Introduction
Cervical cancer is the second most prevalent
malignant tumor in women worldwide and the fourth leading cause of
cancer-associated mortalities among women (1,2).
Surgery, chemotherapy and radiotherapy are the standard treatment
options for patients with cervical cancer; however, the 5 year
survival rate for advanced and recurrent cervical cancer is only
10–20%; with metastasis and chemotherapeutic agent resistance being
the principal causes of treatment failure (3). Cis-diamminedichloroplatinum (II) (more
commonly referred to as cisplatin) is a small-molecular
platinum-based compound that appears to be the most effective
anti-tumor agent in patients with advanced and recurrent cervical
cancer (4). However, either
intrinsic or acquired resistance to cisplatin may develop,
seriously compromising the efficacy of cisplatin (5). Previous studies investigating the
mechanisms of cisplatin resistance have identified a number of
complex resistance mechanisms (6,7). As a
natural defense mechanism developed to protect cells from
environmental toxins, cisplatin resistance requires numerous
epigenetic and genetic alterations that support cell survival,
including cell growth, apoptosis, developmental pathways, DNA
damage repair and cellular accumulation of cisplatin (8).
Tumor necrosis factor α-induced protein 8 (TNFAIP8
or TIPE; additionally termed SCC-S2, NDED or GG2-1) serves as an
anti-apoptotic and pro-oncogenic signaling molecule (9–12). A
previous study suggested that tumor necrosis factor α (TNF-α)
stimulation and the activation of the nuclear factor-κB pathway may
upregulate the mRNA expression levels of TNFAIP8 in head and neck
squamous cell carcinoma cell lines (13). In addition, TNFAIP8 overexpression
was associated with cancer progression and poor prognosis in a
number of different cancer types (12,14–18).
Silencing TNFAIP8 in a variety of cells promoted cellular
apoptosis, attenuated cellular proliferation and the production of
matrix metalloproteinase (MMP)-1 and MMP-9 (19). However, the physiological and
pathophysiological roles of TNFAIP8 in cervical carcinogenesis and
development remain poorly understood. It is unclear how the
expression of TNFAIP8 in cervical cancer promotes cisplatin
resistance.
In the present study, TNFAIP8 expression in cervical
cancer tissues compared with matching adjacent tissues was examined
and it was demonstrated that increased TNFAIP8 was associated with
cisplatin resistance. Additionally, the effect of TNFAIP8
inhibition using short hairpin (sh)RNA on cisplatin-induced
cellular apoptosis in vitro was examined. Subsequent
examination of the potential underlying mechanisms suggested that
TNFAIP8 promoted cisplatin resistance by inhibiting cellular
apoptosis.
Materials and methods
Patient tissue samples
In total, 40 pairs of cervical cancer tissues in
addition to the corresponding adjacent tissues were collected from
40 female patients (39–67 years old) who underwent surgical
resection at the Affiliated Huaihe Hospital of Henan University
(Kaifeng, China) between April 2012 and December 2012. These
patients were pathologically diagnosed with cervical cancer by two
pathologists. All patients had no metastatic tumors, no serious
complications and no other malignant tumors. Prior to cervical
resection, none of the patients had received radiotherapy or
chemotherapy. All patients received cisplatin-based chemotherapy
following surgery. Patients were identified as cisplatin-sensitive
if no neoplasm was found by imaging within 12 months of
chemotherapy, or as cisplatin-resistant if neoplasm was found. The
Committee for Ethical Review at Henan University School of Medicine
(Kaifeng, China) approved the protocol, and written informed
consent was provided by all patients.
Immunohistochemistry
Cervical cancer specimens and adjacent tissues were
fixed with 4% paraformaldehyde overnight at room temperature, and
embedded in paraffin and sectioned at a thickness of 4 µm in the
Department of Pathology, Affiliated Huaihe Hospital of Henan
University. All sectioning was performed using standardized
methods. Sections were deparaffinized in xylene twice for 10 min,
rehydrated in gradient ethanol (100, 90, 80, 70 and 60%) once for 2
min at room temperature and subjected to heat-induced antigen
retrieval and elimination of endogenous peroxidases by boiling in a
water bath for 10 min. Subsequently, sections were blocked with 10%
goat serum (Wuhan Boster Biological Technology, Ltd., Wuhan, China)
at room temperature for 15 min to prevent non-specific adsorption
and incubated with a primary antibody against TNFAIP8 (1:100;
ab195810; Abcam, Cambridge, MA, USA) at 4°C overnight. Sections
were subsequently incubated with a horseradish
peroxidase-conjugated secondary goat anti-rabbit antibody (1:500;
BA1056; Wuhan Boster Biological Technology, Ltd.) for 1.5 h at room
temperature. Subsequently, the samples were stained with
diaminobenzidine for 5 min and counterstained with hematoxylin
(Beyotime Institute of Biotechnology, Haimen, China) for 2 min at
room temperature, and sealed with neutral gum. TNFAIP8 staining was
assessed as previously described (20) with specific modifications. All slides
were independently analyzed with a light microscope (magnification
×100) by two pathologists in a blinded manner and scored based on
staining intensity as follows: i) 0, No staining; ii) 1, weak
staining; iii) 2, moderate staining; and iv) 3, strong staining. If
there were discrepancies between the two pathologists, the final
decision was made by another pathologist.
