Introduction
Malignant glioma accounts for ~40% of all
intracranial tumors, which makes it the most common primary
malignant tumor of the central nervous system (1). Traditionally, gliomas are further
categorized as low-grade glioma (I–II) and high-grade glioma
(III–IV) according to the World Health Organization glioma grading
classification criteria (2).
Glioblastoma (GBM) belongs to the highest grade. Patients with
malignant glioma exhibit poor prognosis. However, currently there
are no well-established methods to inhibit glioma tumor growth. At
present, the comprehensive treatments for patients with glioma
include surgery, radiotherapy and chemotherapy (3). Temozolomide (TMZ) is a conventional
chemotherapy drug for glioma clinical therapy. TMZ has been
reported to exhibit therapeutic benefits in prolonging the survival
of patients with GBM (4). Although
some of the methods show promising results for glioma therapy, the
existence of multiple genetic changes in GBM suggests that a single
drug is incapable of offering a complete solution to the problem
(5,6). Genetic targeted therapy has been
reported to be a potential method of resolving the chemotherapy
resistance of cancer cells, and specific molecule inhibitors
against anomaly genetic targets are now in clinical trials
(7–9).
The nicotinamide-adenine dinucleotide
(NAD)-dependent protein deacetylase sirtuin 1 (SIRT1) is a key
regulator of life span (10–12). The biological function of SIRT1 is
linked to cellular stress, apoptosis and metabolism. SIRT1 has been
demonstrated to be involved in the tumorigenesis of numerous types
of cancer. SIRT1 influences the cell proliferation, differentiation
and apoptosis during tumor growth (13–16).
SIRT1 is overexpressed in several types of cancer (17–20).
However, the biological function of SIRT1 and its regulatory
mechanism in human glioma are rarely studied. In the present study,
the protective effects of SIRT1 on TMZ-induced toxicity were
investigated. The results revealed that SIRT1 was overexpressed in
glioma tissues and cell lines. The Kaplan-Meier analysis
demonstrated a significant association of SIRT1 expression with the
overall survival of patients with glioma. SIRT1 inhibition could
markedly sensitize glioma cell to TMZ in vitro and in
vivo. SIRT1 could regulate intracellular reactive oxygen
species (ROS) generation and the expression of Ki67 and p53 in
glioma. In summary, the present study demonstrated that SIRT1 was
essential in glioma proliferation and chemoresistance. SIRT1 may
serve as a potential therapeutic target for glioma therapy.
Materials and methods
Tissue samples
A total of 29 glioma tissues and 11 normal tissues
from sites adjacent to cancer sites were obtained from 24 men and
16 women aged between 35±2.5 and 65±3.2 years old. The tissues were
obtained as part of surgery carried out between December 2013 and
December 2017 at The First Affiliated Hospital of Shantou
University Medical College (Shantou, China). Tissues were stored at
−80°C prior to RNA or protein extraction. Informed consent from
relevant patients was obtained for tissue collection. The present
study was approved by the Ethics Committee of The First Affiliated
Hospital of Shantou University Medical College Institutional Review
Board (Shantou, China).
Cell lines and lentiviral
transfection
U251 and U87MG cell lines were purchased from the
Cell Bank of Chinese Academy of Sciences (Shanghai, China). A172,
T98 and HEB normal glial cell lines were purchased from the Chinese
Academy of Sciences Cell Bank (Shanghai, China). Short tandem
repeat (STR) profiling was used for cancer cell line testing. The
STR testing result revealed that the U87 cell line used in the
present study matched the American Type Culture Collection (ATCC)
U87MG cell line. Although the ATCC U87MG cell line is suspected to
not be the original GBM cell line from the University of Uppsala,
which was established in 1968, the ATCC U87MG cell line was derived
from the central nervous system, perhaps from another unknown
patient with glioma (21).
Therefore, the ATCC U87MG cell line was appropriate for use in the
present study. All cancer cell lines were cultured in Dulbecco's
modified Eagle's medium (DMEM; Gibco; Thermo Fisher Scientific,
Inc. Waltham, MA, USA), which was supplemented with 10% fetal
bovine serum (Gibco; Thermo Fisher Scientific, Inc.), 1% penicillin
and 1% streptomycin (Gibco; Thermo Fisher Scientific, Inc.). Cells
were cultured at 37°C in a humidified atmosphere with 5%
CO2. A total of 1×106 cells were transiently
transfected with the SIRT1 short hairpin RNA (shSIRT1) lentivirus
using the plent-U6-Puro lentiviral vector kit according to the
manufacturer's protocol (Thermo Fisher Scientific, Inc.).
Subsequently, 6 µg/ml polybrene (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) was used to enhance the transfection
efficiency. Following 24 h transfection, the suspension was
replaced by normal DMEM. U251-shSIRT1 and U87-shSIRT1 cells were
then screened using puromycin (2 µg/ml). After 7 days screening,
the stable U251-shSIRT1 and U87-shSIRT1 cell lines were used for
subsequent experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
For the RT-qPCR assay, total mRNA was extracted from
tumor tissues and cells using the Total RNA Extraction reagent
(Nanjing Sunshine Biotech Co., Ltd., Nanjing, China). Subsequently,
mRNA was reverse transcribed to cDNA using the Reverse TransScript
kit (Takara Bio, Inc., Otsu, Japan). The qPCR assay was performed
using the SYBR® Premix Ex Taq™ II kit (Takara Bio,
Inc.). All procedures were performed according to the
manufacturer's protocols. The RT-qPCR assay was performed using the
Bio-Rad iQ5 Real-Time PCR system (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA), according to the following protocol: Initial
activation step for 10 min at 95°C pre-denaturation, followed by 40
cycles of amplification, with 30 sec denaturation at 95°C, 10 sec
annealing at 60°C and 60 sec extension at 72°C. The primer
sequences used were as follows: SIRT1 forward,
5′-ATCTGACTTTGCTCCCCTTAACC-3′, and reverse,
5′-GGGCCCTGGTTGCAAGA-3′; β-actin was used as an internal reference,
and the primer sequences of β-actin were forward,
5′-CAATGACCCCTTCATTGACC-3′, and reverse, 5′-GACAAGCTTCCCGTTCTCAG-3.
The relative gene expression was quantified using the
2−ΔΔCq method (22).
