Introduction
Oral squamous cell carcinoma (OSCC) is the most
frequent malignant tumor occurring in the oral-maxillofacial
region; OSCC ranks sixth in incidence among all types of tumors
worldwide and represents nearly 3% of all newly diagnosed cancer
cases (1). Although the diagnostic
and therapeutic methods have been improved over the past two
decades, the survival rate of patients with OSCC has not
significantly increased, and the 5-year survival rate remains at
<50% (2). As reported previously,
OSCC is highly malignant, with strong local infiltration and
metastasis, and its inevitable relapse and metastasis constitute
the principal cause of mortality for patients with OSCC (3,4).
Therefore, investigating the molecular mechanism of OSCC metastasis
contributes to identifying an optimal therapeutic target to improve
the prognosis of patients with OSCC.
Long non-coding RNAs (lncRNAs) are a class of
functional RNA molecules with transcripts >200 nucleotides,
which cannot be translated into proteins (5). In recent years, lncRNAs have been
identified to be closely associated with tumor progression by
affecting the growth, apoptosis, infiltration and metastasis of
tumor cells (6). Meanwhile,
epithelial-mesenchymal transition (EMT), one of the effective
processes that endow tumor cells with the ability for invasion and
metastasis, serves an important role in the invasion and metastasis
of epithelium-originated tumors (7).
It has been further confirmed that the presence of EMT markers in
OSCC cells predicts high malignancy and a poor prognosis in
patients with OSCC (8). Accumulating
evidence has suggested that numerous lncRNAs in OSCC, including
urothelial cancer-associated 1 (9),
gastric carcinoma proliferation-enhancing transcript 1 (10) and metastasis-associated lung
adenocarcinoma transcript 1 (MALAT1) (11), may regulate the invasion and migration
of tumor cells and affect the disease progression through mediating
EMT.
Colorectal neoplasia differentially expressed
(CRNDE), which is located at 16q12.2 on the long arm of human
chromosome sixteen, was previously known as LOC388279 or LOC643911
(12). As demonstrated by the
published literature, CRNDE exerts significant effects on the
proliferation, invasion and migration of various types of malignant
tumors, including colorectal cancer (13), ovarian cancer (14) and glioma (15). For instance, the study by Wang et
al (16) detected the
overexpression of CRNDE in glioma, which could facilitate the
growth and migration of glioma cells in vivo and in
vitro. Additionally, Chen et al (17) identified that CRNDE was also
increased, upregulating the expression of nuclear factor-κB and
p-protein kinase B (AKT) via the negative modulation of microRNA
(miR)-384, thereby promoting hepatic carcinoma cell proliferation,
migration and invasion (17).
However, there was no evidence clearly demonstrating whether CRNDE
influences the invasion and migration of OSCC cells through the
regulation of the EMT process. Therefore, the present study was
conducted to provide a novel perspective regarding the targeted
treatment of OSCC in the hope of preventing recurrence and
metastasis, and improving the prognosis of patients with OSCC.
Materials and methods
Ethics statement
The present study was conducted in accordance with
the protocols in the Helsinki Declaration (18), and was approved by Clinical Trial
Ethics Committee of Jingzhou Central Hospital (Jingzhou, China).
All patients involved in the present study were informed of the
experiments and provided written informed consent. The animal
experiments were approved by the Ethics Committee of Jingzhou
Central Hospital, The Second Clinical Medical College, Yangtze
University (Jingzhou, China).
OSCC patients and experimental cell
lines
Between April 2012 and October 2013, OSCC specimens
were collected from 52 patients (including 35 males and 17 females,
aged between 32 and 65 years with a mean age of 58.6±9.1 years) who
received surgical excision in the Department of Stomatology at
Jingzhou Central Hospital. Normal oral mucosa tissue specimens from
25 healthy individuals (including 16 males and 9 females, aged
between 28 and 62 years with a mean age of 57.8±8.9 years) were
obtained as the control group. None of the patients had received
chemotherapy or radiation therapy prior to surgery. The tumor and
normal tissues were examined and confirmed by three pathologists,
and all the specimens were then preserved at −80°C until subsequent
experimentation. The immortalized human oral keratinocyte (HOK)
cell line (catalog no. BNCC340217) and the OSCC cell lines,
Tca8113, SCC-9, TSCCA, CAL-27 and SCC-15, used in the present study
were purchased from the Shanghai Cell Bank of the Chinese Academy
of Sciences (Shanghai, China). The cells were cultured in minimal
essential medium (MEM; Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) supplemented with 10% fetal bovine serum (FBS;
Hyclone; GE Healthcare Life Sciences, Logan, UT, USA), 100 U/ml
penicillin and 50 µg/ml streptomycin, and were incubated with 5%
CO2 at 37°C. When cell confluence reached 80–90%, the
cells were digested with 0.25% trypsin prior to passaging.
In situ hybridization (ISH)
ISH was performed on the basis of the manufacturer's
protocol of a commercial ISH Detection kit (catalogue no. AR0149;
Wuhan Boster Bio Technology, Ltd., Wuhan, China, http://www.boster.com.cn/product/ish_ar0149.html).
In brief, the 4-µm thick paraffin-embedded sections were
deparaffinized with xylene and rehydrated with 100, 90, 70 and 50%
ethanol (5 min each) at room temperature. The samples were digested
with proteinase K and fixed in 4% paraformaldehyde for 10 min at
room temperature, followed by hybridization with a
5′-digoxin-labeled CRNDE probe (Wuhan Boster Bio Technology, Ltd.),
which had the sequence 5′-CCTCAGTTGTCACGCAG-AAG-3′, at 55°C
overnight and subsequent incubation with a horseradish peroxidase
(HRP)-conjugated anti-mouse IgG antibody (1:5,000; part of the ISH
Detection kit) for 30 min at 4°C. Diaminobenzidine was used to
develop the stain with a colorimetric reaction for 30 min at room
temperature. The ISH-stained tissue sections were independently
reviewed and scored by two pathologists in a blinded manner.
Disagreements were resolved by a third pathologist. A simple and
reproducible scoring system based on the intensity and proportion
of CRNDE-positive cells was used according to a previous study
(19).
Cell grouping and transfection
CAL-27 and SCC-15 cells were seeded at a density of
105 cells per well on a 6-well plate 24 h prior to
transfection. Cells were assigned into three groups: The control
group (non-transfected cells), the si-NC group (cells transfected
with control siRNA) and the si-CRNDE group (cells transfected with
CRNDE siRNA). The CRNDE siRNA and control siRNA used in the present
study were provided by Shanghai GenePharma Co., Ltd. (Shanghai,
China). The sequences were as follows: CRNDE siRNA forward,
5′-GUGCUCGAGUGGUUUAAAUTT-3′ and reverse,
5′-AUUUAAACCACUCGAGCACTT-3′; and control siRNA forward,
5′-GCGACGAUCUGCCUAAGAUTT-3′ and reverse,
5′-AUCUUAGGCAGAUCGUCGCTT-3′. The transfection of cells was
performed with Lipofectamine™ 3000 (Thermo Fisher Scientific, Inc.)
for a final concentration of 50 nM siRNA/well, according to the
manufacturer's protocol. Cells were harvested 48 h
post-transfection for further analyses.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from OSCC tumor and normal
tissues, and CAL-27 and SCC-15 cells with a TRIzol reagent kit
(Invitrogen; Thermo Fisher Scientific, Inc.), the optical density
(OD)260/280 ratio was determined with an ultraviolet
spectrophotometer, and the RNA concentration was then calculated.
The RNA specimens were preserved at −80°C for later experiments.
The primer design software Primer 5.0 (Premier Biocoft
International, Palo Alto, CA, USA) was used to design the following
primers: LncRNA CRNDE forward, 5′-CGCGCCCGCGCGGCGGAGGA-3′ and
reverse, 5′-TATGAATTGCAGACTTTGCAGA-3′; and GAPDH forward,
5′-GTCAGCCGCATCTTCTTTTG-3′ and reverse, 5′ GCGCCCAATACGACCAAATC-3′.
