Introduction
Colon cancer is the fourth most common cancer and is
one of the leading causes of cancer-related mortality in the United
States (1–5). Numerous therapies have been developed
with certain therapies involving direct targeting of the cancer,
modulating the immune system or blocking angiogenesis (6–9); however,
>60% have failed in clinical trials for various reasons
(10,11). This high failure rate in clinical
trials has caused there to be increasing research attention on the
tumor microenvironment (TME) and on novel therapies capable of
targeting the TME (12–14). Previous studies have demonstrated that
colon cancer proliferation is not only dependent on cell intrinsic
properties, but is also greatly influenced by the TME (15,16). Colon
cancer cells may also modulate the TME to create a more favorable
environment for proliferation and invasion (17,18).
Therefore, the TME and its components may serve as important
factors in colon cancer proliferation (19,20).
Fibroblasts, as the building blocks of connective tissues, are key
components of the TME (21–23). It has been proposed that
cancer-associated fibroblasts (CAFs) aid tumors to grow and are
associated with all stages of cancer (24,25).
Meanwhile, other research has suggested that normal fibroblasts
inhibit cancer cell proliferation and metastasis at the during
early-stage cancer (26). While
studies into CAFs and their influence on cancer progression have
been conducted (27,28), systematic studies on fibroblasts and
their co-culture effects on the proliferation and drug response of
colon cancer cells are rare. Furthermore, interactions between
colon cancer cells and fibroblasts from different tissues are
relatively unexplored and may provide insight into the
proliferation of early stage or metastatic cancer. To address this
unresolved subject, the present study established 2D and
anchorage-independent colon cancer cell/fibroblast co-culture
systems. Using these, the co-culture effects of fibroblasts on
cancer cell proliferation and apoptosis were studied, with
particular focus on the positive or negative regulatory role of
different fibroblast cell lines on colon cancer growth. In
addition, the response of colon cancer cells to the Wnt/β-catenin
inhibitor XAV939 as well as other anticancer agents, including
5-fluorouracil (5-FU), doxorubicin (DOX), camptothecin (CPT) and
irinotecan (IRI), was examined in the presence or absence of
fibroblasts to determine the co-culture effects.
Materials and methods
Cells and reagents
The SW480, HT-29, DLD-1 and Caco-2 colon cancer cell
lines were obtained from the American Type Culture Collection
(ATCC, Manassas, VA, USA). Lung fibroblasts (WI-38), skin
fibroblasts (BJ) and colon fibroblasts (CCD-18Co) were also
purchased from the ATCC. Ultrapure water (molecular biology grade),
RPMI-1640 medium, fetal bovine serum (FBS), 10× tris-buffered
saline and Dulbecco's phosphate-buffered saline were purchased from
Welgene (Gyeongsan, Korea). RPMI-1640 medium powder, TrypLE Express
and penicillin-streptomycin were purchased from Thermo Fisher
Scientific, Inc., (Waltham, MA, USA). Agar powder, agar
solubilization solution, 8× lysis buffer and Cyqant® GR
dye were purchased from Cell Biolabs, Inc., (San Diego, CA, USA).
Cell culture flasks (25 and 75 cm2), cell culture dishes
(60×15 mm), 24-well clear flat-bottom TC-treated multiwall cell
culture plates and 6.5-mm Transwell® polycarbonate
membrane inserts with 0.4-µm pores were purchased from Corning
(Corning, NY, USA). XAV939, IRI, CPT, DOX and sodium bicarbonate
were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
5-FU and a Muse Caspase-3/7 assay kit were also from Merck KGaA. A
Cell Counting Kit-8 (CCK-8) was purchased from Dojindo Laboratories
(Kumamoto, Japan). Pre-cast gels for western blotting were
purchased from Bio-Rad Laboratories, Inc., (Hercules, CA, USA).
Amersham™ ECL™ Select western blot detection
reagent was purchased from GE Healthcare (Chicago, IL, USA).
Colon cancer cell co-culture with
fibroblasts
The SW480, HT-29, DLD-1 and Caco-2 colon cancer
cells and WI-38, CCD-18Co and BJ fibroblasts were cultured in
RPMI-1640 medium supplemented with 10% FBS and 1%
penicillin-streptomycin at 37°C in a 5% CO2 incubator.
For the co-culture, 30,000 SW480, HT-29, DLD-1 and Caco-2 cells
were seeded onto 24-well plates with 500 µl RPMI-1640 medium.
Additionally, 30,000 WI-38, CCD-18Co and BJ cells were seeded onto
Transwell® inserts with 100 µl RPMI-1640 medium. After 6
h, the fibroblasts seeded onto Transwell® inserts were
transferred to a 24-well plate for co-culturing with colon cancer
cells. As a blank control, 100 µl RPMI-1640 medium without
fibroblasts in Transwell® inserts was prepared. Media
were replaced with fresh RPMI-1640 medium every 2 days. Colon
cancer cell proliferation on day 5 in the presence or absence of
fibroblasts was determined with a CCK-8 assay. Briefly, 50 µl CCK-8
solution was added to each well and incubated for 2 h, following
which the absorbance at 450 nm was measured using a microplate
reader (SpectraMax i3; Molecular Devices, San Jose, CA, USA). The
apoptotic rate of co-cultured colon cancer cells was measured with
a Muse® cell analyzer (Merck KGaA) using the
Muse® Caspase-3/7 Assay kit. Cell images were obtained
using an Olympus IX 73 inverted microscope (Tokyo, Japan).
Anchorage-independent colon cancer
cell culture
To optimize anchorage-independent cell culture,
200–350 µl of 0.1–2.5% agar solution was mixed with RPMI-1640
medium containing 5,000-200,000 colon cancer cells, solidified at
4°C for 3–30 min in 24-well plates and cancer cell proliferation
was assessed. Following optimization, 300 µl of 0.6% agar mixed
with RPMI-1640 medium containing 30,000 colon cancer cells was
solidified for 20 min at 4°C on top of 200 µl of 0.6% agar
containing RPMI-1640 medium. Subsequently, 400 µl RPMI-1640 medium
was added to each well. For fibroblast co-culture, 30,000 WI-38,
CCD-18Co or BJ cells seeded in 24-well Transwell®
inserts were placed on top of anchorage-independent cultured colon
cancer cells. For quantitation of anchorage-independent colon
cancer cell growth, the agar-cell layer was solubilized and
fluorescence at 520 nm was measured following the addition of
CyQuant® dye. The apoptotic rates of the
anchorage-independent cultured colon cancer cells were measured
with the Muse® cell analyzer and the Muse®
Caspase-3/7 assay kit.
Colon cancer cell proliferation in
fibroblast conditioned medium
Fibroblast conditioned RPMI-1640 medium, after 2
days of incubation with 1×106 WI-38, BJ and CCD-18Co
cells, was collected and centrifuged twice at 700 × g for 3 min to
remove dead cells/cellular debris. Supernatants were collected and
diluted with fresh RPMI-1640 medium and incubated with
2D/anchorage-independent colon cancer cells. Fibroblast conditioned
media were replaced every 2 days. After 4 days of colon cancer
cell/fibroblast conditioned medium co-culture, cells were
collected, and rates of proliferation and apoptosis were measured
as above.
