Introduction
Radiotherapy is well known to be used for the
treatment of about half of cancer patients (1–3). However,
radioresistant cancer cells and the tumor recurrence after
radiotherapy have been recognized as serious impediments to the
long-term survival of cancer patients (4,5).
Therefore, to overcome these problems, the biological effects of
ionizing radiation on human tumors have been flourishingly and
diversely investigated up to date. Furthermore, these problems are
closely concerned with tumor microenvironments comprised of
fibroblasts, a variety of inflammatory cells, perivascular cells
and endothelial cells, as well as hypoxic conditions (3). In particular, tumor microenvironments
have been not only shown to positively regulate the tumor
malignancy, but also to greatly influence the radiosensitivity of
cancer cells (3,6). In addition, under tumor
microenvironment, the epithelial-mesenchymal transition (EMT) has
been well shown to play a critical role in the tumor progression
via triggering the motile and invasive activities of cancer cells
to infiltrate into lymph or blood vascular systems (6,7). During
the process of the EMT, the protein levels of epithelial markers
are decreased, but that of mesenchymal markers are increased
(8,9).
The alternations in these marker proteins lead to the loss of
cell-cell interaction, thereby allowing extravasation of cancer
cells from the primary tumors (6,8,9). Thus, EMT can be easily found in the
invasive regions of aggressive cancers (10,11).
Furthermore, many reports indicate that EMT is regarded as a key
mechanism to lead to the generation of cancer stem-like cells and
the resistance of anti-cancer drugs (12–14).
Importantly, endothelial cells have been suggested
to play a pivotal role in the tumor microenvironment as a major
component of the tumor vascular system (15,16).
Furthermore, these cells under tumor microenvironment have been
known to affect cancer stem-like cells through the networks of
growth factors and cytokines (15),
thereby directly regulating the self-renewal of them (17,18).
However, the effects of irradiated endothelial cells on tumor
malignancy and cancer stem-like cells have not been fully
clarified.
In this study, we investigated the differences in
malignant behavior of liver cancer cells in response to irradiated
endothelial cells. We found that 2 Gy irradiation of endothelial
cells enhances EMT of liver cancer cells, and increases the
self-renewal of cancer stem-like cells, whereas both 6 Gy- and
fractionated irradiation (2 Gy × 3 days) do not greatly affect
these events. Our observation also revealed that endothelial cells
play a key role in modulating the malignancy of liver cancer cells
in response to irradiation.
Materials and methods
Cell culture
The human liver cancer HepG2, Hep3B and Huh7 cells
were obtained from the American Type Culture Collection (Manassas,
VA, USA). These liver cancer cell lines were maintained in
Dulbecco's modified Eagle's medium (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) supplemented with 10% (v/v) bovine calf serum,
penicillin (50 U/ml), and streptomycin (50 µg/ml) (all from Gibco;
Thermo Fisher Scientific, Inc., Waltham, MA, USA). Human umbilical
vein endothelial cells (HUVECs) were purchased from ScienCell
Research Laboratories, Inc. (Carlsbad, CA, USA). HUVECs plated on
gelatin-coated 60-mm dishes were cultured in complete endothelial
cell culture medium (ECM; ScienCell Research Laboratories, Inc.)
supplemented with 5% fetal bovine serum, 1% antibiotics and 1%
endothelial cell growth supplement in a humidified 5%
CO2 incubator at 37°C. These cells from passages 2 to 5
were used for experiments.
Irradiation
HUVECs were exposed to γ-rays with a
137Cs irradiation source (Eckert & Ziegler, Berlin,
Germany) at a dose rate of 2.6 Gy/min.
Preparation of conditioned medium
After the medium for endothelial cells was freshly
replaced with new medium, HUVECs were irradiated with various doses
of γ-rays, and then cultured for 24 h. Shortly afterward, the
conditioned medium was harvested, filtered with 0.45-µm filter or
centrifuged for 5 min at 700 × g to remove cells and debris and
then transferred to tubes.
Cell growth curve
The HepG2 cells were plated onto 60-mm dishes at a
density of 2×105 cells/dish, and then treated with the
medium conditioned by HUVECs. At 24, 48 and 72 h after the
treatment, the cells were trypsinized, washed with PBS, and then
the number of HepG2 cells not stained with trypan blue were counted
with a hemocytometer.
Clonogenic cell survival assay
Appropriate numbers of HepG2 cells were plated onto
60-mm dishes, incubated overnight, treated with the medium
conditioned by HUVECs irradiated with various doses of γ-ray and
then cultured for 14 days in a 5% CO2 incubator at 37°C.
The colonies were fixed with 95% methanol and stained with 0.5%
crystal violet. The colonies containing more than 60 cells were
counted. The average of triplicate dishes was calculated for each
sample. The results were normalized according to the plating
efficiencies of the corresponding HepG2 cells treated with the
medium conditioned by non-irradiated HUVECs, and then the surviving
fractions were calculated.
Western blot analysis
Mouse monoclonal antibodies against Vimentin (cat.
no. sc-6260; dilution 1:1,000; incubation time, 1 h at room
temperature), E-cadherin (cat. no. sc-8426; dilution 1:1,000;
incubation time, 1 h at room temperature) and Zeb1 (cat. no.
sc-81428; dilution 1:1,000; incubation time, 1 h at room
temperature) were purchased from Santa Cruz Biotechnology, Inc.
(Santa Cruz, CA, USA). A mouse monoclonal antibody against
N-cadherin (cat. no. 610920; dilution 1:1,000; incubation time, 1 h
at room temperature) was purchased from BD Bioscience (San Jose,
CA, USA). Rabbit monoclonal antibodies against Slug (cat. no. 9585;
dilution 1:1,000; incubation time, 1 h at room temperature) and
Snail (cat. no. 3879; dilution 1:1,000; incubation time, 1 h at
room temperature) were purchased from Cell Signaling Technology,
Inc. (Denver, MA, USA). Mouse monoclonal anti-β-actin (cat. no.
A5441; dilution 1:1,000; incubation time, 1 h at room temperature),
horseradish peroxidase-conjugated anti-mouse IgG (cat. no. A9044;
dilution 1:10,000; incubation time, 1 h at room temperature) and
horseradish peroxidase-conjugated anti-rabbit IgG (cat. no. A0545;
dilution 1:10,000; incubation time, 1 h at room temperature)
antibodies were purchased from Sigma-Aldrich; Merck KGaA. Cells
were treated with lysis buffer [40 mM Tris-HCl (pH 8.0), 120 mM
NaCl, and 0.1% (v/v) NP40] supplemented with protease inhibitor
cocktail tablets (1 tablet/50 ml; Boehringer, Mannheim, Germany),
and centrifuged for 15 min at 12,000 × g at 4°C. 30 µg of proteins
were separated by SDS-PAGE and transferred to nitrocellulose
membranes (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
membranes were blocked with 5% (w/v) nonfat dry milk in
Tris-buffered saline and then incubated for 1 h with primary
antibodies at the dilution of 1:1,000, at room temperature.