RT-PCR was performed on total RNA according to the protocols of the
cDNA transcription kit (Thermo Fisher Scientific, Inc.), and
RT-qPCR was performed following the steps for the SYBR Green PCR
Master Mix kit (Takara Bio, Inc., Otsu, Japan). PCR conditions
included an initial step of 95°C for 10 min, followed by 40 cycles
of 95°C for 15 sec, 55°C for 30 sec and 72°C for 30 sec. The PCR
results were obtained and the relative expression of the target
genes was calculated by the 2−ΔΔCq method (20), with GAPDH as the internal
reference.
Western blotting
The concentrations of proteins extracted from the
CAL-27 and SCC-15 cells were determined with a BCA kit (Wuhan
Boster Biological Technology, Ltd.). Subsequent to the addition of
SDS gel loading buffer (Wuhan Boster Biological Technology, Ltd.),
the proteins were heated for 10 min at 95°C, and 30 µg protein was
loaded into each well of a 10% polyacrylamide gel (Wuhan Boster
Biological Technology, Ltd.). Next, electrophoresis was used for
the separation of proteins, which were transferred to a
polyvinylidene fluoride (PVDF) membrane. The membrane was later
placed in 5% bovine serum albumin (BSA; Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) for 1 h at room temperature, followed by the
addition of primary antibodies overnight at 4°C. The primary
antibodies included GAPDH (1:500 dilution; catalog no. 5174; Cell
Signaling Technology, Inc., Danvers, MA, USA), E-cadherin (1:1,000
dilution; catalog no. 3195; Cell Signaling Technology, Inc.),
glycogen synthase kinase 3β (GSK-3β; 1:1,000 dilution; catalog no.
ab93926; Abcam, Cambridge, UK), phosphorylated (p-)GSK-3β (1:1,000
dilution; catalog no. ab131097; Abcam), β-catenin (1:1,000
dilution; catalog no. ab16051; Abcam), vimentin (1:1,000 dilution;
catalog no. SC-6260; Santa Cruz Biotechnology, Inc., Dallas, TX,
USA), N-cadherin (1:1,000 dilution; catalog no. SC-393933; Santa
Cruz Biotechnology, Inc.) and Snail (1:1,000 dilution; catalog no.
SC-393172; Santa Cruz Biotechnology, Inc.). The next day, the
membrane was rinsed with Tris-buffered saline with Tween-20 (TBST)
three times for 5 min each prior to the addition of the
HRP-conjugated anti-mouse IgG secondary antibody (1:2,000; catalog
no. HS201-01; Beijing Transgen Biotech Co., Ltd., Beijing, China)
for 1 h of incubation at room temperature. Following incubation
with the secondary antibody, the membrane was washed again with
TBST three times for 5 min each. Finally, a chemiluminescence
reagent was used for detection and visualization, and the gray
values of the target bands were analyzed with ImageJ software
version 1.43 (National Institutes of Health, Bethesda, MD, USA),
with GAPDH as the loading control.
Immunofluorescence staining
Briefly, CAL-27 and SCC-15 cells cultured on cover
slips were washed twice with cold PBS and fixed with cold
methanol/acetone (1:1) for 10 min at −20°C. Following washing with
PBS three times, the cells were blocked for 40 min at room
temperature with 0.1% Triton X-100 and 2% normal donkey serum
(Chemicon International; Thermo Fisher Scientific, Inc.) in PBS
buffer, and incubated with the aforementioned specific primary
antibodies, and were then stained at 37°C for 10 min with
fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse
secondary antibody (1:500; catalog no. FI-4000; Vector
Laboratories, Inc., Burlingame, CA, USA) secondary antibody. To
visualize the nuclei, the cells were double-stained with
4,6-diamidino-2-phenylindole for 10 min at 37°C and viewed with a
Nikon Eclipse 80i epifluorescence microscope, which was equipped
with a digital camera (DS-Ri1, Nikon Corporation, Toyko, Japan).
All immunofluorescence images were obtained with identical exposure
settings.
Cell proliferation measurement by MTT
assay
An MTT assay was used to detect cell proliferation.
First, CAL-27 and SCC-15 cells in the logarithmic growth phase were
obtained and seeded into 96-well plates at a density of 3,000
cells/well. Four replicate wells were established for each group.
Subsequently, adherent cells were spread on the bottom and walls of
the plate, MTT was detected at 0, 24, 48 and 72 h post-inoculation
by adding 20 µl of 5 mg/ml MTT solution to each well and incubating
the plate for 4 h at 37°C. The supernatant was discarded, 200 µl
dimethyl sulfoxide (DMSO) was added, and, after 10 min of
oscillation, the absorbance value (OD value) at a wavelength of 570
nm was detected by a microplate reader.
Cell apoptosis determination by flow
cytometry
The rate of cell apoptosis was detected by an
Annexin V kit (BD Biosciences, Franklin Lakes, NJ, USA). CAL-27 and
SCC-15 cells were digested with an appropriate amount of trypsin to
create a single cell suspension, which was centrifuged at 500 × g
for 5 min at 37°C. Next, cells were washed with PBS three times for
5 min each. The supernatant was removed, 100 µl of buffer (provided
in the kit) was added to each Eppendorf tube and 5 µl of propidium
iodide (PI) and FITC was added to the tubes, which were then held
at room temperature for 5 min, followed by the addition of 400 µl
buffer in an ice bath for 30 min and the determination of cell
apoptosis by flow cytometry (FACSCanto II, BD Biosciences, San
Jose, CA, USA). The data were analyzed by CELL Quest 3.0 software
(BD Biosciences).
Cell migration examination by
wound-healing assay
CAL-27 and SCC-15 cells (1×106
cells/well) were seeded in 6-well plates, and a vertical line was
drawn across the middle of the plate with the head of a 200-µl
transfer liquid gun. Cells were subsequently washed with PBS buffer
three times, followed by the addition of 200 µl serum-free MEM. At
0 and 48 h post-incubation at 37°C, the plate was placed under an
inverted microscope for observation and imaging. Cells were counted
in five random fields (magnification, ×200). The distances between
the edges of the scratches were measured by Image-Pro Plus 6.0
software (National Institutes of Health).
Cell invasion detection by Transwell
assay
CAL-27 and SCC-15 cells obtained in the logarithmic
growth phase were subjected to routine digestion and were adjusted
to a cell density of 1×105 cells/ml with serum-free MEM.
Next, the cells were inoculated into the upper chamber of a
Transwell chamber with 100 µl/well Matrigel. After 24 h, the
remaining cells and Matrigel on the upper chamber were removed with
a cotton swab. Next, the cells that had migrated to the lower
chamber, which contained 600 µl Dulbecco's modified Eagle's medium
(Gibco; Thermo Fisher Scientific, Inc.) with 10% FBS, were fixed
for 20 min with 4% paraformaldehyde at 37°C, washed with DPBS 3
times for 2 min each, stained with 0.1% crystal violet for 3–5 min
at 37°C, and rinsed with DPBS 3 times for 2 min each. The
polycarbonate membrane was sliced into pieces and fixed to a slide
with neutral resin, which was inverted under a microscope (×200
magnification) for the observation of 3 randomly selected fields of
view. The number of cells penetrating the membrane was counted
prior to and following transfection in each group.
In vitro generation of xenograft
tumors in nude mice
For this experiment, 15 BALB/c-nu (specific
pathogen-free) male mice, aged 4–6 weeks and weighing 15–18 g, were
purchased from the Institute of Laboratory Animal Science, Chinese
Academy of Medical Sciences and Peking Union Medical College
(Beijing, China). CAL-27 cells were transfected with short hairpin
CRNDE (sh-CRNDE) plasmid or with empty vector according to the
previously outlined transfection protocol and were harvested from
6-well plates, washed with PBS and resuspended at 2×107
cells/ml. The nude mice were divided into three groups (n=5 per
group), namely the control group (implanted with CAL-27 cells
only), the empty vector group (implanted with CAL-27 cells
transfected with empty vector) and the sh-CRNDE group (implanted
with CAL-27 cells transfected with sh-CRNDE plasmid). Subsequently,
100 µl of suspended cells were injected into the right flank of
each mouse. Every 3 days, the length (a) and width (b) of the
tumors were measured by Vernier calipers, and the tumors were
observed for 6 weeks continuously to allow a tumor growth curve to
be drawn. The volume of the tumors was calculated according to the
following formula: V=(ab2)/2. Following the experiment,
the animals were sacrificed and the tumor-bearing specimens were
removed for weighing, formalin fixation, paraffin embedding and
sectioning.