Co-culture effect on cancer cell
proliferation following anti-cancer drug treatment
A total of 30,000 SW480, HT-29, DLD-1 and Caco-2
cells were seeded/mixed with agar in 24-well plates in the
presence/absence of WI-38, BJ or CCD-18Co fibroblasts. High
concentrations of anti-cancer drugs were used to induce strong
reduction in colon cancer cell viability and reveal the effect of
fibroblast co-culture. Specifically, 0.1–100 µM XAV939 was added on
day 2 to the co-cultured colon cancer cells. Cell culture medium
was replaced with fresh RPMI-1640 medium containing 0.1–100 µM of
XAV939 on day 4 and cell proliferation was measured on day 5. For
other anti-cancer drugs, 10 µM 5-FU, IRI, CPT or DOX was added to
the colon cancer cells on day 2, and fresh RPMI-1640 medium
containing equal concentrations of 5-FU, IRI, CPT or DOX was added
on day 4, and cell proliferation was measured on day 5. DLD-1,
HT-29 or Caco-2 cell anti-cancer drug tests were conducted as afore
described with SW480 cells.
Statistical analysis
Statistical analysis was performed using Origin
(v.8.1; OriginLab, Northampton, MA, USA) and GraphPad Prism (v.6;
GraphPad Software, Inc., La Jolla, CA, USA). Each experiment was
performed in triplicate and values were expressed as the mean ±
standard deviation. Statistical significance were examined by
analysis of variance with Tukey's post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Results
Effect of fibroblast co-culture on
colon cancer cell proliferation
To determine the influence of fibroblast co-culture
on colon cancer cell proliferation, a non-contact co-culture system
was established. Briefly, SW480, DLD-1, HT-29 or Caco-2 cells were
seeded onto a 24 well plate, and three different fibroblasts
(WI-38, BJ or CCD-18Co) were placed on inserts and co-cultured with
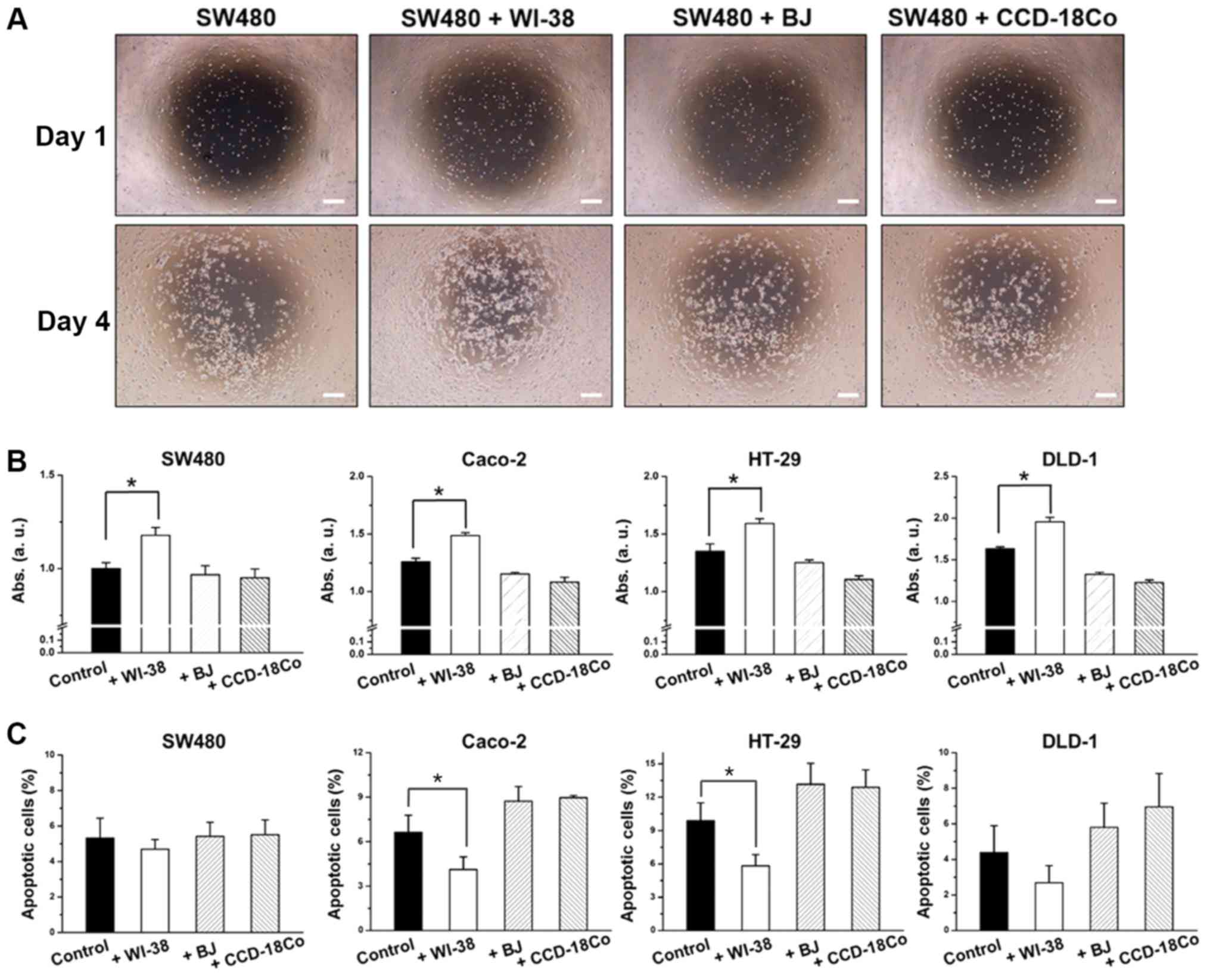
the colon cancer cells. SW480 cells exhibited a 36% increase in
proliferation rate in the presence of WI-38 cells, while showing
little or no changes in proliferation rate in the presence of BJ or
CCD-18Co cells (Fig. 1A and B). WI-38
co-culture decreased SW480 cell apoptosis by 26.3%, while BJ or
CCD-18Co co-culture had little or no effect, compared with the
controls (SW480 cells without fibroblasts; Fig. 1C). Higher proliferation rates for
SW480 cells were observed at the center of the well where
co-cultured WI-38 cells were closer (Fig.
1A). This was not observed for SW480 cells co-cultured with BJ
or CCD-18Co cells. HT-29, DLD-1 and Caco-2 cells co-cultured with
fibroblasts exhibited similar results, wherein WI-38 cells induced
increases in cell proliferation with increased cell proliferation
while BJ and CCD-18Co cells induced decreases in cell proliferation
with decreased cell proliferation (Fig.
1B and C).