Cell culture
The human cervical cancer cell line (HeLa) was
purchased from The Cell Bank of Type Culture Collection of Chinese
Academy of Sciences (Shanghai, China). HeLa cells were cultured in
Dulbecco's modified Eagle's medium (DMEM) (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing certified 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin and 100 µg/ml streptomycin (Beijing Solarbio Science
& Technology Co., Ltd., Beijing, China). A TNFAIP8-silenced
HeLa cell line was established using lentiviral transfection using
a pGLV-U6-Puro vector carrying TNFAIP8 shRNA (Shanghai GenePharma
Co., Ltd., Shanghai, China). Briefly, infectious lentiviral vectors
were harvested form HEK293T cells co-transfected with the
recombinant competent virions (pGLV-U6-shTNFAIP8) and helper
plasmids (pGag/Pol, pRev and pVSV-G) using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) according to the manufacturer's protocol. HeLa
cells were transfected with 109 transducing units/ml of
lentiviruses in fresh transduction medium supplemented with 8 µg/ml
Polybrene (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany). Cells
were cultured in complete medium containing puromycin (2 µg/ml) for
at least 2 weeks prior to being used for experiments. TNFAIP8
expression was determined using both RT-qPCR and western blotting
post-transduction. All cells were cultured in a humidified
incubator containing 5% CO2 in compressed air at
37°C.
Reverse transcription quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from HeLa cells using a
RNAiso Plus extraction kit and reverse-transcribed into cDNA at
42°C for 1 h using the PrimeScript RT reagent kit (Takara
Biotechnology Co., Ltd., Dalian, China). qPCR was performed using
GoTaq qPCR Master Mix (Promega Corporation, Madison, WI, USA). For
detection of TNFAIP8 mRNA expression levels, GAPDH was amplified in
parallel as an internal control. The following primers were used
for qPCR: TNFAIP8 forward, 5′-TCCATCGCCACCACCTTA-3′ and reverse,
5′-CTCTGCCTCCTTCTTGTTTT-3′; GAPDH forward,
5′-GGCAAATTCAACGGCACAGTCA-3′ and reverse,
5′-GTCTCGCTCCTGGAAGATGGTGAT-3′. The qPCR conditions were
denaturation at 95°C for 10 min, followed by 40 cycles of
denaturation at 95°C for 15 sec, annealing at 60°C for 30 sec and
extension at 55°C for 1 min. The gene expression level was
calculated using the 2−∆∆Cq method (21). The expression level in control cells
was considered as 1.
Plate colony-formation assay
TNFAIP8-silenced and control cells were seeded in a
6-well plate with ~500 cells in 2 ml complete culture medium/well.
Cisplatin (5 µg/ml) was added to the medium after 2 days, and the
cells were cultured at 37°C for ~2 weeks. The medium was discarded
and the cells were washed gently with PBS when the clones were
visible. Cells were fixed with 100% methanol for 15 min and stained
with crystal violet for 20 min at room temperature. The clones were
scanned and counted using Image-Pro Plus 6.0 (Media Cybernetics,
Inc., Rockville, MD, USA).
MTS assay
Cell viability was measured with an MTS kit (G5421;
Promega Corporation) according to the manufacturer's protocol.
Cells were seeded at 2×103 cells/well in a 96-well plate
and allowed to attach overnight. Different concentrations (0, 2.5,
5, 10, 20 or 40 µg/ml) of cisplatin were added to the medium for 24
h, or 5 µg/ml cisplatin was added to the medium for 6, 12, 24, 48
or 72 h. Each group contained six well replicates. A total of 20 µl
MTS aqueous solution was added to each well and the cells were
incubated at 37°C for 2 h. Optical density values were measured at
490 nm on a full wavelength multi-function microplate reader
(Thermo Fisher Scientific, Inc.).
Apoptosis analysis
Cells were seeded at 1×105 cells into
each well of a 6-well plate with 2 ml complete culture medium. The
cells were left to attach overnight. Subsequently, cisplatin was
added at a concentration of 5 or 10 µg/ml for 24 h. Cells were
collected and washed twice with DMEM, and apoptotic cells were
stained with Annexin V-fluorescein isothiocyanate and propidium
iodide (Miltenyi Biotec, Inc., Cambridge, MA, USA) according to the
manufacturer's protocol. The cells were detected using a flow
cytometer (FACSCalibur) and analyzed with FlowJo software (version
10; FlowJo LLC, Ashland, OR, USA).
Trypan blue staining
A total of 1×105 cells/well were seeded
into a 6-well plate. Subsequent to cells attaching overnight, the
medium was replaced with fresh medium with or without cisplatin (5
and 10 µg/ml) and incubated for a further 24 h. Cells were
collected and the viability was determined by trypan blue staining
at room temperature for 2 min. Cell numbers were counted manually
using a hemocytometer, and the cell death ratio was calculated as
the proportion of trypan blue-positive cells to the total number of
cells.
Western blot analysis
Cells were harvested, and total protein was
extracted using an appropriate volume of lysis buffer (P0013;
Beyotime Institute of Biotechnology). Supernatants were collected
following centrifugation at 13,000 × g for 10 min at 4°C. The
protein concentration was determined using a bicinchoninic acid
protein assay kit. Equal quantities of 40 µg protein samples were
loaded onto 10% gels for SDS-PAGE. Proteins were subsequently
transferred to polyvinylidene fluoride membranes. Subsequent to
blocking with 5% skimmed milk at room temperature for 1 h, the
membranes were incubated with the following primary antibodies at
4°C overnight: TNFAIP8 (1:1,000; ab195810; Abcam); β-actin
(1:20,000; a1978; Sigma-Aldrich; Merck KGaA); procaspase-3 and
cleaved caspase3 (9662), cleaved caspase-8 (9496),
phosphorylated-p38 (9211), p-38 (9212) and Bcl-2 (4223) at 1:1,000
(Cell Signaling Technology, Inc., Danvers, MA, USA). Blot were then
washed, and incubated with HRP-conjugated secondary antibody
(1:500; BA1056; Wuhan Boster Biological Technology, Ltd.) at room
temperature for 2 h. Protein bands were visualized using an
enhanced chemiluminescence system (Thermo Fisher Scientific,
Inc.).
Statistical analysis
Data are presented as the mean ± standard deviation
of at least three independent experiments. The data was analyzed
using a Student's t-test or one-way analysis of variance followed
by Bonferroni correction using GraphPad Prism 6 (GraphPad Software,
Inc., La Jolla, CA, USA). P<0.05 was considered to indicate a
statistically significant difference.
