Introduction
ZFP91 zinc finger protein (ZFP91) is a
little-studied gene discovered in mouse in 1995 by Saotome et
al (1). In 2003 Unoki et
al (2) found, in a screening-type
study, ZFP91 overexpression in leukemic cells and neoplastic
blood cell lines. These authors were the first to detect
ZFP91 expression (using northern blot method) in several
human tissues and confirm ZFP91 protein presence in human cells-in
cultured colon and endometrial cancer cell lines. They were also
the first to characterize ZFP91 protein structure and potential
properties. It is a protein with a molecular mass of 63.4 kDa,
composed of 570 amino acid residues. It contains five zinc-finger
motives, a leucine zipper, a coiled-coil structure and nuclear
localization sequences. In mammals, it is highly conserved among
species. Based on its structure, ZFP91 was predicted to be
localized in nucleus and to act as a transcription factor (2).
ZFP91 expression is positively regulated by
NF-κB signaling pathway through NF-κB complex binding with
ZFP91 gene's 5′upstream promotor region (3). ZFP91 overexpression, on the other
hand, leads to increased NF-κB signaling pathway activation in a
manner dependent on NF-κB inducing kinase (NIK) presence (4). This kinase regulates the activity of
NF-κB non-canonical (alternative) signaling pathway (5). ZFP91 acts as an atypical E3
ubiquitin-protein ligase in NIK ubiquitinization which results in
NIK stabilization and activation of the non-canonical NF-κB
signaling pathway and its target genes expression (4,6). NIK
activity and its overexpression has been connected to cancer
pathogenesis in e.g., melanoma, pancreatic-, breast- and lung
cancer (7). The potential role of
ZFP91 in above mentioned NIK relationships remains to be
elucidated.
Another important intracellular signaling pathway
besides NF-κB has been recently discovered to be dependent on
ZFP91 expression-the hypoxia inducible factor (HIF-1)
signaling pathway (8). ZFP91
expression was found to be increased in colon cancer and positively
associated with HIF-1α expression. ZFP91 via interaction with
NF-κB/p65 protein binds to HIF-1α promoter region and upregulates
its expression. It was proven that ZFP91 has the potential to
promote proliferation of colon cancer cells in vitro and
tumor growth in vivo via HIF-1α (8). HIF-1 is a key transcription factor
responsible for cellular response to hypoxia and plays a crucial
role in adaptive responses of cancer cells to the hypoxic
microenvironment (9,10). HIF-1α together with NF-κB are two
transcription factors involved on many levels in tumors growth,
progression and resistance to chemotherapy. Novel therapy
strategies based on molecular targets within these factors'
pathways are being investigated (11–13).
Oncogenic properties of ZFP91 were revealed
also in experiments were its expression was inhibited using RNA
interference method. Unoki et al (2) found that ZFP91 inhibition in
colon cancer and endometrial cancer cell lines resulted in
increased apoptotic rate. Lee et al (3), found similarly increased apoptosis in
cultured breast and stomach cancer cell lines. What is more, cells
overexpressing ZFP91 as a result of transfection exhibited
increased growth rate and metastatic potential (3).
To date, ZFP91 expression in human cancers
has been studied almost exclusively in cancer cell lines. On a
protein level, using immunohistochemistry, ZFP91 upregulation in
colon cancer specimens was noted (8).
On a mRNA level, using in situ hybridization technique,
increased ZFP91 mRNA staining was observed in liver-,
prostate- and stomach cancer specimens (3). As reported in our earlier work,
ZFP91 mRNA overexpression was revealed in benign prostate
hyperplasia (BPH) specimens, however without concomitant protein
overexpression. In prostate cancer cell lines ZFP91 protein was
markedly upregulated compared to normal prostate epithelial cells
(14). In most recent work
ZFP91 mRNA was found to be significantly overexpressed in
prostate cancer specimens (15).
To the best of our knowledge, besides above
mentioned colon cancer and prostate studies, expression of
ZFP91 was not analyzed in human tissues and cancers
specimens on a protein level. Similarly, on a mRNA level only
qualitative data regarding ZFP91 expression in several
normal human tissues and few cancer types exist. Taking into
consideration ZFP91 important oncogenic properties, studies
are lacking analyzing this gene expression across a variety of
human normal tissues and cancer types. Presented study is the first
aimed at this gap in our knowledge about ZFP91 biology and
addressing it both on mRNA and protein level.
Materials and methods
Prostate cancer cDNA samples
Cancer Survey cDNA Arrays I, II and III (CSRT101,
CSRT103, CSRT104) from OriGene Technologies (Rockville, USA) were
utilized providing 573 cDNA samples from a variety of human normal
tissues and cancer types. Description of every sample includes
relevant clinical information, full pathology report and RNA
quality data (data available at www.origene.com/qPCR/Tissue-qPCR-Arrays.aspx).
The range of analyzed data was narrowed to organs and cancer types
with a sufficient number of independent samples. Cancer types with
insufficient number of samples e.g. adrenal cancer were excluded.
Moreover, across all three cDNA arrays a fraction of samples
occurred more than once and therefore were treated as technical
repetitions and not independent samples. Final analysis included
397 cDNA samples containing 86 normal tissues and 311 cancer
samples (Table I).
 | Table I.Analyzed types of normal tissue and
cancerous cDNA samples (from the Cancer Survey sets I, II and
III-OriGene). |
Table I.