Detection of EMT-related proteins in
tumor tissues of nude mice by immunohistochemistry
The avidin-biotin-peroxidase complex
immunohistochemical method was utilized to evaluate the expression
of EMT-related proteins in tumor tissues of nude mice. First,
paraffin-embedded sections were dewaxed with 100, 90, 70, and 50%
alcohol solutions (5 min each at 37°C), followed by heat-induced
repair in 0.01 mol/l citrate buffer (pH 6.0), 20 min of endogenous
peroxidase inhibition with 0.3% hydrogen peroxide, 30 min of
incubation at room temperature in 20% normal goat serum (Vector
Laboratories, Inc.) and overnight incubation at 4°C with the
aforementioned primary antibodies, including E-cadherin, vimentin,
N-cadherin and Snail. The sections were then incubated for an
additional 1 h at 37°C, washed with 0.01 mol/l PBS and incubated
for 20 min at 37°C with anti-mouse HRP-conjugated IgG secondary
antibody (1:2,000; catalog no. HS201-01; Transgen Biotech Co.,
Ltd.). After development with 3,3′-diaminobenzidine reagent for 5
min at room temperature, sections were observed for staining under
a light microscope. Finally, hematoxylin was used for 30 sec of
counterstaining; sections were then rinsed with running water for 5
min, hyalinized and mounted with neutral resin prior to observation
under a light microscope (magnification, ×400).
Statistical method
All data were analyzed with the statistical software
package SPSS 21.0 (IBM Corp., Armonk, NY, USA). The results are
presented as the mean ± standard deviation. Comparisons between two
groups of data conforming to a normal distribution were conducted
using unpaired Student's t-test, while the difference among
multiple groups was analyzed by one-way analysis of variance
followed by Tukey's post-hoc test. P<0.05 was considered to
indicate a statistically significant difference.
Results
Expression of CRNDE in human OSCC
tissues and cell lines
The expression of CRNDE in human OSCC tissues and
cell lines was detected by RT-qPCR. As illustrated in Fig. 1A, compared with normal oral mucosa
tissues, human OSCC tissues exhibited a significant increase in
CRNDE expression (P<0.05). Compared with immortalized HOK cells,
the human OSCC cell lines, Tca8113, SCC-9, TSCCA, CAL-27 and
SCC-15, displayed a statistically significant increase in the
expression of CRNDE (all P<0.05; Fig.
1B). Among the OSCC cell lines, CAL-27 and SCC-15 presented
with the most significant upregulation of CRNDE (P<0.05),
therefore, these two cell lines were selected for subsequent
experiments. In addition, the present study confirmed that CRNDE
expression was markedly upregulated in OSCC tissues by in
situ hybridization (Fig. 1C);
CRNDE was identified to be primarily expressed in the cytoplasm,
with expression in 80.77% (42/52) of the OSCC samples, but in only