Fibroblast co-culture and
anchorage-independent colon cancer cell growth
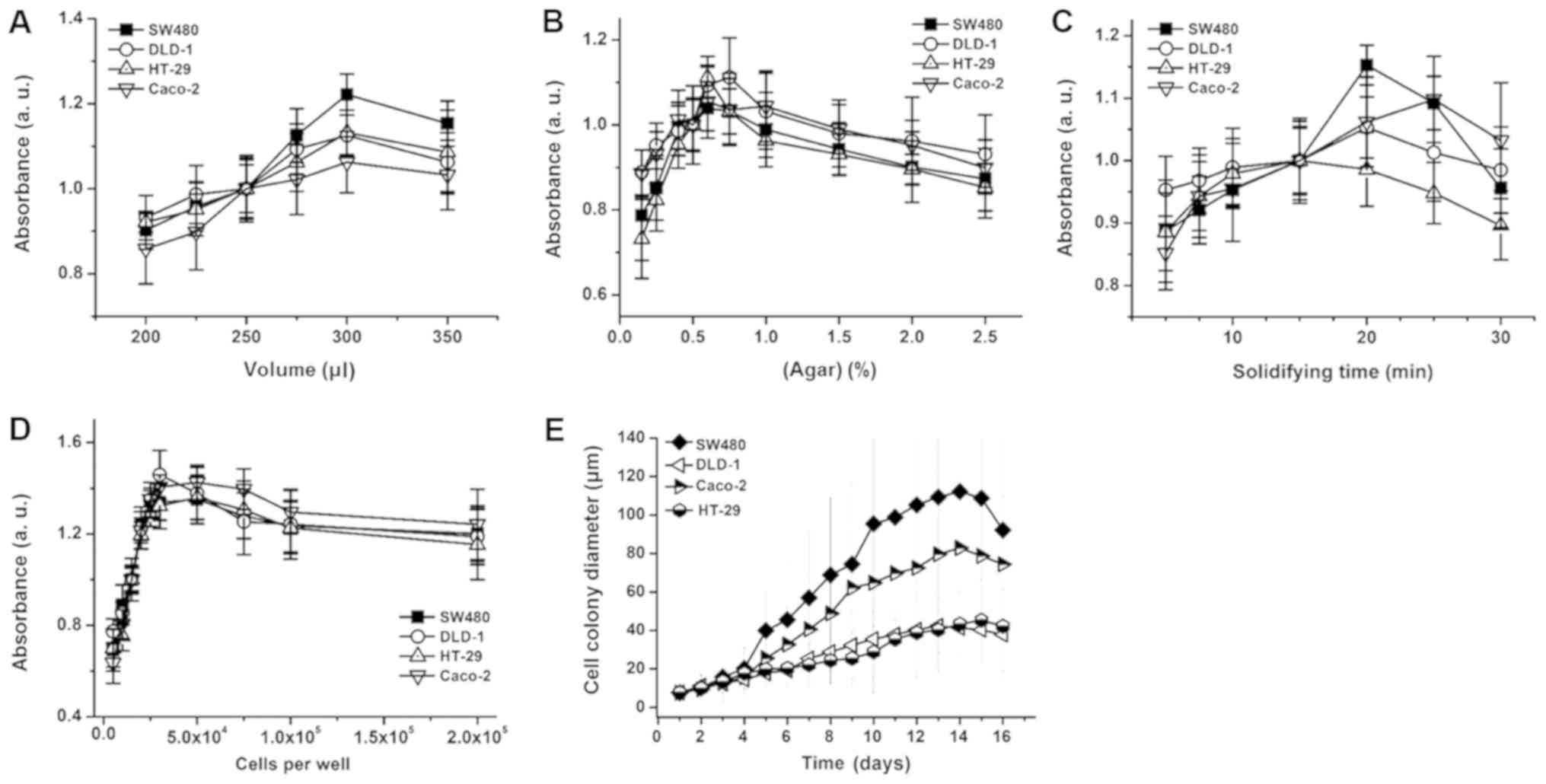
To verify the effects of fibroblasts on 3D colon
cancer cell growth, an anchorage-independent SW480 cell co-culture
was conducted (29). Prior to the
fibroblast co-culture, the agar concentration, matrix volume, agar
solidifying time and colon cancer cell seeding number was optimized
to determine proliferative anchorage-independent cell culture
conditions (Fig. 2A-D). The degree of
colony formation differed by cell type, and SW480 cells exhibited
the highest colony formation efficiency followed by Caco-2, HT-29
and DLD-1 cells (Fig. 2E). Following
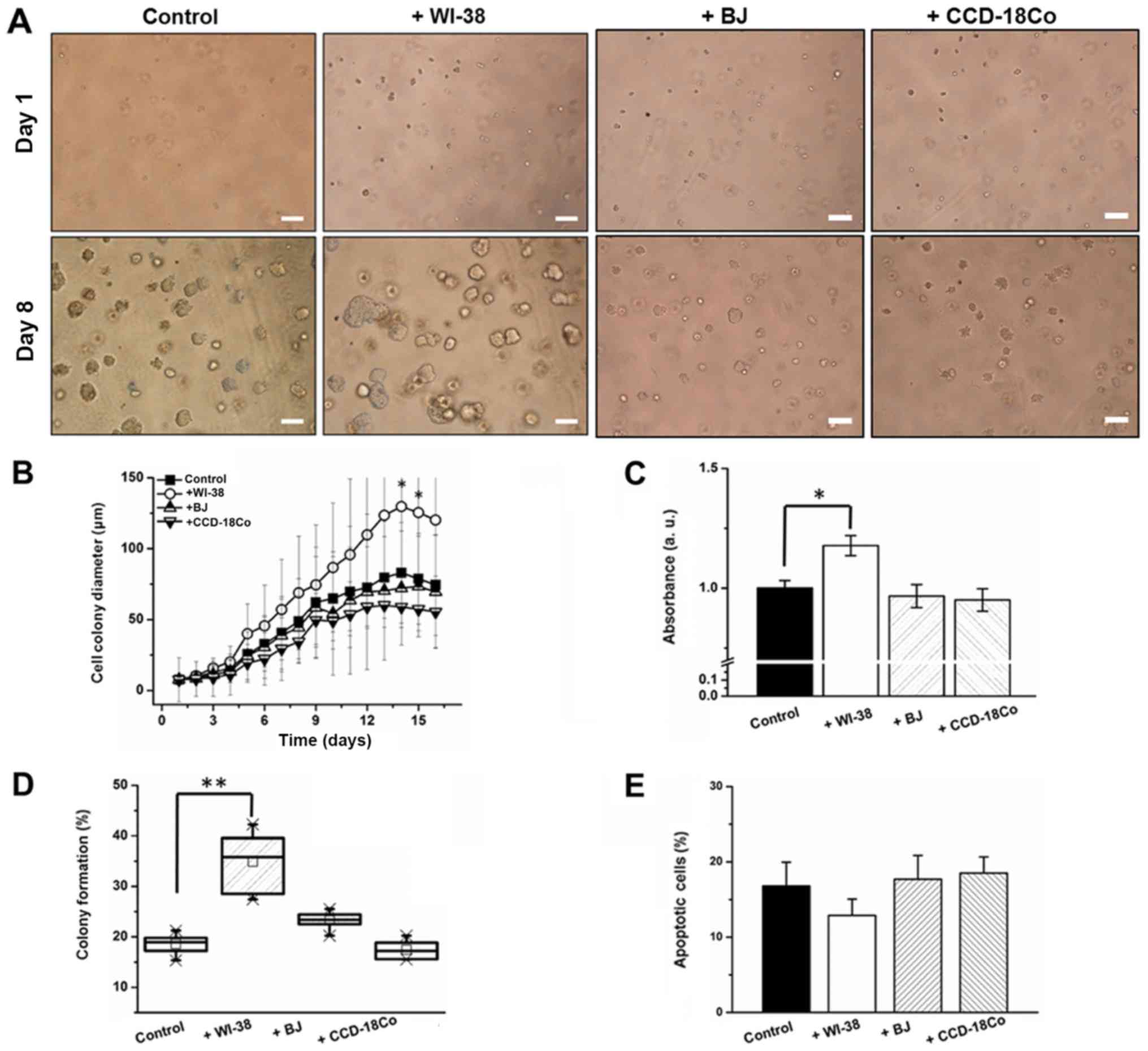
optimization, colon cancer cells in an agar layer were co-cultured
with fibroblasts, and colon cancer cell proliferation, apoptotic
and colony formation rates were monitored. Average cell colony
diameter increased by 39.4% (day 6) and 56.6% (day 14) when SW480
cells were co-cultured with WI-38 cells (Fig. 3A and B). By contrast, BJ or CCD-18Co
co-culture did not induce a significant change in cell colony
diameter (Fig. 3A and B). WI-38 cell
co-culture induced a 24% increase in SW480 cell proliferation,
while co-cultures with BJ and CCD-18Co cells decreased the
apoptotic rate of SW480 cells by 3.2 and 4.3%, respectively, by day
8 (Fig. 3C). WI-38 and BJ cell
co-cultures induced an 84 and 23.3% increase, while that with
CCD-18Co cells induced a 7.8% decrease, in the SW480 cell colony
forming rate (Fig. 3D). WI-38 cell
co-culture induced a 22.4% decrease in the apoptotic rate of
anchorage-independent SW480 cells, whereas BJ and CCD-18Co cell
co-cultures induced a 2.3% decrease and a 17% increase in the SW480
cell apoptotic rate, respectively (Fig.