Analyzed types of normal tissue and
cancerous cDNA samples (from the Cancer Survey sets I, II and
III-OriGene).
| Organ type | Total n | Cancer type | Total n |
|---|
| Breast | 5 | Breast cancer | 30 |
| Colon | 11 | Colon cancer | 16 |
| Endometrium | 5 | Endometrial
cancer | 21 |
| Esophagus | 4 | Esophageal cancer
(adenocarcinoma) | 14 |
| Kidney | 8 | Kidney cancer | 19 |
| Liver | 6 | Liver cancer | 15 |
| Lung | 6 | Lung (non-small
cell carcinoma) | 21 |
| Lymph node | 4 | Lymphoma
(non-Hodgkin) | 33 |
| Ovary | 5 | Ovarian cancer | 27 |
| Pancreas | 5 | Pancreatic cancer
(adenocarcinoma) | 6 |
| Prostate | 6 | Prostate
cancer | 20 |
| Stomach | 5 | Stomach cancer | 8 |
| Testis | 6 | Testicular cancer
(7 seminomas, 10 non-seminomas) | 17 |
| Thyroid | 6 | Thyroid cancer
(papillary and follicular types) | 20 |
| Urothelium | 4 | Urothelial
cancer | 23 |
| – | – | Melanoma | 11 |
| – | – | Sarcoma | 10 |
In order to choose most suitable reference genes to
test a wide range of tissue types, in a preliminary study, in 96
cDNA samples from the Cancer Survey arrays the expression of five
reference genes were tested: Tubulin alpha 1b (TUBA1B),
aminolevulinate, delta-, synthase (ALAS1), β2-microglobulin (B2 M),
actin beta (ACTB) and hypoxanthine phosphoribosyltransferase 1
(HPRT1). Using NormFinder algorithm (16) the expression of TUBA1B and
ALAS1 was found to be the most stable and this pair of genes
was used as a reference for all examined samples.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Analyses were performed as described earlier
(14,17–19).
Briefly, primers were designed by Primer 3 software (Whitehead
Institute for Biomedical Research, Cambridge, USA) and purchased
from the Laboratory of DNA Sequencing and Oligonucleotide Synthesis
(Institute of Biochemistry and Biophysics, Polish Academy of
Sciences, Warsaw, Poland). Primers sequences are listed in the
Table II. RT-qPCR was carried out in
a LightCycler 2.0 thermocycler (Roche Diagnostics, Basel,
Switzerland) with software version 4.05. SYBR Green detection
system was used based on LightCycler FastStart DNA Master SYBR
Green I mix (Roche Diagnostics). PCR reactions were carried out in
20 µl mixtures, containing 4 µl template cDNA, 0.2 µM of each gene
specific primer and 3.5 mM of Mg2+ ions. The RT-qPCR program
included a 10 min denaturation step to activate the Taq DNA
polymerase, followed by a three-step amplification program:
Denaturation at 95.0°C for 9 sec, annealing at 58.0°C for 5 sec,
and extension at 72.0°C for 5 sec. Specificity of the reaction
products was routinely checked by determination of melting points
(0.1°C/s transition rate) and random sample separation in a 2.5%
ethidium bromide/agarose gel. PCR efficiency was assessed by a
serial dilution method. Briefly, products of PCR reactions were
separated in a 2.5% agarose gel and specific bands were extracted
using a DNA gel extraction kit (EMD Millipore, Billerica, MA, USA).
The amount of extracted DNA was estimated spectrophotometrically.
Extracted DNA was diluted (10-fold serial dilutions) in order to
generate a standard curve for efficiency calculation (LightCycler
software v.4.05).
| Table II.Oligonucleotide sequences of sense
and antisense primers. |
Table II.
Oligonucleotide sequences of sense
and antisense primers.
| cDNA | Genbank accession
no. | Primer | Primer sequence
(5′-3′) | Position | PCR product size
(bp) |
|---|
| ZFP91 | NM_053023 | S |
TGTCCTTGCCCATCCTCGCTA | 1,128–1,148 | 190 |
|
|
| A |
ACTCTTGAAGGCCCGAGCAC | 1,298–1,317 |
|
| TUBA1B | NM_006082 | S |
TGGAACCCACAGTCATTGATGA | 430–451 | 135 |
|
|
| A |
TGATCTCCTTGCCAATGGTGTA | 543–564 |
|
| ALAS1 | NM_000688 | S |
AGACATAACATCTACGTGCAA | 2,031–2,051 | 167 |
|
|
| A |
GAATGAGGCTTCAGTTCCA | 2,179–2,197 |
|
| B2M | NM_004048 | S |
CAGCCCAAGATAGTTAAGTG | 385–404 | 262 |
|
|
| A |
CCCTCCTAGAGCTACCTGT | 628–646 |
|
| ACTB | NM_001101 | S |
CAGCCATGTACGTTGCTATCCAG | 473–496 | 151 |
|
|
| A |
GAGGTCCAGACGCAGGATGGCATG | 601–623 |
|
| HPRT1 | NM_000194 | S |
CTCCTCTGCTCCGCCACCG | 103–121 | 218 |
|
|
| A |
TCGAGCAAGACGTTCAGTCC | 301–320 |
|
Cancer protein lysate array
samples
ZFP91 protein expression studies were performed
using a large-scale reverse phase protein array (RPPA)-ProteoScan
Cancer Lysate Array 2.0 from OriGene. It contains 431 protein
lysates of normal and cancer specimens from 11 different tissue
types. The specimens come from accredited academic and medical
institutions in the USA and were collected according to proper
bioethical standards. All samples are provided with detailed
pathology reports and basic clinical information regarding patient
age, gender and disease staging. Array layout, sample location and
associated clinical data for each biospecimen can be found at the
OriGene website (http://www.origene.com).
As for protein extraction, all tissues were
processed within 30 min of ischemia, frozen in OCT embedding agent
at −80°C. The samples were homogenized and extracted in modified
RIPA buffer containing protease inhibitors. Protein content was
determined by BCA assay. All lysates were adjusted to 1 mg/ml using
modified RIPA buffer and diluted to 500, 250, 125 and 62.5 µg/ml in
RIPA buffer. The arrays were printed on Grace-Bio lab SuperNova
nitrocellulose slides using non-contact, inkjet printing
technologies. The total amount deposited for each spot was
approximately 300 pl. The quality of each print was verified by
Syproruby protein staining. Each sample was spotted on a slide at
five different protein concentrations and these were spotted in
triplicate on three different subarrays.