Results
TNFAIP8 overexpression in cervical
cancer tissues is associated with cisplatin resistance
A total of 40 pairs of cervical cancer tissues and
non-tumor adjacent tissues were collected for immunohistochemical
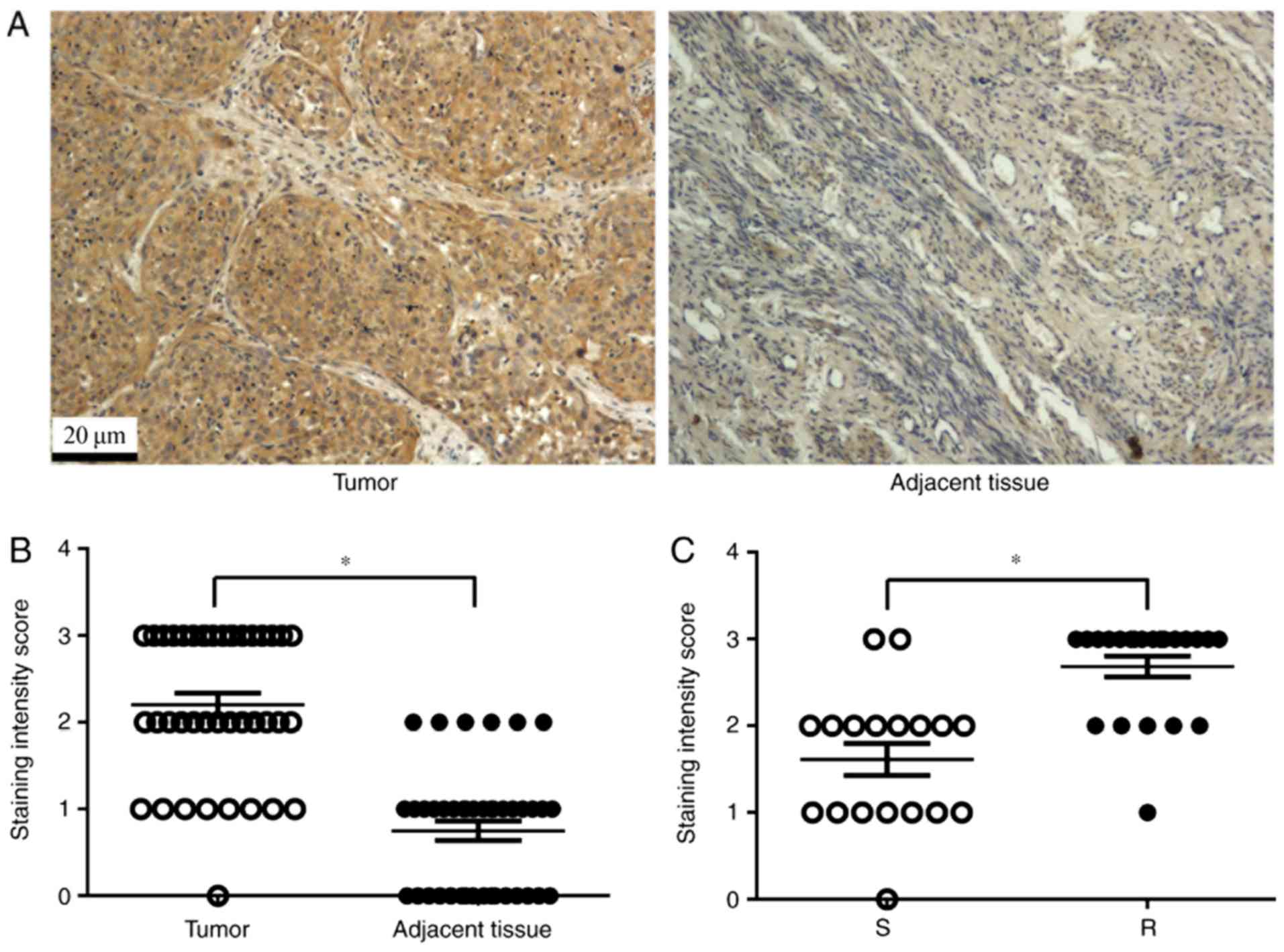
analysis of TNFAIP8 expression. As expected, TNFAIP8 was highly
expressed in cervical cancer tissues, whereas, little or no
staining was observed in the tissues adjacent to the tumor
(Fig. 1A and B). The patients with
high TNFAIP8 expression (18 cases) were significantly more likely
to develop cisplatin resistance (16 cases of the total 22 cases)
compared with patients with low TNFAIP8 expression (Fig. 1C; P<0.05). These data suggested
that TNFAIP8 is an oncogene, which may mediate therapeutic
resistance to cisplatin.