Western blot analysis
Tissues and cells were lysed with
radioimmunoprecipitation assay buffer (Beyotime Institute of
Biotechnology, Shanghai, China), followed by determination of the
total protein concentration by bicinchoninic acid protein assay
(Beyotime Institute of Biotechnology). Total protein samples (20
µg/lane) were loaded and proteins with different molecular weights
were separated on a 10% SDS-PAGE gel. Proteins were transferred
onto polyvinylidene difluoride membranes following electrophoresis.
The membranes were blocked for 1 h at room temperature in 5%
non-fat milk. Subsequently, the membranes were incubated with
primary antibodies at 4°C overnight. The primary antibodies were
anti-SIRT1 (1:2,000 dilution; cat. no. ab110304; Abcam, Cambridge,
UK), anti-Ki67 antibody (1:2,000 dilution; cat. no. ab15580;
Abcam), anti-p53 antibody (1:2,000; cat. no. ab131442; Abcam) and
mouse anti-β-actin antibody (1:2,000; cat. no. ab8226; Abcam).
Following incubation, the membranes were washed with TBS containing
0.1% Tween-20, and incubated with horseradish peroxidase-conjugated
goat anti-mouse (1:4,000 dilution; cat. no. A0216; Beyotime
Institute of Biotechnology) an/d goat anti-rabbit (1:4,000
dilution; cat. no. A0208; Beyotime Institute of Biotechnology)
secondary antibodies at room temperature for 1 h. Subsequently,
membranes were soaked in enhanced chemiluminescence kit (cat. no.
P0018; Beyotime Institute of Biotechnology) for 1 min and
visualized under the chemiluminescence detection system
(ChemiDocXRS; Bio-Rad Laboratories, Inc.).
Cell proliferation assay
Cell proliferation was detected via water soluble
tetrazolium salts 1 (WST-1) assay. Cells (2,000/well) were seeded
in 96-well plates, following culturing for 0, 24, 48 and 72 h,
respectively. A total of 10 µl WST-1 (Dojindo Molecular
Technologies, Inc., Kumamoto, Japan) was added to each well and
incubated for 30 min. Subsequently, absorbance at 490 nm was
detected using a microplate reader (BioTek Instruments, Inc.,
Winooski, VT, USA), according to the manufacturer's protocols. All
assays were repeated at least three times.
TMZ toxicity assay
The SIRT1-specific inhibitor selisistat (EX527) was
purchased from Selleck Chemicals (Houston, TX, USA). Briefly, 200
nM EX527 was added to glioma cell culture medium. Glioma cells were
further cultured at 37°C for 24, 48 and 72 h. U87 and U251 cells
were exposed to TMZ to induce cell death. In order to define TMZ
toxicity, different concentrations of TMZ (0, 50, 100, 150 and 200
µM) were added to 2×106 U87 and U251 cells for 24 h,
prior to detect cell viability with WST-1 assay. The results
allowed determining 100 µM as the working concentration for TMZ.
Cells were then incubated with 100 µM TMZ at 37°C for 0, 24, 48 and
72 h. Subsequently, all the cells treated with EX527 and TMZ were
incubated in WST-1 following the manufacturer's protocol. TMZ (200
µM) was used in the time dependent cell viability assay. All assays
were repeated at least three times.
ROS determination assay
A total of 1×106 cells were cultured in
6-well plates. Prior to ROS detection, cells were washed twice with
PBS. Subsequently, cells were incubated with 5 µM dihydroethidium
(DHE; Vigorous Biotechnology Beijing Co., Ltd., Beijing, China) in
PBS for 30 min at 37°C in the dark. Following incubation, the cells
were washed twice with PBS and fixed in 4% paraformaldehyde for 30
min at room temperature. Following two washes with PBS, images were
captured by fluorescence microscopy (×40 magnification). At least
five images were captured for each group. Relative fluorescence
intensity was calculated by ImageJ software 7.0 (National
Institutes of Health, Bethesda, MD, USA).
Animal studies
A total of 20 nude mice (age, 4 weeks) were
purchased from Vital River Laboratories Co., Ltd. (Beijing, China).
All mice (22±1.5 g) were housed at a controlled temperature
(22–28°C) and humidity (60%) under a 12-h light/dark cycle. Mice
had free access to food and water. The stably transfected negative
control (NC) and shSIRT1 lentivirus U87 cells (2×106 for
each side) were suspended in PBS and implanted subcutaneously into
male Balb/c nude mice. A total of 10 mice received NC lentivirus
and 10 received shSIRT1 lentivirus (subcutaneous injection).
Treatment with 200 nM EX527 was performed for all mice, whereas
treatment with 100 µM TMZ was performed for the 10 normal mice. The
length (a) and width (b) of the tumors were monitored every 2 days.
After 30 days of monitoring, all nude mice were sacrificed. Tumor
volume (V) was calculated using the formula V=ab2/2.
Animal experiments conducted in the present study were approved by
the Ethics Committee of The First Affiliated Hospital of Shantou
University Medical College Institutional Review Board.
Statistical analysis
All the data were expressed as the means ± standard
error of the mean from at least three independent experiments. SPSS
17.0 software (SPSS, Inc., Chicago, IL, USA) was used for data
analysis. Statistical comparisons among different groups were
performed by one-way analysis of variance, followed by Newman-Keuls
comparison test. Kaplan-Meier method was used for the survival
analysis, and Cox regression analysis was used for the univariate
and multivariate analyses. P<0.05 was considered to indicate a
statistically significant difference.