Each organ and its associated cancer type are
represented by approximately 40 samples-25 cancer samples and 15
from normal tissues. In some cases, the number of samples analyzed
was narrowed in order to include only specific cancer-type group
e.g. in case of pancreatic adenocarcinoma. All tissue types and
group sizes are listed in Table
III.
| Table III.Analyzed types of normal tissue and
cancerous protein samples (from Proteoscan Cancer Lysate
Array-OriGene). |
Table III.
Analyzed types of normal tissue and
cancerous protein samples (from Proteoscan Cancer Lysate
Array-OriGene).
| Organ type | Total n | Cancer type | Total n |
|---|
| Breast | 15 | Breast cancer | 24 |
| Colon | 15 | Colon cancer | 25 |
| Kidney | 14 | Kidney cancer | 24 |
| Liver | 15 | Liver cancer | 25 |
| Lung | 15 | Lung cancer
(non-small cell carcinoma) | 21 |
| Lymph node | 15 | Lymphoma
(non-Hodgkin) | 24 |
| Skin | 11 | Melanoma | 19 |
| Ovary | 15 | Ovarian cancer | 24 |
| Pancreas | 9 | Pancreatic cancer
(adenocarcinoma) | 7 |
| Prostate | 13 | Prostate
cancer | 22 |
| Stomach | 12 | Stomach cancer | 23 |
Immunoblotting
The array immunoblotting procedure was optimized
using ProteoScan Assay Optimization Array. After optimization,
following procedure was utilized. The array was hydrated with 10 ml
of ultrapure water for 30 min. Afterwards it was equilibrated with
TBST washing buffer for 5 min, followed by 20 min incubation in
background reducing buffer (0.5% polyvinyl alcohol; Sigma-Aldrich;
Merck KGaA, Darmstadt, Germany). The membrane was quickly washed in
washing buffer and blocked with 5 ml of array blocking buffer for
30 min (StartingBlock T20 (TBS) Blocking Buffer; Thermo Fisher
Scientific, Inc) and again rinsed with washing buffer. Membrane was
then incubated at 4°C with rabbit anti-ZFP91 at 1:50 (sc-102172;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) overnight.
Afterwards, membrane was thoroughly washed and incubated with an