Specific reaction bands were detected using peroxidase-conjugated
secondary antibodies which were at the dilution of 1:10,000, and
proteins were visualized using an enhanced chemiluminescence system
(Amersham Biosciences, Piscataway, NJ, USA).
cDNA preparation and RT2
Profiler PCR Array
Total RNA was extracted from the cells using the
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.), and 1
µg of the isolated total RNAs was reverse transcribed to
complementary DNA (cDNA) with the SuperScript First-Strand
Synthesis System (Gibco; Thermo Fisher Scientific, Inc.), according
to the manufacturer's instructions. The reaction was performed at
25°C for 5 min and then at 42°C for 30 min, and was terminated at
85°C for 5 min. Next, the cDNA was diluted to 100 µl, and then was
used as a template for RT2 Profiler PCR Array Human
cancer stem cells (cat. no. 330231 PAHS-176ZA; Qiagen, Valencia,
CA, USA). The information of transcripts is shown on the
instruction manual of RT2 Profiler PCR Array supplied by
the manufacturer. The transcripts were analyzed using a
CFX96™ Real-Time PCR Detection System (Bio-Rad,
Hercules, CA, USA). PCR amplification was carried out using a
thermal profile of beginning at 95°C for 10 min, followed by 40
cycles at 95°C for 15 sec, and last extension at 60°C for 1 min.
Analysis for the PCR array was performed with a web-based software
(http://dataanalysis.sabiosciences.com/pcr/arrayanalysis.php)
supplied by the manufacturer. The levels of gene expression were
normalized to GAPDH, a housekeeping gene, and shown as the fold
change compared with control.
Migration and invasion assays
Both migration and invasion assays were performed
using the Transwell chamber (8-µm pore size; BD Biosciences). In
total, 2×104 cells were resuspended in serum-free growth
medium for these assays. For the invasion assay, the interior of
the inserts was precoated with 10 mg/ml growth factor-reduced
Matrigel (BD Biosciences). For both assays, the cells were added to
the interior of the inserts. Growth medium supplemented with 10%
(v/v) fetal bovine serum was added to the lower chamber. After
incubation for 24 h, the cells attached on the upper surface of the
filter were removed with a cotton swab. The cells on the lower
surface of the filter were fixed and stained. The number of cells
was determined by counting cells in five microscopic fields per
well. In addition, the cells were imaged by phase contrast
microscopy (Nikon Eclipse 80i; Nikon, Tokyo, Japan).
Sphere forming assay
Cells were grown in serum-free DMEM/F12 (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with B27 (Gibco;
Thermo Fisher Scientific, Inc.), N2 (Gibco; Thermo Fisher
Scientific, Inc.), 20 ng/ml basic fibroblast (Peprotech, London,
UK) and 20 ng/ml epidermal growth factor (Peprotech) onto 24-well
ultra-low attachment plates at 300 cells per well for 7 or 14 days,
and then the size and number of spheres were determined using a
phase-contrast Nikon microscope (TS100; Nikon). To measure the size
of sphere, 12 spheres per group were randomly selected.
Statistical analyses
All data presented as the mean ± standard error of
the mean and are representative of at least three independent
experimental repeats. Statistical analysis were performed with SPSS
ver.18.0 (SPSS, Inc., Chicago, IL, USA). Differences between groups
were analyzed using an unpaired Student's t-test and one-way
analysis of variance (ANOVA) or two-way ANOVA, followed by
Dunnett's post-hoc test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Endothelial cells enhance the
malignancy of liver cancer cells
Endothelial cells under tumor microenvironments have
been known to play a key role in the survival, growth and
malignancy of cancer cells, as well as participation in the
formation of tumor blood vessels (15,16).
Therefore, we investigated whether endothelial cells affect the
malignancy of tumor cells. To examine these effects in
vitro, we used the endothelial cell conditioned medium (ECM)
obtained from 24 h culture of HUVECs. At a ratio of 1:1, ECM was
evenly mixed with DMEM, and then was subsequently treated to HepG2
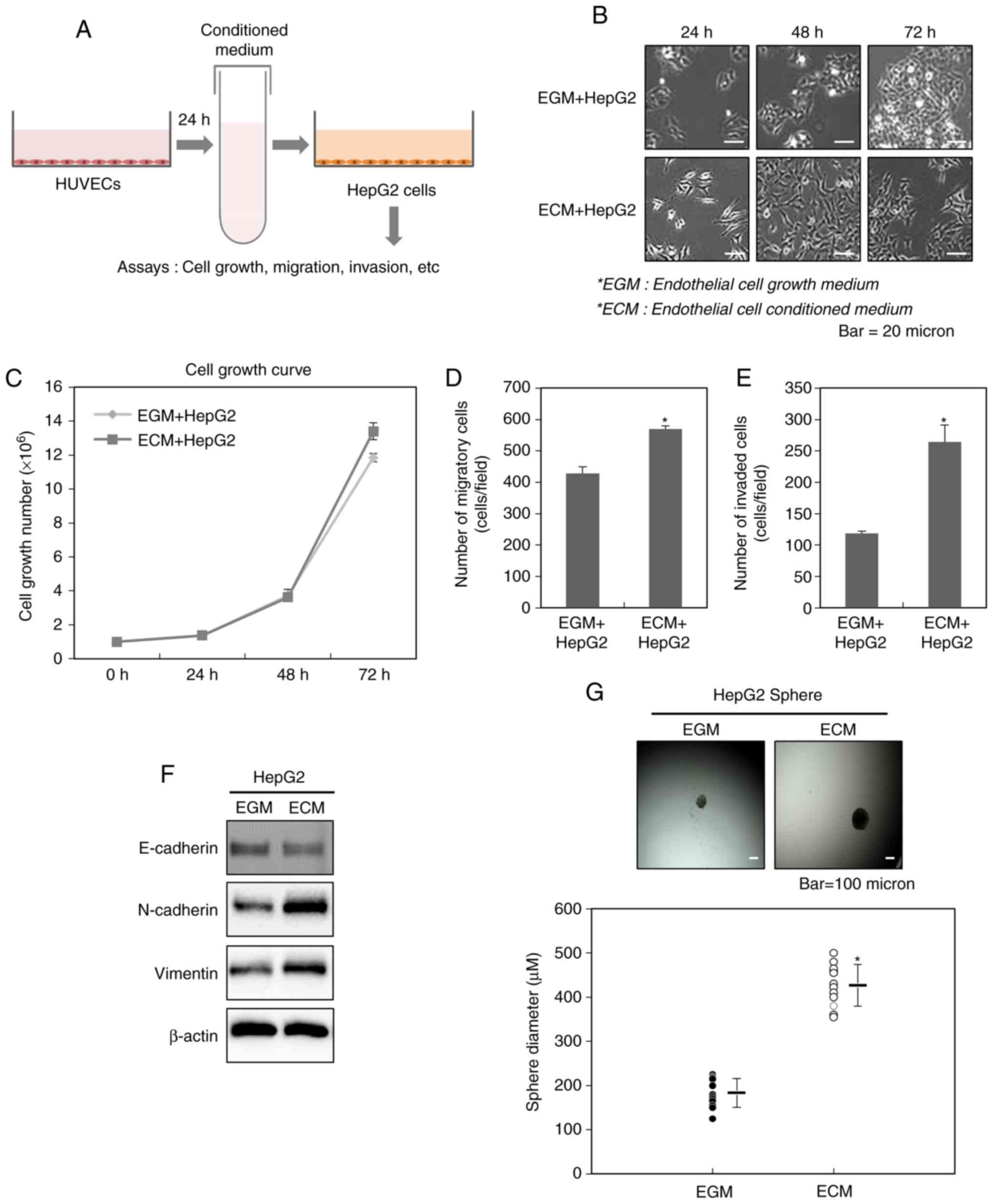
cells (Fig. 1A).