3E).
Colon cancer cell growth in
fibroblast-cultured medium
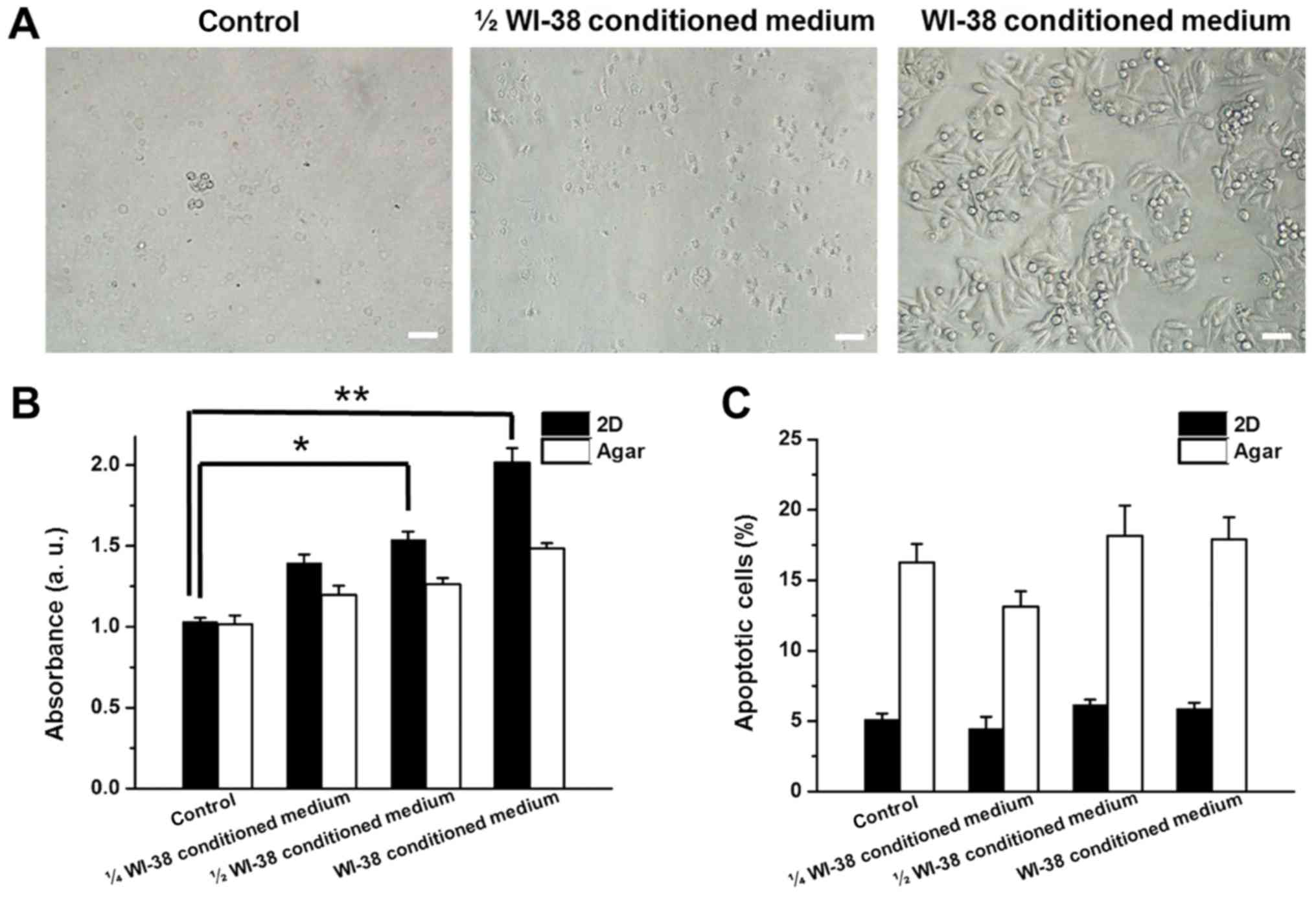
Proliferation and apoptotic rates of SW480 cells
were monitored in the presence of WI-38, BJ or CCD-18Co conditioned
medium to determine whether the co-culture effects were mediated by
factors released from the fibroblasts. WI-38, BJ and CCD-18Co
conditioned media were mixed (1:3, 1:1 ratio) or unmixed with fresh
RPMI-1640 medium and used for 2D/anchorage-independent culture of
SW480 cells. Increasing concentrations of WI-38 conditioned medium
induced higher rates of proliferation in 2D-cultured SW480 cells
(35.3, 49.1 and 95.3% increases for 1/4, 1/2 and full WI-38
conditioned medium, respectively; Fig. 4A
and B). Proliferation of anchorage-independent SW480 cells also
increased by 17.7, 24.3 and 46.2% in the presence of 1/4, 1/2 and
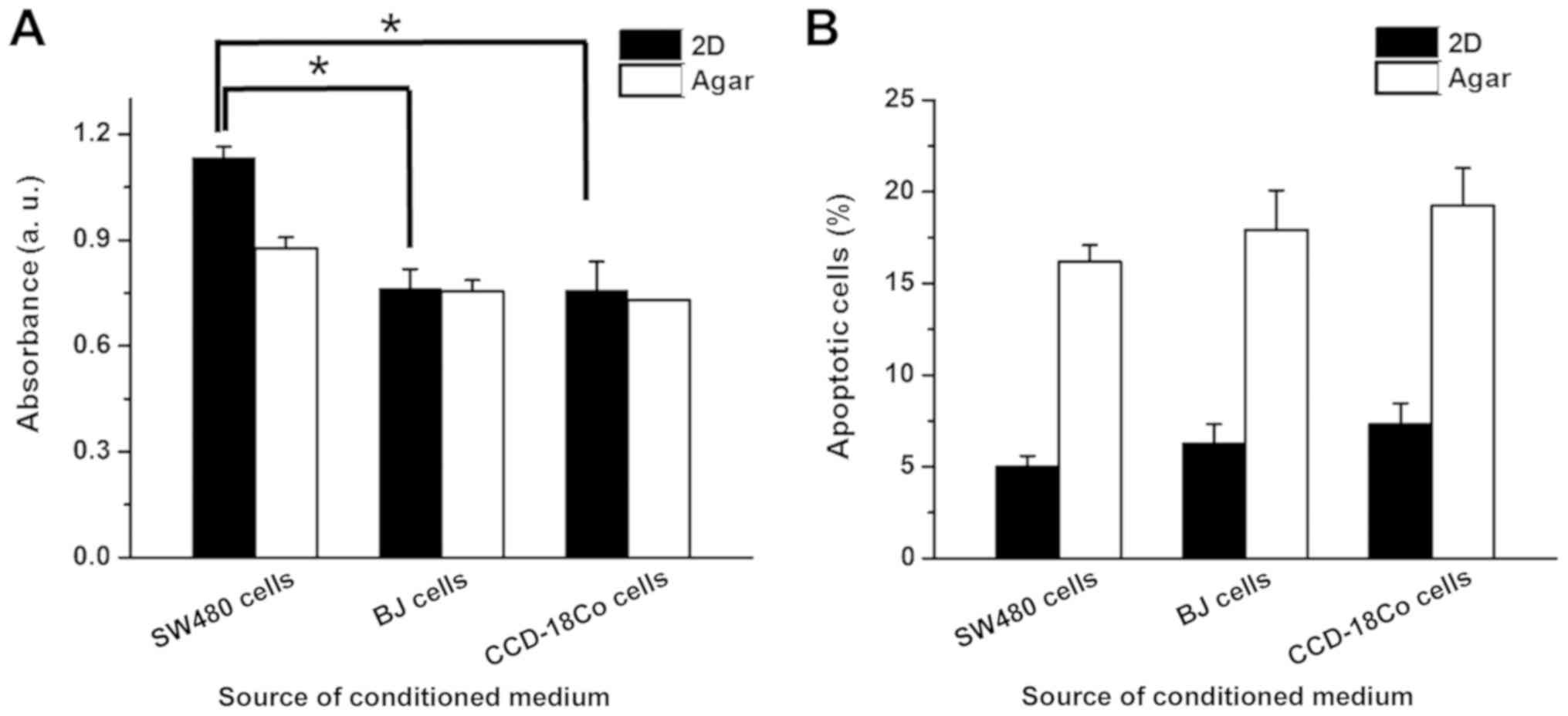
full WI-38-cultured medium, respectively (Fig. 4B). The apoptotic rates of 2D and
anchorage-independent SW480 cells changed by >5% in the presence
of WI-38 conditioned medium (Fig.