anti-rabbit HRP-linked antibody at 1:500 (cat. no. 7074; Cell
Signaling Technology, Inc., Danvers, MA, USA) for 1 h at room
temperature. After washing, membrane was incubated with
Biotinyl-tyramide solution (Tyramide Signal Amplification (TSA™)
from PerkinElmer, Inc., Waltham, MA, USA) for 25 min and washed
again. Streptavidin-Texas red conjugate diluted 1:100 in blocking
buffer was added for 30 min at room temperature. The membrane was
thoroughly washed and finally rinsed for 10 min in ultrapure water.
Afterwards the array was air-dried and scanned using Genepix
microarray scanner. Signal intensities from samples spotted at
maximum protein concentration (1 mg/ml) were found to be falling
best within dynamic range of the detection method for all tested
samples and were utilized for comparative analysis.
Statistical analysis
GraphPad Prism v.5.00 (GraphPad Software, Inc., San
Diego, CA, USA) was used to perform statistical analyses. P<0.05
was considered to indicate a statistically significant difference.
Depending on the number of groups being statistically compared
either a Mann-Whitney or Kruskal-Wallis test was performed with
Dunn's test. The results are presented as the median and the
interquartile range, or as box and whisker plots with the whiskers
representing the 5 and 95th percentile.
Results
ZFP91 mRNA expression pattern in
normal human tissues and cancers
Firstly, cDNA samples from 15 normal human tissues
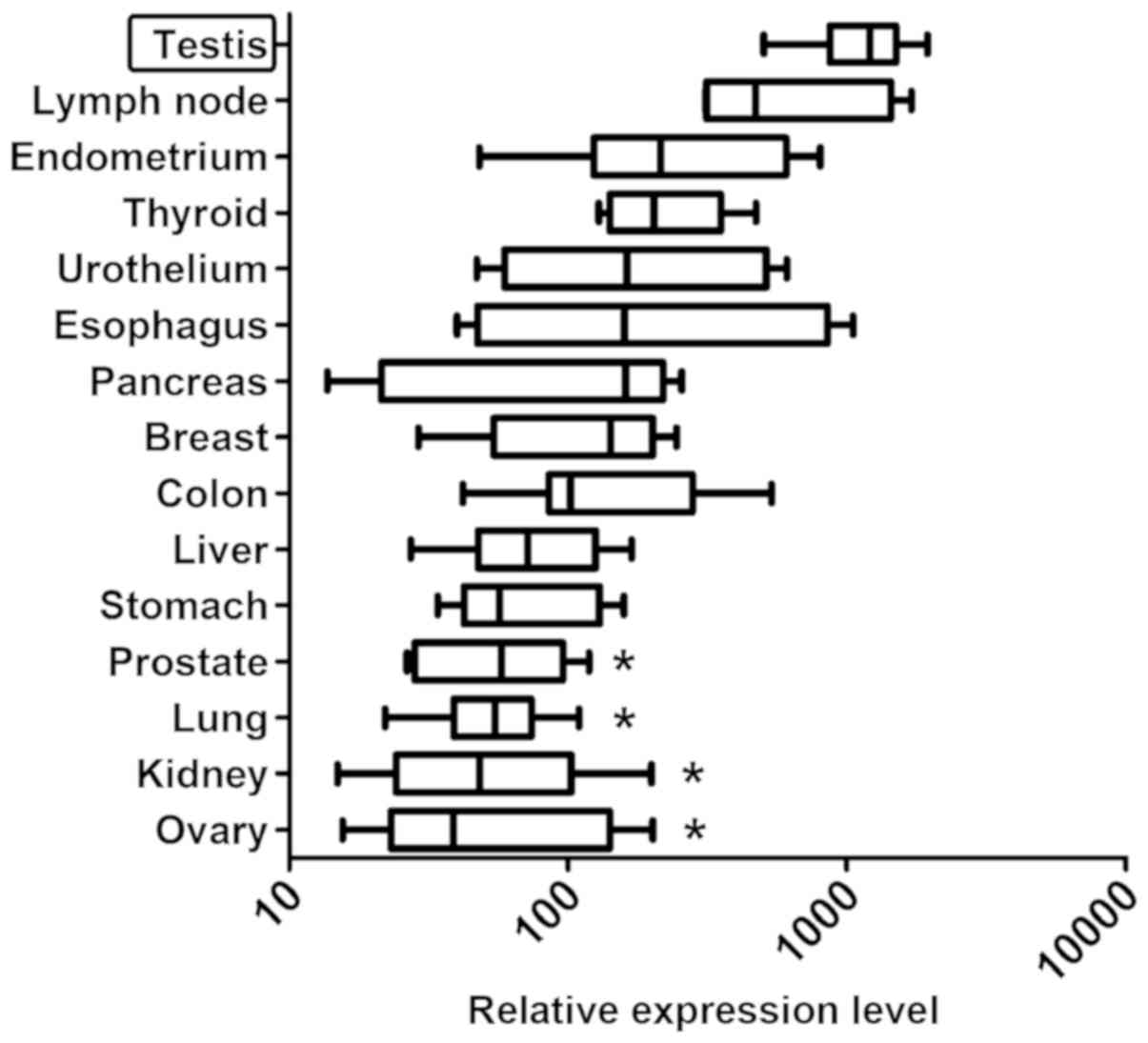
were analyzed. The highest ZFP91 expression was found in
testis (Fig. 1). It was significantly
higher than in prostate, lung, kidney and ovary. The difference
between highest expression-in testis and lowest-in ovary was more
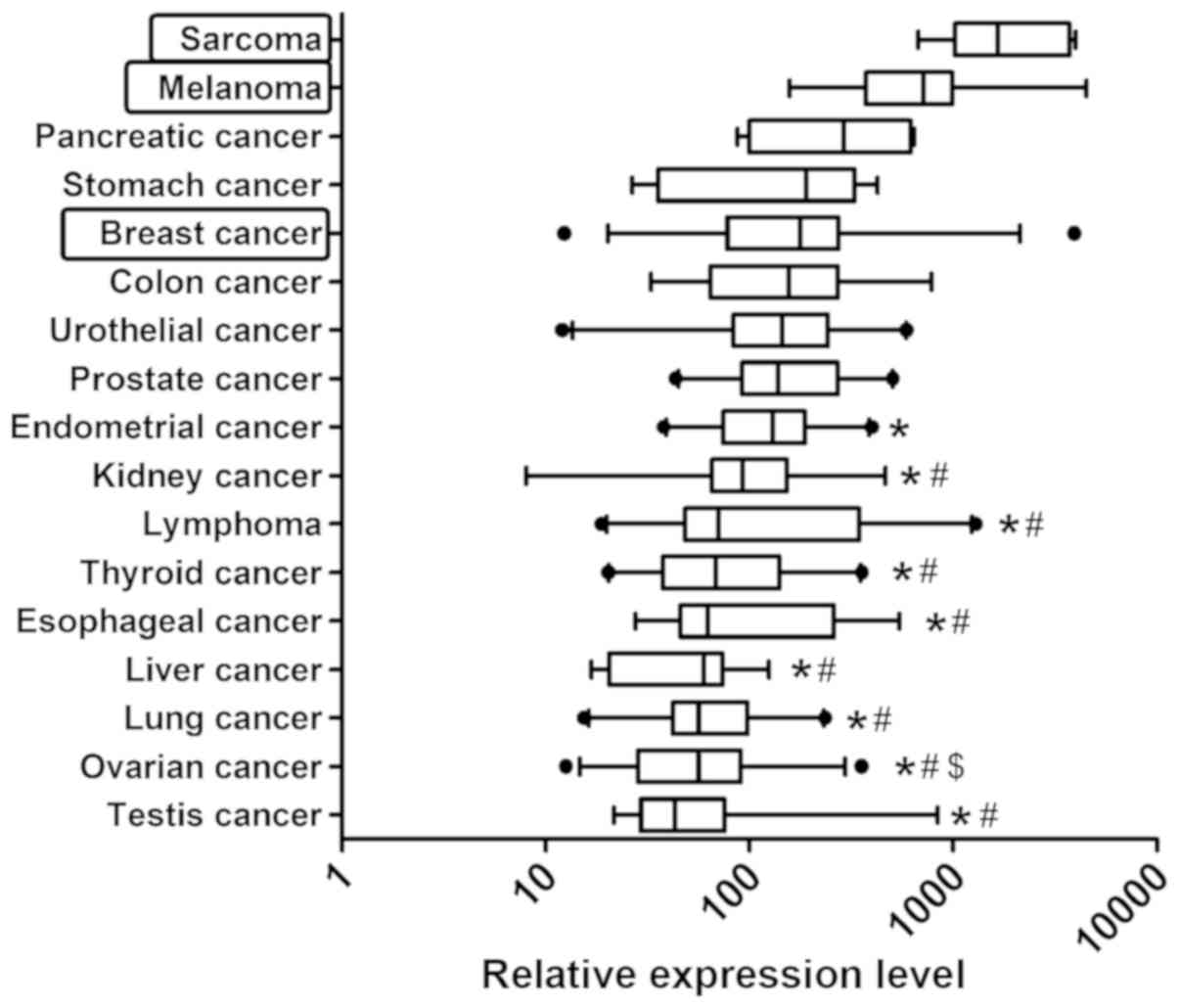
than 10-fold. Among the 17 cancer types analyzed, ZFP91
expression was the highest in sarcomas and melanoma (Fig. 2), while the lowest in testis and
ovarian cancer.
