Introduction
Glioblastoma (GBM) is one of most fatal types of
primary brain tumors with high tumorigenesis, rapid growth and high
invasiveness. GBM is defined as a grade III or IV malignant tumor
by the World Health Organization (WHO) that most frequently
involves the frontal, parietal and temporal lobes (1). Complete resection of tumors is made
difficult as a result of its location and its resistance to
chemical and radical therapies (2).
Therefore, the two-year survival rate of patients with high-grade
malignancy of glioblastoma is <9% (3). Due to the limits of standard therapy, a
novel and effective therapy is needed to improve prognosis.
Targeted molecular therapies have made marked progress in
treatment, with less severe adverse effects and a significant
increase in the cure rate (4).
Therefore, seeking out potential molecular targets for glioblastoma
is becoming an increasing area of interest.
Polycomb group (PcG) proteins, originally discovered
in the Drosophila melanogaster, are abundantly present in
cells, suppress gene transcription through epigenetic remodeling,
and serve as determinants of cell fate, thus serving important
roles in embryonic development, embryonic stem cells (ESC) and
adult stem cells maintenance and tumor formation (5–9). The PcG
family functions as complexes, which are divided into polycomb
repressive complex (PRC) 1 and PRC2 (10). Furthermore, PRC2 regulates histone
H3K27 methyltransferase activity, resulting in the recruitment of
PRC1, transcriptional repression and gene silencing (11,12).
Nervous system polycomb 1 (NSPc1), a novel member of PRC1, was
originally identified in 2001 (13).
The expression of mouse NSPc1 is relatively high and localized to
the nervous system during early embryonic development.
Additionally, NSPc1 serves an important role in stemness
maintenance of octamer (oct)-sex determining region of the Y
chromosome-box 2 (Sox2)-homeobox protein nanog axis by directly
activating the promoter of Oct4 in P19 embryonic carcinoma stem
cells (14) and overexpression of
Nspc1 induces ESC to self-renew independent of leukemia inhibitory
factor in the culture media in vitro (15). Furthermore, Yan et al
(16) recently demonstrated that
although the Nspc1 mutant cells were viable and retained normal
self-renewal capabilities, they presented with severe defects in
differentiation. Additionally, NSPc1 regulates retinoic acid
response element to downregulate p21Waf1/COP1 interacting protin-1,
which is associated with the differentiation and metastasis of
tumor cells (17). As NSPc1
possesses a marked amount of homology with the proto-oncogene and
polycomb protein BMI-1 (13), it may
additionally serve a role in tumorigenesis as a proto-oncogene, as
has been demonstrated in malignant glioma (18).
Long non-coding RNA (lncRNAs) is a variant of
non-coding RNA of 200 bp in length with a lack of an open reading
frame (19). LncRNA was firstly
identified by Okazaki et al (20) in mouse full-length cDNA library.
Recent studies have demonstrated that lncRNAs are involved in
X-chromosome silencing, gene imprinting, chromosome modification,
transcription activation and the occurrence and progression of a
number of diseases (21). Aside from
forming triplets by specifically combining with DNA and RNA, lncRNA
binds to protein to regulate gene expression and possibly cancer
progression. It has been indicated that lncRNA recruits chromosome
to certain loci for gene silence in epigenetic regulation (19).
At present, a number of lncRNAs have been identified
as directing epigenetic silencing through interactions with PcG
proteins during the initiation and progression of cancer (22). It has been demonstrated that the 5′
end of lncRNA, homeobox protein transcript antisense RNA (HOTAIR)
combined with PRC2 drives target gene epigenetically silencing
(23). Furthermore, lncRNA antisense
non-coding RNA in the INK4 locus (ANRIL) in conjunction with PRC1
and PRC2 may regulate H3K27 methylation of the INK4b-ARF-INK4a
locus (24).
The crosstalk between PRC members and lncRNAs may
serve a crucial role in epigenetically silencing gene expression,
resulting in the formation and development of tumors. In the
present study, eight lncRNAs were selected via bioinformatics
(25) as potential NSPc1 interacting
and crosstalking molecules in various types of nervous system tumor
cell lines. The present study aimed to determine among these eight
candidates the potential lncRNAs binding to PRC1 member NSPc1, and
to examine the possible crosstalks between NSPc1 and lncRNAs in
glioma cells. The results demonstrated that metastasis-associated
lung adenocarcinoma transcript 1 (MALAT1), Sox2 overlapping
transcript (SOX2OT) and ANRIL may interact with NSPc1 in U87 glioma
cells, and ANRIL may combine with NSPc1 to affect the proliferation
and apoptosis of U87 glioma cells.