We first observed the changes in the cellular
morphology of HepG2 cells after the treatment with ECM obtained
from HUVECs. As shown in previous reports mentioning these changes
observed during the process of EMT (19–21),
Fig. 1B also showed that the
treatment with ECM for 24, 48 and 72 h leads to the appearance of a
fibroblast-like shape emerging as elongated and dispersed HepG2
cells, compared with control. This result indicates that
endothelial cells may contribute to the acquisition of mesenchymal
traits of liver cancer cells.
We next examined the effect of ECM on the cellular
growth of HepG2 cells. As shown in Fig.
1C, the growth rate of HepG2 cells did not be greatly altered
by the treatment with ECM.
The acquisition of mesenchymal traits of cancer
cells leading to the malignant properties of them is closely
related with the process of EMT (9,12–14). We therefore investigated whether ECM
obtained from HUVECs affects EMT of HepG2 cells. Fig. 1D and E revealed that HepG2 cells
treated with ECM show more migratory and invasive properties,
compared with control. Consistent with these results, we also found
an increase in the expression levels of the mesenchymal markers
N-cadherin and vimentin in HepG2 cells treated with ECM, compared
with control (Fig. 1F).
EMT has been well known to be closely associated to
an increase in the cancer stem-like cell population (9,12–14). Thus, we examined whether ECM obtained
from HUVECs can also change the numbers of cancer stem-like cells.
To fulfill this purpose, we performed sphere forming assay after
the treatment of HepG2 cells with ECM. As shown in Fig. 1G, the cells shows the larger spheres,
compared with control, indicating that endothelial cells can assign
the higher sphere-forming ability to liver cancer cells.
These data collectively suggest that endothelial
cells play a key role in inducing EMT of liver cancer cells and
expanding cancer stem-like cells.
Effect of irradiated endothelial cells
on the cell survival of liver cancer cells
Ionizing radiation influence tumor microenvironment
(22,23). Especially, the tumor vascular system
composing tumor microenvironment has been reported to be highly
sensitive to irradiation (23,24). Thus,
we investigated whether irradiated endothelial cells can affect the
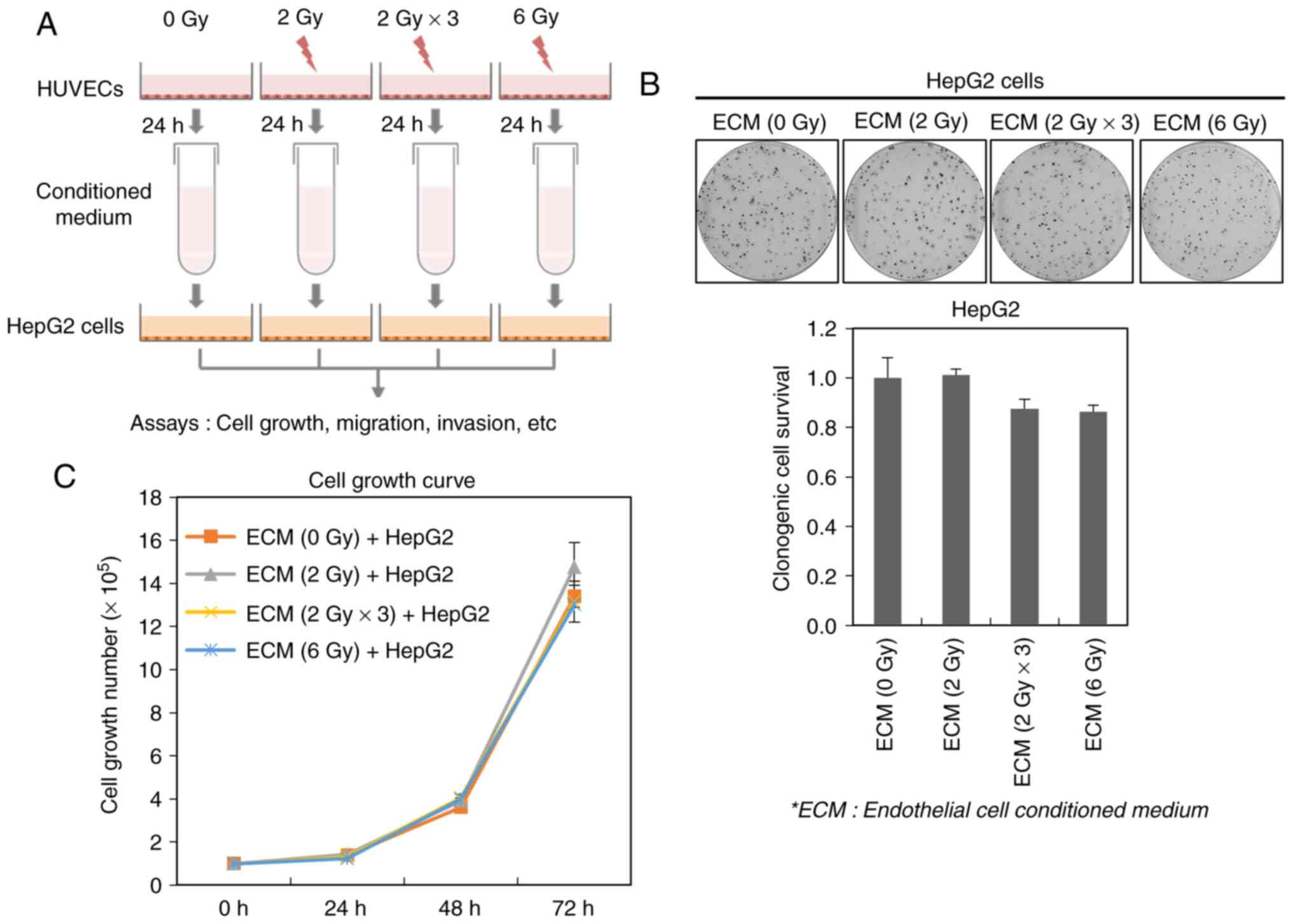
viability of cancer cells. To fulfill this purpose, we acquired ECM
obtained from HUVECs exposed to a single dose (2 or 6 Gy) or
fractionated dose (2 Gy × 3; 2 Gy/day for 3 days) of ionizing
radiation, and then treated it to HepG2 cells (Fig. 2A). Although either the fractionated
dose-irradiated (2 Gy × 3)- or 6 Gy-irradiated ECM slightly
suppressed the clonogenic survival of HepG2 cells, their survival
was not greatly affected by all irradiated ECMs, indicating that
irradiated endothelial cells do not seem to highly influence the
survival of cancer cells (Fig. 2B).