TNFAIP8 increases the proliferation of
cervical cancer cells
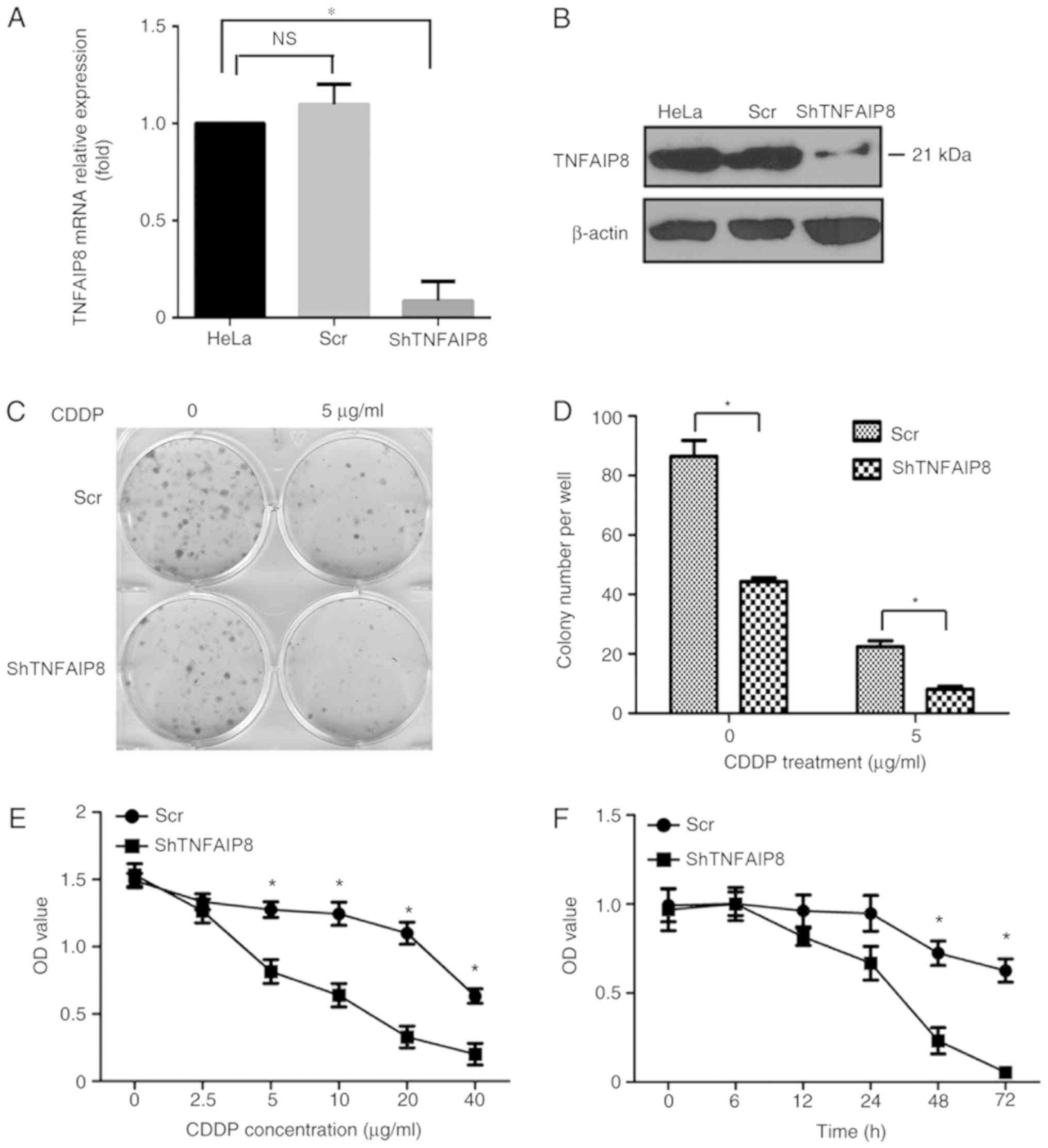
To elucidate the function of TNFAIP8 in the growth
of cervical cancer cells, HeLa cells were transfected with the
plasmid pGLV-shTNFAIP8 or a control vector to generate
TNFAIP8-knockdown and control HeLa cell lines (Fig. 2A and B). Equivalent numbers of each
cell line were seeded into different wells of a flat plate to
evaluate colony formation. After ~2 weeks, visible clones were
stained with crystal violet, and the number of clones was counted
and analyzed. A significant decrease in the proliferation rate of
HeLa cells with decreased expression of TNFAIP8 was observed
(P<0.05). In particular, fewer and smaller clones formed with
TNFAIP8 knockdown cells compared with the control HeLa cells
following treatment with cisplatin (Fig.
2C and D).
Knockdown of TNFAIP8 enhances the
cytotoxicity of cisplatin in HeLa cells
To determine whether TNFAIP8 protein expression in
cervical cancer was associated with cisplatin resistance, an MTS
assay was used to measure cell viability. As approximately all
cells in the shRNA group and control group died within 48 h
following treatment with 10 mg/ml cisplatin (data not shown), cells
were instead treated with 5 mg/ml cisplatin for 6, 12, 24, 48 or 72
h. Only a small number of live HeLa cells were observed in the
TNFAIP8 shRNA group, compared with the numerous cells that were
still viable in the control group after cisplatin treatment for 72
h (Fig. 2F). Treatment with
cisplatin significantly decreased the HeLa cell viability in a dose
and time dependent manner as demonstrated in Fig. 2E and F, respectively.