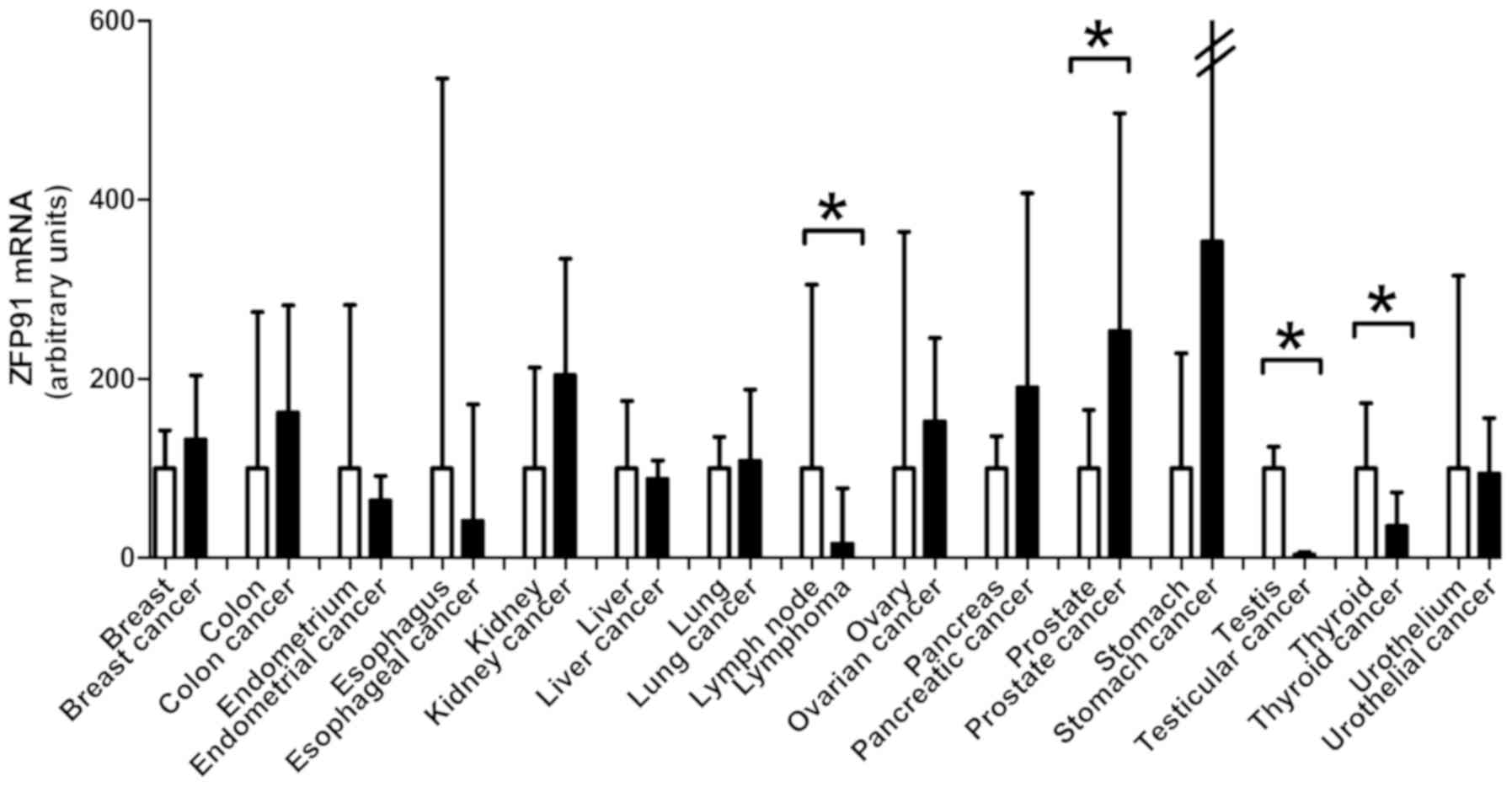
Finally, studied gene expression was compared
between 15 normal tissue types and cancers derived from them
(Fig. 3). Testis cancer deriving from
organ with the highest ZFP91 expression-testis, had its
expression significantly lower. No difference with this regard was
noted between seminomas and non-seminomas. Second organ with
highest ZFP91 expression-lymph nodes, also differed
significantly from analyzed cases of non-Hodgkin lymphomas. In the
case of thyroid cancer, similarly a downregulation of ZFP91
expression compared to normal thyroid was noted. The only cancer
type with elevated ZFP91 mRNA expression compared to normal
tissue was prostate cancer where the results are in accordance with
our earlier reports of ZFP91 upregulation in this disease.
In majority of the analyzed cancer types no significant differences
in ZFP91 mRNA levels were observed between normal tissue and
cancer.
ZFP91 protein levels in 11 human
organs and cancer types
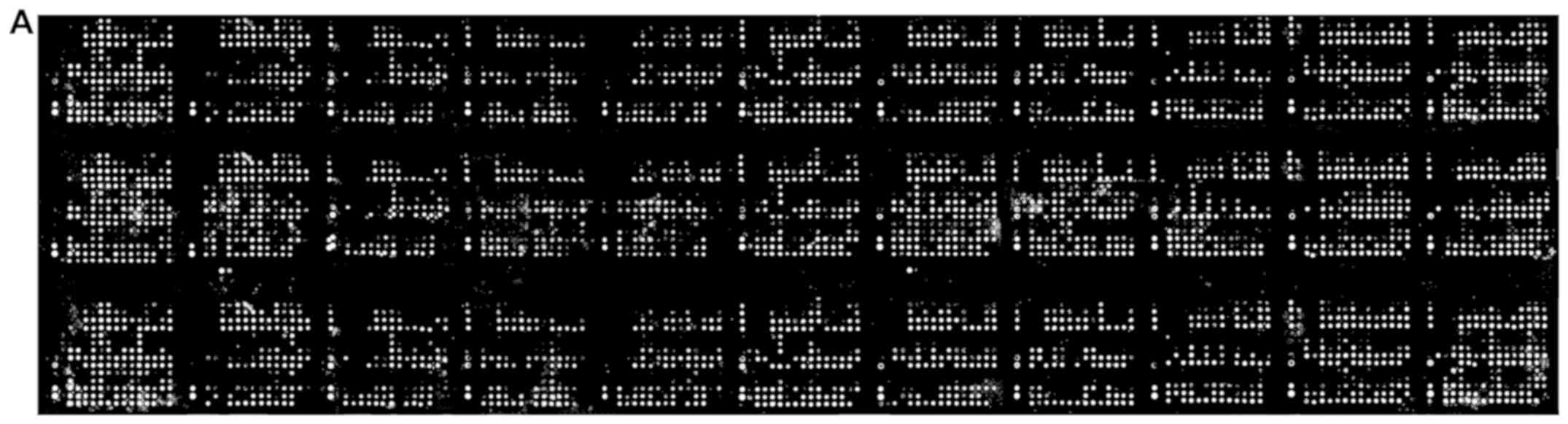
A large scale reverse phase protein array allowed
for analysis of 431 protein lysates from normal and cancer
specimens spanning 11 different tissues (Fig. 4A). ZFP91 protein expression was found
to be ubiquitous and markedly less diverse than ZFP91 mRNA
levels (Fig. 4B). Analyzed tissues
and cancers presented similar ZFP91 protein levels. In most cases
its expression did not differ between normal tissue and cancer
derived from it. What is interesting, neither melanoma with
observed exceptionally high ZFP91 mRNA levels nor prostate
cancer with significant ZFP91 mRNA overexpression were found
to present similar changes on a protein level. On the other hand,
in the case of breast cancer, ovarian cancer and pancreatic cancer
protein analysis revealed significant upregulation of ZFP91 levels,
despite no differences noted in mRNA expression analysis.