In addition, we further found that all irradiated ECMs do not
modulate the growth rate of HepG2 cells (Fig. 2C).
These data suggest that irradiated endothelial cells
do not affect the cell survival and growth of liver cancer
cells.
Irradiated endothelial cells modulates
tumor malignancy of liver cancer cells
To further investigate the connection between
irradiated endothelial cells and liver cancer cells in the tumor
malignancy, we first measured the migratory and invasive abilities
of HepG2 cells after the treatments with ECM obtained from
irradiated HUVECs. As shown in Fig. 3A
and B, we found that 2 Gy-irradiated ECM greatly increases the
migratory and invasive traits of HepG2 cells, compared with the
treatment with ECM obtained from non-irradiated endothelial cells,
whereas 6 Gy- or 2 Gy × 3 (2 Gy/day for 3 days) irradiated ECM does
hardly affect these events. Consistent with the above results, we
further observed that 2 Gy-irradiated ECM greatly increase the
expression levels of the mesenchymal cell markers N-cadherin and
vimentin in HepG2 cells, whereas the expression of the epithelial
cell marker, E-cadherin is not greatly modulated (Fig. 3C). In addition, we investigated
whether 2 Gy-irradiated ECM affects the expression levels of
EMT-regulating transcription factors, Slug, Zeb1 and Snail in HepG2
cells. Although the expression levels of Zeb1 and Snail did not be
change, an increase in the expression levels of Slug was observed
in HepG2 cells (Fig. 3C).
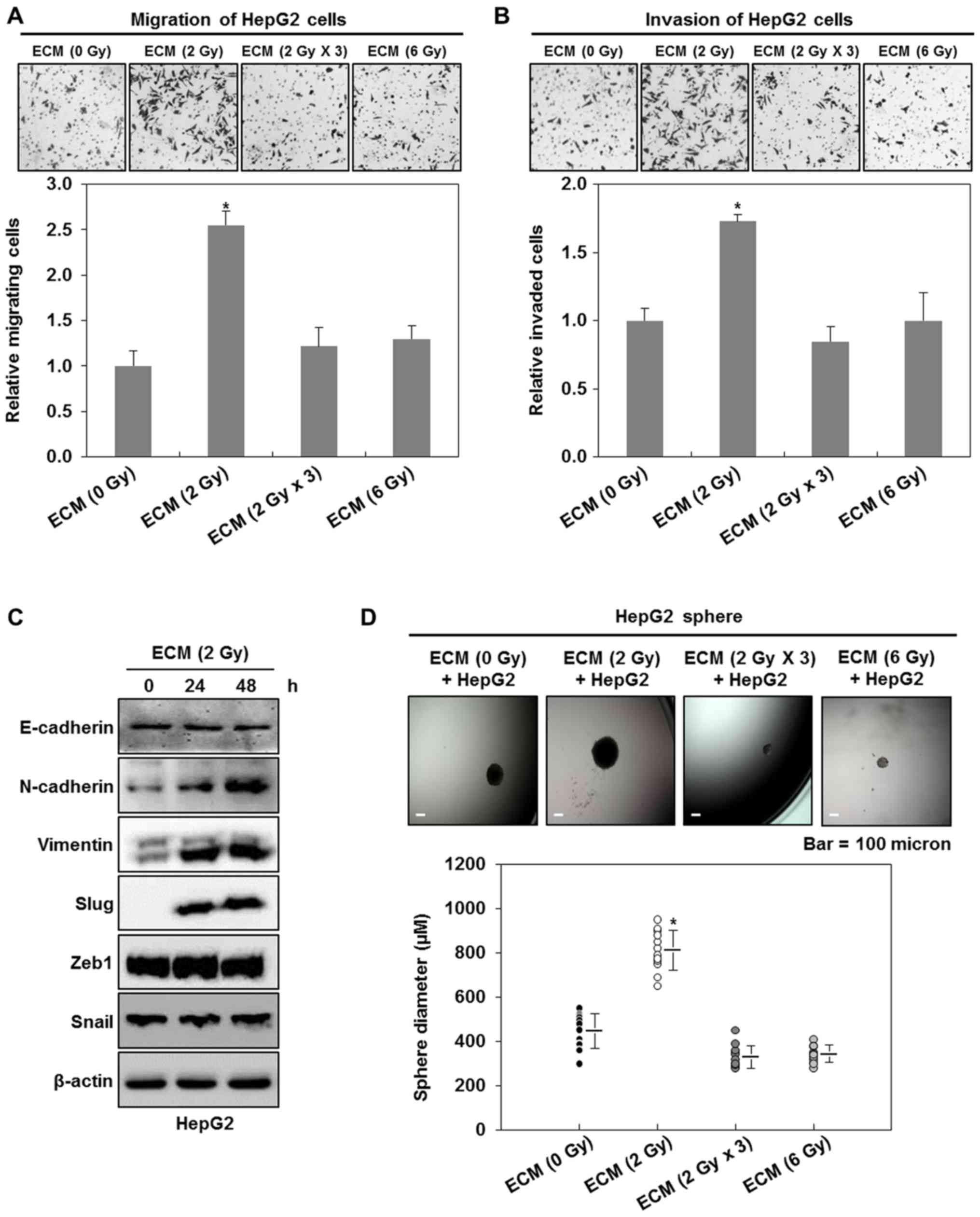 | Figure 3.2 Gy-irradiated endothelial cells
enhance the malignant potential of liver cancer cells. (A)
Migratory and (B) invasive properties of HepG2 cells after the
treatment with ECM obtained from irradiated HUVECs. These
properties of the cells were measured using the Transwell chamber
(×200 magnification). (C) Western blot analysis for the expressions
of E-cadherin, N-cadherin, vimentin, slug, zeb1 and snail after the
treatment with ECM. Experiments were performed in triplicate, and
the data shown are representative of a typical experiment. (D)
Quantification of sphere-forming abilities of HepG2 cells after the
treatment with ECM obtained from irradiated HUVECs. The cells were
grown in DMEM/F12 supplemented with B27, N2, basic fibroblast- and
epidermal growth factor onto 24-well ultra-low attachment plates at
300 cells per well for 7 days, and the size of spheres were
determined. To measure the size of sphere, 12 spheres per group
(n=12/group) were randomly selected. The average size of each
