Introduction
Prostate cancer (PCa) is a malignancy of the urinary
system (1). The probability of newly
diagnosed PCa is high in European and American men (2). Worldwide it has become the second
leading cause of cancer-associated mortality in men (3). Patients with localized PCa are
generally treated with hormones, surgery, and chemotherapy or
radiation therapy. Hormone therapy is effective at early stages;
however, a number of patients slowly develop androgen insensitivity
(4,5). When advanced PCa develops into
castration-resistant PCa and metastasizes to distant sites, taxanes
are often used for treatment (6).
Paclitaxel (PTX) is an alkaloid that has been used
as a first-line treatment for PCa in a clinical setting. It exerts
its antitumor effects via cell cycle arrest (7). Resistance to PTX limits its therapeutic
effect (8). Chemoresistant
metastatic PCa is the most lethal form of cancer in adult men
(9); therefore, more effective
treatments for PCa are required.
Metformin (MET) is an oral anti-diabetic drug,
commonly used to treat type 2 diabetes mellitus (10). It is the most commonly used biguanide
drug, and has a relatively low incidence of toxicity and side
effects (11,12). MET has attracted increased attention
in recent years due to its possible anticancer activity; it
inhibits several tumor types, including PCa (13–15). A
number of in vivo experiments have revealed that MET
directly affects cancer cell growth. Its effects have been observed
in a wide range of cancer cell lines, including PCa cell lines
(16,17). MET induces apoptosis and cell cycle
arrest, reducing cancer cell growth (18,19). A
previous study reported that MET increases sensitivity to
chemotherapy and decreases required chemotherapy drug doses in
various cancer cell lines (20).
Given its excellent safety profile, low cost and
minimal side effects, MET is an attractive candidate as a potential
anticancer agent. Nevertheless, there remains limited knowledge
regarding its anticancer molecular mechanisms. Therefore, the
present study investigated the effects of MET in combination with
PTX on apoptosis of 22RV1, PC-3 and LNCaP cells, as well as the
molecular mechanisms underlying these effects. In the present study
it was demonstrated that MET augmented the effects of PTX.
Materials and methods
Cell culture
Human PCa cell lines 22RV1, PC-3 and LNCaP were
purchased from the Chinese Academy of Sciences Cell Bank (Shanghai,
China). The three cell lines were cultured in RPMI-1640 medium
(Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA),
supplemented with 10% fetal bovine serum (FBS; Gibco; Thermo Fisher
Scientific, Inc.) for 22RV1 and PC-3 cells, and with 12% FBS for
LNCaP cells at 37°C. Finally, a mixture of penicillin and
streptomycin (Beyotime Institute of Biotechnology, Shanghai, China)
at a final concentration of 1% was added.
Reagents and antibodies
MET and PTX were purchased from Beijing Solarbio
Science & Technology Co., Ltd. (Beijing, China). MET was
dissolved in 1X PBS to a concentration of 2 M, and PTX was
dissolved in 100% dimethyl sulfoxide (DMSO) to create a 10 mM stock
solution; these were stored at −20°C. N-acetylcysteine (NAC) and
glutathione disulfide (GSSG) were purchased from Beyotime Institute
of Biotechnology. NAC (100 mM) and GSSG (10 mM) in PBS stock
solutions were stored at −20°C. Antibodies against poly
(ADP-ribose) polymerase (PARP; cat. no. 9542), caspase-3 (cat. no.
9665), caspase-9 (cat. no. 9502), B-cell lymphoma 2 (Bcl-2; cat.
no. 2872), Bcl-2-associated X protein (Bax; cat. no. 2772),
cytochrome c (Cyto-C; cat. no. 11940) and P53 (cat. no.
9284p) were obtained from Cell Signaling Technology, Inc. (Danvers,
MA, USA). GAPDH (cat. no. ab37168) antibody was purchased from
Abcam (Cambridge, UK). Immunoglobulin G-horseradish peroxidase
(IgG-HRP; cat. no. 030181) was purchased from EarthOx Life Sciences
(Millbrae, CA, USA).
Cell viability assay
An MTT assay was used to measure cell viability.
Briefly, PCa cells, PC-3/LNCaP (4×103 cells/well) and
22RV1 (1×104 cells/well), were seeded in 96-well plates
overnight, and were then incubated with various concentrations of
MET and PTX at 37°C for 6, 12, 24, 48 and 72 h. MTT (0.5 mg/ml) was
added to each well. After 4 h of incubation, supernatants were
removed and 150 µl DMSO was added to each well as a solvent. Using
a Multiskan Ascent microplate photometer (EnSpire 2300 Multilabel
Reader; PerkinElmer, Inc., Waltham, MA, USA) absorbance was
measured at 492 nm. DMSO-treated cells (control group) were
regarded as having 100% viability.
Apoptosis assay
Apoptosis was measured using the Apoptosis Detection
kit (BD Pharmingen; BD Biosciences, Franklin Lakes, NJ, USA). Cells
(1×105 cells/well) plated in 6-cm dishes were treated
with MET (5 mM) and PTX (10 nM for PC-3 cells, and 2 µM for 22RV1
and LNCaP cells). After 24 h of treatment, cells were washed with
PBS and harvested. The apoptosis assay was performed according to
the manufacturer's protocol using flow cytometry and the results
were analyzed using BD FACSDiva 6.1 software (BD Biosciences).
Caspase-Glo 3/7 assays
PC-3 cells, LNCaP cells (4×103
cells/well) and 22RV1 cells (1×104 cells/well) were
seeded in 96-well plates and exposed to MET and PTX. Equal volumes
(100 µl) RPMI-1640 medium and caspase-Glo 3/7 reagent (Promega
Corporation, Madison, WI, USA) were added to each well, and the
cells were incubated for 30 min at room temperature in the dark.
Luminescence was measured by a luminometer (Berthold Sirius L;
Titertek-Berthold, Pforzheim, Germany).
Reactive oxygen species (ROS)
detection
Cells were pretreated with an antioxidant, NAC (100
µM), or a pro-oxidant, GSSG (5 µM) for 24 h prior to the addition
of MET. Following treatment with MET (5 mM) and PTX (10 nM for PC-3
cells, and 2 µM for 22RV1 and LNCaP cells) for 12 h, the Reactive
Oxygen Species Assay kit (Beyotime Institute of Biotechnology) was
used. Cells (1×105 cells/well) were collected and
resuspended in serum-free medium containing DCFH-DA (10 µM). Then
cells were incubated at 37°C for 20 min in the dark. ROS levels
were measured by flow cytometry and the results were analyzed using
BD FACSDiva 6.1 software.