Results
SIRT1 expression is upregulated in
glioma tissues and cell lines
To investigate the role of SIRT1 in glioma cell
chemotherapeutic agent resistance, the expression level of SIRT1 in
glioma tissues and cells was detected. The results revealed that
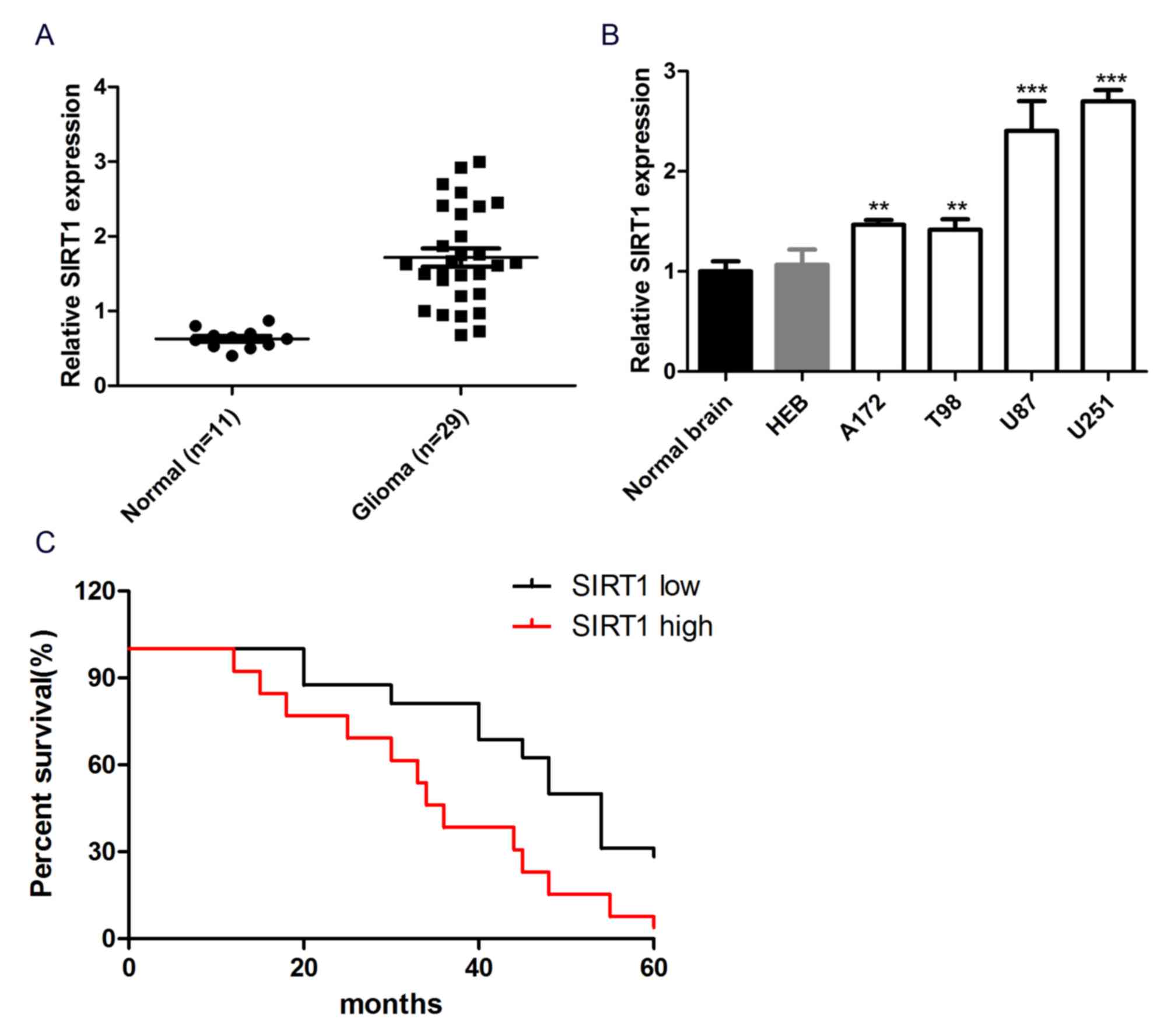
the mRNA expression level of SIRT1 was significantly increased in
29 glioma tissues compared with 11 normal brain tissues (Fig. 1A). In addition, SIRT1 exhibited
significantly higher expression in glioma cell lines (A172, T98,
U87 and U251) than normal human brain glial cell lines (HEB) and
normal brain tissue (Fig. 1B). These
data reflected SIRT1 upregulation in glioma tissues and cell lines.
Kaplan-Meier analysis was performed to study the association
between SIRT1 expression and the prognosis of patients with glioma.
Patients were divided into SIRT1 high and SIRT1 low groups, based
on the mean expression levels of SIRT1 in all samples. The
Kaplan-Meier survival curve revealed that patients with glioma with
high SIRT1 expression had a shorter overall survival than those
with low SIRT1 expression (P<0.05; Fig. 1C), indicating that SIRT1 expression
was associated with the prognosis of patients with glioma. Overall,
SIRT1 exhibited high expression in glioma tissues and cells and was
associated with the prognosis of patients with glioma.
SIRT1 knockdown inhibits glioma cell
proliferation
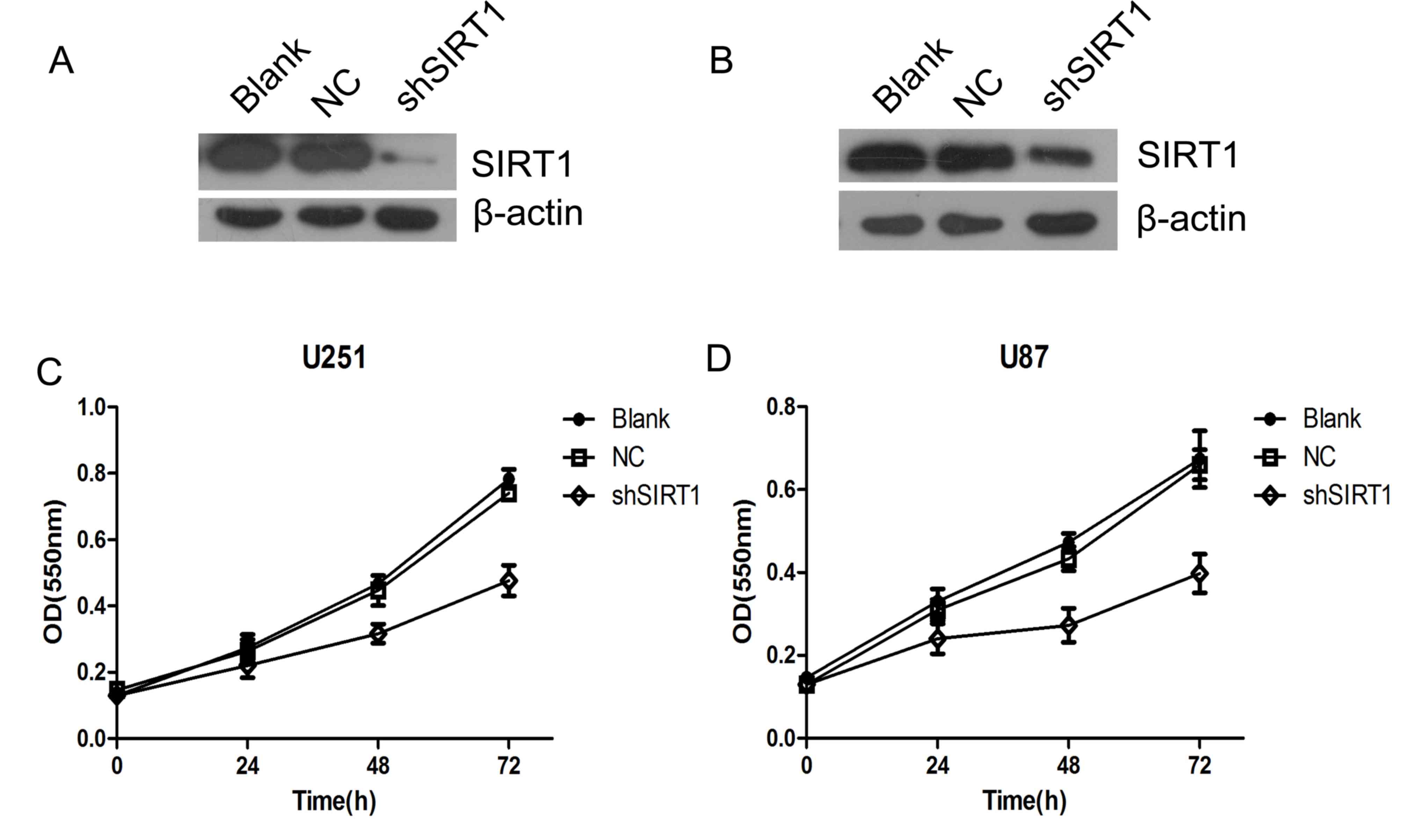
To investigate the biological function of SIRT1 in
glioma growth, SIRT1 was silenced using two short hairpin RNAs in
U251MG (Fig. 2A) and U87MG (Fig. 2B) cells. The proliferation of U87 and
U251 cells were detected by WST-1 cell viability assay. The results
revealed that SIRT1 knockdown significantly inhibited glioma cell
proliferation in U87 and U251 cells (Fig. 2C and D). This suggested that SIRT1 is
essential for glioma cell proliferation.
