Introduction
Lung cancer remains the leading cause of
cancer-associated mortality, annually resulting in >1,000,000
mortalities globally in 2012 (1).
Additionally, ~1,200,000 new cases are diagnosed every year and
prognoses of lung cancer are poor, as estimated worldwide in 2012
(2). Lung adenocarcinoma is the
predominant histological subtype of non-small lung cancer (3). The mean 5-year survival rate of lung
adenocarcinoma is <15% in the urban areas of China as estimated
in 2015, primarily due to the late-stage detection and a paucity of
late-stage treatments (4). The
availability of effective anticancer drugs is considered to be the
key for improvement in the treatment of lung adenocarcinoma.
7, 8-Diacetoxy-4-methylcoumarin (DAMTC) is a
thioderivative of 4-methyl coumarin (5). Coumarins belong to the flavonoid class
of plant secondary metabolite, which have been demonstrated to
exhibit diverse and beneficial biological activities, including
antitumoral, anticoagulant and anti-inflammatory properties
(5). Natural coumarins or synthetic
analogs have attracted intense interest for their applicability as
drugs (6). They have been determined
to have a variety of therapeutic applications, including antitumor
and anti-human immunodeficiency virus therapy (7), and antioxidant (8) and antibacterial (9) applications. Lacy and O'Kennedy
(10) demonstrated that genistein
and esculetin could exert the most potent inhibitory effect on cell
growth of two cell lines, A549, a lung carcinoma cell line, and
MCF-7, a breast carcinoma cell line. Previously, the coumarin
derivative DAMTC was indicated to inhibit cellular proliferation
and induce apoptosis on human non-small cell lung cancer A549 cells
(11). Furthermore, the observations
of Goel et al (12) reported
that upregulation of the nuclear factor-κB, p53 and Akt pathways,
and downregulation of the mitogen activated protein kinase (MAPK)
and Cox-2 pathways were involved in the molecular mechanism of
apoptosis induction by DAMTC in A549 cells. However, the mechanisms
of the anti-proliferative effects of DAMTC in lung adenocarcinoma
are incompletely defined, and further insights into the mechanisms
are required.
Previously, Goel et al (1) used the integrated proteomics and
transcriptomics approach, and identified that DAMTC could regulate
cell motility and cytoskeletal reorganization in lung
adenocarcinoma. In the present study, differentially-expressed
genes (DEGs) were identified in DAMTC-treated lung adenocarcinoma,
compared with DAMTC-untreated controls, using the same gene
expression profile. Comprehensive bioinformatics were used to
analyze the significant pathways and functions, and to construct
the protein-protein interaction (PPI) network, to determine the
critical DEGs. Furthermore, the putative interactions between
signaling pathways were analyzed. The present study aimed to
investigate the potential molecular mechanism underlying
DAMTC-induced apoptosis and inhibition of cell motility in lung
adenocarcinoma.
Materials and methods
Microarray data and data
preprocessing
The gene expression profile of GSE29698, deposited
by Goel et al (1), was
downloaded from the Gene Expression Omnibus database in National
Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov/geo/) based on the
platform of GPL6884 Illumina HumanWG-6 v3.0 expression beadchip. A
total of 6 specimens were applied, including 3 specimens of
DAMTC-treated lung adenocarcinoma cells (A549) and another 3
specimens of DAMTC-untreated A549 cell lines as controls.
The gene expression profile data were preprocessed
using the limma (13) package in
Bioconductor. Following background correction, quantile
normalization and probe summarization, the gene expression matrix
of specimens was received.
DEGs screening
Unpaired Student's t-test (13) in limma package was used to identify
the DEGs in the DAMTC-treated A549 cell group, compared with the
control group. False discovery rate (FDR) (14) was performed for multiple testing
correction using the Benjamini and Hochberg method (15). The threshold for the DEGs was set as
FDR <0.01 and |log2 FC (fold change) |≥2.
Functional and pathway enrichment
analysis of DEGs
Gene Ontology (GO) (16) categories, including biological
process (BP), molecular function (MF) and cellular component (CC),
of the selected DEGs were enriched from GO databases using Database
for Annotation Visualization and Integrated Discovery (DAVID)
(17). Additionally, the pathways of
selected DEGs were enriched using DAVID from Kyoto Encyclopedia of
Genes and Genomes (KEGG) (18)
analysis. P<0.05, as determined by the hypergeometric test
(19), was selected as the
threshold.
Functional annotation of DEGs
Identification of tumor-associated genes (TAGs) and
understanding their functions can be critical for investigating the
roles of genes involved in tumorigenesis. The tumor suppressor gene
(TSGene) database (http://bioinfo.mc.vanderbilt.edu/TSGene/) is a
comprehensive literature-based database that provides detailed
annotations for each TSG. The TAG database (http://www.binfo.ncku.edu.tw/TAG/) is designed to
utilize information from well-characterized oncogenes and tumor
suppressor genes to accelerate cancer research. According to the
data information regarding transcription factors (TFs) from the
TRANSFAC database (20), functional
enrichment of the DEGs for transcription regulation was assessed.