sphere is quantified in the standard deviation and shown in the
representative graph. *P<0.05 vs. control. HUVECs, human
umbilical vein endothelial cells; ECM, endothelial cell culture
medium. (E) Migratory and (F) invasive properties of Hep3B or Huh7
cells after the treatment with ECM obtained from 2 Gy-irradiated
HUVECs. These properties of the cells were measured using the
Transwell chamber (×200 magnification). (G) Quantification of
sphere-forming abilities of Hep3B or Huh7 cells after the treatment
with ECM obtained from 2 Gy-irradiated HUVECs. The cells were grown
in DMEM/F12 supplemented with B27, N2, basic fibroblast- and
epidermal growth factor onto 24-well ultra-low attachment plates at
300 cells per well for 7 days, and the size of spheres were
determined. To measure the size of sphere, 12 spheres per group
(n=12/group) were randomly selected. The average size of each
sphere is quantified in the standard deviation and shown in the
representative graph. Results from three independent experiments
are expressed as mean ± 1 SEM *P<0.05 vs. control. HUVECs, human
umbilical vein endothelial cells; ECM, endothelial cell culture
medium. |
EMT has been well known to be an event leading to an
increase in the population of cancer stem-like cells (9,12–14). Thus, we next investigated whether the
treatments with ECM obtained from irradiated HUVECs also affect the
size of cancer stem-like cells in HepG2 cells. As shown in Fig. 3D, 2 Gy-irradiated ECM induces larger
spheres of HepG2 cells, whereas 6 Gy- or 2 Gy × 3 (2 Gy/day for 3
days)-irradiated ECM does slightly decrease the size of HepG2
spheres, indicating that 2 Gy-irradiated endothelial cells
contribute to the higher sphere-forming ability of liver cancer
cells. When we confirmed these events using the other liver cancer
cell lines, Hep3B and Huh7 cells, we had similar results to those
obtained from HepG2 cells (Fig.
3E-G).
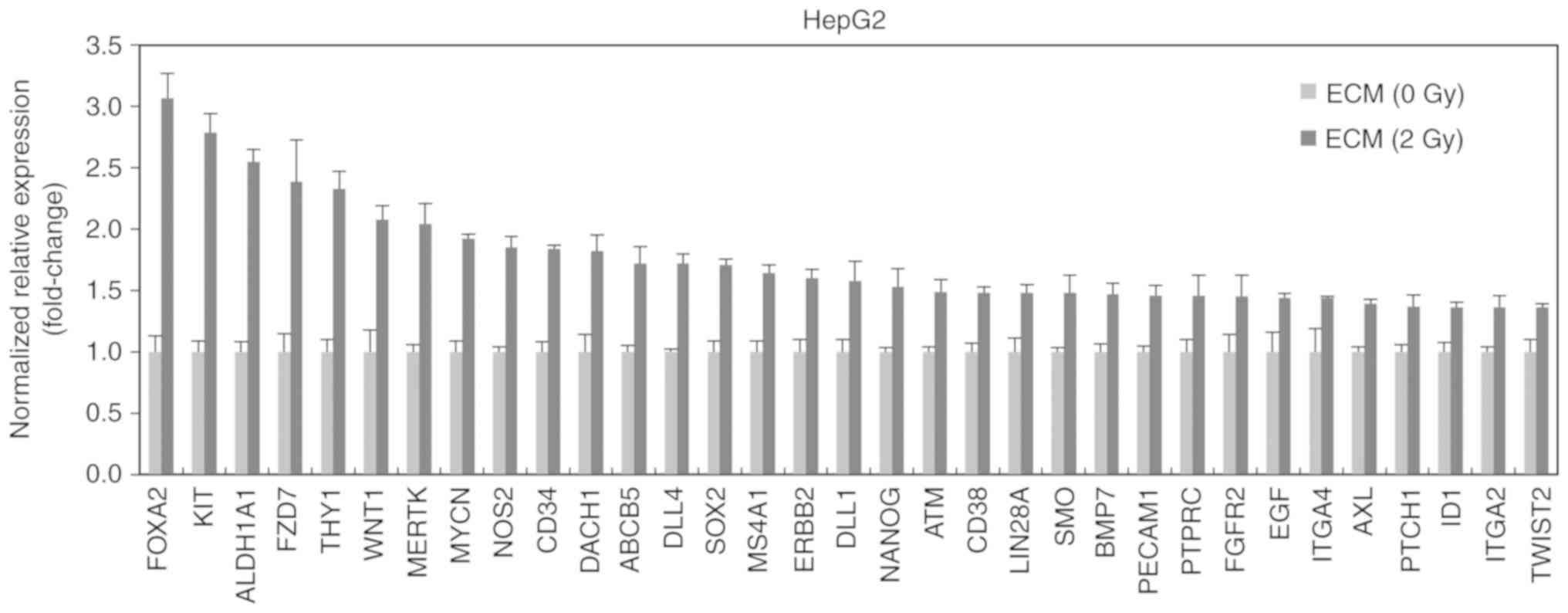
In addition, we determined the level of cancer
stem-like cell-related factors after the treatment with 2
Gy-irradiated ECM in HepG2 cells. 2 Gy-irradiated ECM increased the
mRNA levels of Foxa2, c-Kit, ALDH1A1, FZD7 ((Frizzled 7) and Thy1
(CD90) more than 2 times (Fig. 4).
These factors are already known to contribute to stemness of cancer
cells (25–29).
These results collectively demonstrated that 2
Gy-irradiated endothelial cells can trigger the generation of
cancer stem-like cells which are thought to be closely connected
with the process of EMT.