16.00% (4/25) of the normal oral mucosa tissues.
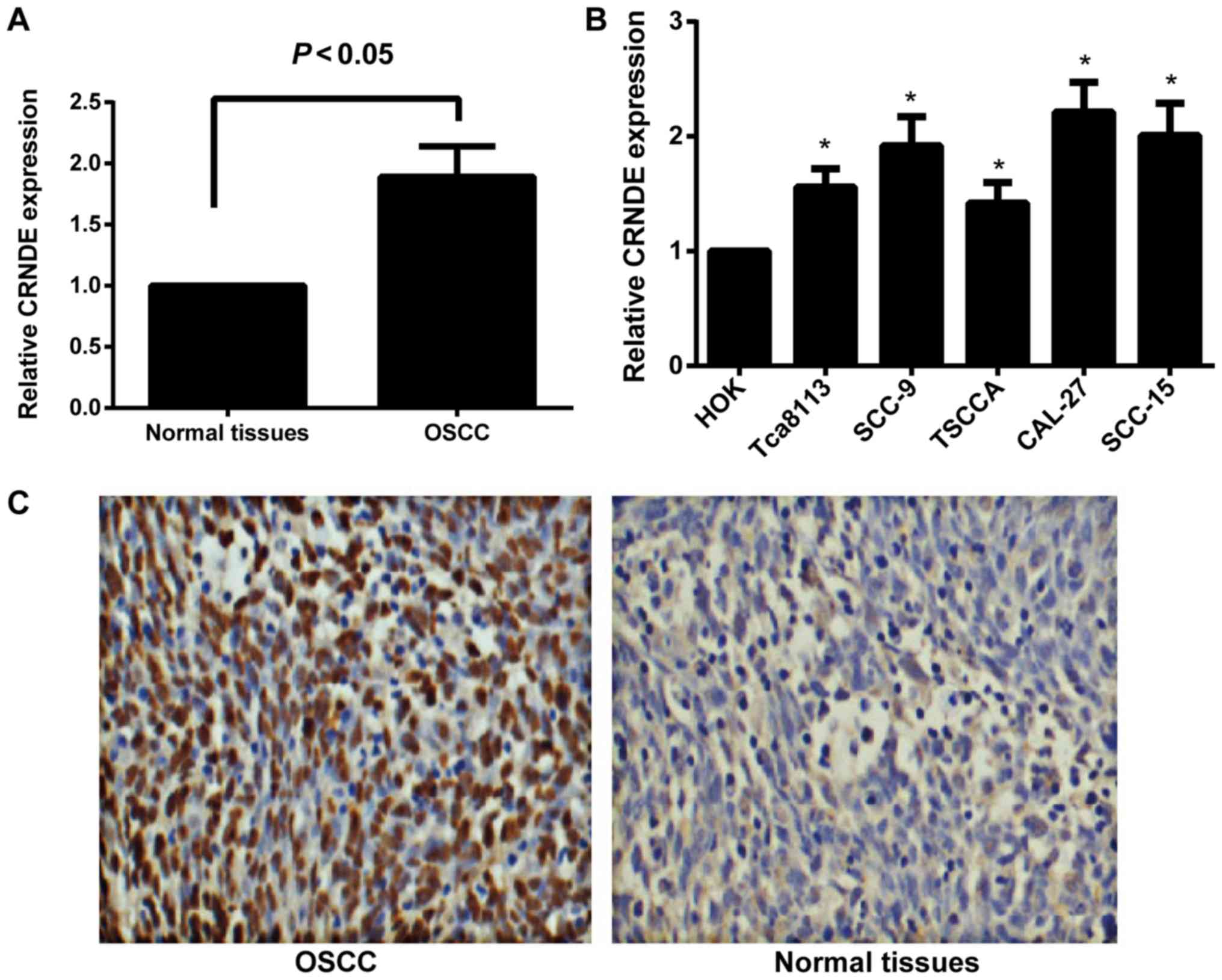 | Figure 1.Expression of lncRNA CRNDE in human
OSCC tissues and cell lines. (A) The relative expression of lncRNA
CRNDE in human OSCC and normal oral mucosa tissues, as determined
by RT-qPCR. (B) The relative expression of lncRNA CRNDE in human
HOK cells and the OSCC cell lines, Tca8113, SCC-9, TSCCA, CAL-27
and SCC-15, as determined by RT-qPCR. *P<0.05 vs. HOK cells. (C)
Expression analysis of CRNDE in human OSCC tissues and normal oral
mucosa tissues by in situ hybridization. Magnification,
×200. lncRNA, long non-coding RNA; CRNDE, colorectal neoplasia
differentially expressed; OSCC, oral squamous cell carcinoma;
RT-qPCR, reverse transcription-quantitative polymerase chain
reaction; HOK, human oral keratinocyte. |
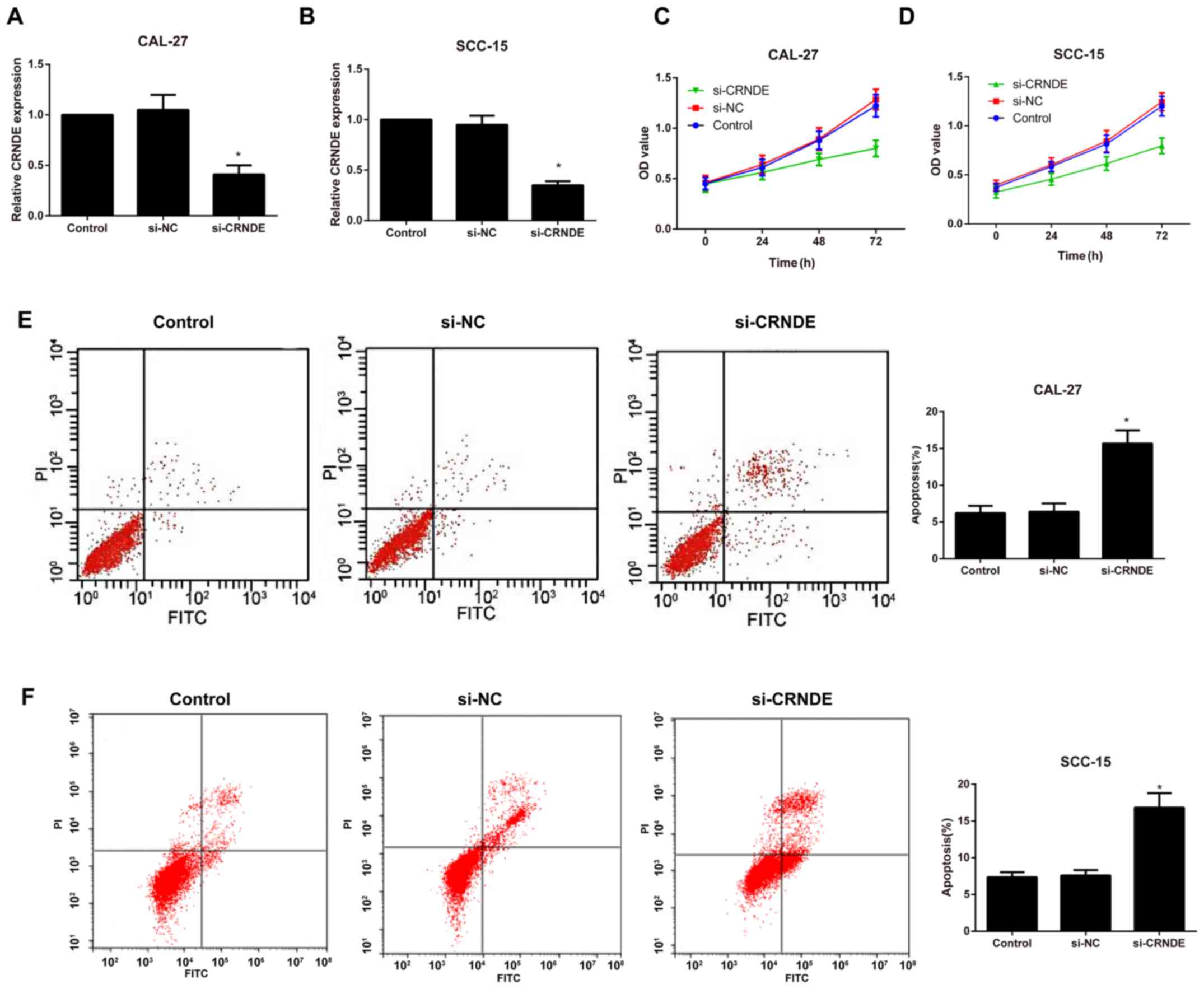
Expression of CRNDE and the effects of
si-CRNDE on proliferation and apoptosis in each group of CAL-27 and
SCC-15 cells
Following the silencing of CRNDE, RT-qPCR was
performed to detect the transfection efficiency in each group of
CAL-27 and SCC-15 cells. As illustrated in Fig. 2A and B the expression of CRNDE was
significantly downregulated in the si-CRNDE group compared with
that in the control and si-NC groups (P<0.05); however, no
observable difference was identified between the control and the
si-NC groups (P>0.05). In addition, cell proliferation and
apoptosis were evaluated by an MTT assay and flow cytometry,
respectively. Compared with the control group, the si-CRNDE group
exhibited a marked decrease in cell proliferation and an
appreciable increase in cell apoptosis (P<0.05; Fig. 2C-F); however, there was no significant
difference between the control group and the si-NC group with
respect to the proliferation and apoptosis rates at each time point
(all P>0.05).
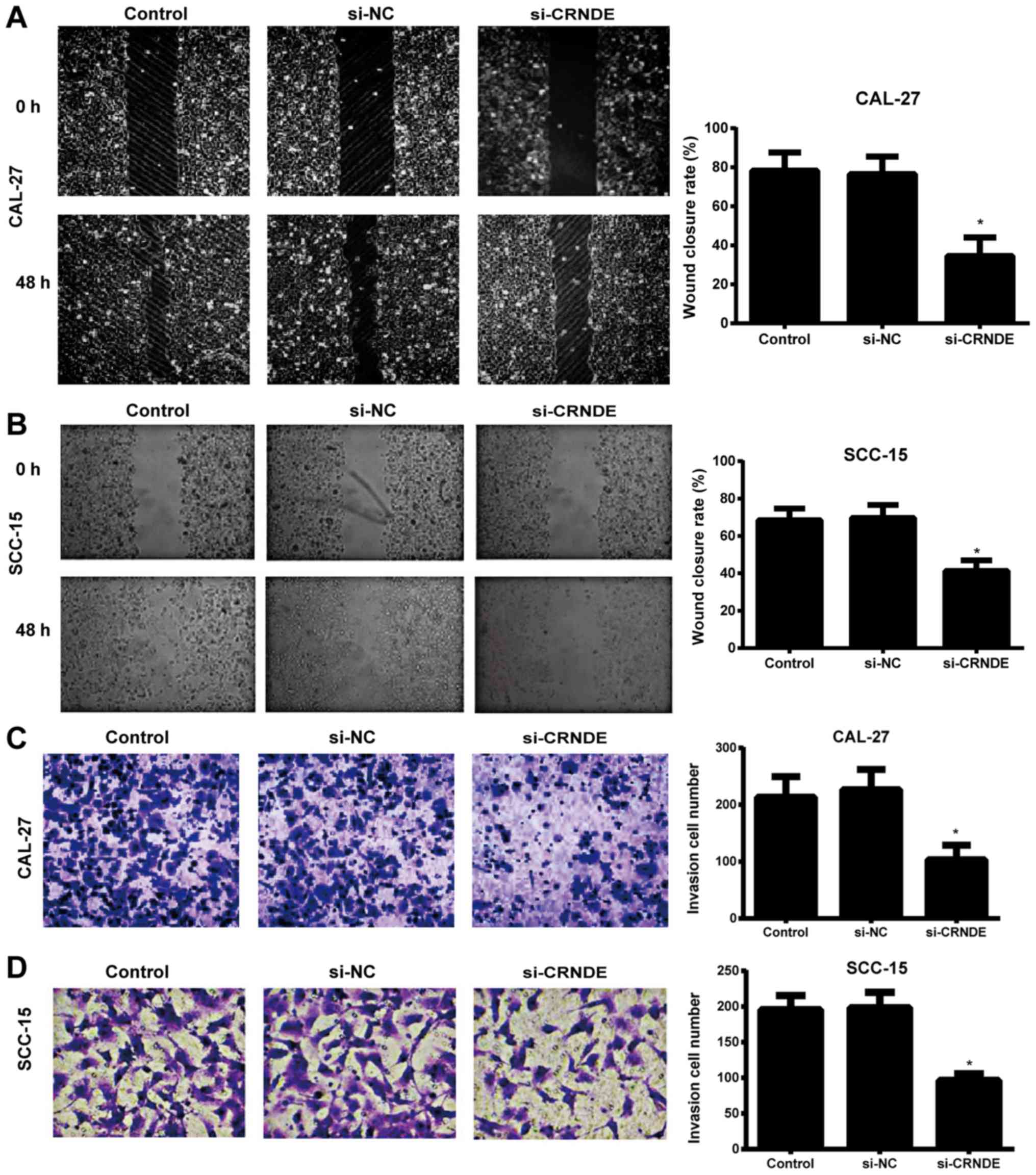
Effects of si-CRNDE on migration and
invasion in each group of CAL-27 and SCC-15 cells
A wound-healing assay was used to detect the
migration ability of each group of CAL-27 and SCC-15 cells. When
the scratch lines at the 0 and 48 h time points were compared, it
was evident that the migration ability of the CAL-27 and SCC-15
cells in the si-CRNDE group was significantly lower than that in
the control and si-NC groups (P<0.05), and that the latter two
groups exhibited no observable difference in cell migration
(P>0.05) (Fig. 3A and B). In
addition, the Transwell assay demonstrated that the number of
invasive CAL-27 and SCC-15 cells in the si-CRNDE group was
decreased significantly compared with that in the control and si-NC
groups (P<0.05); however, there was no significant difference
between the control group and the si-NC group with respect to the
number of invasive cells (P>0.05) (Fig. 3C and D).
