
|
<0.001a |
| N0 | 82 (62.6) | 30 | 52 |
|
|
N1-N3 | 49 (37.4) | 45 | 4 |
|
| Tumor
differentiationb |
|
|
|
<0.001a |
| I | 32 (24.4) | 6 | 26 |
|
| II | 34 (26.0) | 14 | 20 |
|
|
III | 65 (49.6) | 55 | 10 |
|
| Ki67 |
|
|
| 0.085 |
|
<14 | 52 (39.7) | 25 | 27 |
|
|
≥14 | 79 (60.3) | 50 | 29 |
|
| ER |
|
|
|
<0.001a |
| +
(≥1) | 35 (26.7) | 9 | 30 |
|
| −
(<1) | 96 (73.3) | 66 | 26 |
|
| PR |
|
|
| 0.622 |
| +
(≥1) | 110 (84.0) | 64 | 46 |
|
| −
(<1) | 21 (16.0) | 11 | 10 |
|
|
Her2/CerbB2c |
|
|
| 0.411 |
| −,
+ | 91 (69.5) | 49 | 42 |
|
| ++ | 14 (10.7) | 10 | 4 |
|
|
+++ | 26 (19.8) | 16 | 10 |
|
| Histological
type |
|
|
| 0.546 |
|
Invasive ductal carcinoma | 113 (86.3) | 62 | 51 |
|
|
Intraductal carcinoma | 9 (6.9) | 4 | 5 |
|
|
Invasive lobular
carcinoma | 2 (1.5) | 2 | 0 |
|
| Special
carcinoma | 7 (5.3) | 7 | 0 |
|
Construction of tissue microarrays
(TMAs)
A tissue array device (Unitma Quick-Ray; Utigma Co.,
Ltd., Seoul, South Korea) was used to construct the TMAs. For each
case, two 1-mm-diameter cylinders selected from two different areas
were included in the TMAs. Finally, 4 TMA blocks were constructed,
each containing 120 cylinders. Consecutive 4-µm-thick sections were
cut using a microtome and air-dried overnight at room
temperature.
Immunohistochemistry
The tissue samples were fixed win 10%
phosphate-buffered formalin for 6–48 h at room temperature.
Sections (4-µm-thick) were cut from the TMA blocks. All sections
were deparaffinized with xylene and rehydrated by a graded ethanol
series to distilled water. The sections were then heated for
antigenic retrieval in sodium citrate buffer (pH 6.0; WeijiangGene
Chem Co., Ltd., Guangzhou, China) for 3 min at 100°C. Endogenous
peroxidase was subsequently blocked with 0.3%
H2O2 for 15 min at room temperature and
incubated with 5% normal goat serum (Shanghai GeneChem Co., Ltd.,
Shanghai, China) in PBS for 30 min at room temperature. The slides
were incubated with rabbit monoclonal anti-DLC2 antibody (catalog
no. ab126489; 1:150; Abcam, Cambridge, MA, USA) overnight at 4°C.
Following three washes with PBS, the slides were incubated with
anti-rabbit secondary antibody (catalog no. 3900s; 1:200; Cell
Signaling Technology Inc., Danvers, MA, USA) for 1 h at room
temperature. Subsequently, the Non-Biotin HRP Detection system
(Shanghai GeneChem Co., Ltd., Shanghai, China) was used, according
to the manufacturer's protocol. Diaminobenzidine (WeijiangGene Chem
Co., Ltd.) was used for color reaction, and the antibody was
replaced by normal goat serum for negative controls. Slides were
reviewed and scored using a light microscope (magnification, ×400)
by two independent pathologists who were blinded to the data and
affiliated to the Department of Pathology, The First Affiliated
Hospital, Sun Yat-Sen University (Guangzhou, China). Scores given
by the two pathologists were averaged for further comparative
evaluation of DLC2 expression. The slides were counterstained with
haematoxylin at room temperature for 1 min. Immunoreactivity was
scored according to staining intensity of the product as follows:
0, no staining; 1, weak staining (light yellow); 2, moderate
staining (yellow brown) and 3, strong staining (brown). The
percentage of stained cells was classified as follows: 0, no
staining; 1, 1–25% staining; 2, 26–75% staining and 3, >75%
staining. Final scores >4 were defined as positive and scores ≤4
as negative (12).
Cell culture
The human breast cancer cell lines MDA-MB-231 and
MDA-MB-468 were obtained from the American Type Culture Collection
(Manassas, VA, USA). MDA-MB-231 cells were cultured in RPMI-1640
medium supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA) and MDA-MB-468 cells
were cultured in Dulbecco's modified Eagle's medium (DMEM)
supplemented with 10% FBS. All cells were cultured at 37°C with 5%
CO2 for 24 h. All cells were used within 2 months
following receipt or resuscitation of frozen aliquots.
Cell transfection with vectors
Cells were incubated in a humid incubator at 37°C
with 5% CO2 for 24 h, Stable DLC2-knockdown MDA-MB-468 and
MDA-MB-231 cell lines were constructed. A total of 5 µg/ml GV115
plasmid (catalog no. pGCSIL-004; Shanghai GeneChem Co., Ltd.) was
used. Transfection into cells at 30–50% confluency was performed
with Lipofectamine 2000 (Invitrogen; Thermo Fisher Scientific),
according to the manufacturer's protocol. Stable short hairpin
(sh)DLC2 or sh green fluorescent protein (GFP) cells were generated
by infection with shRNA lenti viruses against DLC2 or GFP (Shanghai
GeneChem Co., Ltd.), followed by puromycin (2.5 µg/ml) selection.
The expression level of DLC2 in stable shDLC2 or shGFP breast
cancer cells was verified by western blot analysis following 48
h.
Cell cycle analysis
Cell cycle content was determined by flow cytometry.