Additionally, the selected DEGs were mapped into the TSGene and TAG
database to extract the genes that had transcriptional functions or
functioned as TAGs.
PPI network construction
The PPI network is represented by an undirected
graph with nodes indicating the genes and edges indicating the
mapped interactions of the proteins encoded by the genes (21). The PPI network of the selected genes
was constructed by using data from the Retrieval of Interacting
Genes (STRING) database, which is a comprehensive database
containing functional associations between proteins that are
experimentally derived, as well as associations predicted by
comparative genomics and text mining (22). The interaction pairs with the PPI
combined score >0.4 were selected in this network, which
corresponded to a medium-confidence network (23).
Pathway crosstalk analysis
If more significant protein interactions are
detected between two pathways, these two pathways are probable to
influence or interact with each other (cross-talk). Understanding
the crosstalk between pathways is momentous for understanding the
function of cells and more complex systems (24). In the present study, the KEGG pathway
data and protein interaction data based on the PPI network were
combined to investigate pathway crosstalk in the DAMTC-treated A549
cells. The detail is included in the studies by Li et al
(24) and Liu et al (25).
Results
Identification of DEGs
For the dataset GSE29698, a total of 889 DEGs were
identified in DAMTC-treated A549 cell groups, compared with
DAMTC-untreated controls, including 500 upregulated genes and 389
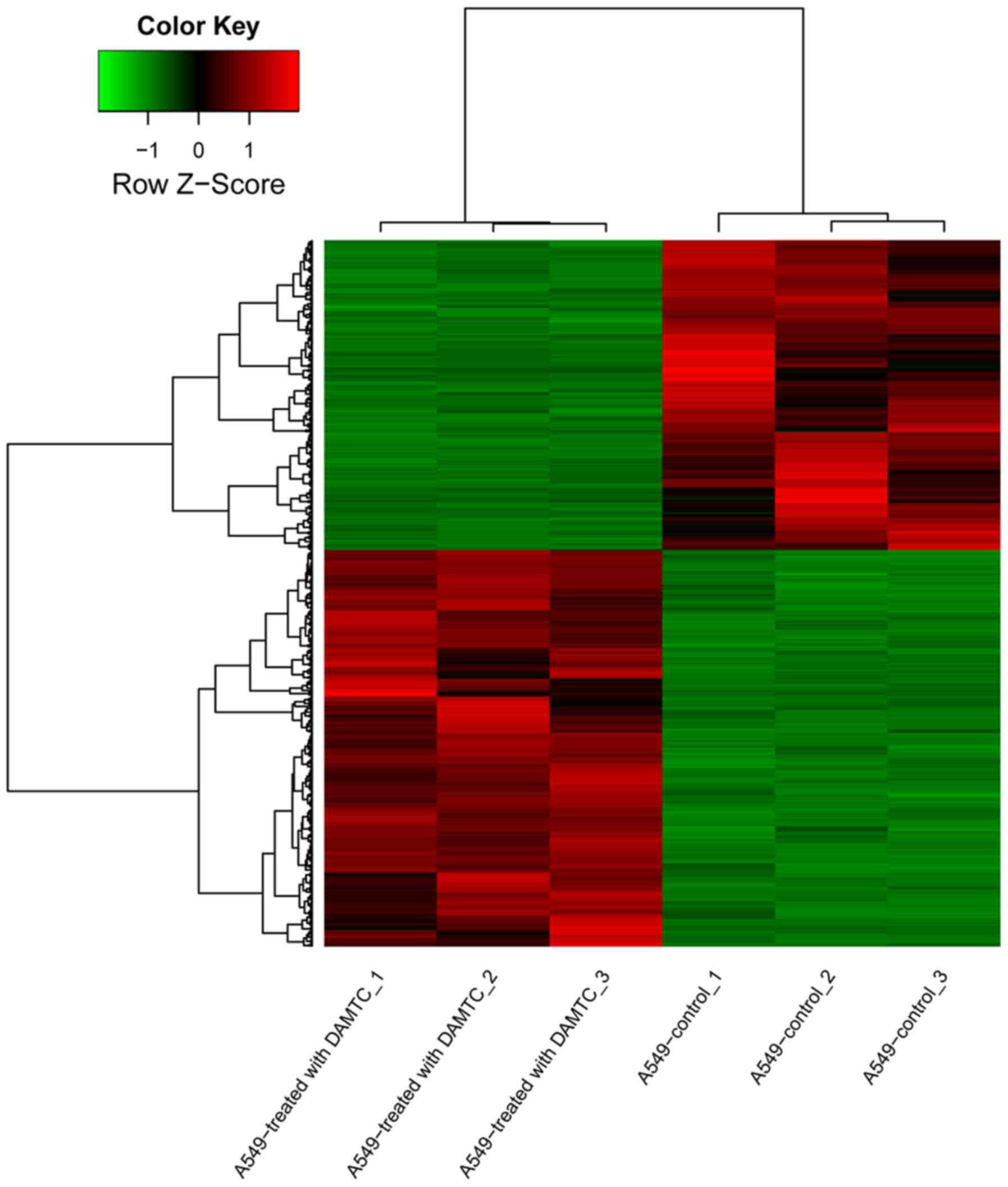
downregulated genes. The heat-map of the DEGs is depicted in
Fig. 1. The results demonstrated
that the DEGs expression pattern could significantly distinguish
the DAMTC-treated A549 cell samples from controls.