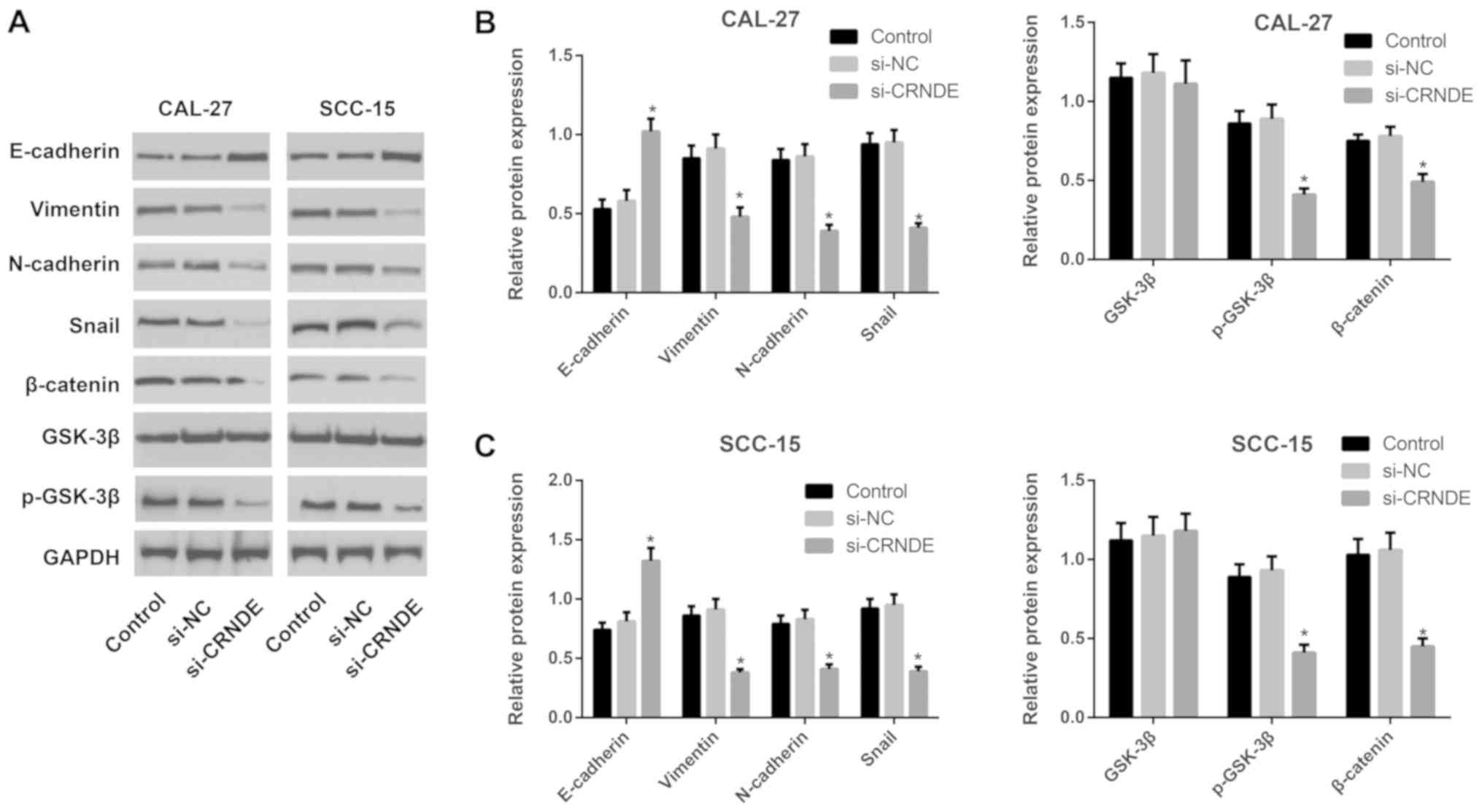
Expression of proteins associated with
the Wnt/β-catenin pathway or with EMT in each group of CAL-27 and
SCC-15 cells
Western blotting was performed to detect the
expression of proteins associated with the Wnt/β-catenin pathway or
with EMT in each group of CAL-27 and SCC-15 cells following
treatment with si-CRNDE (Fig. 4).
Compared with the cells in the control and si-NC groups, those in
the si-CRNDE group demonstrated significantly enhanced expression
of E-cadherin protein (P<0.05), but a significant reduction in
the expression of p-GSK-3β, β-catenin, N-cadherin, vimentin and
Snail protein (all P<0.05). The expression of total GSK-3β
protein did not exhibit a statistically significant difference
among the three groups in the CAL-27 and SCC-15 cells. Furthermore,
the control group was not significantly different from the si-NC
group in terms of the expression of Wnt/β-catenin pathway-related
proteins and EMT-associated proteins.