Discussion
Although radiotherapy is widely known as a highly
efficient treatment for a variety of cancer patients, tumor relapse
and metastasis occurring after radiotherapy still remain as a major
problem to be solved (4,5). In addition, they have been shown to be
the main causes leading to poor survival rates of cancer patients
(4,5).
Tumor malignancy is characterized by them, which are closely
related with the expansion of cancer stem-like cells (12,30). EMT
is considered as a primary mechanism responsible for the expansion
of cancer stem-like cells (9,12,30).
Tumor microenvironment is known to contribute to the
induction of EMT, and the maintenance and survival of cancer
stem-like cells (31,32). Furthermore, the induction of EMT, and
the maintenance and survival of cancer stem-like cells can be
regulated by tumor microenvironment comprised of cancer cells and
diverse stromal cells containing immune cells, fibroblasts and
endothelial cells (12,30–32).
Especially, endothelial cells are suggested to be a key determinant
of tumor microenvironment as they are a main component of vascular
system responsible for delivering oxygen and nutrients to tumor
cells (15,16).
In this study, we preferentially established a
co-cultivation system to find a potential role of endothelial cells
in the malignancy of liver cancer cells using ECM conditioned by
endothelial cells, and then observed that the treatment with ECM
increases the migratory and invasive properties of liver cancer
cells, and leads to the expansion of cancer stem-like cells, as
well as the appearance of a fibroblast-like shape and the
dispersal, indicating that endothelial cells are responsible for
the acquisition of mesenchymal traits of liver cancer cells. This
observation is in good agreement with previous reports that
endothelial cells are involved in both the induction of EMT and the
generation of cancer stem-like phenotype in various tumor types
including glioblastomas, squamous cell carcinomas, colorectal and
breast cancers (18,33–36).
There are many investigations indicating the
functional and morphological changes in irradiated vasculatures in
normal tissues (37–39), but is relatively little known about
the effects of ionizing radiation on tumor vasculatures. However,
it has been already suggested that endothelial cells in tumor
microenvironments are rather sensitive to ionizing radiation so
that irradiation-induced apoptotic cell death of endothelial cells
in tumor vasculatures plays a key role in the reduction of tumor
size caused by irradiation (39–41).
Controversially, it has been reported that ionizing radiation does
not only lead to the capillary-like tube formation of endothelial
cells, but also promotes the metastatic properties of cancer cells
(42,43).
Most of previous studies concerned with tumor
microenvironments have largely focused on the direct influences of
ionizing radiation on the cells, but not the indirect influences of
it (44,45). Thus, additional studies are needed to
clarify the indirect effects of ionizing radiation on the tumor
microenvironments. As part of these studies, we investigated
whether irradiated endothelial cells affect the tumor malignancy of
liver cancer cells through the treatment with ECM conditioned by
endothelial cells irradiated with 2 Gy, fractionated dose (2 Gy ×
3; 2 Gy/day for 3 days) or 6 Gy.
Direct exposure of the cells to ionizing radiation
has been well known to efficiently suppress the clonogenic cell
survival and the cell proliferation (44,45).
However, although the clonogenic cell survival assay showed a
slight decline in the cell survival of liver cancer cells treated
with ECM conditioned by endothelial cells irradiated with
fractionated dose (2 Gy × 3; 2 Gy/day for 3 days) or 6 Gy, we did
not observe the efficient suppression of cell survival in liver
cancer cells after the treatments with all irradiated ECMs.
Previously, many evidences indicated that ionizing radiation can
not only affect the directly irradiated cells, but also influence
the non-irradiated cells surrounding them (3). This biological phenomenon is termed as
the radiation-induced bystander effect (44). In addition, the bystander effect has
been reported to emerge as cellular and molecular events including
DNA damage response, cell cycle arrest, cell growth delay, cell
transformation and cell death in the non-irradiated cells (3,44,45). Unlike these reports, we did not find
the decrease in the survival and proliferation of non-irradiated
liver cancer cells after the treatment with all irradiated ECMs.
These results may be due to the difference in cellular responses of
non-irradiated cells in accordance with cell types, the genetic and
functional traits of the cells.
Many evidences showed that EMT greatly contributes
to tumor malignancy via inducing metastasis and triggering tumor
relapse (6–9). Furthermore, it is significantly
concerned with the generation of cancer stem-like cells (12–14). Thus,
we examined whether irradiated endothelial cells affect the
induction of EMT in liver cancer cells. Interestingly, we found
that ECM conditioned by endothelial cells irradiated with 2 Gy
greatly increases the migratory and invasive properties of liver
cancer cells, as well as inducing mesenchymal markers. Furthermore,
it also efficiently enhanced the sphere-forming ability of liver
cancer cells, and increased the mRNA levels of Foxa2, c-Kit,
ALDH1A1, FZD7 and Thy1 (CD90) known to regulate self-renewal of
cancer stem cells. However, our results showed that either ECM
conditioned by endothelial cells irradiated with fractionated dose
(2 Gy × 3; 2 Gy/day for 3 days) or that with 6 Gy does not greatly
influence the malignancy of liver cancer cells. Particularly, c-Kit
and FZD7 are well known to be the receptors for Stem Cell Factor
(SCF), a cytokine and secreted WNT proteins, respectively (26,28).
Therefore, it can be reasonably assumed that these increased
receptors of liver cancer cells caused by irradiated endothelial
cells may participate in enhancing the induction of malignant liver
cancer cells via interacting with their soluble ligands secreted
from a variety of stromal cells in tumor microenvironment in
vivo. In a good agreement with our assumption, it has been
commonly indicated that soluble factors secreted from endothelial
cells play a key role in tumor malignancy (18,33–36).
Especially, the communications between one cell and the others can
be mediated by these soluble signaling molecules including
cytokines and growth factors (15,17,18). These
factors such as interleukin-6 (IL-6), IL-8, transforming growth
factor-β1 (TGF-β1) and tumor necrosis factor-alpha (TNF-α), have
been reported to play a pivotal role in the cell-to-cell
communications and the bystander effects (46). Moreover, they do not only contribute
to tumor malignancy, but also are induced by ionizing radiation
(46). The principal reason why the
malignant potential of liver cancer cells is raised only by ECM
conditioned by endothelial cells irradiated with 2 Gy but not by
the others may be due to these secreted factors. Thus, it is
necessary to further identify that what kind of them is highly
secreted from endothelial cells in response to 2 Gy irradiation,
and greatly contributes to the malignancy of liver cancer cells,
compared with them secreted from endothelial cells irradiated with
fractionated dose (2 Gy × 3; 2 Gy/day for 3 days) or 6 Gy. In
addition, although we performed the investigation only under three
conditions (2 Gy, fractionated dose (2 Gy/day for 3 days) and 6
Gy), endothelial cells irradiated with various doses of ionizing
radiation are also likely to diversely affect the biological events
of tumor cells. However, there have been no comparative studies
investigating these effects as yet. Thus, further studies are also
needed to precisely define the pivotal role of these irradiated
endothelial cells in the tumor malignancy.