TMZ downregulates the expression of
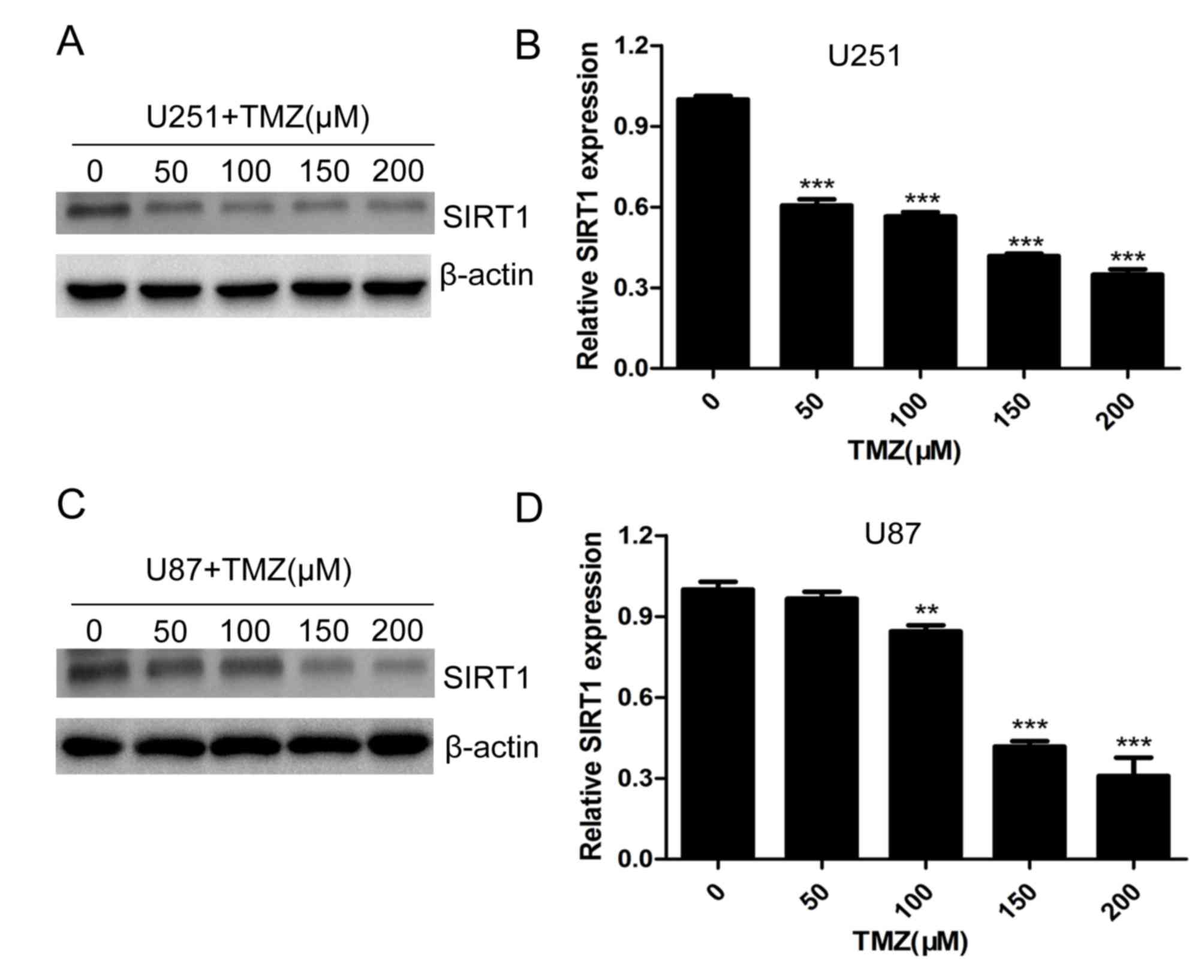
SIRT1 in glioma cells
Temozolomide is one of the most important drugs used
in clinical therapy of glioma (23,24). To
investigate the role of SIRT1 in glioma cell chemotherapeutic agent
resistance, U251 and U87 cells were treated with different
concentrations of TMZ. The protein and mRNA levels of SIRT1 were
significantly decreased with increasing concentrations of TMZ in
U251 (Fig. 3A and B) and U87
(Fig. 3C and D) cells, indicating
that SIRT1 expression was regulated by TMZ treatment.