4C). BJ conditioned medium induced a decrease in 2D and
anchorage-independent cultured SW480 cell proliferation by 33.8 and
15.8%, respectively, while CCD-18Co conditioned medium reduced
proliferation in 2D and anchorage-independent SW480 cells by 34.3
and 18.4%, respectively (Fig. 5A). BJ
and CCD-18Co conditioned media did not induce >5% changes in the
apoptotic rates of 2D and anchorage-independent SW480 cells
(Fig. 5B).
Fibroblast co-culture effect on colon
cancer cell drug responses
Additionally, it was investigated whether fibroblast
co-culture could mediate changes in cancer cell responses to
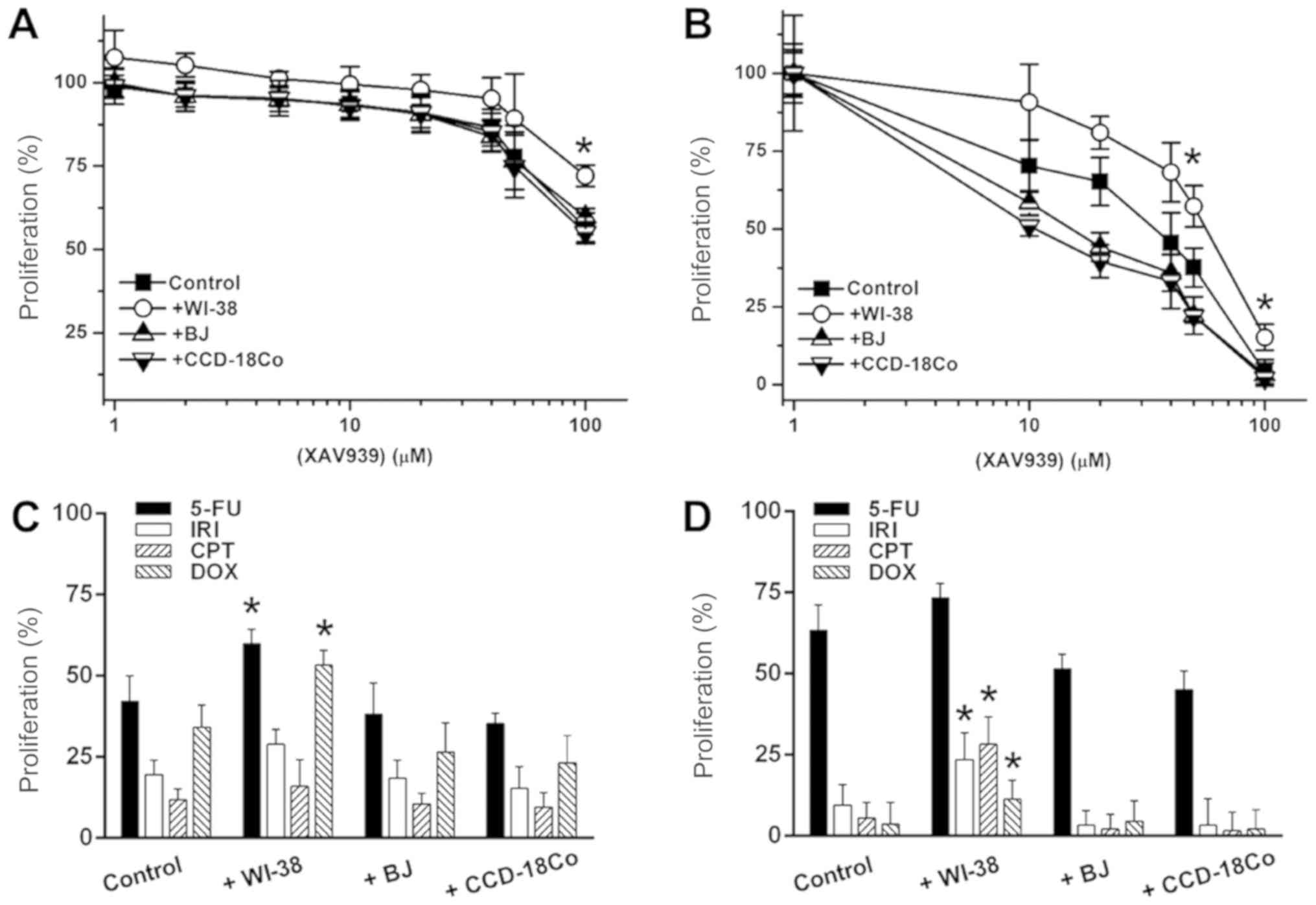
anti-cancer drugs. XAV939 exerted low anti-cancer activity against
2D-cultured SW480 cells, causing a 40.5% decrease in cell
proliferation at 100 µM (Fig. 6A). BJ
and CCD-18Co co-cultures induced 1.1 and 2.1% increases in XAV939
sensitivity, respectively (Fig. 6A).
WI-38 co-culture increased SW480 proliferation by 27.5% against 100
µM XAV939 (Fig. 6A).