It have been already shown that direct exposure of a
variety of cancer cells to ionizing radiation can trigger tumor
malignancy via increasing the migratory and invasive properties of
them, and expanding the population of cancer stem-like cells
(47–50). Similarly to these reports, we found a
possibility that the indirect exposure to ionizing radiation
mediated by irradiated endothelial cells can also elicit the highly
malignant potential from cancer cells.
In summary, we found that 2 Gy irradiation of
endothelial cells influences the increase in the tumor malignancy
of liver cancer cells. Our observations indicate that the distinct
differences in the indirect effects of ionizing radiation on tumor
malignancy may provide a valuable clue to the improvement in the
efficacy of radiotherapy.
Acknowledgements
Not applicable.
Funding
This study was supported by the National Research
Foundation of Korea grant funded by the Korea government (MSIP)
(grant no. 50596-2018).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
SDK performed all functional assays, and contributed
to conception, design, collection and assembly of data. JMY
contributed to data analysis and interpretation, and wrote the
manuscript. MTP contributed to data analysis and interpretation,
and wrote the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Baskar R and Itahana K: Radiation therapy
and cancer control in developing countries: Can we save more lives?
Int J Med Sci. 14:13–17. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Datta NR, Samiei M and Bodis S:
Radiotherapy infrastructure and human resources in Europe-present
status and its implications for 2020. Eur J Cancer. 50:2735–2743.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stapleton S, Jaffray D and Milosevic M:
Radiation effects on the tumor microenvironment: Implications for
nanomedicine delivery. Adv Drug Deliv Rev. 109:119–130. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Begg AC, Stewart FA and Vens C: Strategies
to improve radiotherapy with targeted drugs. Nat Rev Cancer.
11:239–253. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ogawa K, Yoshioka Y, Isohashi F, Seo Y,
Yoshida K and Yamazaki H: Radiotherapy targeting cancer stem cells:
Current views and future perspectives. Anticancer Res. 33:747–754.
2013.PubMed/NCBI
|
|
6
|
Marie-Egyptienne DT, Lohse I and Hill RP:
Cancer stem cells, the epithelial to mesenchymal transition (EMT)
and radioresistance: Potential role of hypoxia. Cancer Lett.
341:63–72. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rankin EB and Giaccia AJ: Hypoxic control
of metastasis. Science. 352:175–180. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
De Craene B and Berx G: Regulatory
networks defining EMT during cancer initiation and progression. Nat
Rev Cancer. 13:97–110. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kim JH, Shim JW, Eum DY, Kim SD, Choi SH,
Yang K, Heo K and Park MT: Downregulation of UHRF1 increases tumor
malignancy by activating the CXCR4/AKT-JNK/IL-6/Snail signaling
axis in hepatocellular carcinoma cells. Sci Rep. 7:27982017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Moreno-Bueno G, Portillo F and Cano A:
Transcriptional regulation of cell polarity in EMT and cancer.
Oncogene. 27:6958–6969. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vasko V, Espinosa AV, Scouten W, He H,
Auer H, Liyanarachchi S, Larin A, Savchenko V, Francis GL, de la
Chapelle A, et al: Gene expression and functional evidence of
epithelial-to-mesenchymal transition in papillary thyroid carcinoma
invasion. Proc Natl Acad Sci USA. 104:2803–2808. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Singh A and Settleman J: EMT, cancer stem
cells and drug resistance: An emerging axis of evil in the war on
cancer. Oncogene. 29:4741–4751. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Asiedu MK, Beauchamp-Perez FD, Ingle JN,
Behrens MD, Radisky DC and Knutson KL: AXL induces
epithelial-to-mesenchymal transition and regulates the function of
breast cancer stem cells. Oncogene. 33:1316–1324. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hwang-Verslues WW, Chang PH, Wei PC, Yang
CY, Huang CK, Kuo WH, Shew JY, Chang KJ, Lee EY and Lee WH: miR-495
is upregulated by E12/E47 in breast cancer stem cells, and promotes
oncogenesis and hypoxia resistance via downregulation of E-cadherin
and REDD1. Oncogene. 30:2463–2474. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Korkaya H, Liu S and Wicha MS: Breast
cancer stem cells, cytokine networks, and the tumor
microenvironment. J Clin Invest. 121:3804–3809. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Jain RK: Normalization of tumor
vasculature: An emerging concept in antiangiogenic therapy.
Science. 307:58–62. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhu TS, Costello MA, Talsma CE, Flack CG,
Crowley JG, Hamm LL, He X, Hervey-Jumper SL, Heth JA, Muraszko KM,
et al: Endothelial cells create a stem cell niche in glioblastoma
by providing NOTCH ligands that nurture self-renewal of cancer
stem-like cells. Cancer Res. 71:6061–6072. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lu J, Ye X, Fan F, Xia L, Bhattacharya R,
Bellister S, Tozzi F, Sceusi E, Zhou Y, Tachibana I, et al:
Endothelial cells promote the colorectal cancer stem cell phenotype
through a soluble form of Jagged-1. Cancer Cell. 23:171–185. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lai JK, Wu HC, Shen YC, Hsieh HY, Yang SY
and Chang CC: Krüppel-like factor 4 is involved in cell scattering
induced by hepatocyte growth factor. J Cell Sci. 125:4853–4864.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Thuault S, Valcourt U, Petersen M,
Manfioletti G, Heldin CH and Moustakas A: Transforming growth
factor-beta employs HMGA2 to elicit epithelial-mesenchymal
transition. J Cell Biol. 174:175–183. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kimura-Tsuchiya R, Ishikawa T, Kokura S,
Mizushima K, Adachi S, Okajima M, Matsuyama T, Okayama T, Sakamoto
N, Katada K, et al: The inhibitory effect of heat treatment against
epithelial-mesenchymal transition (EMT) in human pancreatic
adenocarcinoma cell lines. J Clin Biochem Nutr. 55:56–61. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nguyen DH, Oketch-Rabah HA, Illa-Bochaca
I, Geyer FC, Reis-Filho JS, Mao JH, Ravani SA, Zavadil J, Borowsky
AD, Jerry DJ, et al: Radiation acts on the microenvironment to
affect breast carcinogenesis by distinct mechanisms that decrease
cancer latency and affect tumor type. Cancer Cell. 19:640–651.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Karar J and Maity A: Modulating the tumor
microenvironment to increase radiation responsiveness. Cancer Biol
Ther. 8:1994–2001. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kuonen F, Secondini C and Rüegg C:
Molecular pathways: Emerging pathways mediating growth, invasion,
and metastasis of tumors progressing in an irradiated
microenvironment. Clin Cancer Res. 18:5196–5202. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Peng Q, Qin J, Zhang Y, Cheng X, Wang X,
Lu W, Xie X and Zhang S: Autophagy maintains the stemness of
ovarian cancer stem cells by FOXA2. J Exp Clin Cancer Res.