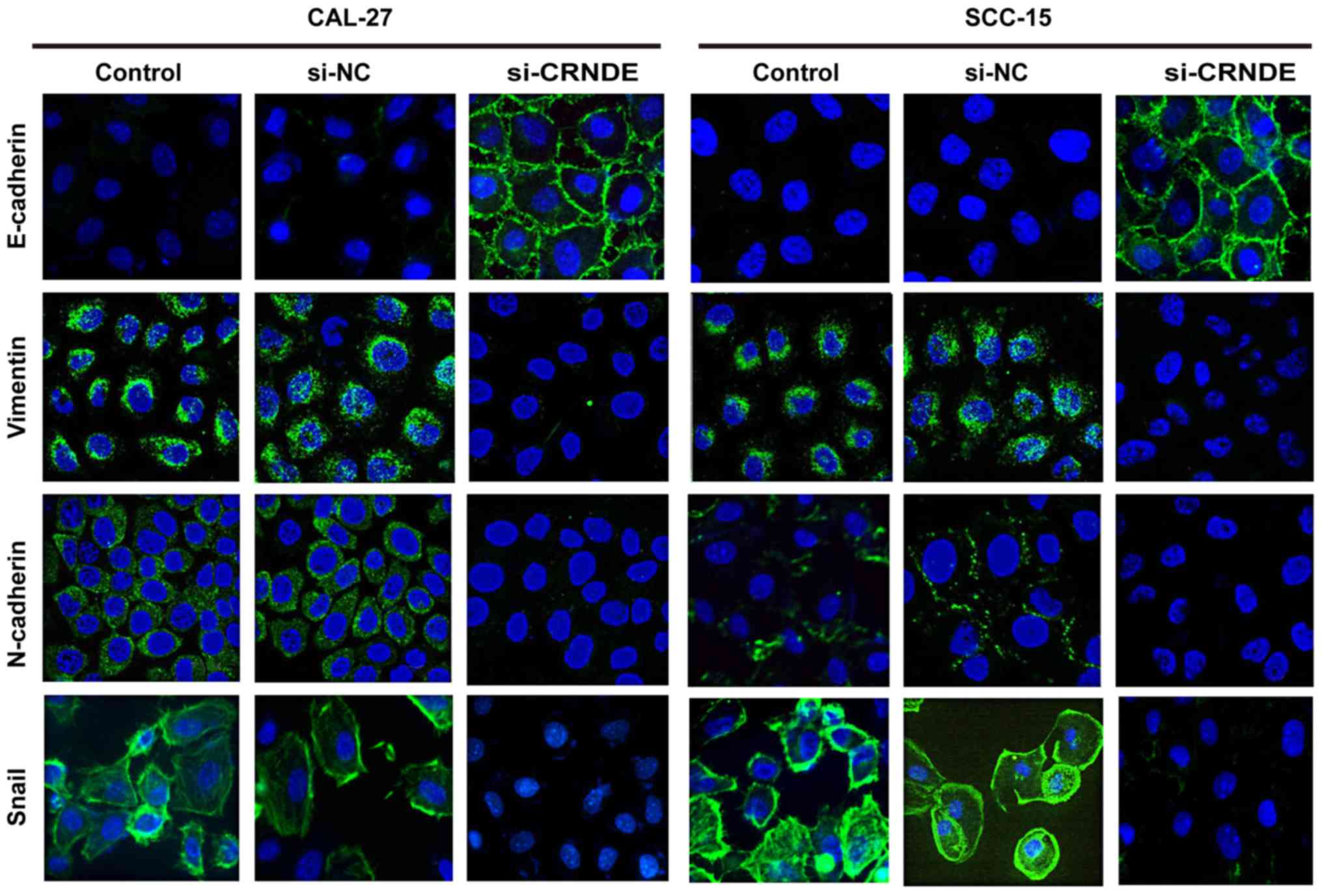
Effects of si-CRNDE on EMT in each
group of CAL-27 and SCC-15 cells
To further investigate the effect of CRNDE targeting
on EMT, immunofluorescence staining was performed to analyze the
expression of epithelial and mesenchymal markers in CAL-27 and
SCC-15 cells. As illustrated in Fig.
5, CAL-27 and SCC-15 cells in the control group displayed a
phenotypic conversion, as demonstrated by the loss of E-cadherin
and the induction of N-cadherin, vimentin and Snail. However,
treatment with CRNDE siRNA resulted in an increase in E-cadherin,
yet a decrease of N-cadherin, vimentin and Snail compared with the
expression of these proteins in the control cells. These results
suggest that downregulation of CRNDE led to a robust blockade of
EMT-like transformation in CAL-27 and SCC-15 cells.
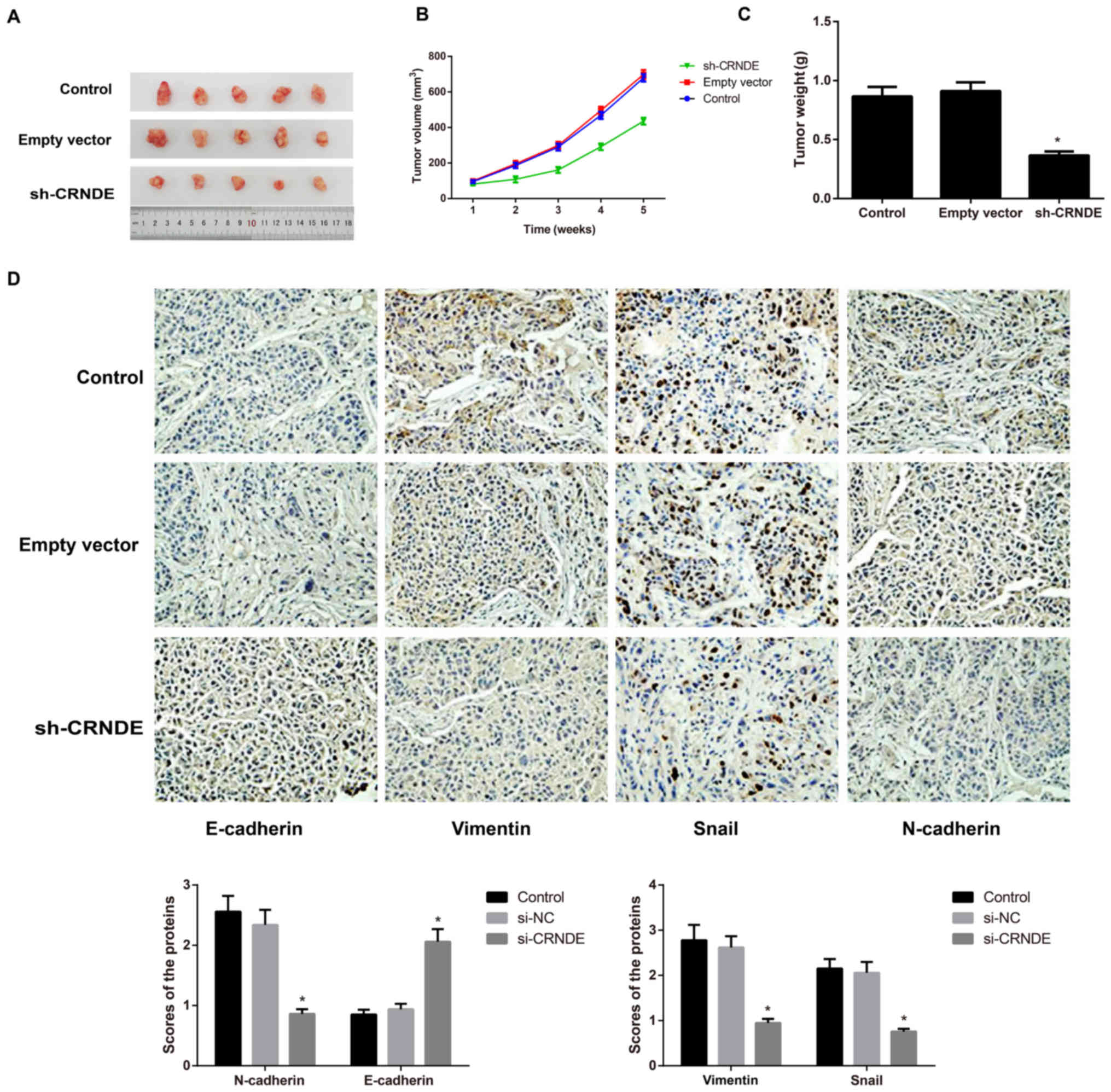
Effect of CRNDE on the growth of
subcutaneous tumors and on EMT in nude mice
Tumors formed ~1 week after tumor cell inoculation
in nude mice, with a tumor formation rate of 100%. Compared with
the control group, the sh-CRNDE group exhibited a noticeably lower
tumor growth rate and a significantly lighter tumor weight
(P<0.05), while the empty vector group did not exhibit an
apparent difference in the two indices (P>0.05) (Fig. 6A-C). Following the detection of
EMT-associated proteins in tumor tissues of nude mice with
immunohistochemical staining, the present study identified that,
compared with the control group and the empty vector group, the
sh-CRNDE group exhibited greatly increased expression of E-cadherin
protein, but appreciably decreased expression of N-cadherin,
vimentin and Snail protein (P<0.05) (Fig. 6D). However, the control group and the
empty vector group were not noticeably different in terms of the
expression of these EMT-related proteins (Fig. 6D).