Both cell lines were collected at 104 cells/well and
fixed in 70% cold ethanol for 24 h at room temperature. Fixed cells
were subsequently washed three times with PBS followed by staining
with 50 µg/ml propidium iodide (catalog no. P1304MP; Thermo Fisher
Scientific, Inc.) for 30 min protected from light at room
temperature. Stained cells were analyzed with an Accuri C6 Plus
flow cytometer (version 3.0; BD Biosciences, Franklin Lakes, NJ,
USA) and data were obtained based on the analysis of 12,000
cells.
Plate clone formation assay
In total, 200 cells/well of each cell line were
seeded in 6-well plates (Corning Incorporated, Corning, NY, USA).
Cells were cultured in DMEM supplemented with 10% FBS for 14 days
at 37°C PBS was flushed twice over 4% paraformaldehyde-fixed cells
after 15 min at room temperature, followed by 30 min of 4% Giemsa
staining at room temperature. The number of clones formed was
determined using a light microscope (magnification, 400×) and ten
fields of view were randomly selected. ImageJ software (v1.8.0;
National Institutes of Health, Bethesda, MD, USA) was used for
quantification.
Soft agar assay
Both cell lines (5,000/well) were suspended in
medium containing 15% FBS and 0.5% agarose and seeded on a layer of
solidified agarose in each 60×15 mm cell-culture dish (Corning
Incorporated). Following 28 days, colony-forming units with cell
numbers ≥15 were counted and colony-forming efficiency was
calculated as the number of colonies/the number of seeded cells. To
view ten randomly selected fields of view a light microscope
(magnification, ×400) was used. ImageJ software (v1.8.0, National
Institutes of Health) was used to analyze data.
Wound healing migration assay
Cells were seeded on 24-well plates and cultured to
100% confluence. The confluent monolayer was wounded with a yellow
pipette tip and was allowed to migrate for 12 h for MDA-MB-468
cells and 20 h for MDA-MB-231 cells. Images were captured at 0 and
12 h in the same position for MDA-MB-468 cells, and at 0 and 20 h
in the same position for MDA-MB-231 cells. Migration distances were
measured, and speed was determined in µm/h using Image J software
(v1.8.0, National Institutes of Health).
Invasion assay
Matrigel (BD Biosciences) was dissolved at 4°C
overnight. Cells were starved in serum-free medium (Gibco; Thermo
Fisher Scientific, Inc.) 24 h prior to assay. Cells were harvested,
centrifuged at 252 × g for 5 min at 4°C and resuspended three times
with culture medium containing 1% FBS at room temperature, and
subsequently brought to a concentration of 106 cells/ml.
Following rehydration, 250 µl medium was removed from Transwell
inserts and 250 µl cell suspensions were added. Inserts were placed
in a 24-well plate, and 500 µl complete RPMI 1640 medium (Gibco;
Thermo Fisher Scientic, Inc.), with 15% FBS, containing 5 µg/ml
fibronectin (R&D Systems, Inc., Minneapolis, MN, USA), as an
adhesive substrate was added to the lower wells. Complete RPMI 1640
medium (500 µl) with 10% serum was added to the upper wells. Plates
were incubated for 24 h at 37°C in a 5% CO2 incubator.
The Transwells (Corning Incorporated) were removed from the 24-well
plates. Cells were stained with crystal violet (0.1%) for 30 min at
room temperature and washed twice in PBS. Non-invasive cells were
scraped off on the top of the Transwell with a cotton swab and the
number of invading cells was counted under a light microscope
(magnification, ×400) in ten randomly selected fields of view.
Western blot analysis
Protein from both cell lines was dissolved in SDS
lysis buffer (Shanghai GeneChem Co., Ltd.), and protein
concentration was determined using a bicinchoninic acid protein
assay kit (Shennengbocai Inc., Shanghai, China), according to the
manufacturer's protocols. A total of 20 µg of each protein sample
was separated by SDS-PAGE (8% gel) and transferred onto
polyvinylidene difluoride membranes overnight at 30 V. The
membranes were subsequently blocked with 5% non-fat dry milk in PBS
containing 0.1% Tween-20 for 1 h at room temperature and incubated
with the following primary antibodies: Anti-DLC2 (ab126489;
dilution, 1:2,000; Abcam), anti-Ras homolog family member A (RhoA;
catalog no. ab54835; dilution, 1:2,000; Abcam), anti-Rac family
small GTPase 1 (Rac-1; catalog no. ab33186; dilution, 1:2,000;
Abcam), anti-cell division cycle 42 (Cdc42; catalog no. ab64533;
dilution, 1:4,000; Abcam), anti-Rho-associated protein kinase
(Rock)-1 (catalog no. ab45171; dilution, 1:8,000; Abcam),
anti-Rock-2 (catalog no. MABN1037; dilution, 1:8,000; EMD
Millipore, Billerica, MA, USA), anti-GAPDH (catalog no. 51332;
dilution, 1:10,000; Cell Signaling Technology Inc.) overnight at
4°C. Following incubation with the primary antibody, the membranes
were washed three times with Tris-buffered saline containing
Tween-20 and incubated with anti-rabbit secondary antibody (catalog
no. 5127; dilution, 1:10,000; Cell Signaling Technology, Inc.) for
2 h at room temperature. The signal was detected by
electrochemiluminescence (EMD Millipore). Images were captured on
X-ray film and protein expression was quantified using ImageJ
software (v1.8.0, National Institutes of Health).
Statistical analysis
All data are presented as the mean ± standard
deviation from three independent experiments. P-values were
calculated by an unpaired Student's t-test or χ2 test
using SPSS for Windows software (version 16.0; SPSS, Inc., Chicago,
IL, USA). The Kaplan-Meier method was used to analyze disease-free
survival time of patients and comparisons were analyzed by log-rank
test. P<0.05 was considered to indicate a statistically
significant difference.