Knockdown of TNFAIP8 promotes cellular
apoptosis and death
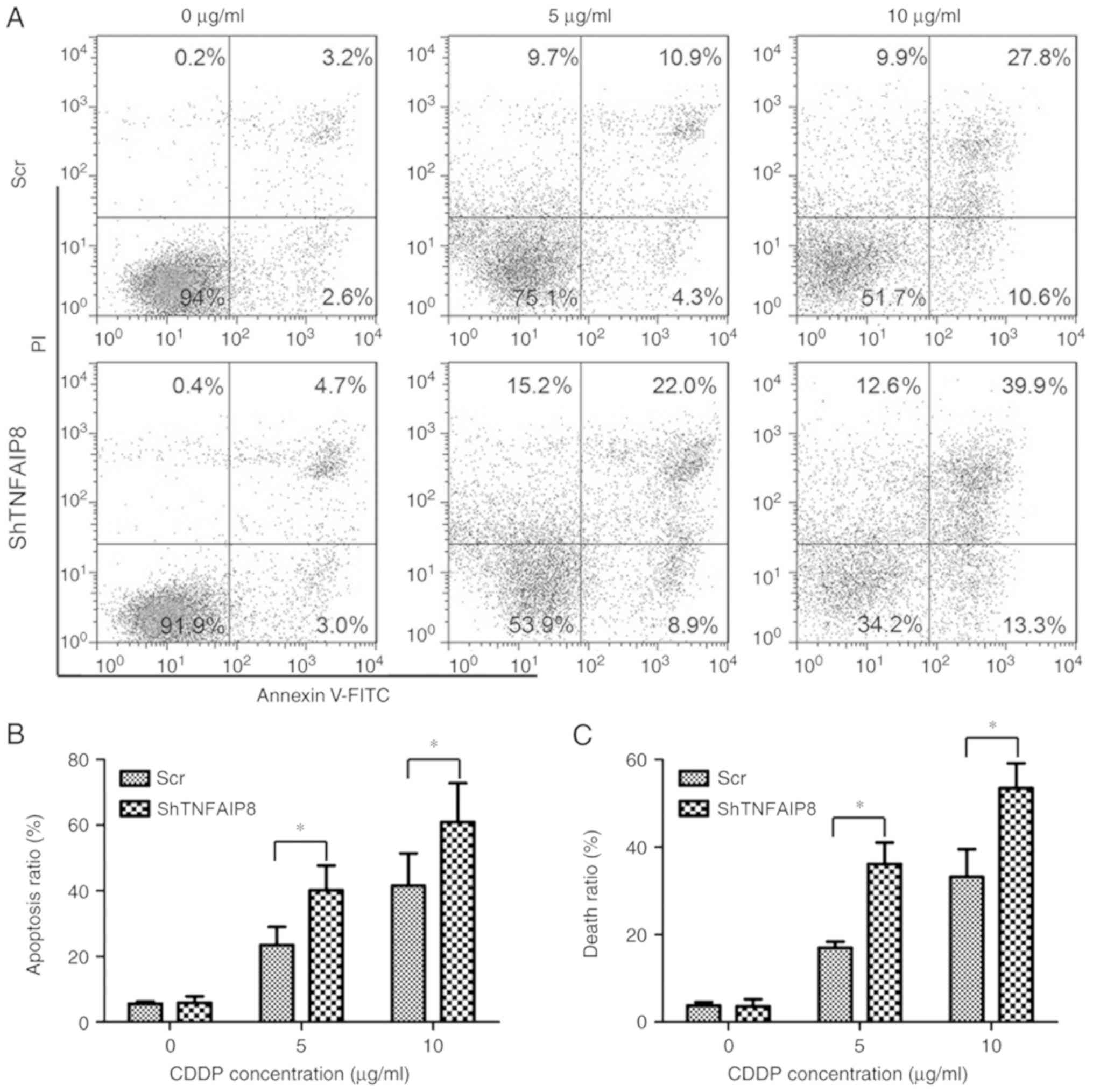
The effect of TNFAIP8 knockdown on apoptosis and
death in HeLa cells was examined. Following treatment with
cisplatin at the stated concentrations for 24 h, cellular apoptosis
was detected using flow cytometry. The percentage of apoptotic HeLa
cells in the TNFAIP8-knockdown group was significantly increased
compared with the control group following treatment with cisplatin
(Fig. 3A and B; P<0.05).
Additionally, the level of cell death, as analyzed by trypan blue
staining, demonstrated results similar to those for cellular
apoptosis (Fig. 3C). Collectively,
the present data demonstrated that silencing TNFAIP8 resulted in an
increase in the cytotoxicity of cisplatin in HeLa cells, suggesting
that TNFAIP8 confers resistance to cisplatin in cervical
cancer.
TNFAIP8 promotes cisplatin resistance
by inhibiting cellular apoptosis
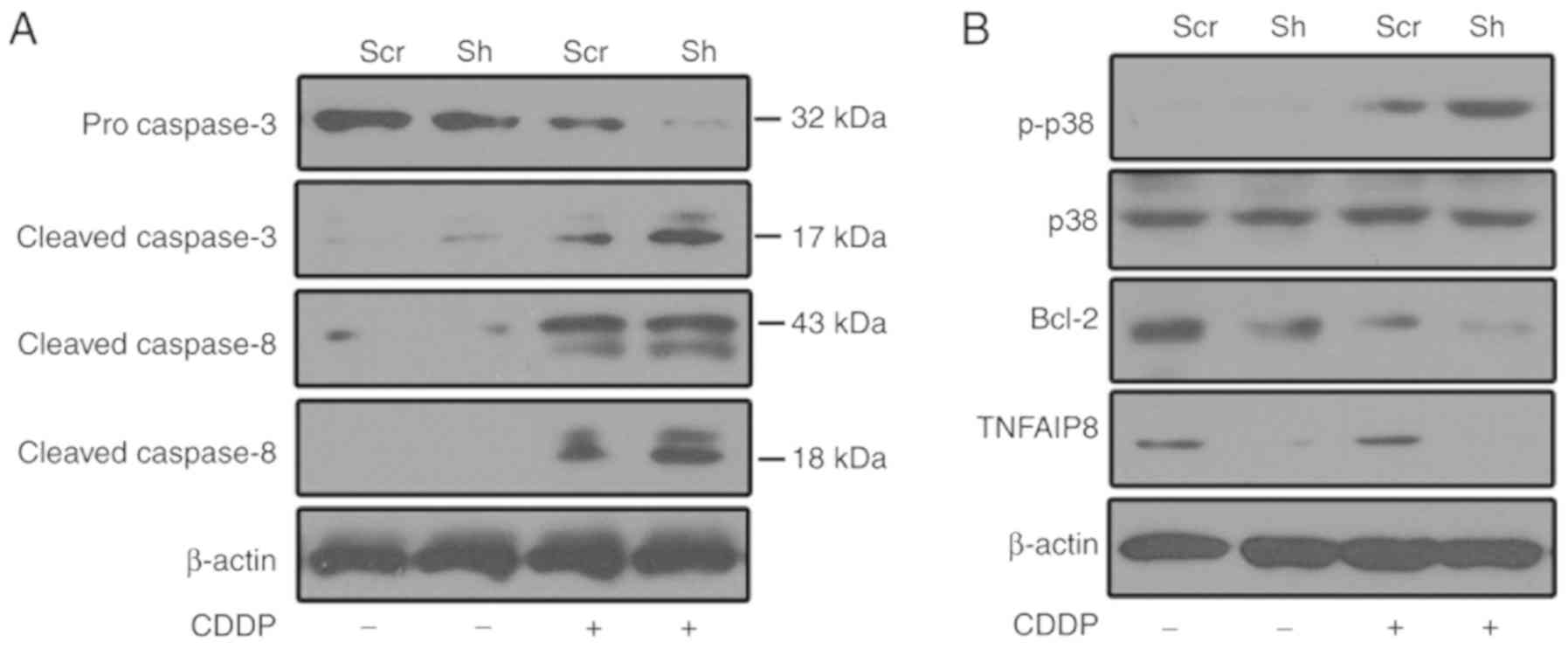
To investigate the underlying mechanisms of TNFAIP8
function in cisplatin resistance, caspase activation was evaluated
following treatment with cisplatin. As presented in Fig. 4A, caspase-3 and −8 were activated
subsequent to treatment with cisplatin, and TNFAIP8 silencing
enhanced caspase activation in HeLa cells following treatment with
cisplatin. Compared with the control group, Bcl-2 expression was
decreased in the TNFAIP8-silenced group following treatment with
cisplatin (Fig. 4B). To determine
whether p38 mitogen-activated protein kinase (MAPK) contributes to
cisplatin resistance in TNFAIP8-silenced HeLa cells, p38 MAPK
activation was assessed. p38 signaling was activated in
cisplatin-treated HeLa cells, and p38 phosphorylation was enhanced
in the TNFAIP8-knockdown group subsequent to treatment with
cisplatin (Fig. 4B). These data
suggested that TNFAIP8 serves as an anti-apoptotic protein against
cisplatin-induced cell death. This effect may aggravate
chemotherapeutic treatment failure in patients with cervical
cancer.
Discussion
To date, four members of the TNFAIP8 family of
proteins have been characterized in mammals; TNFAIP8 (or TIPE),
TNFAIP8-like 1 (TIPE1), TIPE2 and TIPE3 (22). These proteins possess different
biological functions, although they possess high sequence consensus
and share a death effecter domain (DED) (22). TNFAIP8 is the principal member of
this family that is induced by TNF-α in response to environmental
stress (11). It has previously been
identified that TNFAIP8 is overexpressed in a wide range of
different cancer types, and additional previous studies have
demonstrated that it serves as an oncoprotein, as it induces the
development and progression of tumors, and enhances cancer cell
survival, proliferation and metastasis (12,23,24).
However, little is known regarding the biological and pathological
functions of TNFAIP8 in cervical cancer. In the present study, a
significant increase in TNFAIP8 expression was observed in cervical
cancer tissues compared with the adjacent tissues. Furthermore,
when TNFAIP8 was silenced in HeLa cells, cell colony formation and
viability were impaired. This supported the hypothesis that TNFAIP8
serves as an oncogene, and promotes cervical cancer cell
proliferation and survival.
Cisplatin is a widely used chemotherapeutic drug in
the treatment of numerous types of solid tumors, including head and
neck, ovarian, testicular, small-cell lung and cervical cancer
(25). However, cisplatin is limited
as an effective treatment due to dose-associated toxicities and