GO and pathway enrichment analysis of
the DEGs
Functional and pathway enrichment analysis indicated
that upregulated and downregulated DEGs in DAMTC-treated A549 cell
groups were significantly enriched in different GO terms and KEGG
pathways (Tables I and II). GO analysis demonstrated enrichment in
upregulated DEGs involved in BP, including transcription,
DNA-dependent and regulation of cellular metabolic process.
Significant CC ontology terms of upregulated DEGs were associated
with intracellular part. The majority of significant MF ontology
terms of upregulated DEGs were associated with binding function and
enzyme activity, including DNA binding and phosphatase activity.
Furthermore, significant BP ontology terms of downregulated DEGs
were associated with tissue morphogenesis, and morphogenesis of an
epithelium. CC ontology terms of downregulated DEGs were also
associated with intracellular part. Significant MF ontology terms
of downregulated DEGs were associated with organic cyclic compound
binding, and RNA polymerase II transcription corepressor activity
(Table I).
 | Table I.The enriched GO terms of
differentially-expressed genes in 7,
8-diacetoxy-4-methylcoumarin-treated lung cancer groups. |
Table I.
The enriched GO terms of
differentially-expressed genes in 7,
8-diacetoxy-4-methylcoumarin-treated lung cancer groups.
| Expression
changes | Category | GO-ID | Name | Count | P-value |
|---|
| Upregulated | BP | GO:0006351 | Transcription,
DNA-dependent | 153 |
3.30×10−11 |
|
|
| GO:0031323 | Regulation of
cellular metabolic process | 194 |
7.80×10−11 |
|
|
| GO:0060255 | Regulation of
macromolecule metabolic process | 187 |
8.24×10−11 |
|
|
| GO:0051252 | Regulation of RNA
metabolic process | 143 |
1.56×10−10 |
|
|
| GO:0019222 | Regulation of
metabolic process | 207 |
2.08×10−10 |
|
| CC | GO:0005622 | Intracellular | 387 |
2.45×10−10 |
|
|
| GO:0044424 | Intracellular
part | 382 |
1.08×10−9 |
|
|
| GO:0005634 | Nucleus | 219 |
2.68×10−9 |
|
|
| GO:0043227 | Membrane-bounded
organelle | 320 |
5.19×10−9 |
|
|
| GO:0043231 | Intracellular
membrane-bounded organelle | 319 |
5.71×10−9 |
|
| MF | GO:0003677 | DNA binding | 98 |
3.25×10−5 |
|
|
| GO:0016791 | Phosphatase
activity | 18 |
4.00×10−4 |
|
|
| GO:0000988 | Protein binding
transcription factor activity | 28 |
8.00×10−4 |
|
|
| GO:0005515 | Protein
binding | 244 |
11.00×10−3 |
|
|
| GO:0005488 | Binding | 362 |
13.00×10−3 |
| Downregulated | BP | GO:0048729 | Tissue
morphogenesis | 28 |
2.61×10−6 |
|
|
| GO:0002009 | Morphogenesis of an
epithelium | 24 |
3.72×10−6 |
|
|
| GO:0044237 | Cellular metabolic
process | 221 |
7.50×10−5 |
|
|
| GO:0044238 | Primary metabolic
process | 224 |
8.50×10−5 |
|
|
| GO:0048736 | Appendage
development | 12 |
8.77×10−5 |
|
| CC | GO:0044424 | Intracellular
part | 291 |
3.52×10−8 |
|
|
| GO:0043231 | Intracellular
membrane-bounded organelle | 243 |
1.83×10−7 |
|
|
| GO:0005622 | Intracellular | 291 |
2.11×10−7 |
|
|
| GO:0043227 | Membrane-bounded
organelle | 243 |
2.69×10−7 |
|
|
| GO:0043229 | Intracellular
organelle | 257 |
2.01×10−6 |
|
| MF | GO:0097159 | Organic cyclic
compound binding | 154 |
2.31×10−6 |
|
|
| GO:1901363 | Heterocyclic
compound binding | 152 |
2.77×10−6 |
|
|
| GO:0001191 | RNA polymerase II
transcription factor binding transcription factor activity involved
in negative regulation of transcription | 6 |
2.35×10−5 |
|
|
| GO:0001106 | RNA polymerase II
transcription corepressor activity | 5 |
3.99×10−5 |
|
|
| GO:0003714 | Transcription
corepressor activity | 14 |
8.22×10−5 |
| Table II.The enriched pathways of
differentially-expressed genes in 7,
8-diacetoxy-4-methylcoumarin-treated lung cancer groups. |
Table II.