Mitochondrial membrane potential
Cells (1×105 cells/well) were seeded into
6-well plates overnight, and were subsequently treated with MET (5
mM) and PTX (10 nM for PC-3 cells, and 2 µM for 22RV1 and LNCaP
cells) for 12 h. According to the Mitochondrial Membrane Potential
Assay kit (Beyotime Institute of Biotechnology) manufacturer's
protocol, the cells were dyed with JC-1 staining fluid and analyzed
by flow cytometry and the results were analyzed using BD FACSDiva
6.1 software.
ATP levels
The ATP Assay kit (Beyotime Institute of
Biotechnology) was used for detection of ATP levels. Cells
(1×105 cells/well) were treated with MET (5 mM) and PTX
(10 nM for PC-3 cells, and 2 µM for 22RV1 and LNCaP cells) for 12
h. Subsequently, cells were treated with 200 µl lysis buffer
(Beyotime Institute of Biotechnology) and collected by
centrifugation at 12,000 × g for 10 min at 4°C. Subsequently, 50 µl
supernatant and 100 µl ATP detection reagent were mixed. Firefly
luciferase activity was measured using a luminometer (Berthold
Sirius L).
Western blot analysis
Cells were lysed with radioimmunoprecipitation assay
buffer (Beyotime Institute of Biotechnology) following MET and PTX
treatment for 24 h. Total protein was extracted at 4°C and
concentration was determined using a bicinchoninic acid assay.
Proteins (30 µg protein) were separated by 10% SDS-PAGE and
transferred to a polyvinylidene difluoride membrane (EMD Millipore,
Billerica, MA, USA) at 4°C. The membrane was blocked in 5% milk in
Tris-buffered saline with 1% Tween-20 at room temperature for 1 h,
and incubated with primary antibodies overnight at 4°C at the
following dilutions: PARP, 1:1,000; caspase-3, 1:1,000; caspase-9,
1:1,000; Bcl-2, 1:1,000; Bax, 1:1,000; Cyto-C, 1:1,000; P53,
1:1,000; and GAPDH, 1:100,000. The membranes were then probed with
IgG-HRP antibody (dilution, 1:10,000) for 1 h at room temperature.
Finally, the proteins were detected using Enhanced Chemiluminescent
kit (EMD Millipore).
Statistical analysis
The results are presented as the means ± standard
deviation. All experiments were performed in triplicate. Data
analysis was performed using a one-way analysis of variance using
and the least significant difference post hoc test was used to
determine statistical significance. SPSS 16.0 software (SPSS, Inc.,
Chicago, IL, USA) was used for statistical analysis, and figures
were generated using GraphPad Prism 6.0 software (GraphPad
Software, Inc., La Jolla, CA, USA). P<0.05, was considered to
indicate a statistically significant difference. ImageJ 2.0
software (National Institutes of Health, Bethesda, MD, USA) was
used to semi-quantify western blotting images.
Results
Effect of MET on viability of PCa
cells
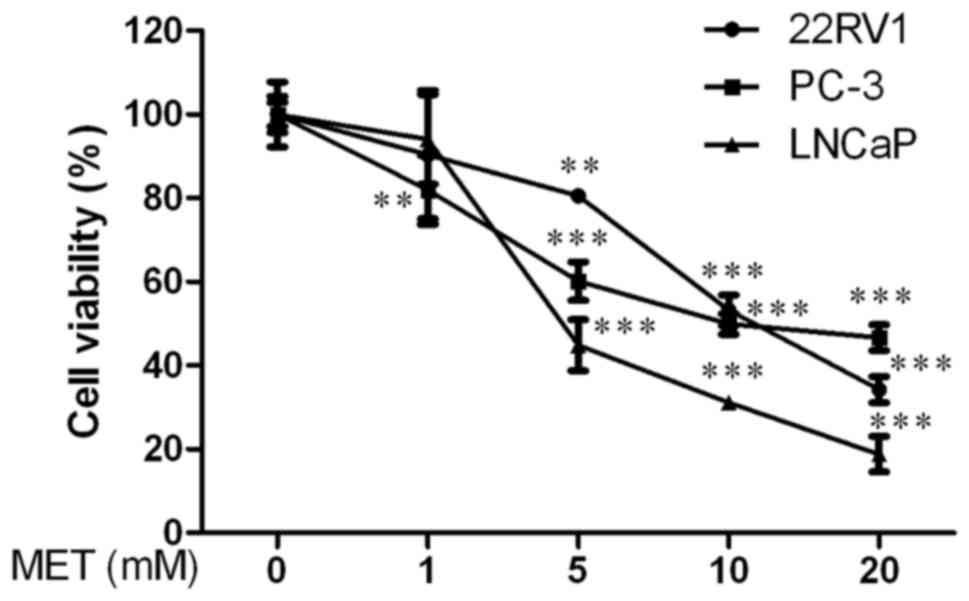
Firstly, the effect of MET on viability of PCa cells
was determined using an MTT assay. 22RV1, PC-3 and LNCaP cells were
treated with various concentrations of MET (0, 1, 5, 10 and 20 mM)
for 48 h. There was a clear reduction in cell viability starting at
5 mM compared with the control group in a dose-dependent manner
(P<0.01, Fig. 1). The half
maximal inhibitory concentration (IC50) value of MET was
12.281±1.809 mM for 22RV1 cells, 2.248±0.352 mM for PC-3 cells and
3.610±0.557 mM for LNCaP cells at 48 h. These data suggested that
MET inhibited the viability of PCa cells in a dose-dependent
manner.
Effects of MET in combination with PTX
on viability of PCa cells
To evaluate whether MET improved the
chemosensitivity of PTX, MET and PTX were administered together to
PCa cells (Fig. 2). The combination
of MET (5 mM) and PTX (1, 2, 5, 10 and 20 nM in PC-3 cells, and
0.5, 1, 2, 4 and 8 µM in 22RV1 and LNCaP cells) exhibited a greater
inhibitory effect on cell viability (Fig. 2A-C) than MET and PTX did
individually. Notably, MET decreased the IC50 of PTX in
PC-3 cells (Fig. 2G). These findings
suggested that MET inhibited PCa cell proliferation and improved
the chemosensitivity of PTX. The concentrations of MET (5 mM) and
PTX (10 nM for PC-3 cells, and 2 µM for 22RV1 and LNCaP cells) were
selected to verify that MET and PTX suppressed cell proliferation
in a time-dependent manner (Fig.
2D-F). Subsequently, MET and PTX-induced growth inhibition in
PCa cells was visualized by microscopy; the cells were treated with
MET (5 mM) and PTX (10 nM for PC-3 cells, and 2 µM for 22RV1 and
LNCaP cells). Cells cultured without these reagents exhibited
characteristic normal growth and shape after 24 h. However,
confluence was markedly reduced for cells treated with MET in
combination with PTX (Fig. 2H). This
finding suggested that MET improved the chemosensitivity of PTX.