Anchorage-independent SW480 cells were more sensitive to treatment
with XAV939 (Fig. 6B); at 10 µM,
XAV939 reduced the proliferation of anchorage-independent SW480
cells by 29.7%, and 100 µM XAV939 decrease SW480 proliferation by
96.1% (Fig. 6B). BJ and CCD-18Co
co-cultures enhanced the anti-cancer activity of 10 µM XAV939 by
16.8 and 27.5%, respectively. By contrast, WI-38 co-culture reduced
XAV939 activity by ~38.2% over a 10–50 µM range. Additionally, the
anti-cancer activity of 5-FU, IRI, CPT and DOX (10 µM single
concentrations) was tested against 2D/anchorage-independent SW480
cells. WI-38 co-culture reduced the anti-cancer activity of IRI by
33.8%, and of Dox by 55.7%, in 2D-cultured SW480 cells, while BJ
and CCD-18Co co-culture only exhibited <10% activity differences
for all 4 drugs (Fig. 6C). For
anchorage-independent SW480 culture, WI-38 co-culture reduced
anti-cancer activity by 17.4 (5-FU) to 48.6 (DOX)%, while BJ and
CCD-18Co co-cultures enhanced anti-cancer activity by on average
16.2 and 23.5% in anchorage-independent SW480 cells, respectively
(Fig. 6D). The effect of co-culture
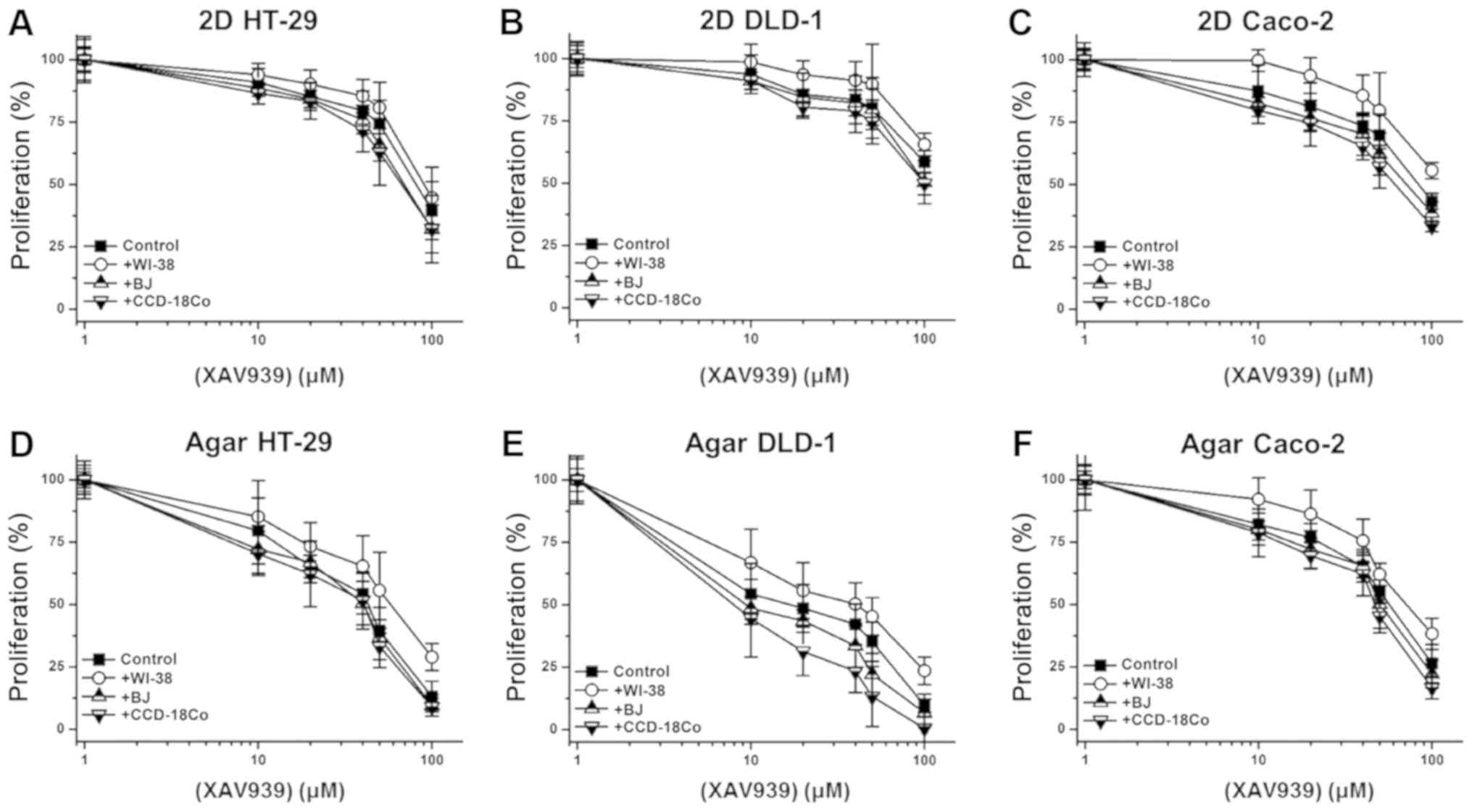
on the anti-cancer activity of XAV939 against the 3 other initial
colon cancer cell lines (HT-29, DLD-1 and Caco-2) was also tested.
Similar to its effect with SW480 cells, WI-38 co-culture reduced
the anti-cancer activity of XAV939 against HT-29, DLD-1 and Caco-2
cells, while BJ or CCD-18Co co-culture only slightly enhanced the
anti-cancer activity (Fig. 7A-F).
Discussion
Previous studies have shown that the CAF secretome
includes angiogenic factors (vascular endothelial growth factor and
fibroblast growth factor 1), growth factors [epidermal growth
factor (EGF) and transforming growth factor-β] and
matrix-modulating factors (matrix metalloproteinases and tissue
inhibitors of metalloproteinases), aiding cancer cells to survive,
proliferate and become invasive (30–32). Other
studies have suggested that normal fibroblasts serve as negative
modulators of cancer and inhibit cancer growth and metastasis
(26,33). Additionally, decreases in the
anti-cancer drug responses of cancer cells co-cultured with
fibroblasts have been observed, and factors produced by co-cultured
fibroblasts have been shown to promote tumor cell proliferation and
resistance against anti-tumor agents (34,35). These
findings warranted investigation into whether different types of
fibroblasts have different impacts on colon cancer cell
proliferation and drug response. In this study we have selected
colon fibroblasts (CCD-18Co), lung fibroblasts (WI-38) and skin
fibroblasts (BJ) for co-culture with colon cancer cells to mimic
the colon cancer microenvironment and two highly metastatic sites
of colon cancer (36,37). Colon cancer cells with low (Caco-2 and
HT-29), medium (SW480) and high (DLD-1) invasiveness (38,39) were
selected. A non-contact co-culture system was adopted using an
insert so that the cancer cell/fibroblast interactions could be
achieved via the sharing of culture medium through micropores.
Furthermore, proliferation, apoptosis for each could be
successfully determined as the two cell types were physically
separated by membranes.
The present data suggested that SW480 cells
co-cultured with WI-38 cells were significantly promoted to undergo
cell proliferation, with little or no changes in apoptotic rate,
compared with controls. To determine whether the co-culture effect
was induced by factors released from the fibroblasts, SW480 cells
were cultured in WI-38 conditioned medium and the effects on colon
cancer cell progression were monitored. SW480 cells exhibited
increased proliferation in the presence of the WI-38 conditioned
medium, as compared with the controls. This suggests that factors
in the WI-38 conditioned medium have a pro-proliferative effect on
colon cancer cells. Additionally, BJ and CCD-18Co co-culture
induced a decrease in SW480 cell proliferation, but did not induce
changes in apoptotic rate. Previous studies have demonstrated that
cancer cell proliferation is not only associated with apoptosis but
also with various other factors, and that the balance between anti-
and pro- apoptotic gene expression overall contributes to tumor
proliferation and progression (40,41). SW480
cells cultured with BJ or CCD-18Co conditioned medium also
exhibited decreased proliferation. This suggests that fibroblast
co-culture influences colon cancer cell proliferation is dependent
on cell type, and its effects on cancer cells are more complex than
simply differentiating between normal fibroblasts and CAFs. Further
tests were conducted to investigate whether fibroblast co-culture
influences anchorage-independent cultured colon cancer cells. Prior
to co-culture, the anchorage-independent culture protocol was
optimized to determine the best conditions for colon cancer cell
proliferation; this was determined to be sensitive to matrix
volume, concentration, solidifying time and the number of cells in
each matrix. Colon cancer cells formed multiple colonies after 3
days of anchorage-independent culture. Different degrees of colony
forming efficiency were observed between 4 different colon cancer
cell types; however, correlations between colon cancer cell
invasiveness and colony formation ability were not observed using
this system. Colon cancer cells that underwent
anchorage-independent culture exhibited an average 3.2-fold
increase in apoptotic rate when compared with those in 2D culture.