The enriched pathways of
differentially-expressed genes in 7,
8-diacetoxy-4-methylcoumarin-treated lung cancer groups.
| Expression
changes | KEGG-ID | Name | n | P-value | Genes |
|---|
| Upregulated | 04115 | p53 signaling
pathway | 9 | 0.0001 | APAF1, CCNG2, CYCS,
GADD45A, MDM2, PPM1D, RPRM, SESN1 and SESN3 |
|
| 00051 | Fructose and
mannose metabolism | 5 | 0.0032 | ALDOC, C12orf5,
HK2, MTMR1 and PFKFB4 |
|
| 05219 | Bladder cancer | 5 | 0.0062 | FGFR3, MAP2K1,
MDM2, PGF and RASSF1 |
|
| 04722 | Neurotrophin
signaling pathway | 9 | 0.0095 | CDC42, GAB1,
MAP2K1, MAP3K3, MAPK7, NFKBIE, NTF3, PDK1 and ZNF274 |
|
| 05200 | Pathways in
cancer | 17 | 0.0103 | BMP2, CCNA1, CDC42,
CYCS, EGLN1, FGF18, FGFR3, FZD1, FZD4, FZD7, MAP2K1, MAX, MDM2,
PGF, PIAS3, RASSF1 and RASSF5 |
|
| 04130 | SNARE interactions
in vesicular transport | 4 | 0.0180 | BNIP1, STX11, STX7
and VAMP1 |
|
| 04144 | Endocytosis | 11 | 0.0265 | CDC42, CXCR4, DNM2,
FGFR3, GIT2, HLA-E, MDM2, RAB11FIP1, SH3GL2, SH3GL3 and VPS37B |
|
| 05211 | Renal cell
carcinoma | 5 | 0.0472 | CDC42, EGLN1, GAB1,
MAP2K1 and PGF |
| Downregulated | 05217 | Basal cell
carcinoma | 6 | 0.0008 | BMP4, FZD2, GLI2,
STK36, TCF7L2 and WNT7B |
|
| 00130 | Ubiquinone and
other terpenoid-quinone biosynthesis | 2 | 0.0079 | COQ5 and COQ7 |
|
| 01100 | Metabolic
pathways | 33 | 0.0130 | ACSS1, AK2,
ATP6V1E2, B3GALT6, BAAT, BCAT1, COQ5, COQ7, CYP2R1, DPAGT1, GGPS1,
GPT2, HKDC1, HLCS, OXSM, PAFAH2, PCK2, PFAS, PIGM, PIGV, PIGW,
PIK3C2B, PLD1, PMM2, POLG2, POLR1B, POLR3H, PSPH, SEPHS2, SLC33A1,
ST3GAL5, ST6GAL1 and SYNJ2 |
|
| 00563 |
Glycosylphosphatidyli-nositol-anchor
biosynthesis | 3 | 0.0132 | PIGM, PIGV and
PIGW |
|
| 04120 | Ubiquitin mediated
proteolysis | 7 | 0.0185 | ANAPC13, BIRC6,
FBXO4, ITCH, KLHL9, UBE2J1 and UBE2Q1 |
|
| 04340 | Hedgehog signaling
pathway | 4 | 0.0255 | BMP4, GLI2, STK36
and WNT7B |
|
| 05200 | Pathways in
cancer | 12 | 0.0300 | BMP4, DAPK1, FZD2,
GLI2, IL6, PLD1, RARB, STAT1, STK36, TCF7L2, TRAF5 and WNT7B |
Additionally, upregulated DEGs were enriched in
different pathways, including the p53 signaling pathway, and
fructose and mannose metabolism pathway. Downregulated DEGs were
also identified to be involved in different pathways, including
ubiquitin mediated proteolysis, and metabolic pathways (Table II).
Function annotation of DEGs
The expression change of TFs, TSGs and other TAGs in
DAMTC-treated A549 cell groups were observed (Table III). The upregulated functional
genes included 34 TFs and 26 TSGs. Subsequently, MAX was
identified to be a TF, while tumor protein p53 inducible nuclear
protein 1 and bone morphogenetic protein 2 were identified to be
TSGs. Additionally, histone deacetylase 3 (HDAC3) was
identified as both a TF and a TSG. Additionally, the downregulated
functional genes included 27 TFs and 18 TSGs. Among them, sex
determining region Y-box 4 and signal transducer and activator of
transcription 1 (STAT1) were identified to be TFs.
| Table III.Functional annotation of
differentially-expressed genes in 7,
8-diacetoxy-4-methylcoumarin-treated lung cancer groups. |
Table III.
