Introduction
Esophageal cancer, including esophageal
adenocarcinoma and esophageal squamous cell carcinoma (ESCC), is
one of the most common types of cancer worldwide (1). In Asia, ESCC is the main type of
esophageal cancer (2). Due to the
lack of an early diagnostic method for ESCC, the mortality rate for
ESCC is very high; the 5-year overall survival rate is <25%
(3). Therefore, a detailed study on
the mechanisms underlying the development and progression of ESCC
is required, in order to improve the prevention, diagnosis and
treatment of this disease.
MicroRNAs (miRNAs/miRs) are a large class of small
non-coding RNAs that regulate gene expression by targeting mRNAs,
and induce mRNA degradation or translational suppression (4). In the human genome, >1,000 miRNAs
have been identified, which are thought to regulate ~30% of all
genes (5). It has previously been
reported that miRNA dysregulation is involved in the development
and progression of cancer (6).
Previous studies have revealed that miR-196a is upregulated in
various types of cancer, including ESCC (7–9). Further
studies have demonstrated that miR-196a promotes tumor progression
and acts as an oncogene in some types of cancer (10,11). For
example, it has been reported that miR-196a promotes cell
proliferation and invasion by targeting homeobox A5 in non-small
cell lung cancer (12). However, the
role of miR-196a in ESCC remains unclear.
The present study demonstrated that miR-196a was
significantly upregulated in human esophageal cancer tissue samples
and in the ESCC cell line EC109. In addition, downregulation of
miR-196a suppressed proliferation, invasion and migration of EC109
cells. The mechanism by which miR-196a affected the EC109 cell
phenotype was further investigated, and the results revealed that
miR-196a acted by targeting Annexin A1 (ANXA1). These results
suggested that miR-196a may be a potential therapeutic target in
ESCC.
Materials and methods
Clinical sample collection
ESCC and adjacent non-cancerous tissues were
harvested from 25 patients (age, 58.3±6.2 years; male, n=18;
female, n=7) from the Second Xiangya Hospital, Central South
University (Changsha, China) between June 2016 and December 2016.
All patients underwent esophageal cancer resection prior to
chemotherapy. After resection, tumor tissues and adjacent non-tumor
tissues were collected and stored at −80°C, and malignancy was
confirmed by pathologists. All experiments were approved by the
Ethics Committees of the Second Xiangya Hospital of Central South
University. All patients provided written informed consent for
their participation.
Cell culture
The human normal esophageal epithelial cell line
Het-1A and the ESCC cell line EC109 were purchased from Shanghai
Institutes for Biological Sciences (Shanghai, China). Het-1A cells
were cultured in Dulbecco's modified Eagle's medium (Gibco; Thermo
Fisher Scientific, Inc., Waltham, MA, USA), and EC109 cells were
cultured in RPMI-1640 medium (Gibco; Thermo Fisher Scientific,
Inc.). All media were supplemented with 10% fetal bovine serum
(FBS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml penicillin
and 100 µg/ml streptomycin. Cells were maintained at 37°C in a
humidified incubator containing 5% CO2.
Detection of miR-196a by reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
Total RNA from tissues, Het-1A cells and EC109 cells
was isolated with TRIzol® reagent (Invitrogen; Thermo
Fisher Scientific, Inc.) and were then converted to cDNA with a
PrimeScript RT Reagent kit (Takara Biotechnology Co., Ltd., Dalian,
China), according to the manufacturer's protocol. SYBR Premix Ex
Taq (Takara Biotechnology Co., Ltd.) was used for the RT-qPCR
assays to detect the expression levels of miR-196a in tissues and
cells. The qPCR reaction was run at 95°C for 10 min, followed by 40
cycles at 95°C for 15 sec for melting, and 60°C for 1 min for
annealing/extension. RNU6 was used as the endogenous control gene
to normalize the expression levels of miR-196a. The qPCR primers
for miR-196a and U6 were as follows: miR-196a, forward
5′-GCGCCCTAGGTAGTTTCATGTT-3′, reverse, 5′-GTGCAGGGTCCGAGGT-3′; and
U6, forward 5′-CTCGCTTCGGCAGCACA-3′ and reverse
5′-AACGCTTCACGAATTTGCGT-3′. Primers were purchased from YRgene Co.,
Ltd. (Changsha, China). The RT-qPCR assays were performed in
triplicate and alterations in miR-196a expression were calculated
using the 2−ΔΔCq method (ABI 7500 Software v2.0.1;
Applied Biosystems; Thermo Fisher Scientific, Inc.) (13).
Transfection of cells with the
miR-196a inhibitor
The miR-196a inhibitor (sense:
5′-CCCAACAACAUGAAACUACCUA-3′) and a negative control (NC) inhibitor
(sense: 5′-CAGUACUUUUGUGUAGUACAA-3′) were purchased from Shanghai
GenePharma Co., Ltd. (Shanghai, China). EC109 cells were plated in
6-well plates at 3×105 cells/well and were cultured for
24 h. The miR-196a inhibitor or inhibitor NC were transfected into
EC109 cells at a final concentration of 150 nM using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol. A
total of 48 h post-transfection, cells were harvested for further
analysis.
Cell proliferation assay
EC109 cells were seeded in 96-well plates at a
density of 5,000 cells/well. The miR-196a inhibitor or inhibitor NC
were then transfected into the cells at a final concentration of
150 nM. After 24-, 48- and 72-h transfection, cell proliferation
was measured using the MTT Assay kit (Beyotime Institute of
Biotechnology, Haimen, China), according to the manufacturer's
protocol. Each experiment was performed three times.
Flow cytometric analysis of
apoptosis
EC109 cell apoptosis was measured using an Annexin
V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis
Detection kit (Nanjing KeyGen Biotech Co., Ltd., Nanjing, China).
Briefly, 5×105 cells were harvested with 0.25% EDTA-free
trypsin, washed twice with PBS and resuspended in 500 µl binding
buffer. Cells were then incubated with 5 µl Annexin V-FITC and 5 µl
PI for 15 min at room temperature in the dark, and apoptosis was
detected with a NovoCyte Flow Cytometer (ACEA Biosciences, Inc.,
San Diego, CA, USA). Each sample was assessed in triplicate.
Transwell cell invasion and migration
assays
The invasive and migratory abilities of EC109 cells
were measured using Transwell assay. Briefly, cells were harvested
and resuspended in serum-free medium. For the migration assay,
2.5×104 cells were seeded into the upper chamber of a
Transwell insert placed in a 24-well plate (8 µm pore; Corning
Inc., Corning, NY, USA). For the invasion assay, 5×104
cells were seeded into the upper chamber of a Transwell insert
coated with Matrigel (BD Biosciences, San Jose, CA, USA). Medium
containing 10% FBS was added into the bottom chamber. After 24-h
incubation at 37°C, cells on the upper surface of the membrane were
removed, and the invaded or migrated cells on the lower surface of
the membrane were fixed with 100% methanol for 15 min at 4°C and
were then stained with 0.1% crystal violet for 15 min at room
temperature. The number of invaded or migrated cells was counted
under a light microscope.
Luciferase reporter assay
The possible target genes of miR-196a were predicted
with TargetScan 7.1 (http://www.targetscan.org) and ANXA1 was selected for
further analysis. A dual-luciferase miRNA target expression plasmid
containing the wild-type 3′-untranslated region fragment of ANXA1,
named pYr-MirTarget-ANXA1-3U, and a corresponding mutant reporter
plasmid containing the mutant miR-196a target site, named
pYr-MirTarget-ANXA1-3U-Mut, were purchased from YRgene Co., Ltd.