Discussion
Earlier findings regarding ZFP91 expression
in normal human tissues showed ZFP91 mRNA presence in all
studied organs, with a particularly high expression in testis
(2). The present study confirmed
ZFP91 expression in tissues not to date examined: lymph
node, endometrium, thyroid gland, urothelium, esophagus, breast and
stomach. Particularly high ZFP91 expression in testis was
also observed in our material. It is difficult to assess the value
of this observation. Studies have proved that some of the mRNA
transcripts found in testis are not present in other tissues and no
evidence of their translation and function can be found (20–22). What
is interesting, in mouse ZFP91 is one of the genes
potentially causing male infertility (23). Unfortunately, this issue has not been
further studied.
Another tissue with relatively high ZFP91
expression is lymph node and leukocytes it contains. This might be
connected with NF-κB signaling pathway activity in these cells.
NF-κB signaling pathway is present in its inducible form in
virtually all cell types (24).
However, in most cells of the immune system it is constitutionally
activated (25–27). The constitutive activation of NF-κB
signaling pathway is necessary to block apoptosis in mature
lymphocytes in phase G0 of cell cycle (28). This suggests that higher ZFP91
expression in lymph node may be a result of higher NF-κB signaling
pathway activity.
Among the 17 cancer types examined, two have shown
markedly increased ZFP91 expression-sarcomas and melanoma.
Overactivity of the NF-κB signaling pathway observed in these
cancer types (29,30) seems not to be a sole cause of this
phenomenon since it is also present in other types of aggressive
cancers (31). The highest expression
of ZFP91 in analyzed cases of sarcomas may reflect unique
features associated with this type of cancer-aggressive,
non-epithelial neoplasm of mesenchymal origin. Only sarcomas among
studied cancer types originate from transformed mesenchymal stem
cells (32). ZFP91 expression
was at the second-highest level in melanoma, a cancer type
possessing some distinct similarities to sarcomas. This cancer type
is also an aggressive neoplasm but of neuroectodermal origin. A
transformation towards mesenchymal phenotype plays, however, an
important role in melanoma's development (33). In the embryogenesis, melanocytes
migrate from the neural crest and this process is facilitated by
transient mesenchymal phenotype. Mature melanocytes present some of
the epithelial cells markers, but their neoplastic transformation
into melanoma cells and melanoma's progression are related to their
change in phenotype towards mesenchymal cells (34). They become more mobile, acquire the
capacity to interact with extracellular matrix, to disseminate and
metastasize (35). A question arises
whether ZFP91 overexpression in sarcomas and melanoma is
connected to their mesenchymal properties or presents just a
concomitant phenomenon.
Comparative analysis of ZFP91 expression in
human organs and respective cancer types revealed significant
differences in a few cases. In testis cancer, non-Hodgkin lymphoma
and thyroid cancer, ZFP91 expression was markedly lower in
cancer samples. This may be connected to the fact that testis,
lymph node and thyroid were all organs with the highest
ZFP91 expression compared to other organs. Decreased
ZFP91 expression in the cancer types stemming from them
seems to be secondary to the changes in cellular phenotype. Further
studies are required to elucidate full physiological role of ZFP91
in human organs. In the case of lymphoma cells a decrease in
ZFP91 expression is particularly interesting, taking into
consideration data showing its overexpression in leukemia cells
(2). ZFP91 overexpression is
therefore not a common feature of all cancers originating from
immune system cells.
Among studied cancer types only in the case of
prostate cancer ZFP91 expression was significantly elevated
in comparison to the unchanged prostatic tissue. Similarly as in
our previous study on this subject (15), the increased (even 10-fold)
ZFP91 mRNA quantity was present only in the fraction of
cancer cases. ZFP91 overexpression on mRNA level was
observed by Lee et al (3) in
cancers of prostate, stomach and liver. In our material, no
differences were noted in cases of stomach and liver cancer. Based
on the obtained results, it can be stated that ZFP91 mRNA
regulation is not a phenomenon typical for oncogenic
transformation.
Results of the reverse phase protein array analysis
are the first to our knowledge to survey ZFP91 expression over a
wide range of tissues and cancer types. Changes in ZFP91
mRNA levels observed in corresponding tissues were not reflected on
a protein level. A discrepancy between mRNA and protein levels is
often encountered (36–39) and in the case of ZFP91 has been
described in our previous works (14,15) as
well as observed by the team of Lee et al (3). At this point of knowledge it seems that
no clear correlation between ZFP91 mRNA and protein levels
exist. Based on previous experiments one can hypothesize that ZFP91
is subjected to posttranscriptional regulation within the cells.