Functional annotation of
differentially-expressed genes in 7,
8-diacetoxy-4-methylcoumarin-treated lung cancer groups.
| Expression
changes | TF | TSG | Other TAG |
|---|
| Upregulated | ATF2, BCL6, CDK7,
ERCC6, EZH2, FOXD1, GATA2, GATA4, GMEB1, GTF2B, HAND1, HDAC3, HEY2,
HOXD10, HOXD11, ING1, IRF7, IRF8, IRX5, ISL1, MAFB, MAFF, MAX,
MEF2A, NFIL3, PAX9, PLAGL1, RFX2, RORA, SOX8, TMF1, VDR, ZNF10 and
ZNF133 | APAF1, ARID3B,
BCL10, BIK, BMP2, BRMS1, CAMTA1, CDKN1C, CDKN2D, DDX3X, EGLN1,
FBXW7, HDAC3, ING1, IRF8, LATS2, NDRG1, PHF6, PLAGL1, PLEKHO1,
PPP1R3C, RASSF1, RASSF5, SLC9A3R1, TMEFF2 and TP53INP1 | CHRM3, FZD7,
LGALS8, MAFB, MAP1A, MAX, PIAS3 and RGS2 |
| Downregulated | AKNA, CEBPG, FOXL2,
FOXQ1, GATA3, GLI2, GTF2E1, HOXB5, HOXB8, HOXC8, MEIS1, MEIS2, MSC,
NFIA, NFIB,NPAS2, NR2E3, NR2F2, NR5A2, PBX1, PRDM1, SMAD6, SOX4,
STAT1, TBX18, TCF7L2 and XBP1 | ABLIM3, ARID1A,
ASXL1, CDH4, COL4A3, DAB2, DAPK1,DFNA5, DKK1, KCNRG, KRIT1, MN1,
MTUS1, NRCAM, PLK2, PRDM1, RARB and TCF7L2 | ATM, PMS1, RHOB,
SSPN and TFAP2A |
PPI network and the pathway crosstalk
analysis
Based on the STRING database, the PPI network was
constructed (Fig. 2). The results
demonstrated that a total of 9 genes had a node degree >16,
including interleukin 6 (IL6; degree, 37), MDM2 oncogene, E3
ubiquitin protein ligase (MDM2; degree, 27), STAT1
(degree, 23), ataxia telangiectasia mutated (degree, 20),
HDAC3 (degree, 19), cell division cycle 42 (CDC42;
degree, 18), cytochrome c, somatic (degree, 17), bone morphogenetic
protein 4 (degree, 17) and MAX (degree, 17).
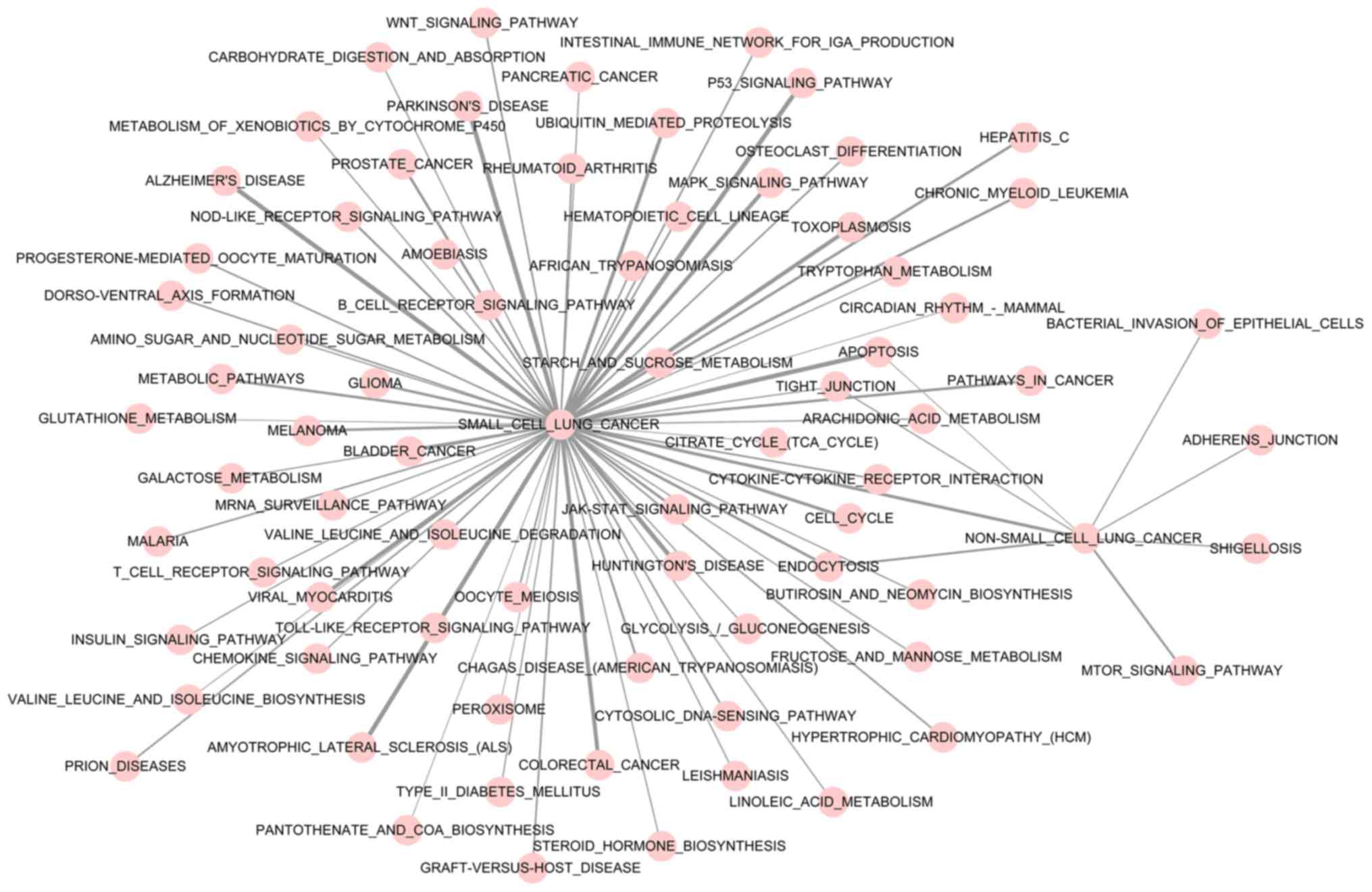
The result of pathway crosstalk analysis is depicted
in Fig. 3. The majority of the
significant pathways were determined to have crosstalk with small
cell lung cancer and non-small cell lung cancer. The apoptosis,
tight junction and endocytosis pathways were determined to be
cross-talking with small cell lung cancer and non-small cell lung
cancer. Additionally, the bacterial invasion of epithelial cells,
adherens junction, shigellosis and mechanistic target of rapamycin
kinase signaling pathways had cross-talk with non-small cell lung
cancer. In addition, small cell lung cancer and non-small cell lung
cancer were also determined to have cross-talk.