The miR-196a mimics (sense, 5′-UAGGUAGUUUCAUGUUGUUGGG-3′; and
antisense, 5′-CAACAACAUGAAACUACCUAUU-3′) and NC mimics (sense,
5′-UUCUCCGAACGUGUCACGUTT-3′; and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′) were purchased from Shanghai
GenePharma Co., Ltd. Briefly, 293 cells (Shanghai Institutes for
Biological Sciences) were seeded in 96-well plates at a density of
5×103 cells/well and were co-transfected with
pYr-MirTarget-ANXA1-3U or pYr-MirTarget-ANXA1-3U-Mut and miR-196a
mimics or mimics NC (50 nM) using Lipofectamine® 2000.
After 48-h transfection, cells were harvested and a luciferase
reporter assay was performed using the Dual-Luciferase Reporter
Assay kit (Promega Corporation, Madison, WI, USA), according to the
manufacturer's protocol. Each experiment was performed three
times.
Western blotting
Total protein was extracted from EC109 cells using
lysis buffer (Beyotime Institute of Biotechnology), and protein
concentrations were measured with a Bicinchoninic Acid Protein
Assay kit (Beyotime Institute of Biotechnology). The proteins (50
µg) were separated by 10% SDS-PAGE and were then transferred to a
0.22-µm nitrocellulose membrane (EMD Millipore, Billerica, MA,
USA). After blocking with 5% skimmed milk for 2 h at room
temperature, membranes were incubated with primary antibodies
against ANXA1 (cat. no. 32934S; Cell Signaling Technology, Inc.,
Danvers, MA, USA), cyclooxygenase 2 (COX2; cat. no. ab15191; Abcam,
Cambridge, MA, USA), matrix metalloproteinase (MMP)-2 (cat. no.
ab97779; Abcam), Snail (cat. no. 3879S; Cell Signaling Technology,
Inc.), E-cadherin (cat. no. 3195S; Cell Signaling Technology, Inc.)
and β-actin (cat. no AT0001; CMCTAG, Inc., Milwaukee, WI, USA) (all
dilutions, 1:1,000) overnight at 4°C with gentle agitation.
Membranes were further incubated for 1 h at room temperature with
the corresponding horseradish peroxidase-conjugated secondary
antibodies: Goat anti-rabbit secondary antibody for ANXA1, COX2,
MMP-2, Snail and E-cadherin (cat. no. sc-2004; 1:5,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) or goat anti-mouse secondary
antibody for β-actin (cat. no. sc-2005; 1:5,000; Santa Cruz
Biotechnology, Inc.). The protein bands were visualized using an
enhanced chemiluminescence system (Beyotime Institute of
Biotechnology) and were semi-quantified using ImageJ2× software
(National Institutes of Health, Bethesda, MD, USA). β-actin was
used as a loading control for normalization.
Transfection of cells with ANXA1 small
interfering (si)RNA
ANXA1 siRNA (sense, 5′-GAGAGAUCUGGCCAAAGACTT-3′; and
antisense, 5′-GUCUUUGGCCAGAUCUCUCTT-3′) or siRNA NC (sense,
5′-UUCUCCGAACGUGUCACGUTT-3′; and antisense,
5′-ACGUGACACGUUCGGAGAATT-3′) was purchased from Shanghai GenePharma
Co., Ltd. and was co-transfected into EC109 cells with miR-196a
inhibitor or inhibitor NC using Lipofectamine® 2000
(Invitrogen; Thermo Fisher Scientific, Inc.), according to the
manufacturer's protocol. The final concentrations of ANXA1 siRNA,
miR-196a inhibitor or inhibitor NC were 150 nM. After 48-h
transfection, cells were harvested, and western blotting, MTT and
Transwell assays were performed.
Effects of miR-196a mimics and ANXA1
siRNA
To confirm the effectiveness of miR-196a mimics, 293
cells were transfected with miR-196a mimics or mimics NC and were
harvested a total of 48 h post-transfection. Subsequently, the
expression levels of miR-196a were detected in 293 cells using
RT-qPCR. RNU6 was used as an endogenous control to normalize the
expression levels of miR-196a. Furthermore, to confirm the
knockdown efficiency of ANXA1 siRNA, EC109 cells were transfected
with ANXA1 siRNA or siRNA NC, and were harvested after 48 h of
transfection. Subsequently, the mRNA expression levels of ANXA1 in
EC109 cells were measured by RT-qPCR. GAPDH was used as an
endogenous control to normalize the mRNA expression levels of
ANXA1. The qPCR primers for ANXA1 and GAPDH were as follows: ANXA1
forward, 5′-CTAAGCGAAACAATGCACAGC-3′; and reverse,
5′-CCTCCTCAAGGTGACCTGTAA-3′; and GAPDH forward,
5′-GGAGCGAGATCCCTCCAAAAT-3′; and reverse,
5′-GGCTGTTGTCATACTTCTCATGG-3′.
Statistical analysis
Data are presented as the means ± standard deviation
of at least three independent experiments. The Student's t-test was
used for the comparison of two groups. Comparisons of means among
multiple groups were determined using one-way analysis of variance
followed by Tukey's post hoc test. All statistical analyses were
performed using SPSS 17.0 software (SPSS, Inc., Chicago, IL, USA).
P<0.05 was considered to indicate a statistically significant
difference.
Results
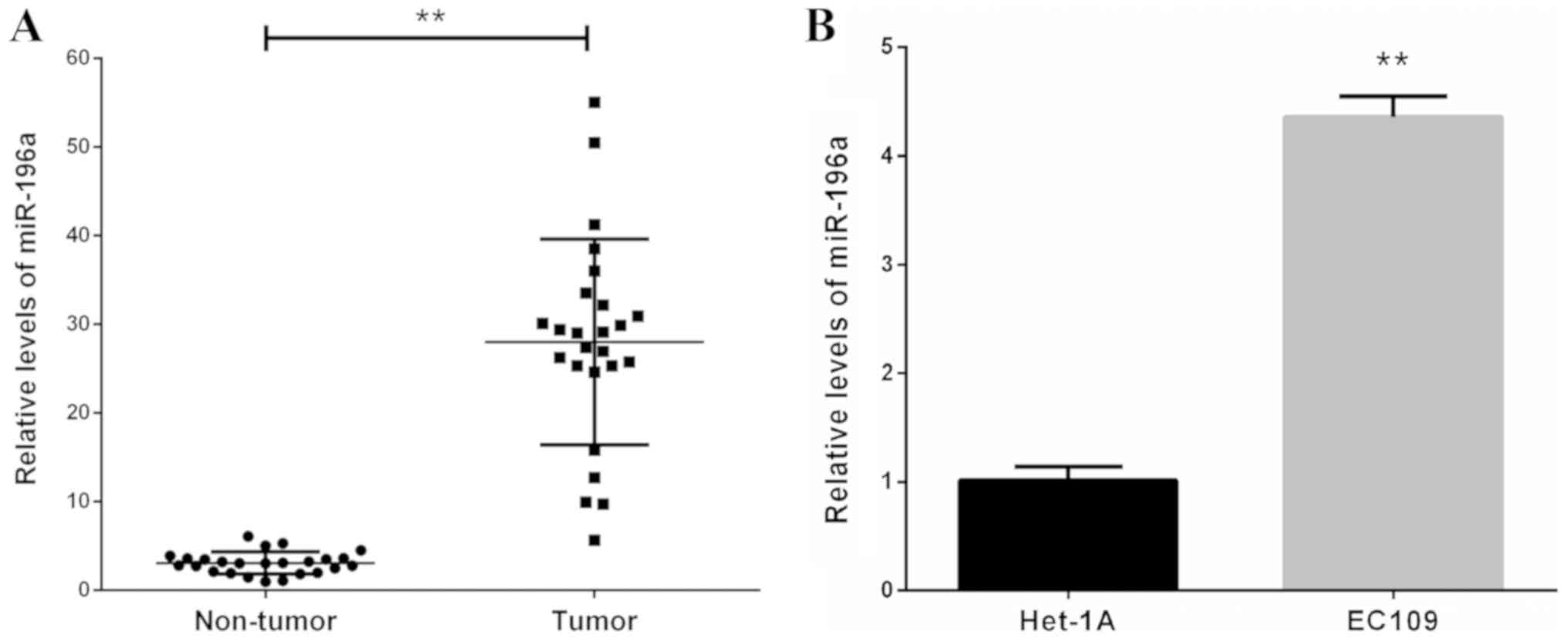
miR-196a is upregulated in human
esophageal cancer clinical tissues and ESCC cells
The expression levels of miR-196a in 25 human
esophageal cancer clinical tissues and corresponding non-cancerous
tissues were measured by RT-qPCR. The results revealed that the
expression levels of miR-196a were markedly upregulated in tumor
tissues compared with in adjacent non-tumor tissues (Fig. 1A). Furthermore, miR-196a expression
levels were significantly increased in the ESCC EC109 cell line
compared with in the Het-1A line cell (Fig. 1B). These data indicated that
upregulation of miR-196a may be involved in the development and
progression of ESCC.