Further experiments in the field should establish whether this
regulation involves changes in ZFP91 protein structure affecting
its stability, particularly in neoplastic cells. Neither
ZFP91 overexpression in melanoma and prostate cancer nor
underexpression in lymphoma was observed on a protein level. On the
other hand, in the case of breast cancer, ovarian cancer and
pancreatic cancer a significant, although limited, protein
upregulation was observed in our material. On a mRNA level these
three types of cancer showed also limited upregulation although
observed changes did not reach statistical significance. What is
important, in the case of colon cancer no ZFP91 protein
upregulation was noted, contrary to the results of the study
performed by Ma et al (8). It
can be hypothesized, that differences in the methodology and in the
material may be the factors responsible for it. Furthermore, the
character of RPPA analysis performed in this study makes it
suitable for survey purposes only and may have limited sensitivity
to detect more subtle changes in protein expression.
Several limitations of the presented study can be
brought up and should be addressed in future experiments. Firstly,
an immunohistochemical staining of ZFP91 protein in cancer tissue
and adjacent healthy tissue would provide an interesting comparison
as well as analysis of paired samples. This was not done in this
study as provided sample sets consisted mostly of unpaired samples.
Another interesting issue would be to analyze ZFP91
expression with regard to cancer sub-types and grading, however
limited sample size of each group prevented us from further
subdivisions. As for relation between ZFP91 expression and
patients' prognosis and survival this could not be addressed in
this study due to lack of such data. It would be interesting to
analyze particular cancer types highlighted by this study e.g.
melanoma, with the use of cell lines and patient samples including
evaluation of patients' prognosis and treatment results.
Presented study is the first to examine ZFP91
expression pattern over a wide range of human tissues and cancer
types. Obtained results indicate that ZFP91 is a
ubiquitously expressed gene with overall stable expression levels.
Revealed differences in studied gene expression, especially high
mRNA levels in melanoma and sarcomas and elevated protein levels in
cancers of breast, ovary and pancreas, are particularly interesting
with regard to ZFP91 oncogenic potential. Further studies are,
however, required to assess biological significance of the observed
changes.
Acknowledgements
Not applicable.
Funding
The present study was supported by a grant from
Ministry of Science and Higher Education of the Republic of Poland
(grant no. DI2011 0219 41).
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
LP, LKM and MR designed the study. LP, KJ, MS, MT
and MR performed the experiments and analyzed the data. LP wrote
the manuscript. LKM and MR revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Saotome Y, Winter CG and Hirsh D: A widely
expressed novel C2H2 zinc-finger protein with multiple consensus
phosphorylation sites is conserved in mouse and man. Gene.
152:233–238. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Unoki M, Okutsu J and Nakamura Y:
Identification of a novel human gene, ZFP91, involved in acute
myelogenous leukemia. Int J Oncol. 22:1217–1223. 2003.PubMed/NCBI
|
|
3
|
Lee JJ, Lee JH, Lee K, Hong Ys and Jin X:
Therapeutic agent for cancer, inflammation and auto-immune disease
containing inhibitor of Zinc Finger Protein 91. US Patent 20:
080,248,024. 2008.
|
|
4
|
Jin X, Jin HR, Jung HS, Lee SJ, Lee JH and
Lee JJ: An atypical E3 ligase zinc finger protein 91 stabilizes and
activates NF-kappaB-inducing kinase via Lys63-linked
ubiquitination. J Biol Chem. 285:30539–30547. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sun SC: Non-canonical NF-κB signaling
pathway. Cell Res. 21:71–85. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Jin HR, Jin X and Lee JJ: Zinc-finger
protein 91 plays a key role in LIGHT-induced activation of
non-canonical NF-κB pathway. Biochem Biophys Res Commun.
400:581–586. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Xiao G and Fu J: NF-κB and cancer: A
paradigm of Yin-Yang. Am J Cancer Res. 1:192–221. 2011.PubMed/NCBI
|
|
8
|
Ma J, Mi C, Wang KS, Lee JJ and Jin X:
Zinc finger protein 91 (ZFP91) activates HIF-1α via NF-κB/p65 to
promote proliferation and tumorigenesis of colon cancer.
Oncotarget. 7:36551–36562. 2016.PubMed/NCBI
|
|
9
|
Mabjeesh NJ and Amir S: Hypoxia-inducible
factor (HIF) in human tumorigenesis. Histol Histopathol.
22:559–572. 2007.PubMed/NCBI
|
|
10
|
Masoud GN and Li W: HIF-1α pathway: Role,
regulation and intervention for cancer therapy. Acta Pharm Sin B.
5:378–389. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Tafani M, Pucci B, Russo A, Schito L,
Pellefrini L, Perrone AG, Villanova L, Salvatori L, Ravenna L,
Petrangeli E and Russo MA: Modulators of HIF1α and NFkB in cancer
treatment: Is it a rational approach for controlling malignant
progression? Front Pharmacol. 4(13)2013.PubMed/NCBI
|
|
12
|
Hoesel B and Schmid JA: The complexity of
NF-κB signaling in inflammation and cancer. Mol Cancer. 12:862013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Hu Y, Liu J and Huang H: Recent agents
targeting HIF-1α for cancer therapy. J Cell Biochem. 114:498–509.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Paschke L, Rucinski M, Ziolkowska A,
Zemleduch T, Malendowicz W, Kwias Z and Malendowicz LK: ZFP91-a
newly described gene potentially involved in prostate pathology.
Pathol Oncol Res. 20:453–459. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Paschke L, Jopek K, Szyszka M, Tyczewska
M, Ziolkowska A, Rucinski M and Malendowicz LK: ZFP91: A
noncanonical NF-κB signaling pathway regulator with oncogenic
properties is overexpressed in prostate cancer. Biomed Res Int.