adverse side-effects, whereas, the intrinsic or acquired resistance
of cells to cisplatin inevitably causes treatment failure (26). Therefore, the two significant
challenges associated with cisplatin therapy are identifying a safe
and tolerable dose, and addressing the issue of cisplatin
resistance. A previous study on the mechanisms of cisplatin
resistance have demonstrated that tumor cells develop resistance as
a natural self-defense mechanism to protect cells from the
cytotoxic effect of cisplatin and to support cell survival
(8). TNFAIP8 is a member of the
Fas-associated death domain-like interleukin-1β-converting enzyme
caspase-8-inhibitory protein family of anti-apoptotic proteins that
contain a DED and as a result, may cause the inhibition of
caspase-mediated cellular apoptosis (9,10).
Overexpression of TNFAIP8 in cancer cells is associated with
enhanced survival and with inhibition of the apoptotic enzymes
caspase-8 and caspase-3 (27). In
the present study, knockdown of TNFAIP8 expression in HeLa cells
inhibited caspase-8 and caspase-3 activation, upon treatment with
cisplatin, demonstrating that TNFAIP8 may be an anti-apoptotic
protein that serves an important role in cellular survival and
cisplatin resistance.
A previous study suggested that dysregulation of the
anti-apoptotic protein Bcl-2 constitutes a risk factor for
tumorigenesis (28). Additionally,
the increased expression of Bcl-2 in a variety of cancer types has
been associated with resistance to chemotherapeutic drugs,
including cisplatin (29). In the
present study, TNFAIP8 knockdown decreased Bcl-2 expression in
cisplatin-treated cervical cancer cells. Along with the observation
that depletion of TNFAIP8 improved cisplatin sensitivity and
promoted cisplatin-induced cellular apoptosis, the present data
suggested that TNFAIP8 is involved in cisplatin resistance by
regulating Bcl-2 expression and suppressing cellular apoptosis.
p38 MAPKs are serine/threonine protein kinases that
are involved in a variety of intracellular signaling events, and
modulate varied biological effects, including proliferation,
apoptosis, differentiation and senescence (30). In mammalian cells, four isoforms of
p38 MAPK have been identified; however, the physiological and
pathophysiological functions of the individual isoforms remain
largely unknown. Despite high amino acid sequence similarity, the
p38 MAPK isoforms present marked differences in tissue expression
and functions. p38α and p38β are ubiquitous, whereas, p38γ and p38δ
are tissue specific (31). The
different p38 isoforms have been demonstrated to exert opposing
effects. For example, p38α suppresses inflammation-associated
initiation of colon tumors; however, contributes to the
proliferation and survival of colon tumor cells (32). In vascular endothelial cells, p38α
mediates pro-apoptotic signaling from transforming growth
factor-β1-induced apoptosis, whereas, p38β relays survival signals
from pro-survival factors, including vascular endothelial growth
factor (33). Given the duality of
p38 MAPK functions in mammalian cells, the role of p38 MAPK was
investigated in TNFAIP8-silenced HeLa cells. In the present study,
it was demonstrated that treatment with cisplatin activated the p38
signaling pathway, and that TNFAIP8 silencing promoted p38
phosphorylation in cisplatin-treated HeLa cells. Additionally, the
present data demonstrated that when TNFAIP8 was silenced,
cisplatin-induced HeLa cell apoptosis and death were enhanced.