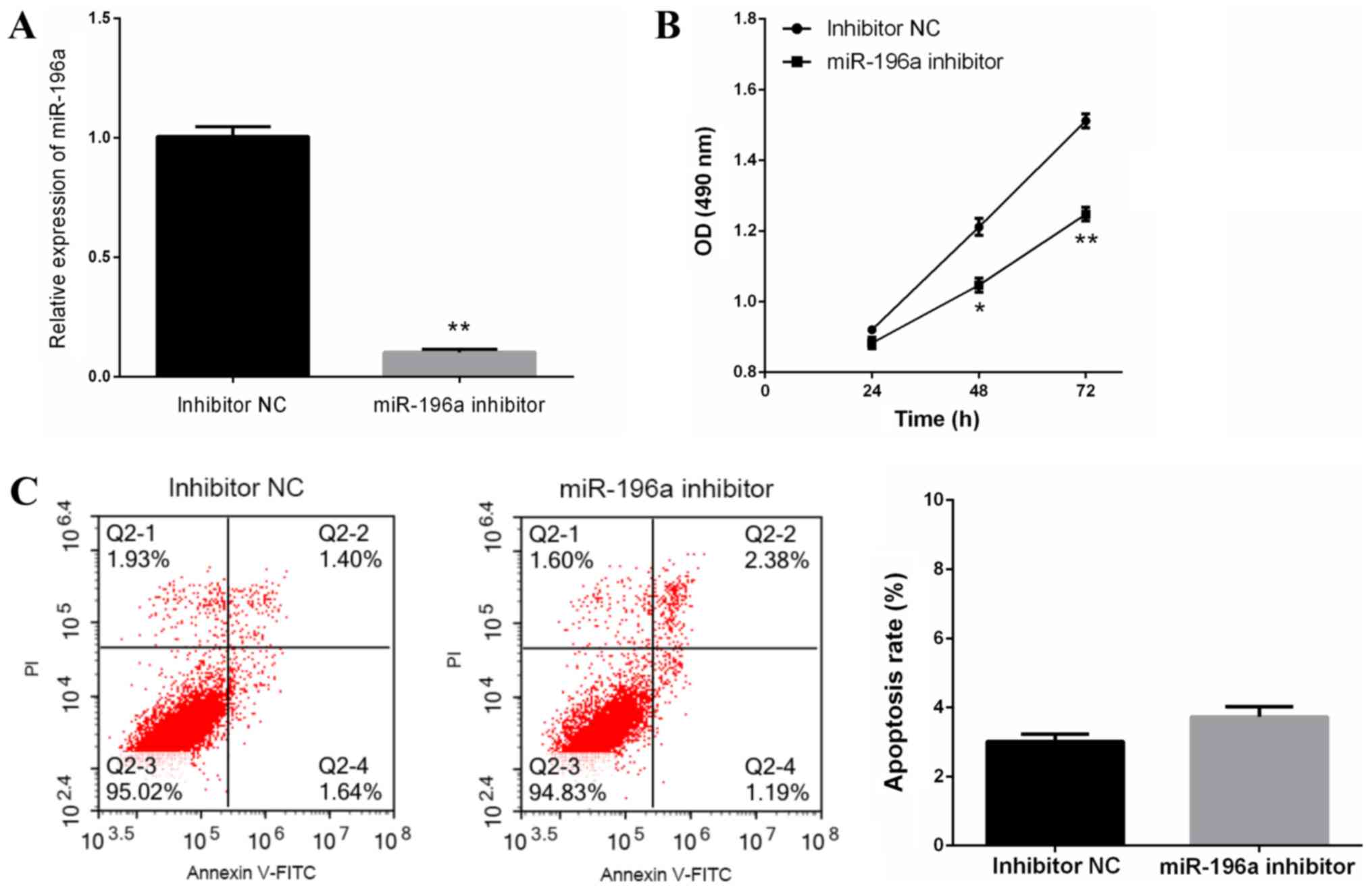
miR-196a knockdown suppresses cell
proliferation without affecting apoptosis of EC109 cells
To explore the role of miR-196a in ESCC progression,
miR-196a inhibitor or inhibitor NC were transfected into EC109
cells, and cell proliferation and apoptosis were measured. miR-196a
inhibitor-induced miR-196a knockdown in EC109 cells was confirmed
by RT-qPCR (Fig. 2A). The results of
the MTT assay revealed that miR-196a downregulation significantly
inhibited EC109 cell proliferation (Fig.
2B); however, flow cytometric analysis revealed that miR-196a
downregulation did not affect EC109 cell apoptosis (Fig. 2C). These data suggested that miR-196a
downregulation inhibited EC109 cell proliferation, but was not
associated with EC109 cell apoptosis.
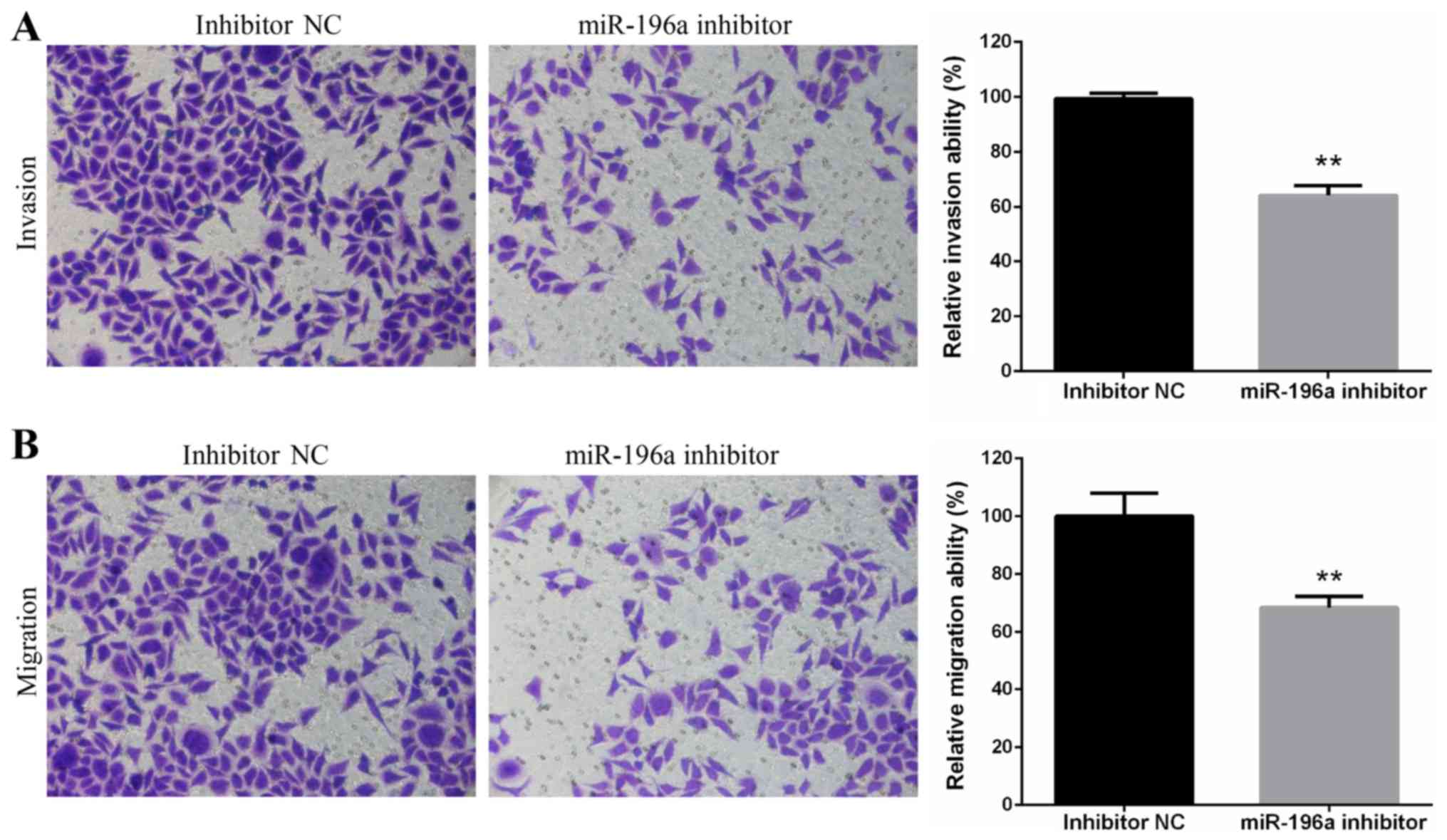
Downregulation of miR-196a suppresses
the invasion and migration of EC109 cells
The invasive and migratory ability of EC109 cells
transfected with miR-196a inhibitor or inhibitor NC were assessed
using Transwell invasion and migration assays. The results
demonstrated that miR-196a silencing significantly suppressed EC109
cell invasion (Fig. 3A) and
migration (Fig. 3B).