MET in combination with PTX suppressed cell proliferation in a
time-dependent manner.
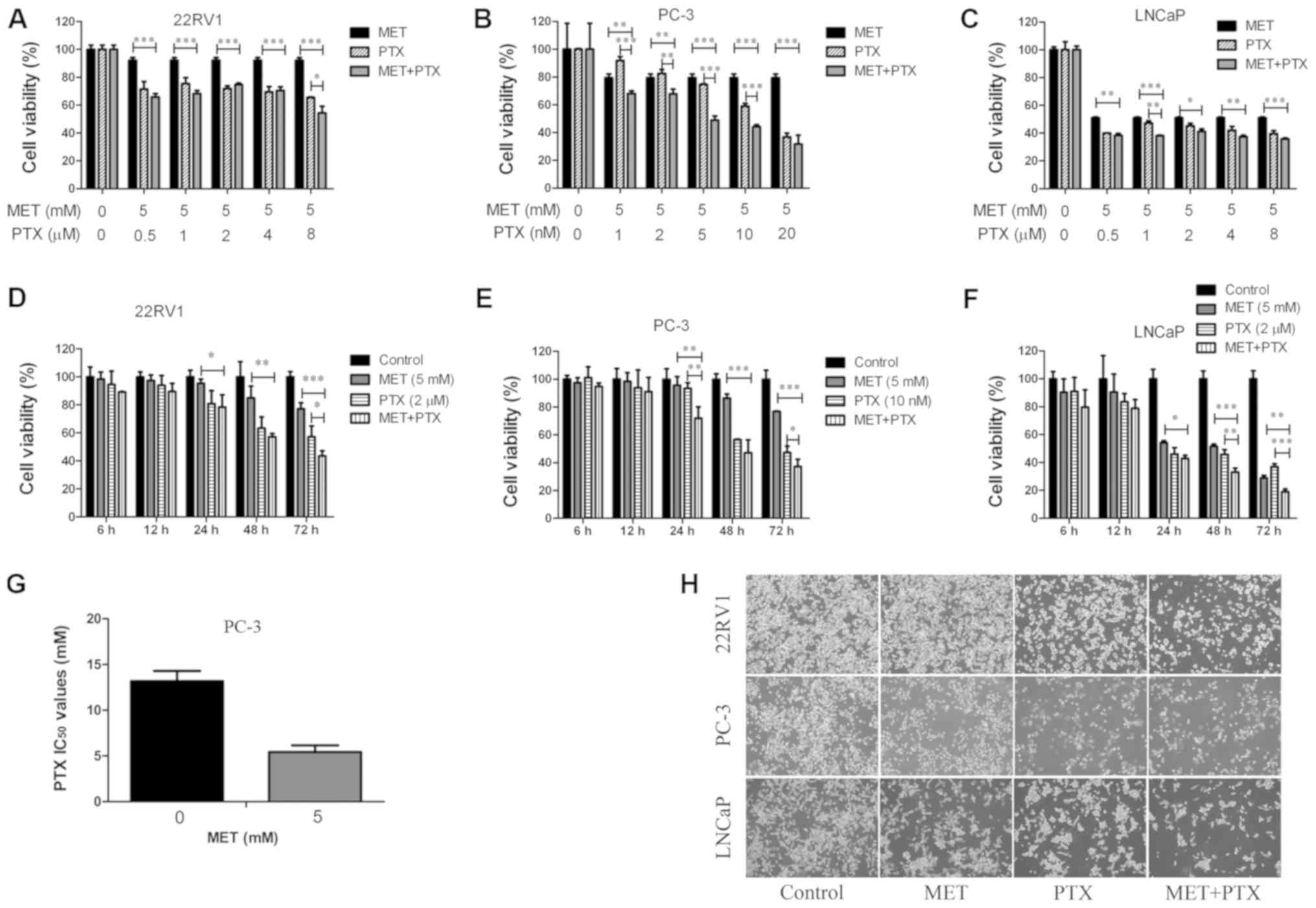 | Figure 2.MET in combination with PTX suppresses
cell proliferation. (A-C) Prostate cancer cells were treated with
MET (5 mM) and PTX (1, 2, 5, 10, 20 nM for PC-3 cells, and 0.5, 1,
2, 4, 8 µM for 22RV1 and LNCaP cells) for 48 h, and viability was
measured by MTT assay. (D-F) Cell viability was measured by MTT
assay following treatment with MET (5 mM) and PTX (10 nM for PC-3
cells, and 2 µM for 22RV1 and LNCaP cells) for 6, 12, 24, 48 and 72
h. (G) Changes in IC50 of PTX following MET treatment in
PC-3 cells. (H) 22RV1, PC-3 and LNCaP cells were treated with MET
(5 mM), PTX (10 nM for PC-3 cells, and 2 µM for 22RV1 and LNCaP
cells) and MET + PTX for 24 h. Images of cells were captured using
inverted contrast microscopy (magnification, ×100). Cells treated
with DMSO were used as the control group with cell viability set at
100%. *P<0.05, **P<0.01, ***P<0.001. DMSO, dimethyl
sulfoxide; IC50, half maximal inhibitory concentration;
MET, metformin; PTX, paclitaxel. |
MET in combination with PTX induces
apoptosis of PCa cells
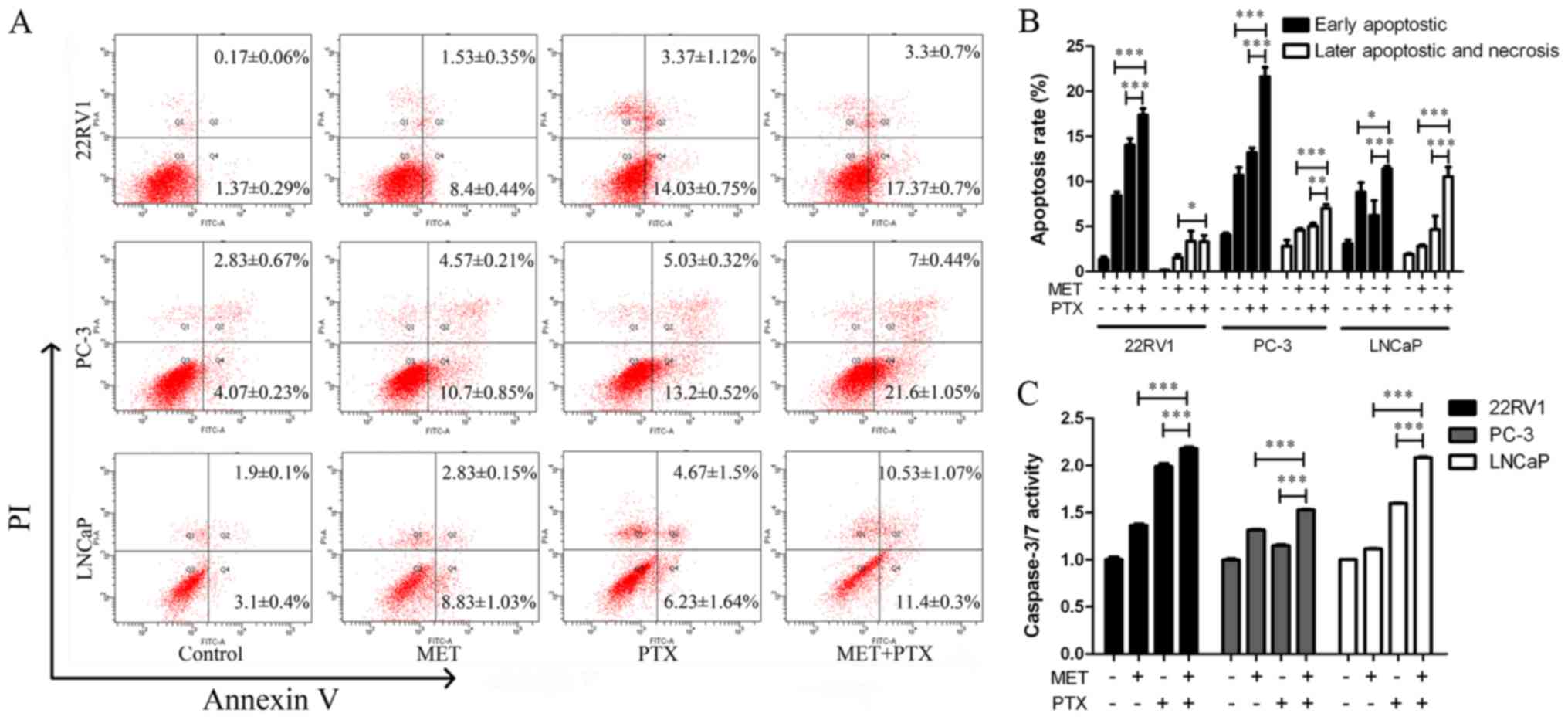
It was also determined whether the augmentation of
cell growth inhibition induced by MET in combination with PTX was