Discussion
LncRNA CRNDE, was initially identified to be highly
expressed in colorectal cancer, and has become an area of
increasing focus for research; CRNDE, overexpressed in other types
of tumor cells, may function as a novel oncogene through its effect
on cellular proliferation, migration, invasion and apoptosis
(12). For instance, in the study by
Jing et al (15), CRNDE was
apparently increased in glioma tissue samples, as determined by
RT-qPCR, which could be considered an independent prognostic factor
associated with the progression of glioma (15). In chronic lymphocytic leukemia, CRNDE
was also detected to be upregulated, and this upregulation was
negatively correlated with the CRNDE methylation level, and
patients with hypermethylation of CRNDE had a significantly worse
prognosis and survival rate than those with hypomethylation of
CRNDE (21). In the present study,
the expression of CRNDE was determined to be markedly increased in
OSCC tissues and cell lines, which was consistent with the results
of the aforementioned studies. In addition, Ellis et al
(22) revealed that CRNDE was
implicated in the epigenetic regulation of chromatin and that its
transcriptional expression was regulated by insulin and
insulin-like growth factors mediating two signaling pathways,
namely, the phosphoinositide 3-kinase/AKT/mechanistic target of
rapamycin (mTOR) and RAF proto-oncogene serine/threonine-protein
kinase/mitogen-activated protein kinase pathways (22). Similarly, as indicated by Wang et
al (16), CRNDE was the most
upregulated lncRNA in gliomas owing to histone acetylation in the
promoter region, which is also influenced by mTOR signaling
(16); these findings further
highlight the oncogenic role of CRNDE in cancer types such as
OSCC.
To better understand the underlying mechanism of
CRNDE in OSCC, CAL-27 cells were selected for transfection with
CRNDE siRNA in vitro in the present study. As a result of
CRNDE silencing, the proliferation, invasion and migration of the
OSCC cells were noticeably reduced, while cell apoptosis was
clearly elevated. Similar to the findings of the present study, the
findings of Gao et al (23)
demonstrated that CRNDE acted as a competing endogenous RNA to
regulate the expression of miR-136 and that knockdown of CRNDE
significantly inhibited the invasion and migration of colorectal
cancer cells (23). Additionally,
downregulation of CRNDE led to a reduction in piwi-like
RNA-mediated gene silencing 4 protein and an elevation in miR-384
levels, consequently resulting in tumor regression (24). In addition, another important finding
from the present study was that the silencing of CRNDE could reduce
the expression of p-GSK-3β and β-catenin, downregulate the protein
expression of N-cadherin, vimentin and Snail, and enhance the
protein expression of E-cadherin, suggesting that the silencing of
CRNDE may slow the EMT process in OSCC through the inhibition of
the Wnt/β-catenin pathway and thereby inhibit the growth, invasion
and migration of OSCC cells. Consistent with this possibility, as
reported by Zhang et al (25),
downregulation of lncRNA H19 in tongue squamous cell carcinoma
(TSCC) cells was able to inhibit the activation of the
β-catenin/GSK3β signaling pathway, thus increasing the expression
of E-cadherin, decreasing the expression of N-cadherin, vimentin,
Snail1, Twist1, and ZEB1, and eventually inhibiting tumor growth.
Moreover, MALAT1 is also able to promote EMT in TSCC cells through
the activation of the Wnt/β-catenin pathway (11).
EMT has been recognized as an inevitable stage of
early tumor metastases and is characterized by common cytological
changes, including downregulation of epithelial cell markers,
including E-cadherin, upregulation of mesenchymal phenotype
markers, including vimentin and N-cadherin, and the increased
expression of EMT-inducing cytokines and transcription factors,
including Snail, Slug and Twist, which results in reduced
intercellular adhesion ability (26,27).
During this process, the tumor cells gradually lose certain
characteristics of epithelial cells and acquire the characteristics
of interstitial cells, which have stronger migration and movement
abilities, eventually giving rise to the proliferation and
metastasis of tumor cells (28,29). Thus
far, it has been demonstrated that the abnormal activation of the
Wnt/β-catenin signaling pathway is involved in the occurrence of
EMT (30). When the pathway is
abnormally activated, Wnt binds to its receptor, frizzled protein
(Frz), and acts on disheveled protein (Dsh) in the cytoplasm in
order to block GSK-3β-mediated phosphorylation/degradation of
β-catenin. The accumulated β-catenin in the cytoplasm then enters
the nucleus to interact with T cell factor/lymphoid enhancer factor
(TCF/LEF), thereby activating the expression of Snail (31,32). In
addition, as an important transcription factor in the EMT process,
Snail can bind to mSin3A and histone deacetylase to form a complex,
which can further reduce the expression of E-cadherin and destroy
the E-cadherin/β-catenin complex on the cell membrane, eventually
enhancing the expression of β-catenin in the cytoplasm and
promoting the transcription of TCF/LEF (33,34).
Therefore, Snail can induce the occurrence of EMT, mediate the
degradation of the extracellular matrix by matrix
metalloproteinase-2/9 and mediate the transcription of numerous
target genes, including c-myc and cyclin D1, thus regulating the
proliferation, apoptosis, invasion and migration of cells (35). Notably, the Wnt/β-catenin pathway,
which can induce the EMT process and promote the invasion and
migration of tumor cells, has been confirmed to be activated in
OSCC (36). In breast cancer cells,
Huan et al (37) identified
the critical role of CRNDE with regard to activation of the
Wnt/β-catenin pathway via direct repression of miR-136. Meanwhile,
CRNDE also promoted the development and chemoresistance of
colorectal cancer through the regulation of Wnt/β-catenin
signaling, as reported by Han et al (38). Considering the aforementioned
findings, silencing CRNDE may regulate the downstream mRNAs and
inhibit the activation of the Wnt/β-catenin pathway, thus
suppressing the EMT process and regulating the proliferation,
migration and invasion of OSCC cells. Nevertheless, the specific
mechanism remains to be further studied in the future. Finally,
through the tumor xenograft experiment, the present study found
that the silencing of CRNDE could slow down the tumor growth rate
and inhibit EMT in tumor-bearing nude mice, which further supported
the hypothesis that CRNDE can affect tumor growth through the
regulation of the EMT process.
In conclusion, the silencing of CRNDE may ameliorate
the EMT process via inactivation of the Wnt/β-catenin signaling
pathway, and may thus inhibit the growth but promote the apoptosis
of human OSCC cells, which provides a novel theoretical basis for
the targeted treatment of OSCC.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Author's contributions
JD and JWM contributed equally to this study. JD and
JWM designed the study and analyzed the data. HM performed the
experiments and generated the figures. JD and JWM drafted and
revised the manuscript. All authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
The present study was approved by Clinical Trial
Ethics Committee of Jingzhou Central Hospital (Jingzhou, China).
All patients involved in the present study provided written
informed consent. The animal experiments were approved by the
Ethics Committee of Jingzhou Central Hospital, The Second Clinical
Medical College, Yangtze University (Jingzhou, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Yang YT, Wang YF, Lai JY, Shen SY, Wang F,
Kong J, Zhang W and Yang HY: Long non-coding RNA UCA1 contributes
to the progression of oral squamous cell carcinoma by regulating
the WNT/β-catenin signaling pathway. Cancer Sci. 107:1581–1589.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Liu Z, Wu C, Xie N and Wang P: Long
non-coding RNA MEG3 inhibits the proliferation and metastasis of
oral squamous cell carcinoma by regulating the WNT/β-catenin
signaling pathway. Oncol Lett. 14:4053–4058. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Leusink FK, van Es RJ, de Bree R,
Baatenburg de Jong RJ, van Hooff SR, Holstege FC, Slootweg PJ,
Brakenhoff RH and Takes RP: Novel diagnostic modalities for
assessment of the clinically node-negative neck in oral
squamous-cell carcinoma. Lancet Oncol. 13:e554–e561. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Zeng G, Xun W, Wei K, Yang Y and Shen H:
MicroRNA-27a-3p regulates epithelial to mesenchymal transition via
targeting YAP1 in oral squamous cell carcinoma cells. Oncol Rep.
36:1475–1482. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Takenaka K, Chen BJ, Modesitt SC, Byrne
FL, Hoehn KL and Janitz M: The emerging role of long non-coding
RNAs in endometrial cancer. Cancer Genet. 209:445–455. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Gibb EA, Brown CJ and Lam WL: The
functional role of long non-coding RNA in human carcinomas. Mol
Cancer. 10:382011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Smith A, Teknos TN and Pan Q: Epithelial
to mesenchymal transition in head and neck squamous cell carcinoma.