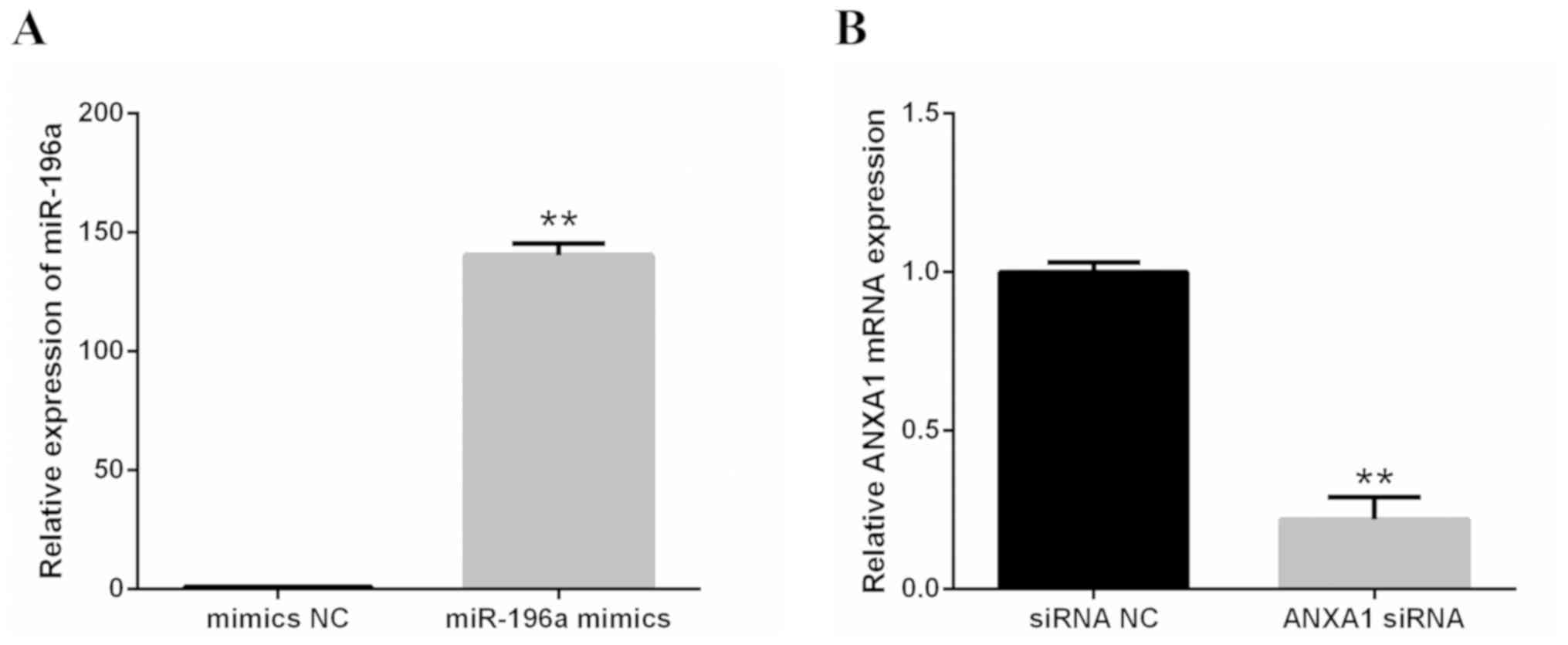
Effects of miR-196a mimics and ANXA1
siRNA
The effectiveness of miR-196a mimics and ANXA1 siRNA
were confirmed by RT-qPCR. The results revealed that miR-196a was
markedly upregulated by miR-196a mimics in 293 cells (Fig. 4A) and the mRNA expression levels of
ANXA1 were significantly silenced by ANXA1 siRNA in EC109 cells
(Fig. 4B).
miR-196a regulates the proliferation,
invasion and migration of EC109 cells by targeting ANXA1
To explore the molecular mechanism underlying the
regulatory effects of miR-196a on EC109 cell proliferation,
invasion and migration, possible target genes of miR-196a were
predicted with TargetScan 7.1. ANXA1 was selected for further
analysis, due to its complementary structure to miR-196a (Fig. 5A). A luciferase reporter assay was
performed to further confirm whether ANXA1 was a direct target gene
of miR-196a. pYr-MirTarget-ANXA1-3U or pYr-MirTarget-ANXA1-3U-Mut
and miR-196a mimics or mimics NC were co-transfected into 293
cells. The results of the luciferase activity assay demonstrated
that miR-196a mimics significantly reduced the luciferase activity
of pYr-MirTarget-ANXA1-3U, but did not affect the luciferase
activity of pYr-MirTarget-ANXA1-3U-Mut (Fig. 5B and C). In addition, western
blotting revealed that miR-196a knockdown via the miR-196a
inhibitor increased the protein expression levels of ANXA1 in EC109
cells (Fig. 5D). These results
indicated that ANXA1 may be a direct target gene of miR-196a. In
addition, following the increase in ANXA1 protein expression, COX2,
MMP-2 and Snail proteins were significantly downregulated, whereas
E-cadherin protein was markedly upregulated (Fig. 5D).