Therefore, it was suggested that p38 MAPK activation likely serves
a negative role in TNFAIP8-mediated cisplatin resistance, and that
regulating p38 MAPK activity may be used to reverse cisplatin
resistance in women with cervical cancer.
Numerous proteins, including cluster of
differentiation (CD)44, CD24, CD38, Fos-related antigen-1, heat
shock protein β-1 and transcription factor SOX-9, have been
implicated in cervical cancer, and dysregulation of these proteins
is closely associated with the occurrence and outcome of cervical
cancer (34–39). However, the etiology of cervical
carcinoma remains elusive. In the present study, it was
demonstrated that in cervical cancer cells, TNFAIP8 overexpression
is associated with resistance to cisplatin, whereas, TNFAIP8
silencing improved cisplatin sensitivity in HeLa cells. The present
data suggested that TNFAIP8 may be considered a promising
therapeutic target for the treatment of cervical cancer.
Acknowledgements
The authors would like to thank Professor Jiang Wu
and Mrs Xi Chen from the Pathology Department of Henan University
Huaihe Hospital for their technical assistance.
Funding
No funding was received.
Availability of data and materials
The datasets used and analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SW and LC conceived the study and wrote the article.
SW and WL designed and performed the experiments, and analyzed the
data. ZW, TC, PW, NL, XL and MC were involved in the design or
execution of a number of experiments. SZ and YL designed and
performed the human tumor studies. YM made suggestions for the
design of the study and contributed to revision of the manuscript
and approved the version to be submitted. All authors have read and
approved the final manuscript.
Ethics approval and consent to
participate
The present study complies with current ethical
considerations and the protocol was approved by the Committee for
Ethical Review at Henan University School of Medicine (Kaifeng,
China). Written informed consent was provided by all patients.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
TNFAIP8
|
tumor necrosis factor α-induced
protein 8
|
|
MAPK
|
mitogen-activated protein kinase
|
|
CDDP
|
Cis-diamminedichloroplatinum
|
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parish SL, Swaine JG, Son E and Luken K:
Determinants of cervical cancer screening among women with
intellectual disabilities: Evidence from medical records. Public
Health Rep. 128:519–526. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Diaz-Padilla I, Monk BJ, Mackay HJ and
Oaknin A: Treatment of metastatic cervical cancer: Future
directions involving targeted agents. Crit Rev Oncol Hematol.
85:303–314. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Takekuma M, Kuji S, Tanaka A, Takahashi N,
Abe M and Hirashima Y: Platinum sensitivity and
non-cross-resistance of cisplatin analogue with cisplatin in
recurrent cervical cancer. J Gynecol Oncol. 26:185–192. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zhu M, Zhou X, Du Y, Huang Z, Zhu J, Xu J,
Cheng G, Shu Y, Liu P, Zhu W, et al: miR-20a induces cisplatin
resistance of a human gastric cancer cell line via targeting CYLD.
Mol Med Rep. 14:1742–1750. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mehta FF, Baik S and Chung SH: Recurrence
of cervical cancer and its resistance to progestin therapy in a
mouse model. Oncotarget. 8:2372–2380. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bi L, Ma F, Tian R, Zhou Y, Lan W, Song Q
and Cheng X: AJUBA increases the cisplatin resistance through hippo
pathway in cervical cancer. Gene. 644:148–154. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Shen DW, Pouliot LM, Hall MD and Gottesman
MM: Cisplatin resistance: A cellular self-defense mechanism
resulting from multiple epigenetic and genetic changes. Pharmacol
Rev. 64:706–721. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kumar D, Whiteside TL and Kasid U:
Identification of a novel tumor necrosis factor-alpha-inducible
gene, SCC-S2, containing the consensus sequence of a death effector
domain of fas-associated death domain-like
interleukin-1beta-converting enzyme-inhibitory protein. J Biol
Chem. 275:2973–2978. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kumar D, Gokhale P, Broustas C,
Chakravarty D, Ahmad I and Kasid U: Expression of SCC-S2, an
antiapoptotic molecule, correlates with enhanced proliferation and
tumorigenicity of MDA-MB 435 cells. Oncogene. 23:612–616. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Patel S, Wang FH, Whiteside TL and Kasid
U: Identification of seven differentially displayed transcripts in
human primary and matched metastatic head and neck squamous cell
carcinoma cell lines: Implications in metastasis and/or radiation
response. Oral Oncol. 33:197–203. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Goldsmith JR and Chen YH: Regulation of
inflammation and tumorigenesis by the TIPE family of phospholipid
transfer proteins. Cell Mol Immunol. 14:10262017. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang LJ, Liu X, Gafken PR, Kioussi C and
Leid M: A chicken ovalbumin upstream promoter transcription factor
I (COUP-TFI) complex represses expression of the gene encoding
tumor necrosis factor alpha-induced protein 8 (TNFAIP8). J Biol
Chem. 284:6156–6168. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xiao M, Xu Q, Lou C, Qin Y, Ning X, Liu T,
Zhao X, Jia S and Huang Y: Overexpression of TNFAIP8 is associated
with tumor aggressiveness and poor prognosis in patients with
invasive ductal breast carcinoma. Hum Pathol. 62:40–49. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sun Z, Liu X, Song JH, Cheng Y, Liu Y, Jia
Y, Meltzer SJ and Wang Z: TNFAIP8 overexpression: A potential
predictor of lymphatic metastatic recurrence in pN0 esophageal
squamous cell carcinoma after Ivor Lewis esophagectomy. Tumour
Biol. 37:10923–10934. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li Y, Jing C, Chen Y, Wang J, Zhou M, Liu
X, Sun D, Mu L, Li L and Guo X: Expression of tumor necrosis factor
α-induced protein 8 is upregulated in human gastric cancer and
regulates cell proliferation, invasion and migration. Mol Med Rep.