associated with an increase in apoptosis of PCa cells. The cells
were treated with MET (5 mM) and PTX (10 nM for PC-3 cells, and 2
µM for 22RV1 and LNCaP cells). After 24 h of treatment, cells were
labeled with Annexin V-fluorescein isothiocyanate/propidium iodide
(PI) and analyzed by flow cytometry. The apoptotic effect of MET +
PTX was much greater than in the single drug groups (Fig. 3A and B). This suggested that MET in
combination with PTX significantly induced early and late apoptosis
of PCa cells.
A caspase-3/7 activity assay was used, as shown in
Fig. 3C. MET in combination with PTX
markedly increased the activity of caspase-3/7. In addition, the
expression levels of apoptosis-associated proteins (PARP, P53,
Bcl-2, Bax, Cyto-C, caspase-3 and caspase-9) were measured using
western blotting. Caspase family members are key proteins in
apoptosis (21). MET in combination
with PTX markedly increased the expression levels of cleaved
caspase-3/9, Bax, P53, Cyto-C and PARP. However, it was identified
that treatment with MET and PTX significantly decreased Bcl-2
expression, compared with levels in the single drug groups
(Fig. 4A and B). As PC-3 cells in
the control group had undetectable expression of Clv-PARP and
Cyto-C, it was not possible to present quantification of the
increase in expression in relation to control expression in these
cells.