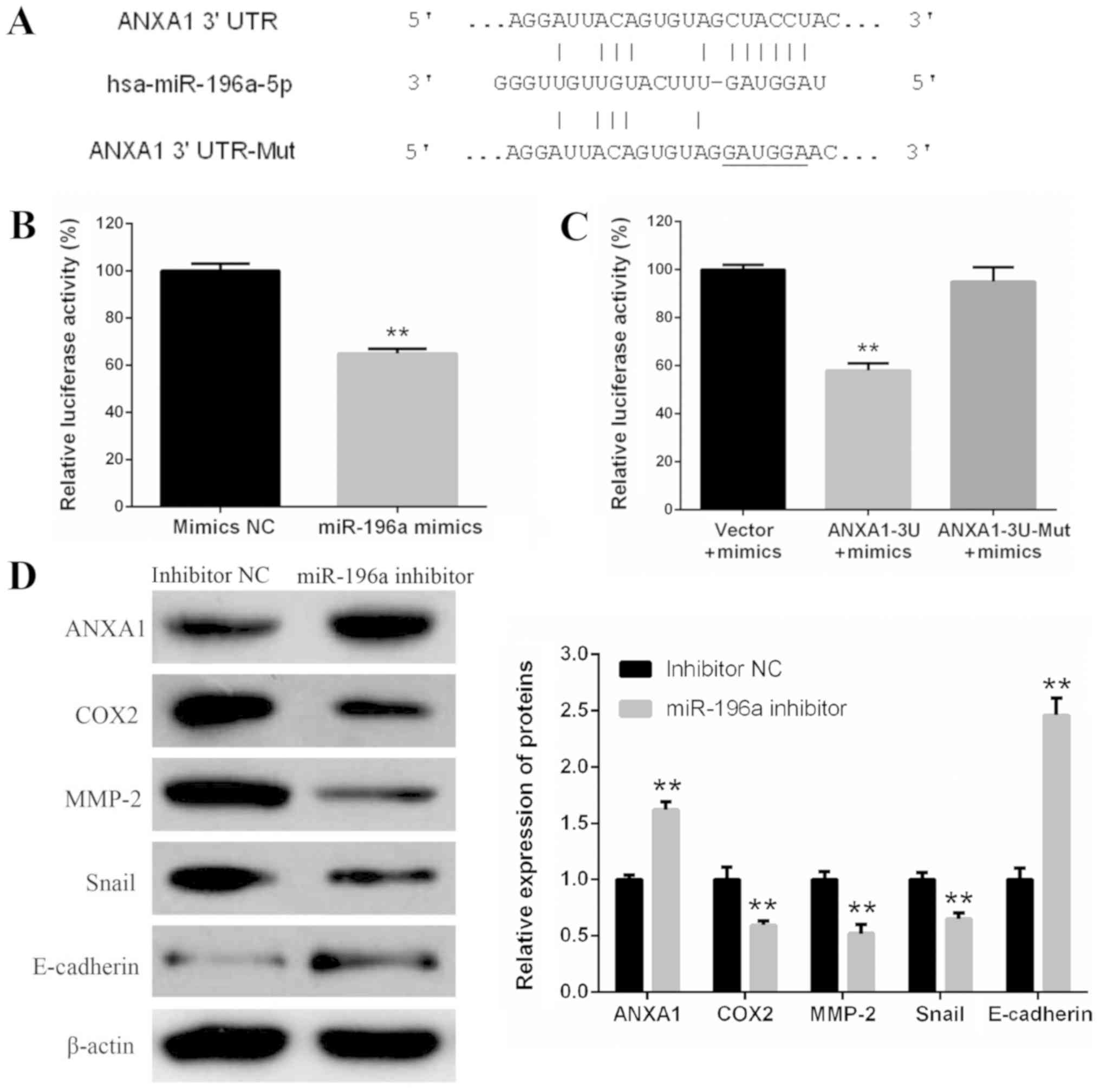 | Figure 5.ANXA1 is a direct target gene of
miR-196a. (A) Human ANXA1 3′UTR fragments containing a wild-type or
Mut miR-196a target site were cloned downstream of the luciferase
reporter gene. (B) 293 cells were co-transfected with
pYr-MirTarget-ANXA1-3U and miR-196a mimics or mimics NC. After 48-h
transfection, dual-luciferase activity was detected. **P<0.01
vs. mimic NC. (C) Luciferase activity assay revealed that miR-196a
mimics reduced the luciferase activity of pYr-MirTarget-ANXA1-3U,
but not pYr-MirTarget-ANXA1-3U-Mut. **P<0.01 vs. vector control.
(D) Western blotting of ANXA1, COX2, MMP-2, Snail and E-cadherin
protein expression in EC109 cells transfected with miR-196a
inhibitor or inhibitor NC. β-actin was used as a control.
**P<0.01 vs. NC. Experiments were performed in triplicate and
data are expressed as the means ± standard deviation. 3′UTR,
3′-untranslated region; ANXA1, Annexin A1; COX2, cyclooxygenase 2;
miR-196a, microRNA-196a; MMP2, matrix metalloproteinase 2; Mut,
mutant; NC, negative control. |
To confirm whether miR-196a regulated EC109 cell
proliferation, invasion and migration via ANXA1, ANXA1 protein
expression was knocked down via siRNA, whereas miR-196a was
inhibited via the miR-196a inhibitor (Fig. 6A). The results revealed that miR-196a
downregulation significantly inhibited EC109 cell proliferation,
invasion and migration; however, ANXA1 protein knockdown reversed
these inhibitory effects (Fig.
6B-D). These results suggested that miR-196a may regulate EC109
cell proliferation, invasion and migration by targeting ANXA1.
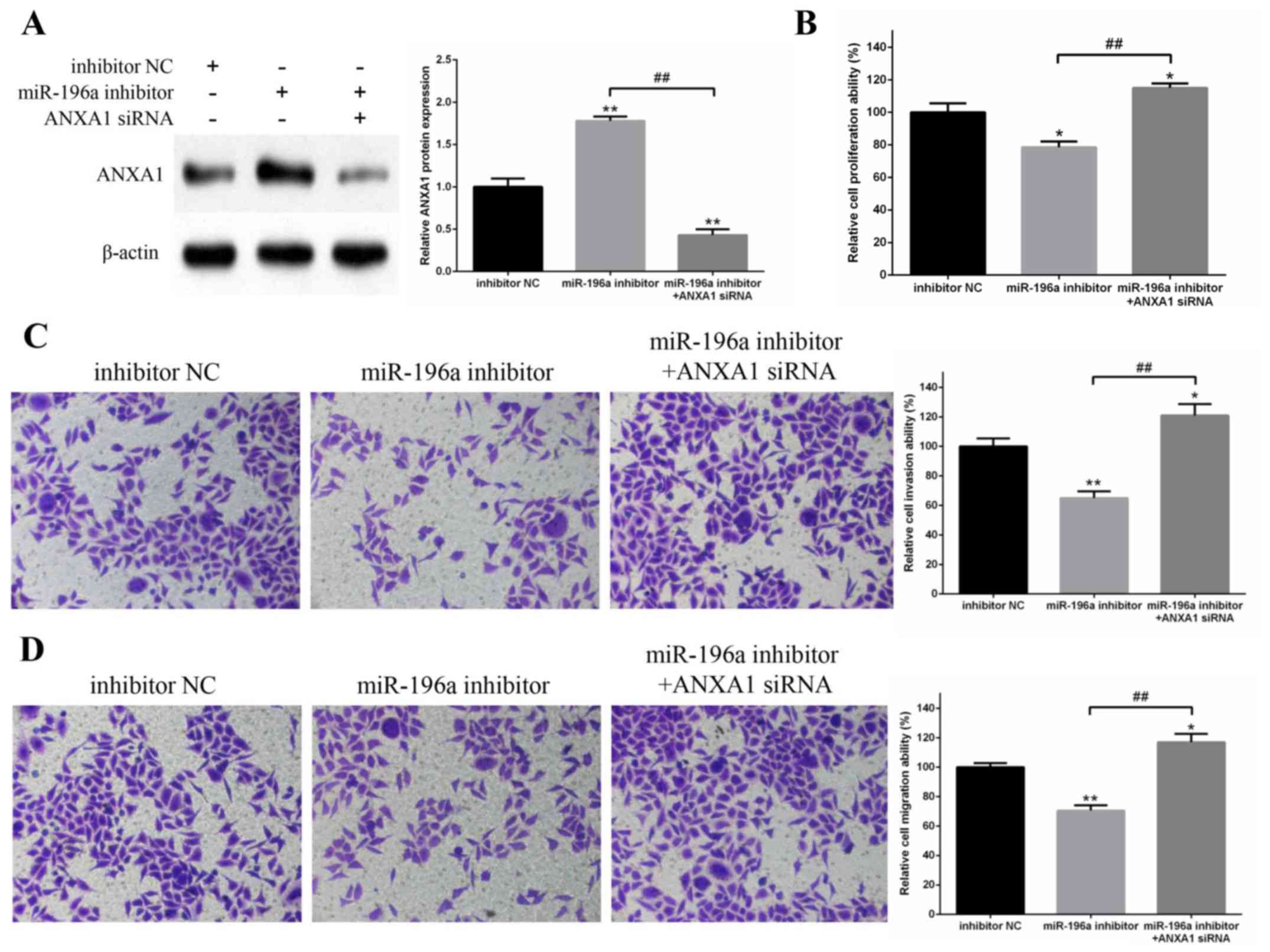 | Figure 6.Knockdown of ANXA1 can reverse the
inhibitory effects of miR-196a downregulation on the proliferation,
invasion and migration of EC109 cells. (A) Protein expression
levels of ANXA1 in EC109 cells were measured by western blotting.