12:2636–2642. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang M, Zhao Q, Wang X, Liu T, Yao G, Lou
C and Zhang Y: TNFAIP8 overexpression is associated with lymph node
metastasis and poor prognosis in intestinal-type gastric
adenocarcinoma. Histopathology. 65:517–526. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liu T, Gao H, Yang M, Zhao T, Liu Y and
Lou G: Correlation of TNFAIP8 overexpression with the
proliferation, metastasis, and disease-free survival in endometrial
cancer. Tumour Biol. 35:5805–5814. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang C, Chakravarty D, Sakabe I, Mewani
RR, Boudreau HE, Kumar D, Ahmad I and Kasid UN: Role of SCC-S2 in
experimental metastasis and modulation of VEGFR-2, MMP-1, and MMP-9
expression. Mol Ther. 13:947–955. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gus-Brautbar Y, Johnson D, Zhang L, Sun H,
Wang P, Zhang S, Zhang L and Chen YH: The anti-inflammatory TIPE2
is an inhibitor of the oncogenic Ras. Mol Cell. 45:610–618. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Sun H, Gong S, Carmody RJ, Hilliard A, Li
L, Sun J, Kong L, Xu L, Hilliard B, Hu S, et al: TIPE2, a negative
regulator of innate and adaptive immunity that maintains immune
homeostasis. Cell. 133:415–426. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lou Y and Liu S: The TIPE (TNFAIP8) family
in inflammation, immunity, and cancer. Mol Immunol. 49:4–7. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fayngerts SA, Wu J, Oxley CL, Liu X,
Vourekas A, Cathopoulis T, Wang Z, Cui J, Liu S, Sun H, et al:
TIPE3 is the transfer protein of lipid second messengers that
promote cancer. Cancer Cell. 26:465–478. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Galluzzi L, Senovilla L, Vitale I, Michels
J, Martins I, Kepp O, Castedo M and Kroemer G: Molecular mechanisms
of cisplatin resistance. Oncogene. 31:1869–1883. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Siddik ZH: Cisplatin: Mode of cytotoxic
action and molecular basis of resistance. Oncogene. 22:7265–7279.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
You Z, Ouyang H, Lopatin D, Polver PJ and
Wang CY: Nuclear factor-kappa B-inducible death effector
domain-containing protein suppresses tumor necrosis factor-mediated
apoptosis by inhibiting caspase-8 activity. J Biol Chem.
276:26398–26404. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang TM, Barbone D, Fennell DA and
Broaddus VC: Bcl-2 family proteins contribute to apoptotic
resistance in lung cancer multicellular spheroids. Am J Respir Cell
Mol Biol. 41:14–23. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Low SY, Tan BS, Choo HL, Tiong KH, Khoo AS
and Leong CO: Suppression of BCL-2 synergizes cisplatin sensitivity
in nasopharyngeal carcinoma cells. Cancer Lett. 314:166–175. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chang L and Karin M: Mammalian MAP kinase
signalling cascades. Nature. 410:37–40. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cuadrado A and Nebreda AR: Mechanisms and
functions of p38 MAPK signalling. Biochem J. 429:403–417. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gupta J, del Barco Barrantes I, Igea A,
Sakellariou S, Pateras IS, Gorgoulis VG and Nebreda AR: Dual
function of p38α MAPK in colon cancer: Suppression of
colitis-associated tumor initiation but requirement for cancer cell
survival. Cancer Cell. 25:484–500. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ferrari G, Terushkin V, Wolff MJ, Zhang X,
Valacca C, Poggio P, Pintucci G and Mignatti P: TGF-β1 induces
endothelial cell apoptosis by shifting VEGF activation of
p38MAPK from the prosurvival p38β to proapoptotic p38α.
Mol Cancer Res. 10:605–614. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xiao S, Zhou Y, Yi W, Luo G, Jiang B, Tian
Q, Li Y and Xue M: Fra-1 is downregulated in cervical cancer
tissues and promotes cervical cancer cell apoptosis by p53
signaling pathway in vitro. Int J Oncol. 46:1677–1684. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wobus M, Kuns R, Wolf C, Horn LC, Köhler
U, Sheyn I, Werness BA and Sherman LS: CD44 mediates constitutive
type I receptor signaling in cervical carcinoma cells. Gynecol
Oncol. 83:227–234. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Liao S, Xiao S, Zhu G, Zheng D, He J, Pei
Z, Li G and Zhou Y: CD38 is highly expressed and affects the
PI3K/Akt signaling pathway in cervical cancer. Oncol Rep.
32:2703–2709. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wu JH, Liang XA, Wu YM, Li FS and Dai YM:
Identification of DNA methylation of SOX9 in cervical cancer using
methylated-CpG island recovery assay. Oncol Rep. 29:125–132. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Dobo C, Stavale JN, Lima Fde O, Ribeiro
DA, Arias V, Gomes TS and Oshima CT: HSP27 is commonly expressed in
cervical intraepithelial lesions of Brazilian women. Asian Pac J
Cancer Prev. 14:5007–5010. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pei Z, Zhu G, Huo X, Gao L, Liao S, He J,
Long Y, Yi H, Xiao S, Yi W, et al: CD24 promotes the proliferation
and inhibits the apoptosis of cervical cancer cells in
vitro. Oncol Rep. 35:1593–1601. 2016. View Article : Google Scholar : PubMed/NCBI
|