Results
DLC2 is downregulated in breast cancer
and is associated with tumor differentiation and lymph node
metastasis
The expression of DLC2 protein was detected by
immunohistochemistry in TMAs containing 131 cases of breast cancer
tissue and adjacent normal tissue. The clinicopathological data
were collected and analyzed. The results indicated that the DLC2
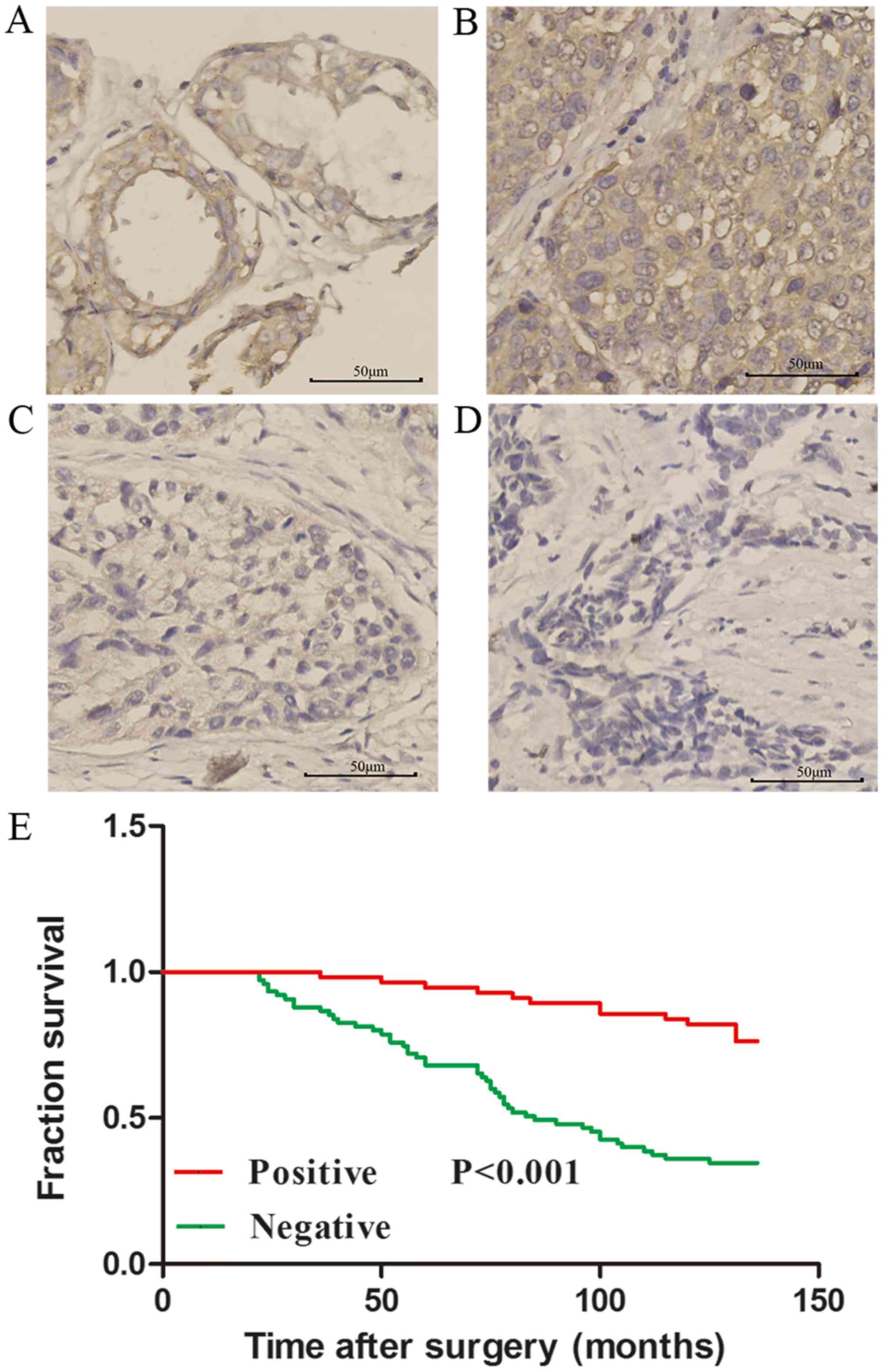
expression level in breast cancer tissue was lower compared with
the adjacent normal tissue (P<0.05; Table I; Fig.
1). The association between DLC2 expression and
clinicopathogical parameters was analyzed and it was identified
that lower expression of DLC2 in breast cancer tissue was
significantly associated with tumor differentiation (P<0.001)
and lymph node metastasis (P<0.001; Table I). The positive rate of DLC2
expression was 81.34% (26/32) in well differentiated grade I
(13) breast cancer and was markedly
higher (P>0.05) compared with poorly differentiated grade III
breast cancer at 15.38% (10/65) (Table
I). Cancer grade was determined using a cancer grading system
as previously described (13). The
positive rate of DLC2 expression in breast cancer with lymphatic
metastasis was 8.26% (4/49) compared with 63.41% (52/82) in breast
cancer without lymphatic metastasis (Table I). The results also indicated that the
DLC2 expression was associated with the expression of estrogen
receptor (P<0.001), which is important for the prognosis and
treatment of breast cancer. However, there was no statistical
significant difference between the expression of DLC2 and other
clinical features, including age, histological type, tumor size and
the expression of progesterone receptor, Ki-67 and CerbB2 (Table I).
Expression of DLC2 in breast cancer is
associated with patient overall survival (OS) rate
Following a median follow-up of 85 months (range,
22–136 months), 61 (53.4%) patients experienced disease recurrence
(Fig. 1E). Low expression of DLC2 was
significantly associated with a poorer OS rate (P<0.001).