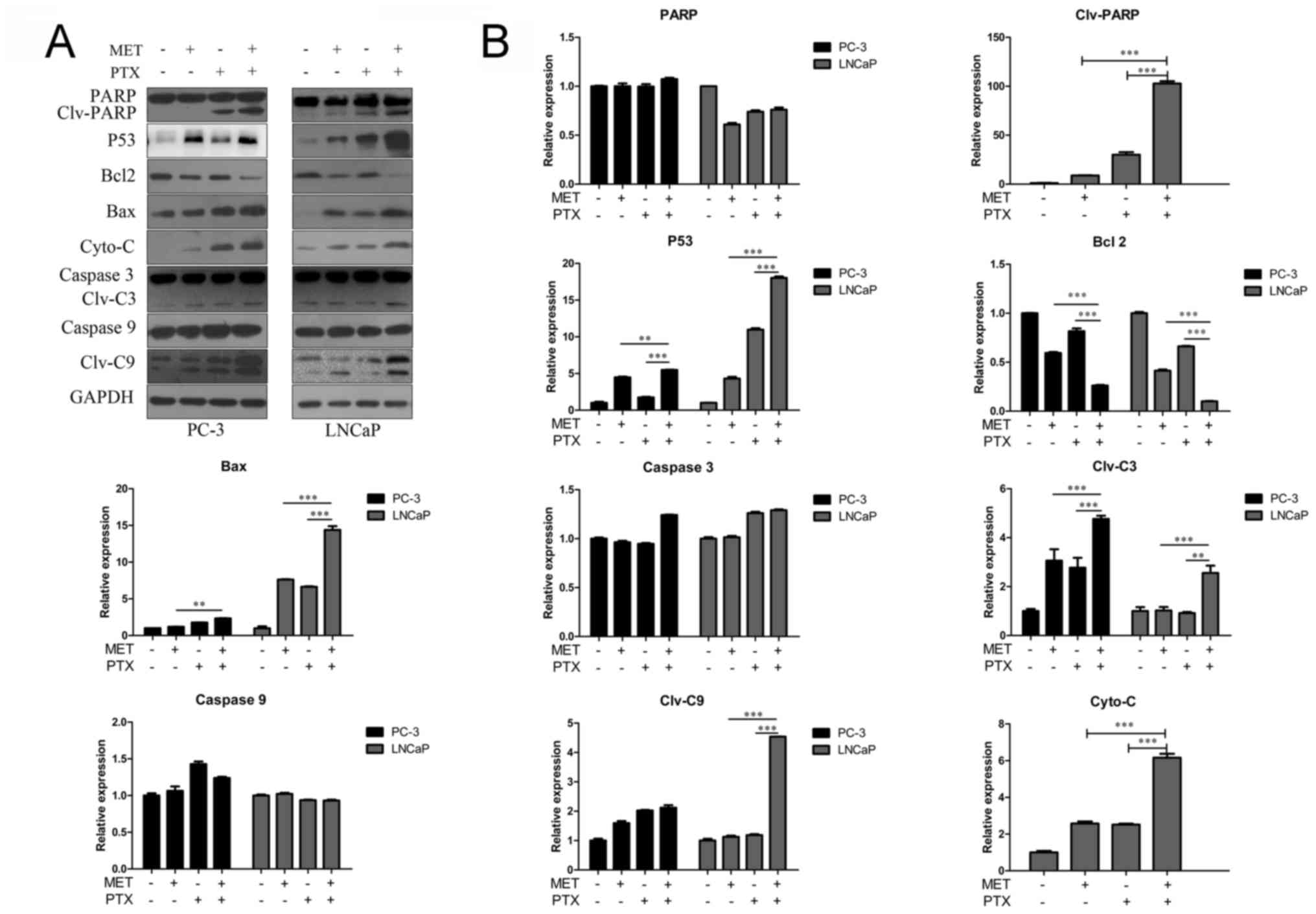 | Figure 4.Western blot analysis of
apoptosis-associated proteins in prostate cancer cells. Cells were
treated with MET (5 mM), PTX (10 nM for PC-3 cells, and 2 µM for
LNCaP cells) and MET + PTX for 24 h. (A) Western blot analysis was
used to detect the expression of apoptosis-associated proteins. (B)
Semi-quantification of western blotting. **P<0.01,
***P<0.001. Bax, Bcl-2-associated X protein; Bcl-2, B-cell
lymphoma 2; C3, caspase-3; C9, caspase-9; Clv, cleaved; Cyto-C,
cytochrome c; MET, metformin; PARP, poly (ADP-ribose)
polymerase; PTX, paclitaxel. |
MET induces growth suppression and
apoptosis of PCa cells via the production of ROS
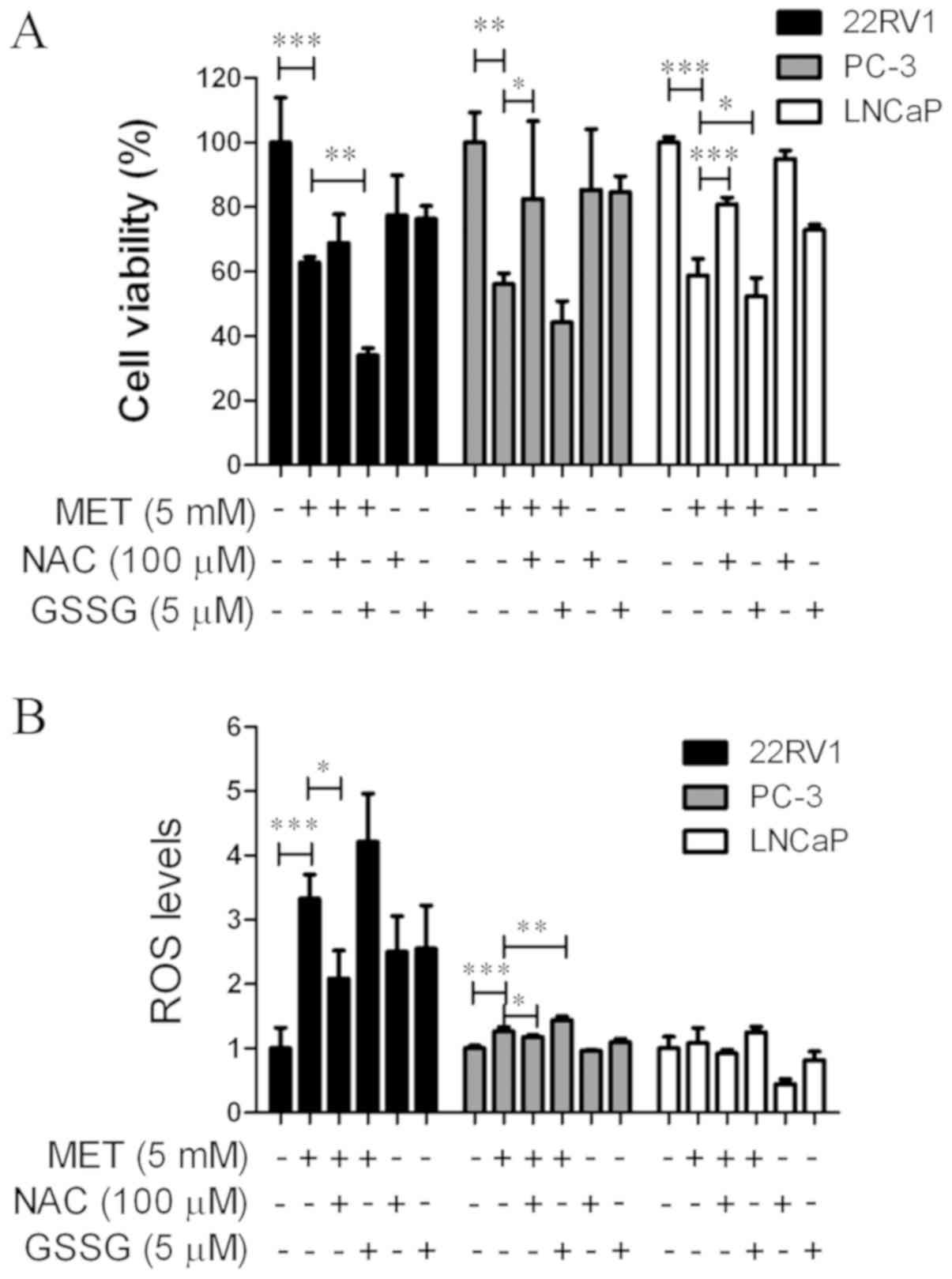
An MTT assay and a Reactive Oxygen Species Assay kit
were used to determine whether MET induced growth suppression and
apoptosis via elevation of intracellular ROS levels. Cells were
pretreated with an antioxidant, NAC (100 µM), or a prooxidant, GSSG
(5 µM), for 24 h prior to the addition of MET. NAC and GSSG in the
cells were then removed and MET was added for another 24 h. NAC
blocked MET-induced cell growth suppression (Fig. 5A) and increased ROS levels (Fig. 5B). GSSG augmented MET-induced cell
growth inhibition and promoted ROS production. This suggested that
MET inhibited the growth of PCa cells via the production of
ROS.