Discussion
The worldwide incidence rate of lung cancer is high
(26). It is important to develop
novel, more effective treatment modalities for dealing with this
disease. DAMTC is a potential inducer of apoptosis and an inhibitor
of cell growth (1). Thus,
identifying the molecular mechanism underlying the effects of DAMTC
is imperative. In the present study, microarray analysis
demonstrated that 500 upregulated DEGs and 389 downregulated DEGs
were identified in DAMTC-treated A549 cells, compared with
DAMTC-untreated controls. Among the DEGs, IL6, which was
downregulated, was a hub protein with the highest degree in the PPI
network. The upregulated DEGs MDM2, CDC42 and MAX,
which were also hub genes, had different functions and were
involved in different pathways, including the p53 signaling and
endocytosis pathways. Additionally, the apoptosis, tight junction,
and endocytosis pathways were determined to cross-talk with small
cell lung cancer and non-small cell lung cancer.
IL6 encodes a multifunctional cytokine, which
can function in inflammation and the maturation of B cells
(27). Hodge et al (28) demonstrated that the release of
various survival factors, including IL6, serve to block apoptosis
in cancer cells during the inflammatory process, keeping them alive
in toxic environments. Additionally, studies have demonstrated that
drugs designed to restore apoptosis exhibit potential to
effectively treat numerous cancer types, including inhibitors of
apoptosis, protein inhibitors and MDM2 antagonists (29,30).
Furthermore, in keeping with the previous study, our study
identified that IL6 was downregulated in DAMTC-treated A549 groups
and was involved in the pathways in cancer (Table I). Thus, it was indicated that DAMTC
may induce cancer cell apoptosis through targeting IL6 in
lung adenocarcinoma.
Additionally, the other upregulated DEG MAX
was enriched in the pathways in cancer and identified to be a TAG
in the present study. MAX is a member of the basic helix-loop-helix
leucine zipper family of TFs and could interact with other family
members, including v-myc avian myelocytomatosis viral oncogene
homolog (MYC), MAX interactor 1, dimerization protein and mothers
against decapentaplegic (MAD), to form homodimers or heterodimers
(31). The study by Grandori et
al (32) demonstrated that the
MYC/MAX/MAD network was comprised of a group of TFs controlling
cell cycle differentiation, progression and death. Additionally,
Zhu et al (33) reported that
levels of MAD1 rapidly decrease upon mitogen stimulation, and this
degradation could be regulated by the MAPK pathway. Furthermore,
Goel et al (34) reported
that DAMTC could induce apoptosis through the mitochondrial pathway
by modulating the MAPK pathway. Thus, it was indicated that DAMTC
may induce cell apoptosis by upregulating the expression of
MAX and modulating the MAPK pathway in lung
adenocarcinoma.
Furthermore, pathway crosstalk analysis in the
present study demonstrated that apoptosis, tight junction and
endocytosis had cross-talks with non-small and small lung cancer.
Additionally, the present study also revealed that MDM2 and
CDC42, which were another two hub genes in the PPI network,
were identified to be involved in different pathways, including the
p53 signaling and endocytosis pathways. MDM2 encodes a
nuclear-localized E3 ubiquitin ligase that mediates ubiquitination
of p53 (35). Oren (36) demonstrated that proteins encoded by
one or more p53 target genes could serve an essential role in
causing p53-mediated apoptosis. This was consistent with the
observations by Goel et al (12), demonstrating that DAMTC could induce
apoptosis by modulating the p53 pathway in A549 cells. For the
other DEG, CDC42 is a small GTPase of the Rho-subfamily, which can
regulate signaling pathways that control numerous cellular
functions, including cell endocytosis, migration and cell cycle
progression (37). Georgiou et
al (38) reported that
CDC42 could act together with par-6 family cell polarity
regulator and protein kinase C in the regulation of Arp2/3-mediated
endocytosis to control local adherens junction stability,
modulating actin filament dynamics (38). Additionally, Mosesson et al
(39) revealed that aberrant
endocytosis of transmembrane proteins could contribute to malignant
transformation. Furthermore, Goel et al (34) demonstrated that DAMTC could inhibit
cell motility in A549 cells. Collectively, it was confirmed that
DAMTC could induce cancer cell apoptosis by modulating the p53
pathway. In addition, it was indicated that DAMTC may inhibit cell
motility through targeting CDC42 and affecting the
endocytosis pathway in lung adenocarcinoma.