It is speculated that higher apoptotic rate in
anchorage-independent culture may be attributed to the hypoxic
condition of the system (42).
Following protocol optimization, colon cancer cells
in an agar matrix were co-cultured with WI-38, BJ and CCD-18Co
cells. WI-38 cells not only induced an increase in colon cancer
cell proliferation, but also induced and increase in colon cancer
cell colony size and colony forming rate. BJ and CCD-18Co
fibroblasts decreased colon cancer cell proliferation while having
little or no effect on colony forming efficiency. It was further
evaluated whether fibroblast co-culture modulated the anti-cancer
drug response of colon cancer cells. The Wnt signaling inhibitor
XAV939 was selected as a test drug to apply to colon cancer cells,
since aberrant WNT signaling is recognized as an early event in the
development of colorectal cancer (43). BJ and CCD-18Co co-cultures caused
>3% changes in the response of cancer cells to XAV939, while
WI-38 co-culture significantly reduced the XAV939-sensitivity of
colon cancer cells. 2D and anchorage independent SW480 cultures
exhibited disparate responses to XAV939. Kim et al (44) suggested that this difference can be
attributed by differential expression of lactate dehydrogenase A
and gelsolin. The effect of 5-FU, IRI, CPT and DOX on the colon
cancer-fibroblast co-culture system was also tested to investigate
the relationship between different signaling pathways and cell
proliferation in the anchorage independent condition. It has been
reported that 5-FU induces mitochondrial apoptosis in colon cancer
cells lacking p53, while IRI, CPT and DOX inhibit topoisomerase
resulting in DNA damage and apoptosis (45–47).
Against these anti-cancer agents (5-FU, IRI, CPT and DOX), WI-38
co-culture also reduced the colon cancer cell drug response. By
contrast, BJ and CCD-18Co co-cultures increased sensitivity to all
5 drugs tested, further supporting their role as negative
regulators of cancer cell proliferation. Similarly, WI-38
conditioned medium enhanced colon cancer cell proliferation while
causing little or no decrease in the rate of apoptosis, whereas
BJ/CCD-18Co medium reduced colon cancer cell proliferation.
However, the mechanism underlying the interaction between colon
cancer cells and fibroblasts are not yet fully understood, and our
group is currently investigating protein, metabolite and lipid
expression patterns associated with colon cancer growth in the
presence of fibroblasts. As p53 might play on important role in the
cancer microenvironment, we are currently investigating its role in
cancer proliferation.
In conclusion, fibroblasts may act as positive or
negative regulators of colon cancer cell proliferation depending on
the fibroblast cell type. Factors released by different types of
non-cancerous fibroblasts may have an impact on cancer cell
proliferation, apoptosis and drug responses. Thus, when targeting
TME to treat specific cancers, fibroblasts in the tissue should be
carefully examined and either positive/negative modulation of the
TME should be applied depending on the prevailing
fibroblast-mediated regulation of cancer.
Acknowledgements
The authors would like to thank Dr. Sang Dal Rhee,
Dr. Ki Young Kim and Dr. Hyejin Nam (Korea Research Institute of
Chemical Technology, Daejeon, Republic of Korea) for their helpful
discussions. The authors would also like to thank Mr. Jonghyum Kim
and Professor Tae-Young Kim (both of Gwangju Institute of Science
and Technology, Gwangju, Republic of Korea) for assisting with the
acquisition of data.
Funding
The present study was financially supported by the
R&D Convergence Program of National Research Council of Science
& Technology (grant no. CAP-15-10-KRICT), and the Korea
Research Institute of Chemical Technology core project (grant no.
KK1703-F00).
Availability of data and materials
The datasets used during the present study are
available from the corresponding author upon reasonable
request.
Authors' contributions
BK, DuK and KRK conceived and designed the study.
BK, HJ, DaK and DuK performed the experiments. BK wrote the
manuscript. BK, HJ, DaK, DuK and KRK reviewed and edited the
manuscript. All authors read and approved the manuscript and agree
to be accountable for all aspects of the research in ensuring that
the accuracy or integrity of any part of the work are appropriately
investigated and resolved. All authors have given approval to the
final version of the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interest
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jung KW, Won YJ, Kong HJ, Oh CM, Cho H,
Lee DH and Lee KH: Cancer statistics in Korea: Incidence,
mortality, survival, and prevalence in 2012. Cancer Res Treat.
47:127–141. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Wang D, Rai B, Qi F, Liu T, Wang J, Wang X
and Ma B: Influence of the Twist gene on the invasion and
metastasis of colon cancer. Oncol Rep. 39:31–44. 2018.PubMed/NCBI
|
|
4
|
Duan L, Deng L, Wang D, Ma S, Li C and
Zhao D: Treatment mechanism of matrine in combination with
irinotecan for colon cancer. Oncol Lett. 14:2300–2304. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Li K, Guo J, Wu Y, Jin D, Jiang H, Liu C
and Qin C: Suppression of YAP by DDP disrupts colon tumor
progression. Oncol Rep. 39:2114–2126. 2018.PubMed/NCBI
|
|
6
|
De Roock W, De Vriendt V, Normanno N,
Ciardiello F and Tejpar S: KRAS, BRAF, PIK3CA, and PTEN mutations:
Implications for targeted therapies in metastatic colorectal
cancer. Lancet Oncol. 12:594–603. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Moriarity A, O'Sullivan J, Kennedy J,
Mehigan B and McCormick P: Current targeted therapies in the
treatment of advanced colorectal cancer: A review. Ther Adv Med
Oncol. 8:276–293. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hagan S, Orr MC and Doyle B: Targeted
therapies in colorectal cancer-an integrative view by PPPM. EPMA J.
4:32013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Heinemann V, Douillard JY, Ducreux M and
Peeters M: Targeted therapy in metastatic colorectal cancer-an
example of personalized medicine in action. Cancer Treat Rev.
39:592–601. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hwang TJ, Carpenter D, Lauffenburger JC,
Wang B, Franklin JM and Kesselheim AS: Failure of investigational
drugs in late-stage clinical development and publication of trial
results. JAMA Intern Med. 176:1826–1833. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hay M, Thomas DW, Craighead JL, Economides
C and Rosenthal J: Clinical development success rates for
investigational drugs. Nat Biotechnol. 32:40–51. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun Y: Tumor microenvironment and cancer
therapy resistance. Cancer Lett. 380:205–215. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fang H and Declerck YA: Targeting the
tumor microenvironment: From understanding pathways to effective
clinical trials. Cancer Res. 73:4965–4977. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chen F, Zhuang X, Lin L, Yu P, Wang Y, Shi
Y, Hu G and Sun Y: New horizons in tumor microenvironment biology:
Challenges and opportunities. BMC Med. 13:452015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Peddareddigari VG, Wang D and Dubois RN:
The Tumor microenvironment in colorectal carcinogenesis. Cancer
Microenviron. 3:149–166. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhou W, Xu G, Wang Y, Xu Z, Liu X, Xu X,
Ren G and Tian K: Oxidative stress induced autophagy in cancer
associated fibroblast enhances proliferation and metabolism of
colorectal cancer cells. Cell Cycle. 16:73–81. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
O'Toole A, Michielsen AJ, Nolan B, Tosetto
M, Sheahan K, Mulcahy HE, Winter DC, Hyland JM, O'Connell PR,
Fennelly D, et al: Tumour microenvironment of both early- and
late-stage colorectal cancer is equally immunosuppressive. Br J
Cancer. 111:927–932. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jacobs J, Smits E, Lardon F, Pauwels P and
Deschoolmeester V: Immune checkpoint modulation in colorectal
cancer: What's new and what to expect. J Immunol Res.