SIRT1 inhibition increases TMZ
sensitivity of glioma cells
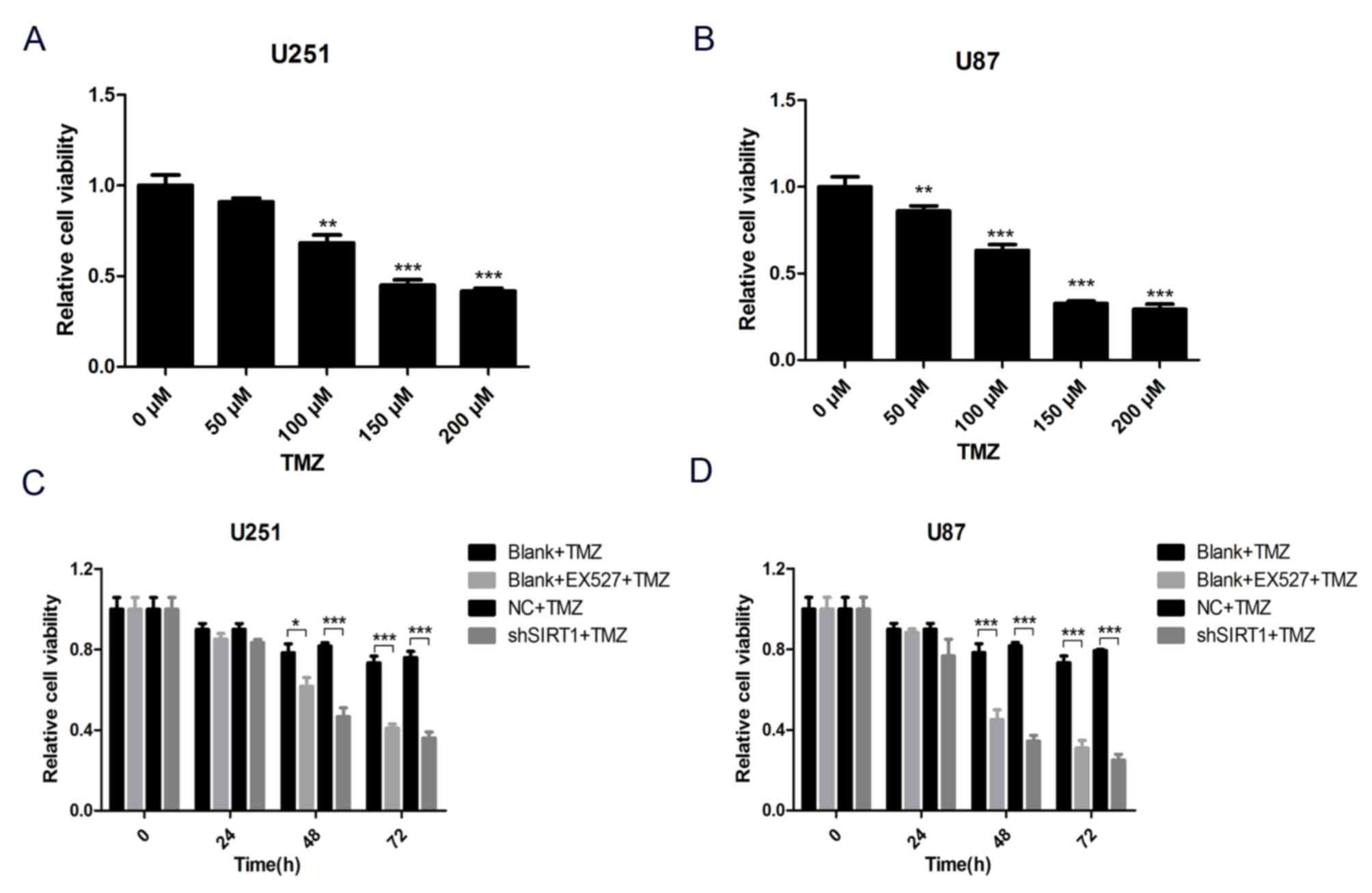
To comprehensively evaluate the association between
SIRT1 and glioma cell TMZ sensitivity, a SIRT1 specific inhibitor,
EX527, was used to inhibit SIRT1 protein activity in U251 and U87
cells. Subsequently, cells were treated with different
concentrations of TMZ for 24 h and cell viability was assessed by
WST-1 assay. The results demonstrated that cell viability decreased
with TMZ treatment in U251 and U87 cells (Fig. 4A and B). To avoid any extra toxicity
TMZ may induce, 100 µM was selected as the working concentration of
TMZ in subsequent tests. The result of the WST-1 assay demonstrated
that SIRT1 activity inhibition markedly decreased viability of U251
and U87 cells treated with TMZ, as did SIRT1 knockdown (Fig. 4C and D). These results demonstrated
that SIRT1 was indispensable for glioma cell TMZ sensitivity.
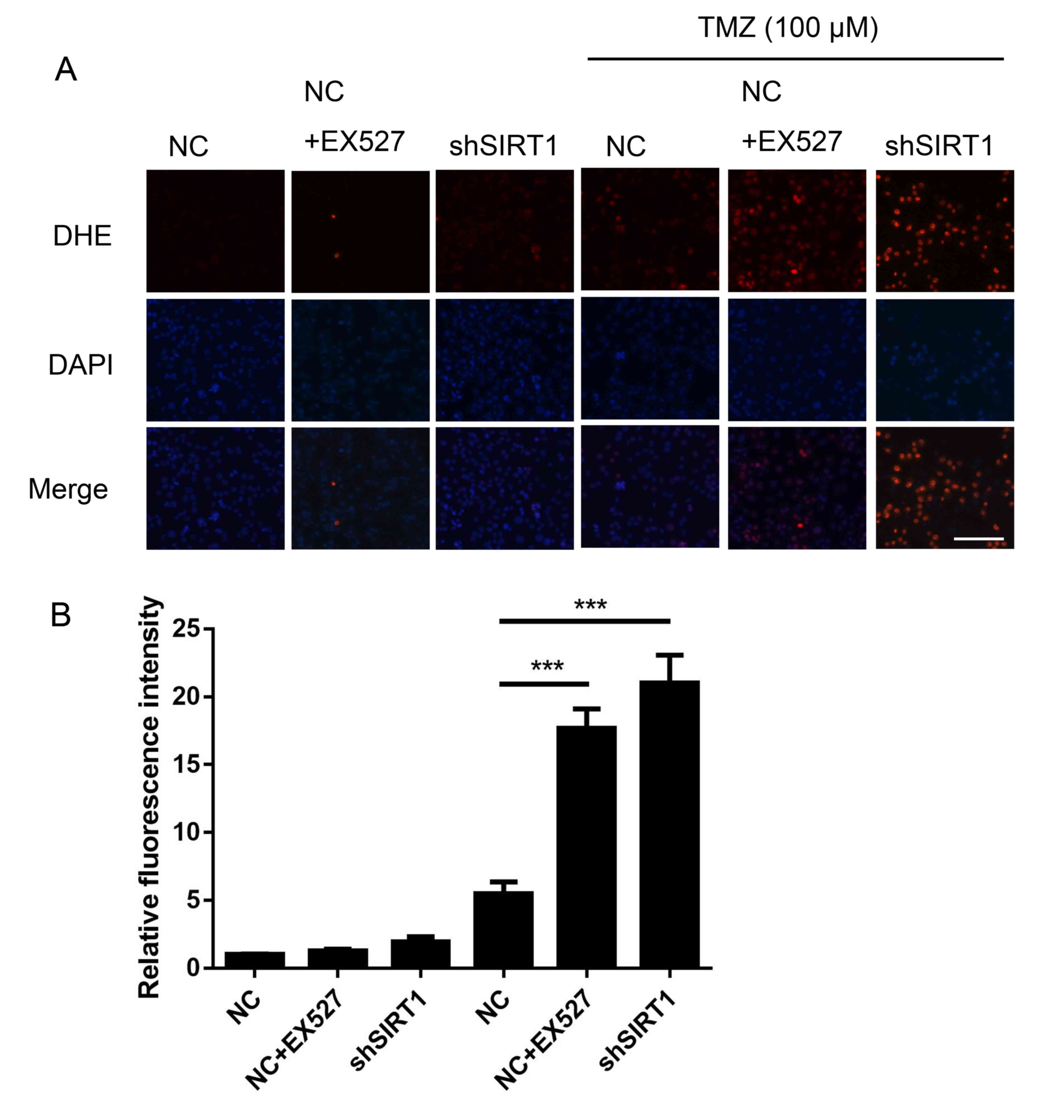
SIRT1 inhibition increases ROS
generation in glioma
TMZ has been reported to increase ROS production
(3), suggesting that ROS are a key
factor in TMZ mediated glioma clinical therapy. DHE staining was