Patients with negative DLC2 expression had a 10-year OS rate of
34.7% [95% confidence interval (CI), 17.2–51.0], whereas those with
positive DLC2 expression had a 10-year OS rate of 78.6% (95% CI,
62.65–93.9; P<0.001; Fig. 1E).
These results indicated that DLC2 expression may be an independent
prognostic marker for OS rate in patients with breast cancer.
Knockdown of DLC2 promotes
proliferation, migration and invasion in breast cancer cells
The aforementioned results of DLC2 expression in
breast cancer tissue strongly indicate that DLC2 may serve as a
tumor suppressor in the development of breast cancer. To analyze
the function of DLC2 in breast cancer cells, DLC2-silencing
(shDLC2) and control (shGFP) stable cell lines in MDA-MB-468 and
MDA-MB-231 were generated. Flow cytometry was performed to analyze
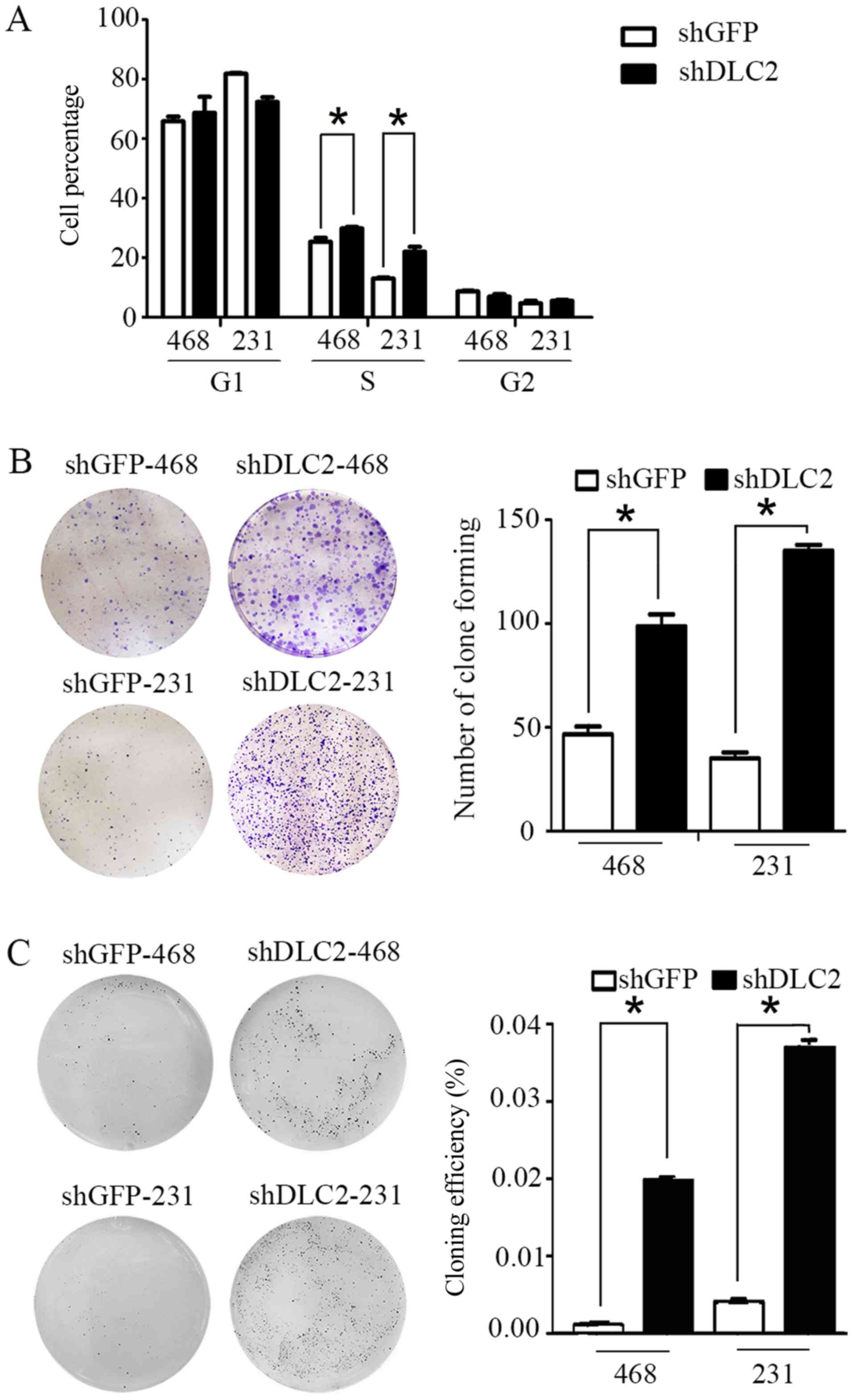
the cell cycle status of shDLC2 and shGFP stable cell lines.
Results indicated that the number of cells entering S phase
significantly increased for the shDLC2 group compared with the
shGFP group, when DLC2 was knocked down in MDA-MB-468 and
MDA-MB-231 cells (P<0.05; Fig.
2A). In MDA-MB-468, cell plate cloning experiments indicated
that the shDLC2 stable cell line exhibited significantly increased
cell clone formation ability compared with the shGFP cell line
(P<0.05; Fig. 2B). In addition,
soft agar cloning experiments indicated that the colony forming
efficiency was 1.98% in shDLC2 stable cell line, compared with
0.66% in the shGFP group (P<0.05; Fig.
2C). Similar results were observed in MDA-MB-231 cells
(Fig. 2B and C).