In conclusion, the present study determined a
network of genes, including IL6, MDM2, CDC42 and MAX,
targeted by DAMTC that participated in different pathways were
involved in the mechanism of DAMTC-treated lung adenocarcinoma.
DAMTC may induce cell apoptosis by targeting IL6 and
MAX, which was involved in the MAPK pathway in lung
adenocarcinoma. Additionally, DAMTC may inhibit cell motility
through targeting CDC42 and affecting the endocytosis
pathway in lung adenocarcinoma. These activities may contribute to
the anti-carcinogenic action of DAMTC. Due to the relatively small
number of samples in the present study, further studies using a
larger sample size to validate and determine the role of the
potential molecular mechanism identified are required. Ongoing
studies may emphasize the potential of DAMTC as an anticancer
therapeutic.
Acknowledgements
Not applicable.
Funding
Not applicable.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available on reasonable request from the corresponding
author.
Authors' contributions
BW conceived and designed the study. YC and YK
acquired the data. XL and HF analyzed and interpreted the data. SZ
and YK performed statistical analysis. BW drafted the manuscript.
YC and TZ revised the manuscript for important intellectual
content. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Goel A, Chhabra R, Ahmad S, Prasad A,
Parmar V, Ghosh B and Saini N: DAMTC regulates cytoskeletal
reorganization and cell motility in human lung adenocarcinoma cell
line: An integrated proteomics and transcriptomics approach. Cell
Death Dis. 3:e4022012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel R, DeSantis C, Virgo K, Stein K,
Mariotto A, Smith T, Cooper D, Gansler T, Lerro C, Fedewa S, et al:
Cancer treatment and survivorship statistics, 2012. CA Cancer J
Clin. 62:220–241. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kaisermann MC, Trajman A and Madi K:
Evolving features of lung adenocarcinoma in Rio de Janeiro, Brazil.
Oncol Rep. 8:189–192. 2001.PubMed/NCBI
|
|
4
|
Imielinski M, Berger AH, Hammerman PS,
Hernandez B, Pugh TJ, Hodis E, Cho J, Suh J, Capelletti M,
Sivachenko A, et al: Mapping the hallmarks of lung adenocarcinoma
with massively parallel sequencing. Cell. 150:1107–1120. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Borges F, Roleira F, Milhazes N, Santana L
and Uriarte E: Simple coumarins and analogues in medicinal
chemistry: Occurrence, synthesis and biological activity. Curr Med
Chem. 12:887–916. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Musa MA, Cooperwood JS and Khan MO: A
review of coumarin derivatives in pharmacotherapy of breast cancer.
Curr Med Chem. 15:2664–2679. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kostova I, Raleva S, Genova P and Argirova
R: Structure-activity relationships of synthetic coumarins as HIV-1
inhibitors. Bioinorg Chem Appl. 682742006.PubMed/NCBI
|
|
8
|
Kontogiorgis CA and Hadjipavlou-Litina DJ:
Synthesis and biological evaluation of novel coumarin derivatives
with a 7-azomethine linkage. Bioorg Med Chem Lett. 14:611–614.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Appendino G, Mercalli E, Fuzzati N,
Arnoldi L, Stavri M, Gibbons S, Ballero M and Maxia A:
Antimycobacterial coumarins from the sardinian giant fennel
(Ferula communis). J Nat Prod. 67:2108–2110. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lacy A and O'Kennedy R: Studies on
coumarins and coumarin-related compounds to determine their
therapeutic role in the treatment of cancer. Curr Pharm Des.
10:3797–3811. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Goel A, Prasad A, Parmar V, Ghosh B and
Saini N: Antiproliferative effects of 7, 8 diacetoxy 4 methyl
coumarin and 7, 8 diacetoxy 4 methyl thiocoumarin in human lung
adenocarcinoma cell line. Clin Cancer Res. 13:C38. 2007.
|
|
12
|
Goel A, Prasad AK, Parmar VS, Ghosh B and
Saini N: Apoptogenic effect of 7, 8-diacetoxy-4-methylcoumarin and
7, 8-diacetoxy-4-methylthiocoumarin in human lung adenocarcinoma
cell line: Role of NF-kappaB, Akt, ROS and MAP kinase pathway. Chem
Biol Interact. 179:363–374. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Smyth GK: Limma: Linear models for
microarray data. In: Bioinformatics and Computational Biology
Solutions Using R and Bioconductor. Gentleman R, Carey VJ, Huber W,
Irizarry RA and Dudoit S: Springer; New York, NY: pp. 397–420.