MET in combination with PTX suppresses
cell growth and induces apoptosis by increasing ROS production,
decreasing mitochondrial membrane potential and decreasing ATP
levels in PCa cells
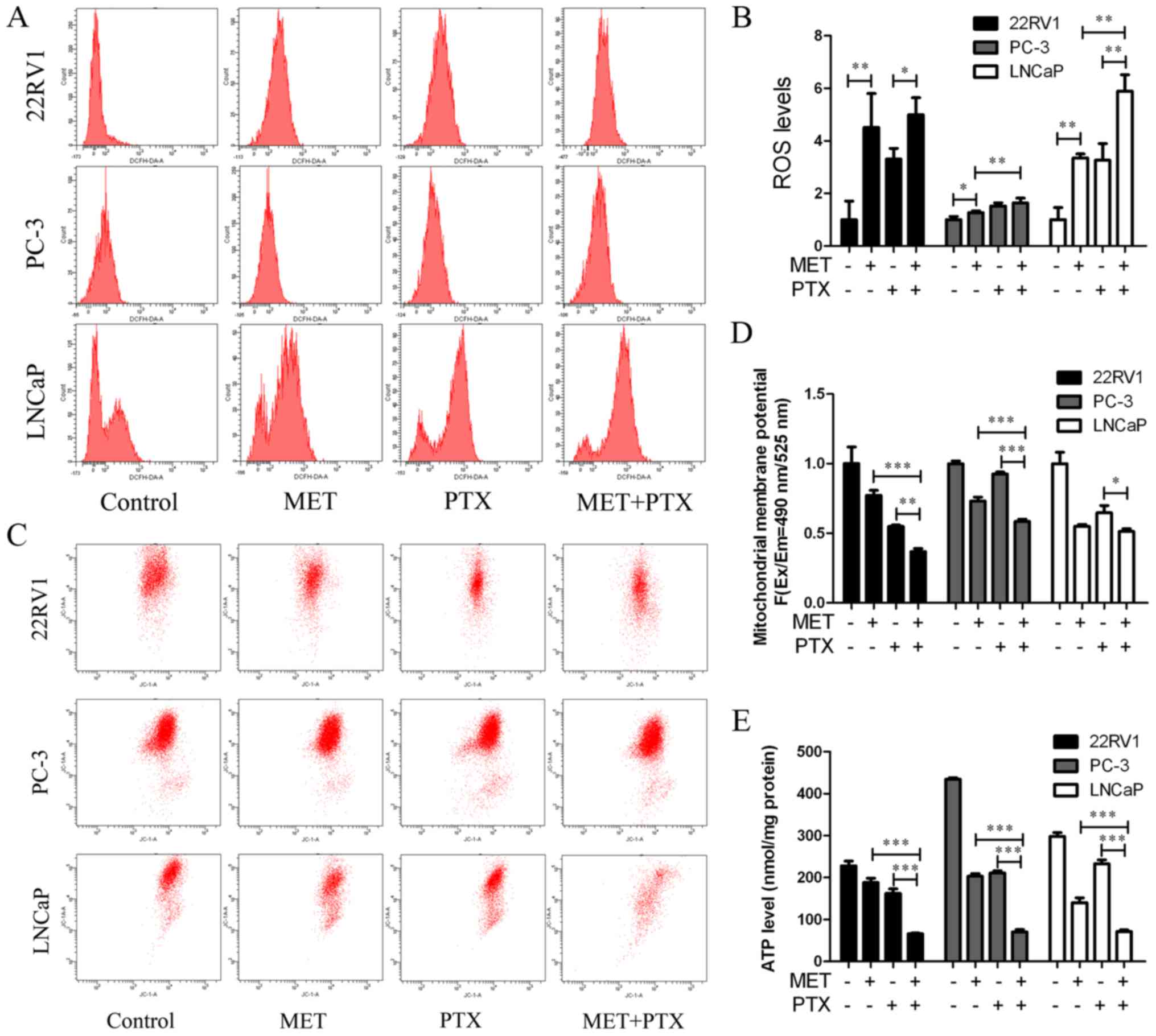
22RV1, PC-3 and LNCaP cells were treated with MET (5
mM) in combination with PTX (10 nM in PC-3 cells, 2 µM in 22RV1 and
LNCaP cells) for 12 h, and ROS production was measured. MET + PTX
significantly increased the production of ROS in LNCaP cells
(Fig. 6A and B). The imbalance of
ROS may promote mitochondrial dysfunction and lead to
mitochondria-mediated apoptosis. To evaluate the dysfunction in
mitochondrial energy production, the mitochondrial membrane
potential was measured. It is known that mitochondrial damage
during apoptosis alters the mitochondrial membrane potential and
intracellular levels of ATP in PCa cells. The present study
identified that the mitochondrial membrane potential (Fig. 6C and D) and intracellular levels of
ATP (Fig. 6E) were decreased by MET
+ PTX, and levels were significantly decreased in the MET + PTX
group compared with levels in the single drug groups. This
suggested that apoptosis of PCa cells mediated by MET + PTX was
associated with damage to the mitochondrial membrane.
Discussion
Chemotherapeutic regimens are commonly used to
inhibit tumor growth; nevertheless, these often have side effects.
Chemotherapy drugs not only have side effects but cancer cells also
often develop resistance to chemotherapeutic agents, limiting their
efficacy. Previously, MET was identified as an attractive
anticancer adjuvant drug combined with chemotherapeutic drugs,
which may improve treatment efficacy and lower the dose of
chemotherapeutic agents required.
In the present study, the antitumor activity of MET
+ PTX was evaluated in PCa cells. MET exhibited potential growth
inhibitory activity against PCa cells, as determined using the MTT
assay. MET and PTX exhibited enhanced ability to reduce tumor
proliferation and growth. It was demonstrated that PTX and MET, on
their own or in combination, exhibited anti-proliferative effects
against cultured PCa cell lines in a time- and dose-dependent
manner. However, there were differential sensitivities, in terms of
effectiveness of the treatment dosages, among the cell lines. LNCaP
is an early stage androgen-dependent PCa cell line. whereas 22RV1
and PC-3 are androgen-independent PCa cell lines. PC-3 cells have
no androgen receptor and exhibit moderate metastatic potential.
Therefore, the cell lines exhibit different sensitivity to
drugs.
A number of reports have suggested that
chemotherapeutic agents exert anti-proliferative effects by
inducing apoptosis. It was observed that MET-treated cells exhibit
reduced levels of ROS-mediated matrix membrane potential (22). Mitochondria serve a key role in ROS
production (23); the present data
suggested that MET + PTX induced apoptosis via increasing
intracellular ROS levels, and reducing mitochondrial membrane
potential and ATP. Notably, an increase in ROS serves a role in the
effect of MET + PTX on PCa cells. In the future, we aim to explore
whether antioxidant pretreatment can inhibit the effect of MET +
PTX on cell proliferation and apoptosis. In the present study,
antioxidant NAC and prooxidant GSSG were used to verify that MET
increased intracellular ROS levels in PCa cells. NAC pretreatment
led to a decrease in MET-mediated production of ROS in 22RV1 and
PC-3 compared to MET treatment alone, and had no effect in LNCaP
cells. GSSG had no effect on ROS levels in PC-3 and LNCaP cells
compared with MET group. In the present study, antioxidant NAC
attenuated and prooxidant GSSG increased the effect of MET on ROS
production in PCa cells. As is commonly known, the androgen
receptor serves an important role in the development of PCa. LNCaP
is an early stage androgen-dependent growth PCa cell line, PC-3
cells have no androgen receptor, and 22RV1 is an
androgen-independent growth PCa cell line. A previous study
demonstrated that physiological stimulation of the androgen
receptor increases ROS production (24). The androgen receptor may be an
important target to investigate differences among the three studied
cell lines concerning ROS levels in response to NAC and GSSG.