β-actin was used as a control. The results demonstrated that ANXA1
protein expression was significantly upregulated by miR-196a
inhibitor and markedly downregulated by ANXA1 siRNA. (B)
Proliferation of EC109 cells was detected by MTT assay. The
miR-196a inhibitor suppressed the proliferation of EC109 cells,
whereas co-transfection with ANXA1 siRNA reversed this inhibitory
effect. (C) Invasive and (D) migratory abilities of EC109 cells
were determined by Transwell cell invasion and migration assays.
Magnification, ×200. Downregulation of miR-196a significantly
suppressed the invasion and migration of EC109 cells, whereas
knockdown of ANXA1 reversed these inhibitory effects. Experiments
were performed in triplicate and data are expressed as the means ±
standard deviation. *P<0.05, **P<0.01 vs. inhibitor NC;
##P<0.01, as indicated. ANXA1, Annexin A1; miR-196a,
microRNA-196a; NC, negative control; siRNA, small interfering
RNA. |
Discussion
Previous studies have reported that miR-196a is
upregulated in various types of cancer, including gastric cancer,
gastrointestinal stromal tumor, colon cancer, pancreatic cancer,
non-small-cell lung carcinoma, laryngeal cancer and cervical
carcinoma (14–16). Further studies have revealed that
miR-196a is likely to act as an oncogene in some types of cancer,
since it can promote cancer cell proliferation, invasion and
migration, and suppress tumor cell apoptosis (17,18). For
example, miR-196a promotes cell proliferation in gastric cancer by
targeting cyclin-dependent kinase inhibitor 1B (19). In colorectal cancer, miR-196a
stimulates the oncogenic phenotype of tumor cells (20). It has also been reported that
miR-196a inhibits cell apoptosis, and promotes cell proliferation
and invasion by targeting the inhibitor of growth family member 5
in pancreatic cancer (21).
Furthermore, Maru et al (18)
reported that miR-196a is upregulated in esophageal adenocarcinoma,
and Fendereski et al (9)
described an overexpression of miR-196a in ESCC. However, to the
best of our knowledge, few studies have examined the role of
miR-196a in regulation of the ESCC phenotype, and the role of
miR-196a in ESCC remains unclear.
The present study demonstrated that miR-196a was
markedly upregulated in ESCC tissues and EC109 cells compared with
in non-cancerous tissues and normal cells, respectively. These data
suggested that miR-196a may function as an oncogene in ESCC.
Furthermore, the regulatory role of a miR-196a inhibitor on EC109
cell phenotype was examined, and the results revealed that miR-196a
downregulation inhibited proliferation, invasion and migration of
EC109 cells. These results further suggested that miR-196a may be
involved in the development and progression of ESCC.
The molecular mechanisms underlying the regulatory
effects of miR-196a on the EC109 cell phenotype were investigated.
The results of the luciferase reporter assay and western blotting
demonstrated that ANXA1 was a direct target gene of miR-196a.
ANXA1, a member of the Annexin family, is a calcium-dependent
phospholipid binding protein (22).
It is involved in inflammation, cell proliferation, apoptosis,
tumorigenesis and tumor progression (23–25), and
is aberrantly expressed in various types of malignancy. It is
overexpressed in breast, liver and pancreatic cancer, whereas it is
downregulated in nasopharyngeal carcinoma, thyroid carcinoma,
prostatic cancer and ESCC (26–28).
Previous studies have demonstrated that ANXA1 is likely to function
as an anti-oncogene in some types of cancer, including ESCC. For
example, upregulation of ANXA1 expression inhibits cell growth and
induces cell apoptosis in DU145 human prostate cancer cells
(29). Xia et al (30) and Hu et al (31) reported that ANXA1 is significantly
downregulated in ESCC. Paweletz et al (32) discovered that a loss of ANXA1 is
involved in the tumorigenesis of esophageal and prostate carcinoma.
Álvarez-Teijeiro et al (33)
reported that ANXA1 is downregulated in head and neck squamous cell
carcinoma (HNSCC) and is a direct target of miR-196a/b in
HNSCC-derived cell lines; however, the authors did not investigate
the effects of ANXA1 or miR-196a/b on HNSCC cell phenotypes and the
corresponding molecular mechanism. In the present study, miR-196a
directly regulated the protein expression levels of ANXA1, and
miR-196a downregulation increased ANXA1 expression and inhibited
proliferation, invasion and migration of EC109 cells. Furthermore,
ANXA1 protein knockdown reversed the inhibitory effects of miR-196a
on EC109 cell proliferation, invasion and migration. These results,
combined with previous findings, suggested that miR-196a may
regulate the proliferation, invasion and migration of ESCC cells by
targeting ANXA1.
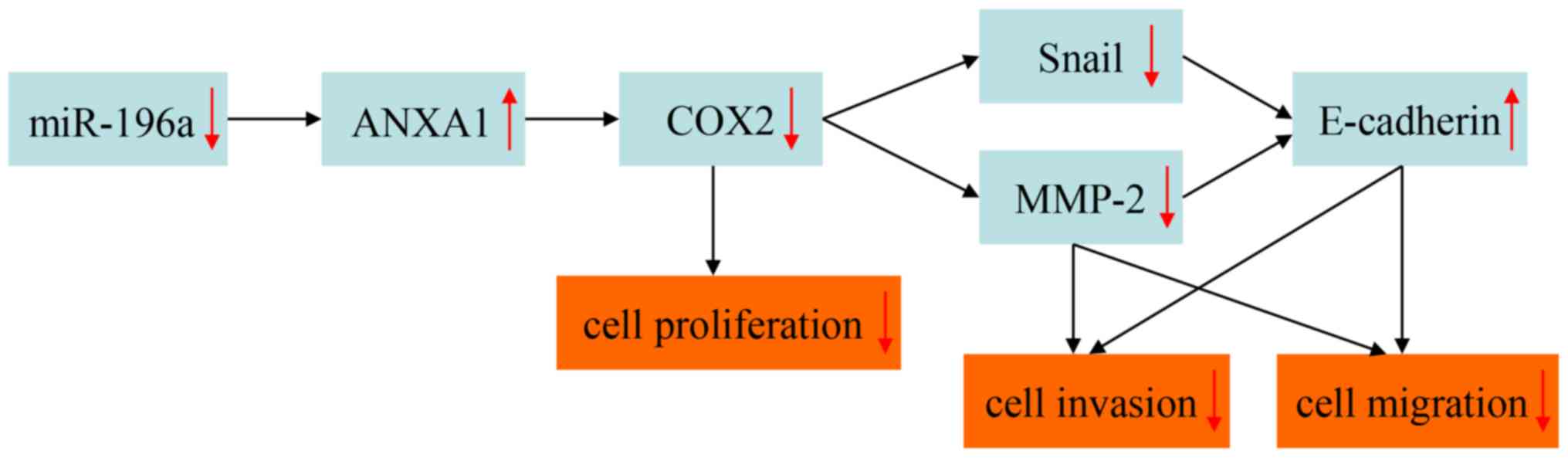
COX2, MMP-2 and Snail were downregulated and
E-cadherin was upregulated in EC109 cells transfected with miR-196a
inhibitor. These results further suggested that miR-196a may indeed
affect ESCC cell phenotypes.