2005
|
|
14
|
Reiner-Benaim A: FDR Control by the BH
Procedure for two-sided correlated tests with implications to gene
expression data analysis. Biom J. 49:107–126. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Benjamini Y and Hochberg Y: Controlling
the false discovery rate: A practical and powerful approach to
multiple testing. J R Stat Soc Series B (Methodological).
57:289–300. 1995. View Article : Google Scholar
|
|
16
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: Gene ontology: Tool for the unification of biology. The Gene
Ontology Consortium. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kanehisa M and Goto S: KEGG: Kyoto
encyclopedia of genes and genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Du P, Feng G, Flatow J, Song J, Holko M,
Kibbe WA and Lin SM: From disease ontology to disease-ontology
lite: Statistical methods to adapt a general-purpose ontology for
the test of gene-ontology associations. Bioinformatics. 25:i63–i68.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Matys V, Fricke E, Geffers R, Gössling E,
Haubrock M, Hehl R, Hornischer K, Karas D, Kel AE, Kel-Margoulis
OV, et al: TRANSFAC: Transcriptional regulation, from patterns to
profiles. Nucleic Acids Res. 31:374–378. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Köhler S, Bauer S, Horn D and Robinson PN:
Walking the interactome for prioritization of candidate disease
genes. Am J Hum Genet. 82:949–958. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Franceschini A, Szklarczyk D, Frankild S,
Kuhn M, Simonovic M, Roth A, Lin J, Minguez P, Bork P, von Mering C
and Jensen LJ: STRING v9. 1: Protein-protein interaction networks,
with increased coverage and integration. Nucleic Acids Res.
41:(Database Issue). D808–D815. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Von Mering C, Huynen M, Jaeggi D, Schmidt
S, Bork P and Snel B: STRING: A database of predicted functional
associations between proteins. Nucleic Acids Res. 31:258–261. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li Y, Agarwal P and Rajagopalan D: A
global pathway crosstalk network. Bioinformatics. 24:1442–1447.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu ZP, Wang Y, Zhang XS and Chen L:
Identifying dysfunctional crosstalk of pathways in various regions
of Alzheimer's disease brains. BMC Syst Biol. 4 Suppl 2:S112010.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Cooper S and Spiro SG: Small cell lung
cancer: Treatment review. Respirology. 11:241–248. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Atreya R, Mudter J, Finotto S, Müllberg J,
Jostock T, Wirtz S, Schütz M, Bartsch B, Holtmann M, Becker C, et
al: Blockade of interleukin 6 trans signaling suppresses T-cell
resistance against apoptosis in chronic intestinal inflammation:
Evidence in crohn disease and experimental colitis in vivo. Nat
Med. 6:583–588. 2000. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hodge DR, Hurt EM and Farrar WL: The role
of IL-6 and STAT3 in inflammation and cancer. Eur J Cancer.
41:2502–2512. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fesik SW: Promoting apoptosis as a
strategy for cancer drug discovery. Nat Rev Cancer. 5:876–885.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Nicholson DW: From bench to clinic with
apoptosis-based therapeutic agents. Nature. 407:810–816. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lüscher B: Function and regulation of the
transcription factors of the Myc/Max/Mad network. Gene. 277:1–14.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Grandori C, Cowley SM, James LP and
Eisenman RN: The Myc/Max/Mad network and the transcriptional
control of cell behavior. Annu Rev Cell Dev Biol. 16:653–699. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhu J, Blenis J and Yuan J: Activation of
PI3K/Akt and MAPK pathways regulates Myc-mediated transcription by
phosphorylating and promoting the degradation of Mad1. Proc Natl
Acad Sci USA. 105:6584–6589. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Goel A, Chhabra R, Ahmad S, Prasad AK,
Parmar VS, Ghosh B and Saini N: DAMTC regulates cytoskeletal
reorganization and cell motility in human lung adenocarcinoma cell
line: An integrated proteomics and transcriptomics approach. Cell
Death Dis. 3:e4022012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Stevenson LF, Sparks A, Allende-Vega N,
Xirodimas DP, Lane DP and Saville MK: The deubiquitinating enzyme
USP2a regulates the p53 pathway by targeting Mdm2. EMBO J.
26:976–986. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Oren M: Decision making by p53: Life,
death and cancer. Cell Death Differ. 10:431–442. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Pula G and Poole AW: Critical roles for
the actin cytoskeleton and cdc42 in regulating platelet integrin
alpha2beta1. Platelets. 19:199–210. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Georgiou M, Marinari E, Burden J and Baum
B: Cdc42, Par6 and aPKC regulate Arp2/3-mediated endocytosis to
control local adherens junction stability. Curr Biol. 18:1631–1638.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Mosesson Y, Mills GB and Yarden Y:
Derailed endocytosis: An emerging feature of cancer. Nat Rev
Cancer. 8:835–850. 2008. View Article : Google Scholar : PubMed/NCBI
|