Therefore, we aim to further investigate whether the androgen
receptor is involved in the effect of antioxidant NAC and
prooxidant GSSG on ROS levels in PC-3, 22RV1 and LNCaP cells.
Increasing intracellular ROS, and reducing
mitochondrial membrane potential and ATP induces mitochondrial
damage via Cyto-C release from the mitochondria, which in turn
activates downstream caspase activity. Bcl-2 family proteins
(Bcl-2, Bax and Bcl-2 homologous antagonist killer) participate in
the apoptotic pathway leading to cell death. PTX sensitivity is
determined by the anti-apoptotic protein Bcl-extra large (25,26). PTX
downregulates Bcl-2 and activates caspases and PARP (27,28),
resulting in induction of apoptosis. Annexin V and PI staining and
analysis by flow cytometry demonstrated that MET + PTX induced PCa
cell apoptosis. Finally, the effects of MET + PTX on the expression
levels of various proteins, including caspase-3/9, Bax, Bcl2, PARP,
Cyto-C and P53, were studied by western blot analysis. MET +
PTX-treated cells exhibited decreased expression of Bcl-2 protein,
and increased expression of caspase-3/9, Bax, PARP, Cyto-C and P53
proteins. Taken together, these data suggested that MET + PTX
suppressed proliferation and induced apoptosis of human PCa cells
via ROS, promoting expression of the pro-apoptotic protein P53, and
inducing mitochondrial damage. P53 promoted expression of Bax,
inhibited expression of Bcl-2 and mitochondrial damage, and Bcl-2
promoted Cyto-C release from mitochondria. This resulted in the
activation of caspase-dependent apoptotic pathways (Fig. 7). A limitation of the present study
is that it was limited to in vitro data; therefore, an in
vivo study will be seriously considered in the future.
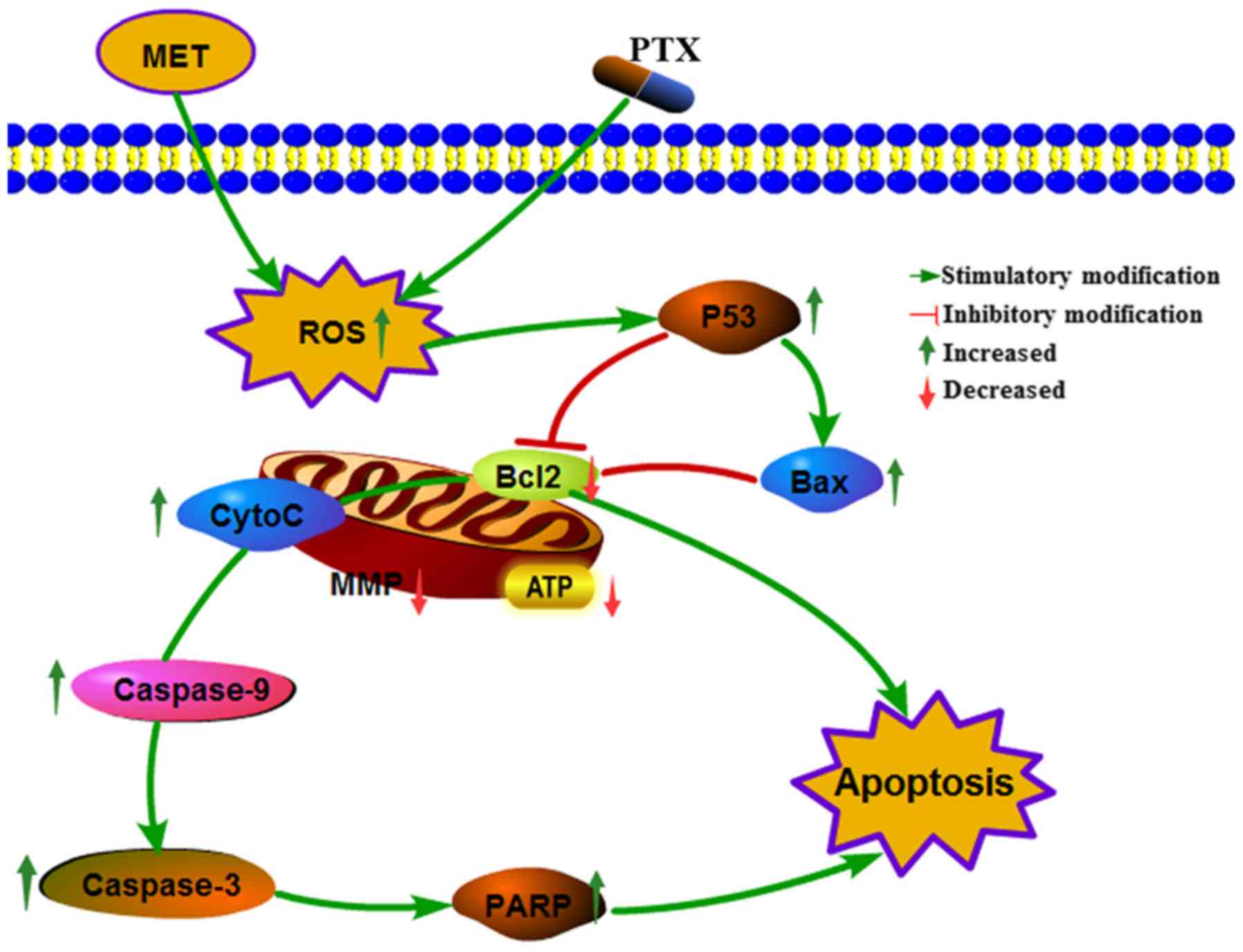 | Figure 7.Cellular pathway of the effects of MET
+ PTX-induced growth inhibition and apoptosis of PCa cells. MET +
PTX increased oxidative stress, and decreased MMP and ATP levels in
PCa cells. MET + PTX upregulated the production of P53, PARP,
capase3/9, Bax, Cyto-C, and downregulated the production of Bcl-2,
promoting the release of Cyto-C, increasing caspase-3/7 activities,
and potentiating apoptosis in PCa cells. Bax, Bcl-2-associated X
protein; Bcl-2, B-cell lymphoma 2; CytoC, cytochrome c; MET,
metformin; MMP, mitochondrial membrane potential; PARP, poly
(ADP-ribose) polymerase; PCa, prostate cancer; PTX, paclitaxel. |
In conclusion, this study demonstrated that MET
combined with PTX suppressed cell growth and induced apoptosis of
PCa cells via mitochondria-mediated apoptotic pathways. These
findings provide promising insights into novel, potential
therapeutic strategies for PCa.