COX2 is overexpressed in various types of cancer,
including ESCC, and is involved in the development and progression
of tumors (34). Numerous studies
reported that COX2 inhibition suppresses proliferation, invasion
and migration of tumor cells (35,36). Gao
et al (37) revealed that the
loss of COX2 following ANXA1 overexpression inhibits cell
proliferation and invasion in gastric cancer cells. Hannon et
al (38) and Croxtall et
al (39) also reported that
ANXA1 negatively regulates COX2 expression. In the present study,
COX2 expression was decreased alongside the upregulation of ANXA1
in EC109 cells. This finding suggested that miR-196a may regulate
cell proliferation in ESCC by targeting ANXA1, which may further
modulate COX2 expression.
E-cadherin is a cell adhesion molecule and a key
marker of epithelial-mesenchymal transition, which can suppress
tumor invasion and migration (40).
In addition, Snail can decrease the expression of E-cadherin by
binding to the E-box of the E-cadherin promoter (41). Furthermore, MMP-2 can promote the
invasion and migration of tumor cells by degrading the
extracellular matrix (42). MMPs can
also weaken the effect of E-cadherin by mediating E-cadherin
ectodomain shedding (43). Previous
studies have reported that MMP-2 and Snail are overexpressed in
various types of cancer, including ESCC, whereas E-cadherin is
downregulated (40–42). It has also been reported that COX2
downregulation decreases MMP-2 and Snail expression, and increases
E-cadherin expression, in tumor cells, and that these alterations
result in suppression of tumor cell invasion and migration
(44,45). These findings, combined with the
results of the present study, provided evidence supporting the
hypothesis that miR-196a may regulate the invasion and migration of
ESCC cells by targeting ANXA1, which may further modulate MMP-2,
Snail and E-cadherin expression by regulating COX2 expression.
In conclusion, the present study demonstrated that
miR-196a was upregulated in ESCC tumor tissues and cells, and may
function as an oncogene in ESCC. In addition, miR-196a
downregulation inhibited cell proliferation, invasion and
migration, but did not affect apoptosis of EC109 cells. The
molecular mechanism by which miR-196a may regulate EC109 cell
phenotypes was also investigated. The hypothetical mechanism of
regulation is presented in Fig. 7.
To the best of our knowledge, this mechanism has not yet been
reported in the literature. Further studies investigating the
associated cell signaling pathways are required. In particular,
whether miR-196a induces the degradation of ANXA1 mRNA and the
molecular mechanism underlying miR-196a-induced ANXA1 mRNA
degradation remain to be explored. Furthermore, the role of ANXA1
in regulating COX2 expression levels remains to be
investigated.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
CH and JP designed the study. CH, LL, XW and YZ
performed the experiments. JH and DL analyzed the data. CH wrote
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The Ethics Committees of the Second Xiangya
Hospital, Central South University approved the study, and all
patients provided written informed consent prior to their
inclusion.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Kamangar F, Qiao YL, Schiller JT, Dawsey
SM, Fears T, Sun XD, Abnet CC, Zhao P, Taylor PR and Mark SD: Human
papillomavirus serology and the risk of esophageal and gastric
cancers: Results from a cohort in a high-risk region in China. Int
J Cancer. 119:579–584. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lagergren J: Oesophageal cancer in 2014:
Advances in curatively intended treatment. Nat Rev Gastroenterol
Hepatol. 12:74–75. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Guo Y, Chen Z, Zhang L, Zhou F, Shi S,
Feng X, Li B, Meng X, Ma X, Luo M, et al: Distinctive microRNA
profiles relating to patient survival in esophageal squamous cell
carcinoma. Cancer Res. 68:26–33. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Chen K and Rajewsky N: The evolution of
gene regulation by transcription factors and microRNAs. Nat Rev
Genet. 8:93–103. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Fendler A, Jung M, Stephan C, Honey RJ,
Stewart RJ, Pace KT, Erbersdobler A, Samaan S, Jung K and Yousef
GM: miRNAs can predict prostate cancer biochemical relapse and are
involved in tumor progression. Int J Oncol. 39:1183–1192.
2011.PubMed/NCBI
|
|
7
|
Darda L, Hakami F, Morgan R, Murdoch C,
Lambert DW and Hunter KD: The role of HOXB9 and miR-196a in head
and neck squamous cell carcinoma. PLoS One. 10:e01222852015.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Li HL, Xie SP, Yang YL, Cheng YX, Zhang Y,
Wang J, Wang Y, Liu DL, Chen ZF, Zhou YN and Wu HY: Clinical
significance of upregulation of mir-196a-5p in gastric cancer and
enriched KEGG pathway analysis of target genes. Asian Pac J Cancer
Prev. 16:1781–1787. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Fendereski M, Zia MF, Shafiee M, Safari F,
Saneie MH and Tavassoli M: MicroRNA-196a as a potential diagnostic
biomarker for esophageal squamous cell carcinoma. Cancer Invest.
35:78–84. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Niinuma T, Suzuki H, Nojima M, Nosho K,
Yamamoto H, Takamaru H, Yamamoto E, Maruyama R, Nobuoka T, Miyazaki
Y, et al: Upregulation of miR-196a and HOTAIR drive malignant
character in gastrointestinal stromal tumors. Cancer Res.
72:1126–1136. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lu YC, Chang JT, Liao CT, Kang CJ, Huang
SF, Chen IH, Huang CC, Huang YC, Chen WH, Tsai CY, et al:
OncomiR-196 promotes an invasive phenotype in oral cancer through
the NME4-JNK-TIMP1-MMP signaling pathway. Mol Cancer. 13:2182014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu XH, Lu KH, Wang KM, Sun M, Zhang EB,
Yang JS, Yin DD, Liu ZL, Zhou J, Liu ZJ, et al: MicroRNA-196a
promotes non-small cell lung cancer cell proliferation and invasion
through targeting HOXA5. BMC Cancer. 12:3482012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hui AB, Shi W, Boutros PC, Miller N,
Pintilie M, Fyles T, McCready D, Wong D, Gerster K, Waldron L, et
al: Robust global micro-RNA profiling with formalin-fixed
paraffin-embedded breast cancer tissues. Lab Invest. 89:597–606.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kong X, Du Y, Wang G, Gao J, Gong Y, Li L,
Zhang Z, Zhu J, Jing Q, Qin Y and Li Z: Detection of differentially
expressed microRNAs in serum of pancreatic ductal adenocarcinoma
patients: miR-196a could be a potential marker for poor prognosis.
Dig Dis Sci. 56:602–609. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu P, Xin F and Ma CF: Clinical
significance of serum miR-196a in cervical intraepithelial
neoplasia and cervical cancer. Genet Mol Res. 14:17995–18002. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jin C, Zhang Y and Li J: Upregulation of
miR-196a promotes cell proliferation by downregulating
p27kip1 in laryngeal cancer. Biol Res. 49:402016.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Maru DM, Singh RR, Hannah C, Albarracin
CT, Li YX, Abraham R, Romans AM, Yao H, Luthra MG, Anandasabapathy
S, et al: MicroRNA-196a is a potential marker of progression during
Barrett's metaplasia-dysplasia-invasive adenocarcinoma sequence in
esophagus. Am J Pathol. 174:1940–1948. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sun M, Liu XH, Li JH, Yang JS, Zhang EB,
Yin DD, Liu ZL, Zhou J, Ding Y, Li SQ, et al: miR-196a is
upregulated in gastric cancer and promotes cell proliferation by
downregulating p27(kip1). Mol Cancer Ther. 11:842–852. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Schimanski CC, Frerichs K, Rahman F,
Berger M, Lang H, Galle PR, Moehler M and Gockel I: High miR-196a
levels promote the oncogenic phenotype of colorectal cancer cells.