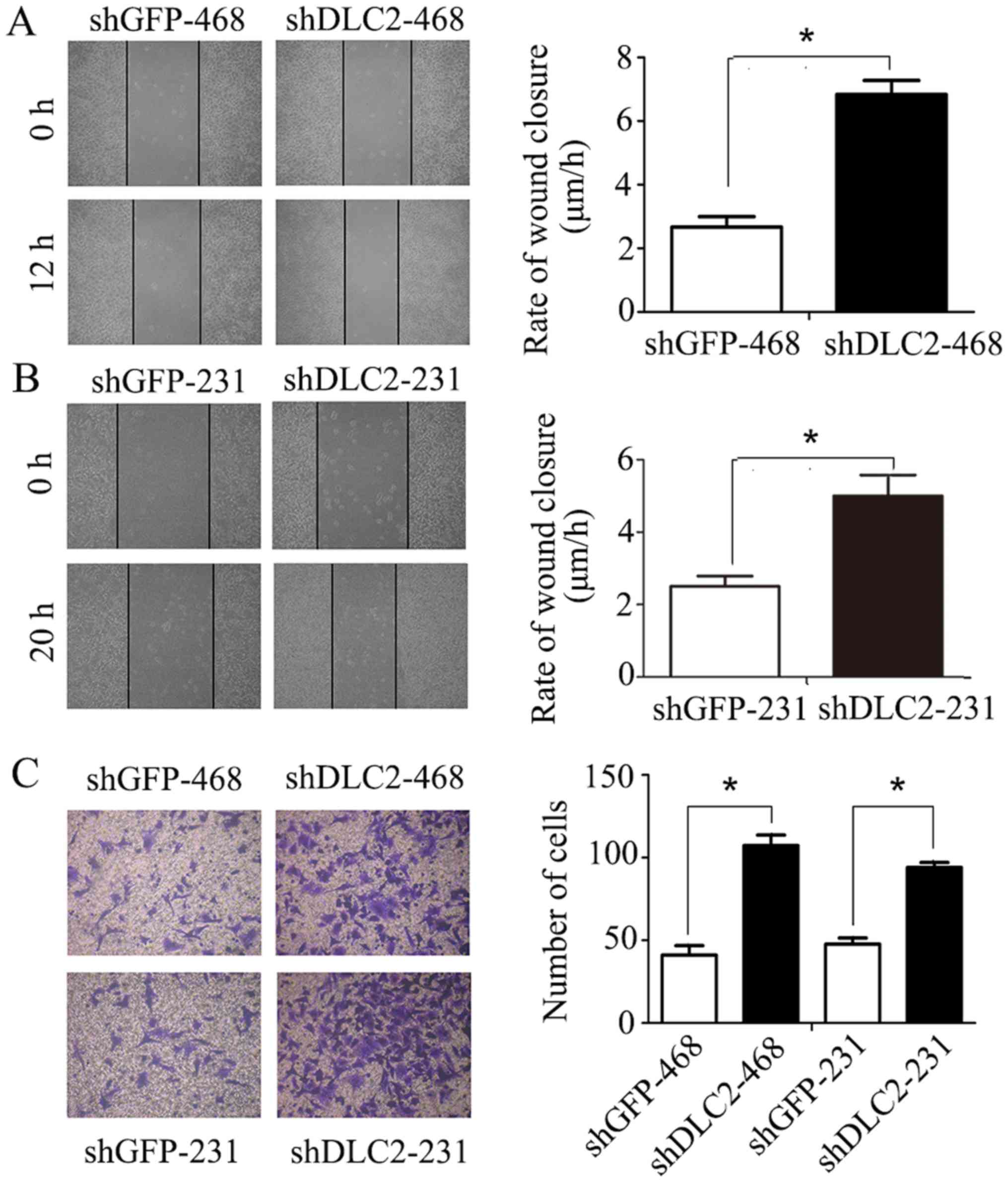
In wound healing migration assay, silencing of DLC2
in MDA-MB-468 and MDA-MB-231 cells significantly promoted cell
invasion at 12 and 20 h, respectively, following scratching
(P<0.05; Fig. 3A and B). In
addition, the Transwell invasion assay indicated that
downregulation of DLC2 in breast cancer cells significantly
increased migration, particularly in MDA-MB-468 cells (P<0.05;
Fig. 3C).