Acknowledgements
Not applicable.
Funding
This study was funded by The National Natural
Science Funds of China (grant no. 81272833).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YZ and JL designed the experiments. YZ, XZ, HT and
DY performed the experiments. YZ and XZ participated in data and
statistical analyses. YZ and JL wrote the article and prepared
figures. JL provided the financial support. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MET
|
metformin
|
|
PTX
|
paclitaxel
|
|
NAC
|
N-acetylcysteine
|
|
GSSG
|
glutathione disulfide
|
|
ROS
|
reactive oxygen species
|
References
|
1
|
Yang Y and Wu XH: Study on the influence
of metformin on castration-resistant prostate cancer PC-3 cell line
biological behavior by its inhibition on PLCε gene-mediated
Notch1/Hes and androgen receptor signaling pathway. Eur Rev Med
Pharmacol Sci. 21:1918–1923. 2017.PubMed/NCBI
|
|
2
|
Crawford ED: Epidemiology of prostate
cancer. Urology. 62:3–12. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sandblom G and Varenhorst E: Incidence
rate and management of prostate carcinoma. Biomed Pharmacother.
55:135–143. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zitzmann S, Mier W, Schad A, Kinscherf R,
Askoxylakis V, Kramer S, Altmann A, Eisenhut M and Haberkorn U: A
new prostate carcinoma binding peptide (DUP-1) for tumor imaging
and therapy. Clin Cancer Res. 11:139–146. 2005.PubMed/NCBI
|
|
6
|
Chi K, Hotte SJ, Joshua AM, North S, Wyatt
AW, Collins LL and Saad F: Treatment of mCRPC in the
AR-axis-targeted therapy-resistant state. Ann Oncol. 26:2044–2056.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Weaver BA: How Taxol/paclitacel kills
cancer cells. Mol Biol Cell. 25:2677–2681. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sobue S, Mizutani N, Aoyama Y, Kawamoto Y,
Suzuki M, Nozawa Y, Ichihara M and Murate T: Mechanism of
paclitaxel resistance in a human prostate cancer cell line, PC3-PR,
and its sensitization by cabazitaxel. Biochem Biophys Res Commun.
479:808–813. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yang Y, Ma Y, Sheng J, Huang Y, Zhao Y,
Fang W, Hong S, Tian Y, Xue C and Zhang L: A multicenter,
retrospective epidemiologic survey of the clinical features and
management of bone metastatic disease in China. Chin J Cancer.
35:402016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Hatoum D and McGowan EM: Recent advances
in the use of metformin: Can treating diabetes prevent breast
cancer? Biomed Res Int. 2015:5484362015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dowling RJ, Goodwin PJ and Stambolic V:
Understanding the benefit of metformin use in cancer treatment. BMC
Med. 9:332011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gou S, Cui P, Li X, Shi P, Liu T and Wang
C: Low concentrations of metformin selectively inhibit
CD133+ cell proliferation in pancreatic cancer and have
anticancer action. PLoS One. 8:e639692013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Noto H, Goto A, Tsujimoto T and Noda M:
Cancer risk in diabetic patients treated with metformin: A
systematic review and meta-analysis. PLoS One. 7:e334112012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Pollak MN: Investigating metformin for
cancer prevention and treatment: The end of the beginning. Cancer
Discov. 2:778–790. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Del Barco S, Vazquez-Martin A, Cufí S,
Oliveras-Ferraros C, Bosch-Barrera J, Joven J, Martin-Castillo B
and Menendez JA: Metformin: Multi-faceted protection against
cancer. Oncotarget. 2:896–917. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zakikhani M, Dowling R, Fantus IG,
Sonenberg N and Pollak M: metformin Is an AMP kinase-dependent
growth inhibitor for breast cancer cells. Cancer Res.
66:10269–10273. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Goodwin PJ, Ligibel JA and Stambolic V:
Metformin in breast cancer: Time for action. J Clin Oncol.
27:3271–3273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wang LW, Li ZS, Zou DW, Jin ZD, Gao J and
Xu GM: metformin induces apoptosis of pancreatic cancer cells.
World J Gastroenterol. 14:7192–7198. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shank JJ, Yang K, Ghannam J, Cabrera L,
Johnston CJ, Reynolds RK and Buckanovich RJ: Metformin targets
ovarian cancer stem cells in vitro and in vivo. Gynecol Oncol.
127:390–397. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Iliopoulos D, Hirsch HA and Struhl K:
Metformin decreases the dose of chemotherapy for prolonging tumor
remission in mouse xenografts involving multiple cancer cell types.
Cancer Res. 71:3196–3201. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Abedi H, Aghaei M, Panjehpour M and
Hajiahmadi S: Mitochondrial and caspase pathways are involved in
the induction of apoptosis by IB-MECA in ovarian cancer cell lines.
Tumor Biol. 35:11027–11039. 2014. View Article : Google Scholar
|
|
22
|
Cheng G and Lanza-Jacoby S: Metformin
decreases growth of pancreatic cancer cells by decreasing reactive
oxygen species: Role of NOX4. Biochem Biophys Res Commun.
465:41–46. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ding H, Han C, Guo D, Chin YW, Ding Y,
Kinghorn AD and D'Ambrosio SM: Selective induction of apoptosis of
human oral cancer cell lines by avocado extracts via a ROS-mediated
mechanism. Nutr Cancer. 61:348–356. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Khandrika L, Kumar B, Koul S, Maroni P and
Koul HK: Role of Oxidative Stress in Prostate Cancer. Cancer Lett.
282:125–136. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Harley ME, Allan LA, Sanderson HS and
Clarke PR: Phosphorylation of Mcl-1 by CDK1-cyclin B1 initiates its
Cdc20-dependent destruction during mitotic arrest. EMBO J.
29:2407–2420. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Castilla C, Flores ML, Medina R,
Perez-Valderrama B, Romero F, Tortolero M, Japón MA and Sáez C:
Prostate cancer cell response to paclitaxel is affected by
abnormally expressed securin PTTG1. Mol Cancer Ther. 13:2372–2383.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tudor G, Aguilera A, Halverson DO, Laing
ND and Sausville EA: Susceptibility to drug-induced apoptosis
correlates with differential modulation of Bad, Bcl-2 and Bcl-xL
protein levels. Cell Death Differ. 7:574–586. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lim SJ, Choi MK, Kim MJ and Kim JK:
Alpha-tocopheryl succinate potentiates the paclitaxel-induced
apoptosis through enforced caspase 8 activation in human H460 lung
cancer cells. Exp Mol Med. 41:737–745. 2009. View Article : Google Scholar : PubMed/NCBI
|