Oral Oncol. 49:287–292. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
da Silva SD, Morand GB, Alobaid FA, Hier
MP, Mlynarek AM, Alaoui-Jamali MA and Kowalski LP:
Epithelial-mesenchymal transition (EMT) markers have prognostic
impact in multiple primary oral squamous cell carcinoma. Clin Exp
Metastasis. 32:55–63. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xiao C, Wu CH and Hu HZ: LncRNA UCA1
promotes epithelial-mesenchymal transition (EMT) of breast cancer
cells via enhancing Wnt/beta-catenin signaling pathway. Eur Rev Med
Pharmacol Sci. 20:2819–2824. 2016.PubMed/NCBI
|
|
10
|
Liu H, Zhen Q and Fan Y: LncRNA GHET1
promotes esophageal squamous cell carcinoma cells proliferation and
invasion via induction of EMT. Int J Boil Markers. 32:e403–e408.
2017. View Article : Google Scholar
|
|
11
|
Liang J, Liang L, Ouyang K, Li Z and Yi X:
MALAT1 induces tongue cancer cells' EMT and inhibits apoptosis
through Wnt/β-catenin signaling pathway. J Oral Pathol Med.
46:98–105. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Graham LD, Pedersen SK, Brown GS, Ho T,
Kassir Z, Moynihan AT, Vizgoft EK, Dunne R, Pimlott L, Young GP, et
al: Colorectal Neoplasia Differentially Expressed (CRNDE), a novel
gene with elevated expression in colorectal adenomas and
adenocarcinomas. Genes Cancer. 2:829–840. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu T, Zhang X, Yang YM, Du LT and Wang
CX: Increased expression of the long noncoding RNA CRNDE-h
indicates a poor prognosis in colorectal cancer, and is positively
correlated with IRX5 mRNA expression. Onco Targets Ther.
9:1437–1448. 2016.PubMed/NCBI
|
|
14
|
Szafron LM, Balcerak A, Grzybowska EA,
Pienkowska-Grela B, Podgorska A, Zub R, Olbryt M, Pamula-Pilat J,
Lisowska KM, Grzybowska E, et al: The putative oncogene, CRNDE, is
a negative prognostic factor in ovarian cancer patients.
Oncotarget. 6:43897–43910. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Jing SY, Lu YY, Yang JK, Deng WY, Zhou Q
and Jiao BH: Expression of long non-coding RNA CRNDE in glioma and
its correlation with tumor progression and patient survival. Eur
Rev Med Pharmacol Sci. 20:3992–3996. 2016.PubMed/NCBI
|
|
16
|
Wang Y, Wang Y, Li J, Zhang Y, Yin H and
Han B: CRNDE, a long-noncoding RNA, promotes glioma cell growth and
invasion through mTOR signaling. Cancer Lett. 367:122–128. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chen Z, Yu C, Zhan L, Pan Y, Chen L and
Sun C: LncRNA CRNDE promotes hepatic carcinoma cell proliferation,
migration and invasion by suppressing miR-384. Am J Cancer Res.
6:2299–2309. 2016.PubMed/NCBI
|
|
18
|
Riis P: The helsinki declaration of the
world medical association (WMA). Ethical principles of medical
research involving human subjects. Pol Merkur Lekarski. 36:298–301.
2014.(In Polish).
|
|
19
|
Jiang H, Wang Y, Ai M, Wang H, Duan Z,
Wang H, Zhao L, Yu J, Ding Y and Wang S: Long noncoding RNA CRNDE
stabilized by hnRNPUL2 accelerates cell proliferation and migration
in colorectal carcinoma via activating Ras/MAPK signaling pathways.
Cell Death Dis. 8:e28622017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Subhash S, Andersson PO, Kosalai ST,
Kanduri C and Kanduri M: Global DNA methylation profiling reveals
new insights into epigenetically deregulated protein coding and
long noncoding RNAs in CLL. Clin Epigenetics. 8:1062016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ellis BC, Graham LD and Molloy PL: CRNDE,
a long non-coding RNA responsive to insulin/IGF signaling,
regulates genes involved in central metabolism. Biochim Biophys
Acta 1843. 372–386. 2014.
|
|
23
|
Gao H, Song X, Kang T, Yan B, Feng L, Gao
L, Ai L, Liu X, Yu J and Li H: Long noncoding RNA CRNDE functions
as a competing endogenous RNA to promote metastasis and oxaliplatin
resistance by sponging miR-136 in colorectal cancer. OncoTargets
Ther. 10:205–216. 2017. View Article : Google Scholar
|
|
24
|
Zheng J, Liu X, Wang P, Xue Y, Ma J, Qu C
and Liu Y: CRNDE promotes malignant progression of glioma by
attenuating miR-384/PIWIL4/STAT3 axis. Mol Ther. 24:1199–1215.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang DM, Lin ZY, Yang ZH, Wang YY, Wan D,
Zhong JL, Zhuang PL, Huang ZQ, Zhou B and Chen WL: IncRNA H19
promotes tongue squamous cell carcinoma progression through
β-catenin/GSK3β/EMT signaling via association with EZH2. Am J
Transl Res. 9:3474–3486. 2017.PubMed/NCBI
|
|
26
|
Liu Y: Epithelial to mesenchymal
transition in renal fibrogenesis: Pathologic significance,
molecular mechanism, and therapeutic intervention. J Am Soc
Nephrol. 15:1–12. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee MY and Shen MR: Epithelial-mesenchymal
transition in cervical carcinoma. Am J Transl Res. 4:1–13.
2012.PubMed/NCBI
|
|
28
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tse JC and Kalluri R: Mechanisms of
metastasis: Epithelial-to-mesenchymal transition and contribution
of tumor microenvironment. J Cell Biochem. 101:816–829. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chiurillo MA: Role of the Wnt/β-catenin
pathway in gastric cancer: An in-depth literature review. World J
Exp Med. 5:84–102. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhou BP, Deng J, Xia W, Xu J, Li YM,
Gunduz M and Hung MC: Dual regulation of Snail by
GSK-3beta-mediated phosphorylation in control of
epithelial-mesenchymal transition. Nat Cell Biol. 6:931–940. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Guo J, Xia N, Yang L, Zhou S, Zhang Q,
Qiao Y and Liu Z: GSK-3β and vitamin D receptor are involved in
β-catenin and snail signaling in high glucose-induced
epithelial-mesenchymal transition of mouse podocytes. Cell Physiol
Biochem. 33:1087–1096. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Scheel C and Weinberg RA: Cancer stem
cells and epithelial- mesenchymal transition: Concepts and
molecular links. Semin Cancer Biol. 22:396–403. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Müller T, Bain G, Wang X and Papkoff J:
Regulation of epithelial cell migration and tumor formation by
beta-catenin signaling. Exp Cell Res. 280:119–133. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shan Y, Zhang L, Bao Y, Li B, He C, Gao M,
Feng X, Xu W, Zhang X and Wang S: Epithelial-mesenchymal
transition, a novel target of sulforaphane via COX-2/MMP2, 9/Snail,
ZEB1 and miR-200c/ZEB1 pathways in human bladder cancer cells. J
Nutr Biochem. 24:1062–1069. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang LP, Chen SW, Zhuang SM, Li H and Song
M: Galectin-3 accelerates the progression of oral tongue squamous
cell carcinoma via a Wnt/β-catenin-dependent pathway. Pathol Oncol
Res. 19:461–474. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Huan J, Xing L, Lin Q, Xui H and Qin X:
Long noncoding RNA CRNDE activates Wnt/β-catenin signaling pathway
through acting as a molecular sponge of microRNA-136 in human
breast cancer. Am J Transl Res. 9:1977–1989. 2017.PubMed/NCBI
|
|
38
|
Han P, Li JW, Zhang BM, Lv JC, Li YM, Gu
XY, Yu ZW, Jia YH, Bai XF, Li L, et al: The lncRNA CRNDE promotes
colorectal cancer cell proliferation and chemoresistance via
miR-181a-5p-mediated regulation of Wnt/β-catenin signaling. Mol
Cancer. 16:92017. View Article : Google Scholar : PubMed/NCBI
|