DLC2 may regulate breast cancer cell
proliferation and invasion via the RhoGTPase pathway
As DLC2 is a RhoGTPase activation protein-containing
molecule, the present study investigated whether the observed
results in DLC2-knockdown breast cancer cell lines were dependent
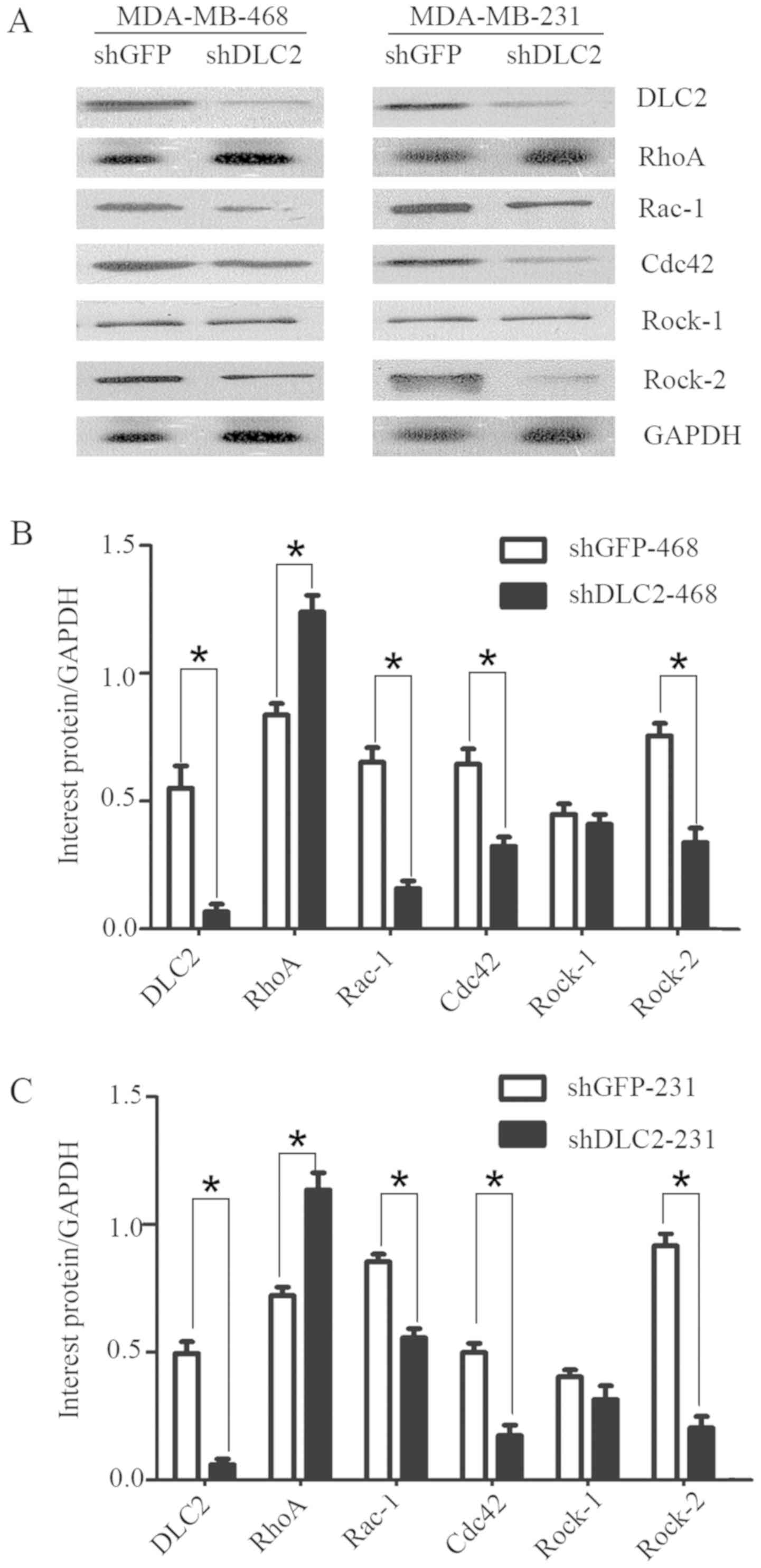
on the RhoGTPase pathway. Western blot analysis indicated that the
expression of RhoA was enhanced in the breast cancer cell lines
MDA-MB-468 and MDA-MB-231, following knockdown of DLC2 (Fig. 4A). In addition, Rac-1, Cdc42, and
Rock-2 were downregulated (Fig. 4A).
However, the expression of Rock-1 exhibited no obvious change
(Fig. 4A). Densitometry analysis
indicated that DLC2 may regulate these cellular processes through
the RhoGTPase pathway, including regulation of Rac-1, Cdc42, and
Rock-2 (Fig. 4B and C).
Discussion
DLC2 has been identified as a tumor suppressor gene
and is significantly downregulated in a wide range of human tumors
(7,10), including liver cancer, lung cancer,
colon cancer and rectal cancer (8).
In breast cancer, it has been reported that significant DLC2
downregulation was exhibited in 73% of tumors by reverse
transcription-quantitative polymerase chain reaction (10). In the present study, DLC2 expression
was detected in 131 cases of breast cancer by immunohistochemistry.
The present study, to the best of our knowledge, has used the
largest sample to investigate the protein expression of DLC2 in
breast cancer. The results suggest that downregulation of DLC2 is
associated with tumor differentiation, and is overall in agreement
with previous reports (10,14). In addition, the results of the present
study indicate that the downregulation of DLC2 is significantly
associated with poor prognosis in patients with breast cancer. This
indicates DLC2 may be a prognostic marker for patients with breast
cancer. In addition, the results of the present study indicated
that lower expression of DLC2 in breast cancer may promote
proliferation and viability. Similar observations were reported
previously in various tumor types. El-Sitt et al (15) reported that upregulation of DLC2
expression resulted in a limited number of astrocytoma cells in S
and G2 phase. The studies by Leung et al
(16,17) reported that silencing of DLC2 in liver
cancer cells led to an increased proliferation rate. In addition,
studies have demonstrated that DLC1 and DLC3, DLC family members
together with DLC2, are also involved in the regulation of
proliferation (15,18–22). These
results indicate that DLC2 serves as a tumor suppressor in
tumorigenesis.
Furthermore, the results of the present study
indicated that DLC2 affected tumorigenesis, and influenced
metastasis in breast cancer. It was indicated that decreased
expression of DLC2 was associated with breast cancer metastasis.
However, the results of the present study, in addition to those of
previous studies (14,23), indicated that downregulation of DLC2
increased the migratory and invasive ability of breast cancer
cells. We hypothesized that this was associated with DLC2 function
in focal adhesion, which is required for cell movement, and that
the deletion of DLC2 may cause this change in cell movement. This
may suggest that DLC2 participates in tumor metastasis. In our
previous study, it was identified that DLC2 was involved in
angiogenesis (11). Angiogenesis is
important for tumor metastasis (6),
therefore providing additional evidence that DLC2 may be involved
in tumor angiogenesis and regulate tumor metastasis in breast
cancer. DLC1, a DCL family member, was reported to regulate tumor
angiogenesis in prostate cancer, through the hypoxia-inducible
factor 1-vascular endothelial growth factor pathway (24). However, the underlying molecular
mechanism of DLC2 remains unknown.