36:1712017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hassan HT: c-Kit expression in human
normal and malignant stem cells prognostic and therapeutic
implications. Leuk Res. 33:5–10. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Condello S, Morgan CA, Nagdas S, Cao L,
Turek J, Hurley TD and Matei D: β-Catenin-regulated ALDH1A1 is a
target in ovarian cancer spheroids. Oncogene. 34:2297–2308. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chakrabarti R, Wei Y, Hwang J, Hang X,
Andres Blanco M, Choudhury A, Tiede B, Romano RA, DeCoste C,
Mercatali L, et al: ΔNp63 promotes stem cell activity in mammary
gland development and basal-like breast cancer by enhancing Fzd7
expression and Wnt signalling. Nat Cell Biol. 16:1004–1015, 1-13.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang ZF, Ho DW, Ng MN, Lau CK, Yu WC, Ngai
P, Chu PW, Lam CT, Poon RT and Fan ST: Significance of CD90+ cancer
stem cells in human liver cancer. Cancer Cell. 13:153–166. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Scheel C and Weinberg RA: Cancer stem
cells and epithelial-mesenchymal transition: Concepts and molecular
links. Semin Cancer Biol. 22:396–403. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Carnero A and Lleonart M: The hypoxic
microenvironment: A determinant of cancer stem cell evolution.
Bioessays. 38 Suppl 1:S65–S74. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
van der Horst G, Bos L and van der Pluijm
G: Epithelial plasticity, cancer stem cells, and the
tumor-supportive stroma in bladder carcinoma. Mol Cancer Res.
10:995–1009. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sharma A and Shiras A: Cancer stem
cell-vascular endothelial cell interactions in glioblastoma.
Biochem Biophys Res Commun. 473:688–692. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Charles N, Ozawa T, Squatrito M, Bleau AM,
Brennan CW, Hambardzumyan D and Holland EC: Perivascular nitric
oxide activates notch signaling and promotes stem-like character in
PDGF-induced glioma cells. Cell Stem Cell. 6:141–152. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhang Z, Dong Z, Lauxen IS, Filho MS and
Nör JE: Endothelial cell-secreted EGF induces epithelial to
mesenchymal transition and endows head and neck cancer cells with
stem-like phenotype. Cancer Res. 74:2869–2881. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Sigurdsson V, Hilmarsdottir B,
Sigmundsdottir H, Fridriksdottir AJ, Ringnér M, Villadsen R, Borg
A, Agnarsson BA, Petersen OW, Magnusson MK and Gudjonsson T:
Endothelial induced EMT in breast epithelial cells with stem cell
properties. PLoS One. 6:e238332011. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li YQ, Ballinger JR, Nordal RA, Su ZF and
Wong CS: Hypoxia in radiation-induced blood-spinal cord barrier
breakdown. Cancer Res. 61:3348–3354. 2001.PubMed/NCBI
|
|
38
|
Wilson CM, Gaber MW, Sabek OM, Zawaski JA
and Merchant TE: Radiation-induced astrogliosis and blood-brain
barrier damage can be abrogated using anti-TNF treatment. Int J
Radiat Oncol Biol Phys. 74:934–941. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Park MT, Oh ET, Song MJ, Kim WJ, Cho YU,
Kim SJ, Han JY, Suh JK, Choi EK, Lim BU, et al: The
radiosensitivity of endothelial cells isolated from human breast
cancer and normal tissue in vitro. Microvasc Res. 84:140–148. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Fuks Z and Kolesnick R: Engaging the
vascular component of the tumor response. Cancer Cell. 8:89–91.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Garcia-Barros M, Paris F, Cordon-Cardo C,
Lyden D, Rafii S, Haimovitz-Friedman A, Fuks Z and Kolesnick R:
Tumor response to radiotherapy regulated by endothelial cell
apoptosis. Science. 300:1155–1159. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Asuthkar S, Velpula KK, Nalla AK, Gogineni
VR, Gondi CS and Rao JS: Irradiation-induced angiogenesis is
associated with an MMP-9-miR-494-syndecan-1 regulatory loop in
medulloblastoma cells. Oncogene. 33:1922–1933. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Zhou YC, Liu JY, Li J, Zhang J, Xu YQ,
Zhang HW, Qiu LB, Ding GR, Su XM, Mei-Shi and Guo GZ: Ionizing
radiation promotes migration and invasion of cancer cells through
transforming growth factor-beta-mediated epithelial-mesenchymal
transition. Int J Radiat Oncol Biol Phys. 81:1530–1537. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Prise KM and O'Sullivan JM:
Radiation-induced bystander signalling in cancer therapy. Nat Rev
Cancer. 9:351–360. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Powathil GG, Munro AJ, Chaplain MA and
Swat M: Bystander effects and their implications for clinical
radiation therapy: Insights from multiscale in silico experiments.
J Theor Biol. 401:1–14. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Najafi M, Fardid R, Hadadi G and Fardid M:
The mechanisms of radiation-induced bystander effect. J Biomed Phys
Eng. 4:163–172. 2014.PubMed/NCBI
|
|
47
|
Kim RK, Cui YH, Yoo KC, Kim IG, Lee M,
Choi YH, Suh Y and Lee SJ: Radiation promotes malignant phenotypes
through SRC in breast cancer cells. Cancer Sci. 106:78–85. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Gomez-Casal R, Bhattacharya C, Ganesh N,
Bailey L, Basse P, Gibson M, Epperly M and Levina V: Non-small cell
lung cancer cells survived ionizing radiation treatment display
cancer stem cell and epithelial-mesenchymal transition phenotypes.
Mol Cancer. 12:942013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kim RK, Suh Y, Cui YH, Hwang E, Lim EJ,
Yoo KC, Lee GH, Yi JM, Kang SG and Lee SJ: Fractionated
radiation-induced nitric oxide promotes expansion of glioma
stem-like cells. Cancer Sci. 104:1172–1177. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Su Z, Li G, Liu C, Ren S, Tian Y, Liu Y
and Qiu Y: Ionizing radiation promotes advanced malignant traits in
nasopharyngeal carcinoma via activation of epithelial-mesenchymal
transition and the cancer stem cell phenotype. Oncol Rep. 36:72–78.
2016. View Article : Google Scholar : PubMed/NCBI
|