performed to assess the association between SIRT1 and intracellular
ROS generation in glioma. As shown in Fig. 5, shSIRT1 significantly increased the
ROS level in U251 cells treated with TMZ, by ~4-fold compared with
only TMZ treatment. Simultaneously, EX527 increased the ROS level
in U251 cells (Fig. 5). The results
suggested that SIRT1 was an important regulator for glioma cell ROS
generation.
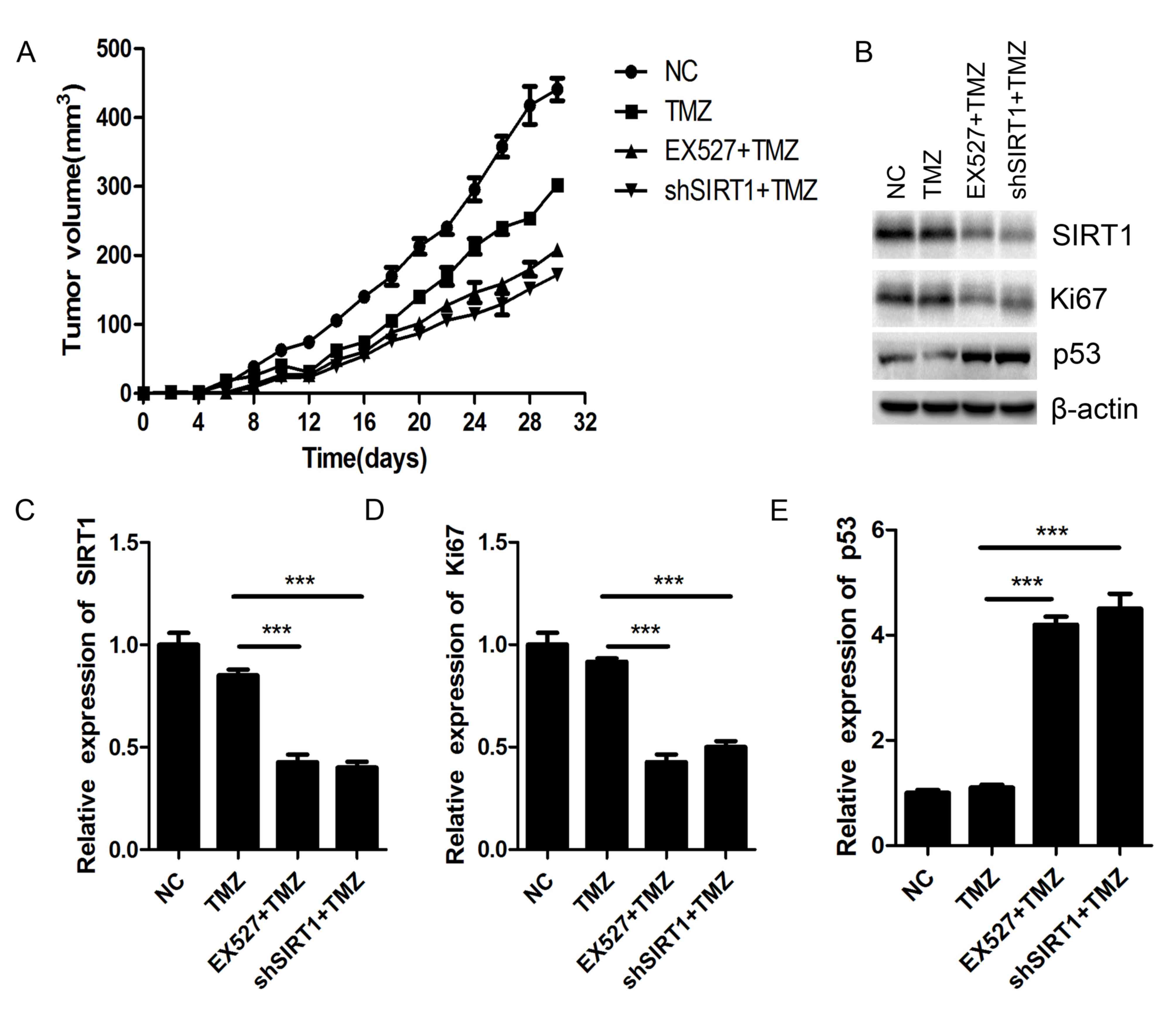
SIRT1 inhibition sensitizes glioma to
TMZ in vivo
Nude mice xenograft was performed to investigate the
biological effect of SIRT1 on glioma chemosensitivity in
vivo. The SIRT1 knockdown U87MG cells and the NC U87MG cells
were implanted into nude mice subcutaneously. After 30 days of TMZ
(100 µM) treatment, the tumor volumes of shSIRT1 U87MG
cells-implanted mice were smaller than those of NC U87MG
cells-implanted mice. Simultaneously, EX527 treatment exhibited
similar inhibitory effects on glioma tumor growth as shSIRT1
(Fig. 6A). Both shSIRT1 and EX527
treatment significantly decreased SIRT1 expression in the mice
(Fig. 6B and C). The protein level
of Ki67 greatly decreased in SIRT1 knockdown and EX527 treated
tumors compared with the NC tumors (Fig.