DLC2 belongs to the RhoGTPase activation protein
family, and the members of this family primarily regulate
RhoGTPase, including RhoA, Rac1, Cdc42 and Rock expression
(25). In our previous research, it
was indicated that DLC2 regulated angiogenesis depending on RhoA
(9). Alternative studies have also
demonstrated that DLC2 serves an important function through the
RhoA pathway. It has been reported that, during pancreatic
development, Rho signaling was restricted by DLC2 (26). In liver cancer, DLC2 has been reported
to suppress tumor cell proliferation and colony formation in a
RhoGAP-dependent manner (17). In
astrocytoma, the regulation of RhoA in focal adhesion was essential
in cell motility (27). In addition,
in breast cancer, it was indicated that miR-125b affected the
metastatic activities of cells. The regulation of α-smooth muscle
actin by miR-125b has been reported to be dependent on the
DLC2-RhoA-Rock signaling pathway (28). In addition, Kaplan-Meier survival
analysis performed in a previous study that the mRNA expression of
DLC2 and its competing endogenous (ce)RNAs, including cadherin-5
(CDH5), homeobox D1 (HOXD1) and homeoboxD10 (HOXD10), is associated
with survival rate of patients with breast cancer. DLC2 3′
untranslated region (UTR) suppressed metastasis via inhibiting
epithelial-mesenchymal transition (EMT) in breast cancer (29). It was also indicated that the C-C
motif chemokine receptor 2 (CCR2) 3′ UTR inhibited cell metastasis,
by repressing EMT. The CCR2 3′ UTR has also been reported to serve
as a metastatic suppressor by acting as a ceRNA for DLC2 and
therefore, inhibiting the RhoA-Rock-1-MLC-filamentous actin pathway
in breast cancer cells (30). In
addition, the DLC2 3′ UTR has been demonstrated to promote
apoptosis and, therefore, DLC2 may serve as a ceRNA for Bcl-2
modifying factor (BMF) to promote apoptosis (31). The present study indicated that
silencing of DLC2 in breast cancer resulted in changes of Rac-1,
Cdc42 and Rock-2, therefore suggesting that not only RhoA, but also
other RhoGTPases, may be involved in breast cancer. The specific
underlying mechanisms and pathways involved require further
investigation.
In conclusion, low expression of DLC2 in breast
cancer tissue is associated with tumor differentiation, lymph node
metastasis and poor prognosis. Loss of DLC2 promotes proliferation,
migration and invasion of breast cancer cells, depending on the
RhoGTPase pathway. These results indicate that DLC2 is a tumor
suppressor in breast cancer. Future in vivo studies to
confirm the results of the present in vitro experiments are
scheduled to be performed.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81102021), the
Natural Science Foundation of Guangdong Province (grant no.
2014A030313032) and the Specialized Research Fund for the Doctoral
Program of Higher Education (grant no. 20110171120076).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LX and YL designed this study. ZY performed the
experiments and was a major contributor towards the writing of the
manuscript. HC analyzed and interpreted the data. MS performed the
immunohistochemistry experiments. YZ was responsible for the
collection and analysis of clinical data. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
The First Affiliated Hospital of Sun Yat-sen
University (Guangzhou, China) Ethics Committee approved the current
study. All methods were performed in accordance with the relevant
guidelines and regulations. All patients provided written informed
consent prior to participation in the present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
DLC2
|
deleted in liver cancer 2
|
|
TMA
|
tissue microarray
|
|
FBS
|
fetal bovine serum
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
Rock
|
Rho-associated protein kinase
|
|
OS
|
overall survival
|
References
|
1
|
Gray JM, Rasanayagam S, Engel C and Rizzo
J: State of the evidence 2017: An update on the connection between
breast cancer and the environment. Environ Health. 16:942017.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Law AMK, Lim E, Ormandy CJ and
Gallego-Ortega D: The innate and adaptive infiltrating immune
systems as targets for breast cancer immunotherapy. Endocr Relat
Cancer. 24:X12017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li Y, Li S, Meng X, Gan RY, Zhang JJ and
Li HB: Dietary natural products for prevention and treatment of
breast cancer. Nutrients. 9(pii): E7282017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Boix-Perales H, Borregaard J, Jensen KB,
Ersbøll J, Galluzzo S, Giuliani R, Ciceroni C, Melchiorri D,
Salmonson T, Bergh J, et al: The European medicines agency review
of pertuzumab for the treatment of adult patients with
HER2-positive metastatic or locally recurrent unresectable breast
cancer: Summary of the scientific assessment of the committee for
medicinal products for human use. Oncologist. 19:766–773. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Folkman J: Angiogenesis: An organizing
principle for drug discovery. Nat Rev Drug Discov. 6:273–286. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Carmeliet P: Angiogenesis in health and
disease. Nat Med. 9:653–660. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ching YP, Wong CM, Chan SF, Leung TH, Ng
DC, Jin DY and Ng IO: Deleted in liver cancer (DLC) 2 encodes a
RhoGAP protein with growth suppressor function and is
underexpressed in hepatocellular carcinoma. J Biol Chem.
278:10824–10830. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vitiello E, Ferreira JG, Maiato H, Balda
MS and Matter K: The tumour suppressor DLC2 ensures mitotic
fidelity by coordinating spindle positioning and cell-cell
adhesion. Nat Commun. 5:58262014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chan FK, Chung SS, Ng IO and Chung SK: The
RhoA GTPase-activating protein DLC2 modulates RhoA activity and
hyperalgesia to noxious thermal and inflammatory stimuli.