World J Gastroenterol. 15:2089–2096. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu M, Du Y, Gao J, Liu J, Kong X, Gong Y,
Li Z, Wu H and Chen H: Aberrant expression miR-196a is associated
with abnormal apoptosis, invasion, and proliferation of pancreatic
cancer cells. Pancreas. 42:1169–1181. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Xiao Y, Ouyang C, Huang W, Tang Y, Fu W
and Cheng A: Annexin A1 can inhibit the in vitro invasive ability
of nasopharyngeal carcinoma cells possibly through Annexin
A1/S100A9/Vimentin interaction. PLoS One. 12:e01743832017.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sugimoto MA, Vago JP, Teixeira MM and
Sousa LP: Annexin A1 and the resolution of inflammation: Modulation
of neutrophil recruitment, apoptosis and clearance. J Immunol Res.
2016:82392582016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Boudhraa Z, Bouchon B, Viallard C, D'Incan
M and Degoul F: Annexin A1 localization and its relevance to
cancer. Clin Sci (Lond). 130:205–220. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Suh YE, Raulf N, Gäken J, Lawler K, Urbano
TG, Bullenkamp J, Gobeil S, Huot J, Odell E and Tavassoli M:
MicroRNA-196a promotes an oncogenic effect in head and neck cancer
cells by suppressing Annexin A1 and enhancing radioresistance. Int
J Cancer. 137:1021–1034. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Huang X, Wang Q, Li T, Li F and Lang J:
The negative expression of Annexin A1 in esophageal squamous cell
carcinoma is associated with poor outcome. Int J Radiat Oncol. 87
(Suppl):S288–S289. 2013. View Article : Google Scholar
|
|
27
|
Shen D, Nooraie F, Elshimali Y, Lonsberry
V, He J, Bose S, Chia D, Seligson D, Chang HR and Goodglick L:
Decreased expression of Annexin A1 is correlated with breast cancer
development and progression as determined by a tissue microarray
analysis. Hum Pathol. 37:1583–1591. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hsiang CH, Tunoda T, Whang YE, Tyson DR
and Ornstein DK: The impact of altered Annexin I protein levels on
apoptosis and signal transduction pathways in prostate cancer
cells. Prostate. 66:1413–1424. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mu D, Gao Z, Guo H, Zhou G and Sun B:
Sodium butyrate induces growth inhibition and apoptosis in human
prostate cancer DU145 cells by up-regulation of the expression of
Annexin A1. PLoS One. 8:e749222013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Xia SH, Hu LP, Hu H, Ying WT, Xu X, Cai Y,
Han YL, Chen BS, Wei F, Qian XH, et al: Three isoforms of Annexin I
are preferentially expressed in normal esophageal epithelia but
down-regulated in esophageal squamous cell carcinomas. Oncogene.
21:6641–6648. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hu N, Flaig MJ, Su H, Shou JZ, Roth MJ, Li
WJ, Wang C, Goldstein AM, Li G, Emmert-Buck MR and Taylor PR:
Comprehensive characterization of Annexin I alterations in
esophageal squamous cell carcinoma. Clin Cancer Res. 10:6013–6022.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Paweletz CP, Ornstein DK, Roth MJ, Bichsel
VE, Gillespie JW, Calvert VS, Vocke CD, Hewitt SM, Duray PH,
Herring J, et al: Loss of Annexin 1 correlates with early onset of
tumorigenesis in esophageal and prostate carcinoma. Cancer Res.
60:6293–6297. 2000.PubMed/NCBI
|
|
33
|
Álvarez-Teijeiro S, Menéndez ST,
Villaronga MÁ, Pena-Alonso E, Rodrigo JP, Morgan RO, Granda-Díaz R,
Salom C, Fernandez MP and García-Pedrero JM: Annexin A1
down-regulation in head and neck squamous cell carcinoma is
mediated via transcriptional control with direct involvement of
miR-196a/b. Sci Rep. 7:67902017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu X, Li P, Zhang ST, You H, Jia JD and
Yu ZL: COX-2 mRNA expression in esophageal squamous cell carcinoma
(ESCC) and effect by NSAID. Dis Esophagus. 21:9–14. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shao Y, Li P, Zhu ST, Yue JP, Ji XJ, He Z,
Ma D, Wang L, Wang YJ, Zong Y, et al: Cyclooxygenase-2, a potential
therapeutic target, is regulated by miR-101 in esophageal squamous
cell carcinoma. PLoS One. 10:e01406422015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Shao Y, Li P, Zhu ST, Yue JP, Ji XJ, Ma D,
Wang L, Wang YJ, Zong Y, Wu YD and Zhang ST: miR-26a and miR-144
inhibit proliferation and metastasis of esophageal squamous cell
cancer by inhibiting cyclooxygenase-2. Oncotarget. 7:15173–15186.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gao Y, Chen Y, Xu D, Wang J and Yu G:
Differential expression of ANXA1 in benign human gastrointestinal
tissues and cancers. BMC Cancer. 14:5202014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hannon R, Croxtall JD, Getting SJ,
Roviezzo F, Yona S, Paul-Clark MJ, Gavins FN, Perretti M, Morris
JF, Buckingham JC and Flower RJ: Aberrant inflammation and
resistance to glucocorticoids in Annexin 1−/− mouse.
FASEB J. 17:253–255. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Croxtall JD, Newman SP, Choudhury Q and
Flower RJ: The concerted regulation of cPLA2, COX2, and lipocortin
1 expression by IL-1beta in A549 cells. Biochem Biophys Res Commun.
220:491–495. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Dohadwala M, Yang SC, Luo J, Sharma S,
Batra RK, Huang M, Lin Y, Goodglick L, Krysan K, Fishbein MC, et
al: Cyclooxygenase-2-dependent regulation of E-cadherin:
Prostaglandin E(2) induces transcriptional repressors ZEB1 and
snail in non-small cell lung cancer. Cancer Res. 66:5338–5345.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen Z, Liu M, Liu X, Huang S, Li L, Song
B, Li H, Ren Q, Hu Z, Zhou Y and Qiao L: COX-2 regulates E-cadherin
expression through the NF-κB/Snail signaling pathway in gastric
cancer. Int J Mol Med. 32:93–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li Y, Ma J, Guo Q, Duan F, Tang F, Zheng
P, Zhao Z and Lu G: Overexpression of MMP-2 and MMP-9 in esophageal
squamous cell carcinoma. Dis Esophagus. 22:664–667. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
McGuire JK, Li Q and Parks WC: Matrilysin
(matrix metalloproteinase-7) mediates E-cadherin ectodomain
shedding in injured lung epithelium. Am J Pathol. 162:1831–1843.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Fujii R, Imanishi Y, Shibata K, Sakai N,
Sakamoto K, Shigetomi S, Habu N, Otsuka K, Sato Y, Watanabe Y, et
al: Restoration of E-cadherin expression by selective Cox-2
inhibition and the clinical relevance of the
epithelial-to-mesenchymal transition in head and neck squamous cell
carcinoma. J Exp Clin Cancer Res. 33:402014. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kurihara Y, Hatori M, Ando Y, Ito D,
Toyoshima T, Tanaka M and Shintani S: Inhibition of
cyclooxygenase-2 suppresses the invasiveness of oral squamous cell
carcinoma cell lines via down-regulation of matrix
metalloproteinase-2 production and activation. Clin Exp Metastasis.
26:425–432. 2009. View Article : Google Scholar : PubMed/NCBI
|