6B and D), while p53 expression was markedly increased in SIRT1
knockdown and EX527 treated mice tumors (Fig. 6B and E). As Ki67 is closely
associated with cell proliferation and p53 is the most important
tumor suppressor, SIRT1 may regulate glioma proliferation by
regulating the expression of Ki67 and p53.
Discussion
During past decades, malignant glioma has been one
of the most common primary malignant tumors of the brain. Although
techniques for the clinical therapy of glioma have developed in
previous years, the prognosis of patients with glioma remains poor
(25). The high incidence and low
survival rate of patients with glioma prompts exploration of novel
therapeutic strategies. Molecularly targeted therapy is considered
to be a potential approach. The identification of novel molecular
targets is currently being investigated extensively (26).
SIRT1 is a NAD-dependent protein deacetylase
involved in numerous pivotal biological processes, including energy
metabolism, DNA repair and mitochondrial homeostasis (27). These biological processes are crucial
for cell growth, differentiation, migration and survival, which
makes SIRT1 a key regulator in numerous diseases (28). At present, the role of SIRT1 in
cancer has been preliminarily studied, and SIRT1 has been reported
as an oncogene in a number of cases (29,30). In
the present study, tissue analysis demonstrated that SIRT1 was
overexpressed in glioma tissues compared with non-tumor tissues.
Additionally, SIRT1 was overexpressed in glioma cell lines compared
with normal brain cells. The Kaplan-Meier survival analysis
demonstrated that high expression levels of SIRT1 were associated
with poor prognosis. These findings demonstrated that SIRT1 may
serve a vital role in glioma tumorigenesis.
Currently, TMZ is the most efficient chemotherapy
agent for clinical therapy of GBM. It can extend the median
survival rate by 12–15 months (31).
However, long-term TMZ treatment may induce cancer cell resistance.
Damage to glioma cells, induced by TMZ, can be repaired by
O6-Methylguanine-DNA methyltransferase (MGMT), thus
inducing cancer cell resistance, while methylation of the MGMT
promoter leads to an increase in TMZ sensitivity (32). Additional molecular mechanisms of TMZ
resistance in glioma cells remain unknown. In the present study,
SIRT1 knockdown or SIRT1 activity inhibition greatly sensitized
glioma cells to TMZ treatment in vitro and in vivo,
indicating that SIRT1 was an important regulator of TMZ resistance
in glioma cells. Notably, the glioma cell lines treated with TMZ
exhibited MGMT promoter hypermethylation status (33). An intracellular ROS detection assay
suggested that SIRT1 knockdown or treatment with SIRT1 inhibitor
significantly increased glioma cell ROS level following treatment
with TMZ, indicating that SIRT1 was closely associated with
intracellular ROS generation.
In conclusion, SIRT1 was demonstrated to be
overexpressed in glioma tissues and cells and SIRT1 may regulate
glioma TMZ sensitivity by regulating intracellular ROS generation.
The present study illuminated the role and regulatory mechanism of
SIRT1 in glioma tumor growth and chemotherapy resistance. SIRT1 may
serve as a novel promising therapeutic target for clinical therapy
of glioma in the future.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
YMX, HWC and RL conceived and designed the
experiments. HWC, RL, ZWZ, JHH and YMX developed the methodology.
HWC, RL, ZHZ and QTW performed the experiments. HWC, RL, ZHZ and
QTW performed the analysis and interpretation of the data. HWC and
RL wrote, reviewed and revised the manuscript. YMX supported study
supervision. All authors have read and approved the manuscript.
Ethics approval and consent to
participate
Informed consent from relevant patients was obtained
for all tissue sample obtaining procedures. The tissue sample
obtaining procedures were approved by the ethics committee of The
First Affiliated Hospital of Shantou University Medical College
Institutional Review Board (Shantou, China). Animal experiments
conducted in the present study were approved by the Ethics
Committee of First Affiliated Hospital of Shantou University
Medical College Institutional Review Board.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Dunn GP, Rinne ML, Wykosky J, Genovese G,
Quayle SN, Dunn IF, Agarwalla PK, Chheda MG, Campos B, Wang A, et
al: Emerging insights into the molecular and cellular basis of
glioblastoma. Genes Dev. 26:756–784. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: The 2007
WHO classifcation of tumours of the central nervous system. Acta
Neuro Pathol. 114:97–109. 2007.
|
|
3
|
Cloughesy TF, Cavenee WK and Mischel PS:
Glioblastoma: From molecular pathology to targeted treatment. Annu
Rev Pathol. 9:1–25. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer Clin. 67:7–30. 2017. View Article : Google Scholar
|
|
5
|
Liao AY, Shi RR, Jiang YL, Tian S, Li P,
Song F, Qu Y, Li J, Yun H and Yang X: SDF-1/CXCR4 axis regulates
cell cycle progression and epithelial-mesenchymal transition via
up-regulation of survivin in glioblastoma. Mol Neurobiol.
53:210–215. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Altieri R, Fontanella M, Agnoletti A,
Pnciani PP, Spena G, Crobeddu E, Pilloni G, Tardivo V, Lanotte M,
Zenga F, et al: Role of nitric oxide in glioblastoma therapy:
Another step to resolve the terrible puzzle. Transl Med UniSa.