2016:69635822016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Andersen CL, Jensen JL and Ørntoft TF:
Normalization of real-time quantitative reverse transcription-PCR
data: A model-based variance estimation approach to identify genes
suited for normalization, applied to bladder and colon cancer data
sets. Cancer Res. 64:5245–5250. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Paschke L, Zemleduch T, Rucinski M,
Ziolkowska A, Szyszka M and Malendowicz LK: Adiponectin and
adiponectin receptor system in the rat adrenal gland: Ontogenetic
and physiologic regulation, and its involvement in regulating
adrenocortical growth and steroidogenesis. Peptides. 31:1715–1724.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Szyszka M, Paschke L, Tyczewska M,
Rucinski M, Grabowska P and Malendowicz LK: Lack of expression of
preproorexin and orexin receptors genes in human normal and
prostate cancer cell lines. Folia Histochem Cytobiol. 53:333–341.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jopek K, Celichowski P, Szyszka M,
Tyczewska M, Milecka P, Malendowicz LK and Rucinski M:
Transcriptome profile of rat adrenal evoked by gonadectomy and
testosterone or estradiol replacement. Front Endocrinol (Lausanne).
8:262017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Dobner PR, Kislauskis E, Wentworth BM and
Villa-Komaroff L: Alternative 5′exons either provide or deny an
initiator methionine codon to the same alpha-tubulin coding region.
Nucleic Acids Res. 15:199–218. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Garrett JE, Collard MW and Douglass JO:
Translational control of germ cell-expressed mRNA imposed by
alternative splicing: Opioid peptide gene expression in rat testis.
Mol Cell Biol. 9:4381–4389. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Stallard BJ, Collard MW and Griswold MD: A
transferrinlike (hemiferrin) mRNA is expressed in the germ cells of
rat testis. Mol Cell Biol. 11:1448–1453. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Trachtulec Z, Mnuková-Fajdelová M, Hamvas
RM, Gregorová S, Mayer WE, Lehrach HR, Vincek V, Forejt J and Klein
J: Isolation of candidate hybrid sterility 1 genes by cDNA
selection in a 1.1 megabase pair region on mouse chromosome 17.
Mamm Genome. 8:312–316. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lenardo MJ and Baltimore D: NF-kappa B: A
pleiotropic mediator of inducible and tissue-specific gene control.
Cell. 58:227–229. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kolenko V, Bloom T, Rayman P, Bukowski R,
Hsi E and Finke J: Inhibition of NF-kappa B activity in human T
lymphocytes induces caspase-dependent apoptosis without detectable
activation of caspase-1 and −3. J Immunol. 163:590–598.
1999.PubMed/NCBI
|
|
26
|
Ward C, Chilvers ER, Lawson MF, Pryde JG,
Fujihara S, Farrow SN, Haslett C and Rossi AG: NF-kappaB activation
is a critical regulator of human granulocyte apoptosis in vitro. J
Biol Chem. 274:4309–4318. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Griffin GE, Leung K, Folks TM, Kunkel S
and Nabel GJ: Activation of HIV gene expression during monocyte
differentiation by induction of NF-kappa B. Nature. 339:70–73.
1989. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bureau F, Vanderplasschen A, Jaspar F,
Minner F, Pastoret PP, Merville MP, Bours V and Lekeux P:
Constitutive nuclear factor-kappaB activity preserves homeostasis
of quiescent mature lymphocytes and granulocytes by controlling the
expression of distinct Bcl-2 family proteins. Blood. 99:3683–3691.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Horiuchi K, Morioka H, Nishimoto K, Suzuki
Y, Susa M, Nakayama R, Kawai A, Sonobe H, Takaishi H, Ozaki T, et
al: Growth suppression and apoptosis induction in synovial sarcoma
cell lines by a novel NF-kappaB inhibitor,
dehydroxymethylepoxyquinomicin (DHMEQ). Cancer Lett. 272:336–344.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Madonna G, Ullman CD, Gentilcore G,
Palmieri G and Ascierto PA: NF-κB as potential target in the
treatment of melanoma. J Transl Med. 10:532012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lin Y, Bai L, Chen W and Xu S: The
NF-kappaB activation pathways, emerging molecular targets for
cancer prevention and therapy. Expert Opin Ther Targets. 14:45–55.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Xiao W, Mohseny AB, Hogendoorn PC and
Cleton-Jansen AM: Mesenchymal stem cell transformation and sarcoma
genesis. Clin Sarcoma Res. 3:102013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mikesh LM, Kumar M, Erdag G, Hogan KT,
Molhoek KR, Mayo MW and Slingluff CL Jr: Evaluation of molecular
markers of mesenchymal phenotype in melanoma. Melanoma Res.
20:485–495. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Alonso SR, Tracey L, Ortiz P, Pérez-Gómez
B, Palacios J, Pollán M, Linares J, Serrano S, Sáez-Castillo AI,
Sánchez L, et al: A high-throughput study in melanoma identifies
epithelial-mesenchymal transition as a major determinant of
metastasis. Cancer Res. 67:3450–3460. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Grunert S, Jechlinger M and Beug H:
Diverse cellular and molecular mechanisms contribute to epithelial
plasticity and metastasis. Nat Rev Mol Cell Biol. 4:657–665. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Maier T, Güell M and Serrano L:
Correlation of mRNA and protein in complex biological samples. FEBS
Lett. 583:3966–3973. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Maier T, Schmidt A, Güell M, Kühner S,
Gavin AC, Aebersold R and Serrano L: Quantification of mRNA and
protein and integration with protein turnover in a bacterium. Mol
Syst Biol. 7:5112011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Taniguchi Y, Choi PJ, Li GW, Chen H, Babu
M, Hearn J, Emili A and Xie XS: Quantifying E. coli proteome and
transcriptome with single-molecule sensitivity in single cells.
Science. 329:533–538. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vogel C and Marcotte EM: Insights into the
regulation of protein abundance from proteomic and transcriptomic
analyses. Nat Rev Genet. 13:227–232. 2012. View Article : Google Scholar : PubMed/NCBI
|