2015:1580382015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Colangelo T, Polcaro G, Muccillo L,
D'Agostino G, Rosato V, Ziccardi P, Lupo A, Mazzoccoli G, Sabatino
L and Colantuoni V: Friend or foe?: The tumour microenvironment
dilemma in colorectal cancer. Biochim Biophys Acta Rev Cancer.
1867:1–18. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Cammarota R, Bertolini V, Pennesi G, Bucci
EO, Gottardi O, Garlanda C, Laghi L, Barberis MC, Sessa F, Noonan
DM and Albini A: The tumor microenvironment of colorectal cancer:
Stromal TLR-4 expression as a potential prognostic marker. J Transl
Med. 8:1122010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Marsh T, Pietras K and McAllister SS:
Fibroblasts as architects of cancer pathogenesis. Biochim Biophys
Acta. 1832:1070–1078. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kalluri R: The biology and function of
fibroblasts in cancer. Nat Rev Cancer. 16:582–598. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kalluri R and Zeisberg M: Fibroblasts in
cancer. Nat Rev Cancer. 6:392–401. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lotti F, Jarrar AM, Pai RK, Hitomi M,
Lathia J, Mace A, Gantt GA Jr, Sukhdeo K, DeVecchio J, Vasanji A,
et al: Chemotherapy activates cancer-associated fibroblasts to
maintain colorectal cancer-initiating cells by IL-17A. J Exp Med.
210:2851–2872. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ostman A and Augsten M: Cancer-associated
fibroblasts and tumor growth-bystanders turning into key players.
Curr Opin Genet Dev. 19:67–73. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Iacopino F, Angelucci C and Sica G:
Interactions between normal human fibroblasts and human prostate
cancer cells in a co-culture system. Anticancer Res. 32:1579–1588.
2012.PubMed/NCBI
|
|
27
|
Tao L, Huang G, Song H, Chen Y and Chen L:
Cancer associated fibroblasts: An essential role in the tumor
microenvironment. Oncol Lett. 14:2611–2620. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shiga K, Hara M, Nagasaki T, Sato T,
Takahashi H and Takeyama H: Cancer-associated fibroblasts: Their
characteristics and their roles in tumor growth. Cancers (Basel).
7:2443–2458. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zabransky DJ, Yankasakas CL, Cochran RL,
Wong HY, Croessmann S, Chu D, Kavuri SM, Red Brewer M, Rosen DM,
Dalton WB, et al: HER2 missense mutations have distinct effects on
oncogenic signaling and migration. Proc Natl Acad Sci USA.
112:E6205–E6214. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hwang RF, Moore T, Arumugam T,
Ramachandran V, Amos KD, Rivera A, Ji B, Evans DB and Logsdon CD:
Cancer-associated stromal fibroblasts promote pancreatic tumor
progression. Cancer Res. 68:918–926. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liao D, Luo Y, Markowitz D, Xiang R and
Reisfeld RA: Cancer associated fibroblasts promote tumor growth and
metastasis by modulating the tumor immune microenvironment in a 4T1
murine breast cancer model. PLoS One. 4:e79652009. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cheng HH, Chu LY, Chiang LY, Chen HL, Kuo
CC and Wu KK: Inhibition of cancer cell epithelial mesenchymal
transition by normal fibroblasts via production of
5-methoxytryptophan. Oncotarget. 7:31243–31256. 2016.PubMed/NCBI
|
|
33
|
de Toledo M, Anguille C, Roger L, Roux P
and Gadea G: Cooperative anti-invasive effect of Cdc42/Rac1
activation and ROCK inhibition in SW620 colorectal cancer cells
with elevated blebbing activity. PLoS One. 7:e483442012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Majety M, Pradel LP, Gies M and Ries CH:
Fibroblast influence survival and therapeutic response in a 3d
Co-culture model. PLoS One. 10:e01279482015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yamada T, Matsumoto K, Wang W, Li Q,
Nishioka Y, Sekido Y, Sone S and Yano S: Hepatocyte growth factor
reduces susceptibility to an irreversible epidermal growth factor
receptor inhibitor in EGFR-T790M mutant lung cancer. Clin Cancer
Res. 16:174–183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Villeneueve PJ and Sundaresan RS: Surgical
management of colorectal lung metastasis. Clin Colon Rectal Surg.
22:233–241. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Nesseris I, Tsamakis C, Gregoriou S,
Ditsos I, Christofidou E and Rigopoulos D: Cutaneous metastasis of
colon adenocarcinoma: Case report and review of the literature. An
Bras Dermatol. 88:(6 Suppl 1). S56–S58. 2013. View Article : Google Scholar
|
|
38
|
Liu Y, Zhang F, Zhang XF, Qi LS, Yang L,
Guo H and Zhang N: Expression of nucleophosmin/NPM1 correlates with
migration and invasiveness of colon cancer cells. J Biomed Sci.
19:532012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mouradov D, Sloggett C, Jorissen RN, Love
CG, Li S, Burgess AW, Arango D, Strausberg RL, Buchanan D, Wormald
S, et al: Colorectal cancer cell lines are representative models of
the main molecular subtypes of primary cancer. Cancer Res.
74:3238–3247. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Labi V and Erlacher M: How cell death
shapes cancer. Cell Death Dis. 6:e16752015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wong RS: Apoptosis in cancer: From
pathogenesis to treatment. J Exp Clin Cancer Res. 30:872011.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Trédan O, Galmarini CM, Patel K and
Tannock IF: Drug Resistance and the solid tumor microenvironment. J
Natl Cancer Inst. 99:1441–1454. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Segditsas S and Tomlinson I: Colorectal
cancer and genetic alterations in the Wnt pathway. Oncogene.
25:7531–7537. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Kim YE, Jeon HJ, Kim D, Lee SY, Kim KY,
Hong J, Maeng PJ, Kim KR and Kang D: Quantitative proteomic
analysis of 2D and 3D cultured colorectal cancer cells: Profiling
of tankyrase inhibitor xav939-induced proteome. Sci Rep.
8:132552018. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Pagliara V, Saide A, Mitidieri E,
d'Emmanuele di Villa Bianca R, Sorrentino R, Russo G and Russo A:
5-FU targets rpL3 to induce mitochondrial apoptosis via
cystathionine-β-synthase in colon cancer cells lacking p53.
Oncotarget. 7:50333–50348. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Fuchs C, Mitchell EP and Hoff PM:
Irinotecan in the treatment of colorectal cancer. Cancer Treat Rev.
32:491–503. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Weinländer G, Kornek G, Raderer M, Hejna
M, Tetzner C and Scheithauer W: Treatment of advanced colorectal
cancer with doxorubicin combined with two potential
multidrug-resistance-reversing agents: High-dose oral tamoxifen and
dexverapamil. J Cancer Res Clin Oncol. 123:452–455. 1997.
View Article : Google Scholar : PubMed/NCBI
|