12:54–59. 2014.PubMed/NCBI
|
|
7
|
De Paepe A, Vandeneede N, Strens D and
Specenier P: The economics of the treatment and follow-up of
patients with glioblastoma. Value Health. 18:A4482015. View Article : Google Scholar
|
|
8
|
Lv B, Yang X, Lv S, Wang L, Fan K, Shi R,
Wang F, Song H, Ma X, Tan X, et al: CXCR4 signaling induced
epithelial-mesenchymal transition by PI3K/AKT and ERK pathways in
glioblastoma. Mol Neurobiol. 52:1263–1268. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lin SJ, Defossez PA and Guarente L:
Requirement of NAD and SIR2 for life-span extension by calorie
restriction in Saccharomyces cerevisiae. Science. 289:2126–2128.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Ford J, Jiang M and Milner J:
Cancer-specific functions of SIRT1 enable human epithelial cancer
cell growth and survival. Cancer Res. 65:10457–10463. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Smith JS, Brachmann CB, Celic I, Kenna MA,
Muhammad S, Starai VJ, Avalos JL, Escalante-Semerena JC, Grubmeyer
C, Wolberger C and Boeke JD: A phylogenetically conserved
NAD(+)-dependent protein deacetylase activity in the Sir2 protein
family. Proc Natl Acad Sci USA. 97:6658–6663. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Imai S, Armstrong CM, Kaeberlein M and
Guarente L: Transcriptional silencing and longevity protein Sir2 is
an NAD-dependent histone deacetylase. Nature. 403:795–800. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saunders LR and Verdin E: Sirtuins:
Critical regulators at the crossroads between cancer and aging.
Oncogene. 26:5489–5504. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Baylin SB and Ohm JE: Epigenetic gene
silencing in cancer-a mechanism for early oncogenic pathway
addiction? Nat Rev Cancer. 6:107–116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen WY, Wang DH, Yen RC, Luo J, Gu W and
Baylin SB: Tumor suppressor HIC1 directly regulates SIRT1 to
modulate p53-dependent DNA-damage responses. Cell. 123:437–448.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Choi HN, Bae JS, Jamiyandorj U, Noh SJ,
Park HS, Jang KY, Chung MJ, Kang MJ, Lee DG and Moon WS: Expression
and role of SIRT1 in hepatocellular carcinoma. Oncol Rep.
26:503–510. 2011.PubMed/NCBI
|
|
17
|
Huffman DM, Grizzle WE, Bamman MM, Kim JS,
Eltoum IA, Elgavish A and Nagy TR: SIRT1 is significantly elevated
in mouse and human prostate cancer. Cancer Res. 67:6612–6618. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jang KY, Kim KS, Hwang SH, Kwon KS, Kim
KR, Park HS, Park BH, Chung MJ, Kang MJ, Lee DG and Moon WS:
Expression and prognostic significance of SIRT1 in ovarian
epithelial tumours. Pathology. 41:366–371. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Jang KY, Hwang SH, Kwon KS, Kim KR, Choi
HN, Lee NR, Kwak JY, Park BH, Park HS, Chung MJ, et al: SIRT1
expression is associated with poor prognosis of diffuse large
B-cell lymphoma. Am J Surg Pathol. 32:1523–1531. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Qu Y, Zhang J, Wu S, Li B, Liu S and Cheng
J: SIRT1 promotes proliferation and inhibits apoptosis of human
malignant glioma cell lines. Neurosci Lett. 525:168–172. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Allen M, Bjerke M, Edlund H, Nelander S
and Westermark B: Origin of the U87MG glioma cell line: Good news
and bad news. Sci Transl Med. 8:354re32016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lopez-Valero I, Saiz-Ladera C, Torres S,
Hernández-Tiedra S, García-Taboada E, Rodríguez-Fornés F, Barba M,
Dávila D, Salvador-Tormo N, Guzmán M, et al: Targeting glioma
initiating cells with a combined therapy of cannabinoids and
temozolomide. Biochem Pharmacol. 157:266–274. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yamashita AS, da Costa Rosa M, Borodovsky
A, Festuccia WT, Chan T and Riggins GJ: Demethylation and
epigenetic modification with 5-Azacytidine reduces IDH1 mutant
glioma growth in combination with Temozolomide. Neuro Oncol.
2018.
|
|
25
|
Costa PM, Cardoso AL, Nóbrega C, Pereira
de Almeida LF, Bruce JN, Canoll P and Pedroso de Lima MC:
MicroRNA-21 silencing enhances the cytotoxic effect of the
antiangiogenic drug sunitinib in glioblastoma. Hum Mol Genet.
22:904–918. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hung AL, Maxwell R, Theodros D, Belcaid Z,
Mathios D, Luksik AS, Kim E, Wu A, Xia Y, Garzon-Muvdi T, et al:
TIGIT and PD-1 dual checkpoint blockade enhances antitumor immunity
and survival in GBM. Oncoimmunology. 7:e14667692018. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ruan Y, Dong CL, Patel J, Duan C, Wang X,
Wu X, Cao Y, Pu L, Lu D, Shen T and Li J: SIRT1 suppresses
doxorubicin-induced cardiotoxicity by regulating the oxidative
stress and p38MAPK pathways. Cell Physiol Biochem. 35:1116–1124.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chen HC, Lu Q, Fei XF, Shen L, Jiang D and
Dai D: miR-22 inhibits the proliferation, motility, and invasion of
human glioblastoma cells by directly targeting SIRT1. Tumor Biol.
37:6761–6768. 2016. View Article : Google Scholar
|
|
29
|
Zhou YF, Zhou Z, Zhang W, Hu X, Wei H,
Peng J and Jiang S: SIRT1 inhibits adipogenesis and promotes
myogenic differentiation in C3H10T1/2 pluripotent cells by
regulating Wnt signaling. Cell Biosci. 5:612015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Shuang T, Wang M, Zhou Y and Shi C:
Over-expression of Sirt1 contributes to chemoresistance and
indicates poor prognosis in serous epithelial ovarian cancer (EOC).
Med Oncol. 32:2602015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Linz U: Commentary on Effects of
radiotherapy with concomitant and adjuvant temozolomide versus
radiotherapy alone on survival in glioblastoma in a randomised
phase III study: 5-year analysis of the EORTC-NCIC trial (Lancet
Oncol. 2009;10:459-466). Cancer. 116:1844–1846. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gaspar N, Marshall L, Perryman L, Bax DA,
Little SE, Viana-Pereira M, Sharp SY, Vassal G, Pearson AD, Reis
RM, et al: MGMT-independent temozolomide resistance in pediatric
glioblastoma cells associated with a PI3-kinase-mediated HOX/stem
cell gene signature. Cancer Res. 70:9243–9252. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pyko IV, Nakada M, Sabit H, Teng L,
Furuyama N, Hayashi Y, Kawakami K, Minamoto T, Fedulau AS and
Hamada J: Glycogen synthase kinase 3β inhibition sensitizes human
glioblastoma cells to temozolomide by affecting O6-methylguanine
DNA methyltransferase promoter methylation via c-Myc signaling.
Carcinogenesis. 34:2206–2217. 2013. View Article : Google Scholar : PubMed/NCBI
|