Neurosignals. 20:112–126. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ullmannova V and Popescu NC: Expression
profile of the tumor suppressor genes DLC-1 and DLC-2 in solid
tumors. Int J Oncol. 29:1127–1132. 2006.PubMed/NCBI
|
|
11
|
Lin Y, Chen NT, Shih YP, Liao YC, Xue L
and Lo SH: DLC2 modulates angiogenic responses in vascular
endothelial cells by regulating cell attachment and migration.
Oncogene. 29:3010–3016. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yakirevich E, Magi-Galluzzi C, Grada Z, Lu
S, Resnick MB and Mangray S: Cadherin 17 is a sensitive and
specific marker for metanephric adenoma. Am J Surg Pathol.
39:479–486. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lakhani SR, Ellis IO, Schnitt SJ, Tan PH
and van de Vijver MJ: WHO classification of tumours of the breast.
Fourth edition. Int Agency Res Cancer. 9–27. 2016.
|
|
14
|
Hanna S, Khalil B, Nasrallah A, Saykali
BA, Sobh R, Nasser S and El-Sibai M: StarD13 is a tumor suppressor
in breast cancer that regulates cell motility and invasion. Int J
Oncol. 44:1499–1511. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
El-Sitt S, Khalil BD, Hanna S, El-Sabban
M, Fakhreddine N and El-Sibai M: DLC2/StarD13 plays a role of a
tumor suppressor in astrocytoma. Oncol Rep. 28:511–518. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Leung TH, Yam JW, Chan LK, Ching YP and Ng
IO: Deleted in liver cancer 2 suppresses cell growth via the
regulation of the Raf-1-ERK1/2-p70S6K signalling pathway. Liver
Int. 30:1315–1323. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Leung TH, Ching YP, Yam JW, Wong CM, Yau
TO, Jin DY and Ng IO: Deleted in liver cancer 2 (DLC2) suppresses
cell transformation by means of inhibition of RhoA activity. Proc
Natl Acad Sci USA. 102:15207–15212. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yuan BZ, Zhou X, Durkin ME, Zimonjic DB,
Gumundsdottir K, Eyfjord JE, Thorgeirsson SS and Popescu NC: DLC-1
gene inhibits human breast cancer cell growth and in vivo
tumorigenicity. Oncogene. 22:445–450. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yuan BZ, Jefferson AM, Baldwin KT,
Thorgeirsson SS, Popescu NC and Reynolds SH: DLC-1 operates as a
tumor suppressor gene in human non-small cell lung carcinomas.
Oncogene. 23:1405–1411. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhou X, Thorgeirsson SS and Popescu NC:
Restoration of DLC-1 gene expression induces apoptosis and inhibits
both cell growth and tumorigenicity in human hepatocellular
carcinoma cells. Oncogene. 23:1308–1313. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lukasik D, Wilczek E, Wasiutynski A and
Gornicka B: Deleted in liver cancer protein family in human
malignancies (Review). Oncol Lett. 2:763–768. 2011.PubMed/NCBI
|
|
22
|
Kawai K, Kiyota M, Seike J, Deki Y and
Yagisawa H: START-GAP3/DLC3 is a GAP for RhoA and Cdc42 and is
localized in focal adhesions regulating cell morphology. Biochem
Biophys Res Commun. 364:783–789. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng L, Li X, Chou J, Xiang C, Guo Q,
Zhang Z, Guo X, Gao L, Xing Y and Xi T: StarD13 3′-untranslated
region functions as a ceRNA for TP53INP1 in prohibiting migration
and invasion of breast cancer cells by regulating miR-125b
activity. Eur J Cell Biol. 97:23–31. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shih YP, Liao YC, Lin Y and Lo SH: DLC1
negatively regulates angiogenesis in a paracrine fashion. Cancer
Res. 70:8270–8275. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tcherkezian J and Lamarche-Vane N: Current
knowledge of the large RhoGAP family of proteins. Biol Cell.
99:67–86. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Petzold KM, Naumann H and Spagnoli FM: Rho
signalling restriction by the RhoGAP Stard13 integrates growth and
morphogenesis in the pancreas. Development. 140:126–135. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Khalil BD, Hanna S, Saykali BA, El-Sitt S,
Nasrallah A, Marston D, El-Sabban M, Hahn KM, Symons M and El-Sibai
M: The regulation of RhoA at focal adhesions by StarD13 is
important for astrocytoma cell motility. Exp Cell Res. 321:109–122.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Tang F, Zhang R, He Y, Zou M, Guo L and Xi
T: MicroRNA-125b induces metastasis by targeting STARD13 in MCF-7
and MDA-MB-231 breast cancer cells. PLoS One. 7:e354352012.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li X, Zheng L, Zhang F, Hu J, Chou J, Liu
Y, Xing Y and Xi T: STARD13-correlated ceRNA network inhibits EMT
and metastasis of breast cancer. Oncotarget. 7:23197–23211.
2016.PubMed/NCBI
|
|
30
|
Hu J, Li X, Guo X, Guo Q, Xiang C, Zhang
Z, Xing Y, Xi T and Zheng L: The CCR2 3′UTR functions as a
competing endogenous RNA to inhibit breast cancer metastasis. J
Cell Sci. 130:3399–3413. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Guo X, Xiang C, Zhang Z, Zhang F, Xi T and
Zheng L: Displacement of Bax by BMF Mediates STARD13 3′UTR-induced
breast cancer cells apoptosis in an miRNA-depedent manner. Mol
Pharm. 15:63–71. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wolff AC, Hammond ME, Hicks DG, Dowsett M,
McShane LM, Allison KH, Allred DC, Bartlett JM, Bilous M,
Fitzgibbons P, et al: Recommendations for human epidermal growth
factor receptor 2 testing in breast cancer: American Society of
Clinical Oncology/College of American Pathologists clinical
practice guideline update. J Clin Oncol. 31:3997–4013. 2013.
View Article : Google Scholar : PubMed/NCBI
|