Materials and methods
Cell culture and induction of
differentiation
U87 cells (derived from malignant glioma of unknown
origin; WHO grade IV) and H4 cells (derived from low-grade
malignant glioma; WHO grade II) were acquired from American Type
Culture Collection (Manassas, VA, USA). U251 cells (derived from
glioblastoma astrocytoma, additionally known as U-373; WHO grade
III) were obtained from the Type Culture Collection of the Chinese
Academy of Medical Science (Shanghai, China). All the above cell
lines used have been authenticated using STR profiling. The cell
lines were cultured in Dulbecco's modified Eagle's medium (Gibco;
Thermo Fisher Scientific Inc., Waltham, MA, USA) supplemented with
10% fetal bovine serum (Gibco; Thermo Fisher Scientific Inc.) at
37°C in an incubator containing 5% CO2. Differentiation
of U87 cells was induced by adding 1 µM of all-trans retinoic acid
(ATRA) (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) to the
medium.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from cells using
TRIzol® Reagent (Invitrogen; Thermo Fisher Scientific
Inc.). RT-qPCR was performed with the primer pair sequences for
lncRNAs and NSPc1 designed as follows: MALAT1, forward
5′-AAAGCAAGGTCTCCCCACAAG-3′, reverse 5′-GGTCTGTGCTAGATCAAAAGGCA-3′;
SOX2OT, forward 5′-AGGCATAGCATCCACCCTAA-3′, reverse
5′-AGCCCTCACACCTCCTTACTT-3′; HOTAIR, forward
5′-GGTAGAAAAAGCAACCACGAAGC-3′, reverse
5′-ACATAAACCTCTGTCTGTGAGTGCC-3′; H19, forward
5′-TTCAAAGCCTCCACGACTCT-3′, reverse 5′-GCTCACACTCACGCACACTC-3′;
ANRIL forward 5′-GAAGAAGCAAAAGCGGAAAC-3′, reverse
5′-GGACCCAGAGGGAGGTAAAT-3′; alu-mediated CDKN1A/P21 transcriptional
regulator (APTR), forward 5′-GTGGGTATCAGGGAAGAACC-3′, reverse
5-′CAGGAAAAATGGAAGGGAAA-3′; maternally expressed 3 (MEG3), forward
5′-GGGAGTGGAAAGAAACA-3′, reverse 5′-TGTGGATGGCTTGTGCTAAA-3; ADAMTS9
antisense RNA 2 (ADAMTS9-AS2), forward 5′-AACACAAACACCTTGGGTA-3,
reverse 5′-GGGTCTCTTGGCTTCGTAT-3′; NSPc1, forward
5′-GAGACACAGCCACTGCTCAA-3′, reverse 5′-TGCACATGCTGAGGGTTTAG-3′; and
GAPDH, forward, 5′-GAAGGTCGGAGTCAACGGATT-3′ and reverse
5′-CGCTCCTGGAAGATGGTGAT-3′. RT-qPCR was achieved on Step one Plus™
System (Applied Biosystems; Thermo Fisher Scientific Inc.) using
SYBR Premix EX Taq™ (Takara Biotechnology Co., Ltd., Dalian,
China), according to the manufacturer's protocol. The amplification
conditions were 95°C for 30 sec, followed by 40 cycles of 95°C for
5 sec and 60°C for 1 min. Relative gene expression was normalized
to the housekeeping gene GAPDH. The relative expression fold change
was calculated using the 2−ΔΔCq method (26). The RT-qPCR products of NSPc1 and
GAPDH were also removed for electrophoresis in 2% agarose gel, and
were further visualized using a UV Transilluminator Gel Docu 2000
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). All
quantifications are the averages of at least three biological
replicates.
Western blot analysis
Western blot analysis was performed as described
previously (14). Cells were lysed
using radioimmunoprecipitation assay buffer (cat. no. P0013B;
Beyotime Institute of Biotechnology, Nanjing, China) and the
protein concentration was determined using a bicinchonininc acid
assay kit (cat. no. P0009; Beyotime Institute of Biotechnology).
Proteins (20 µg) were separated by 10% SDS-PAGE, and then
transferred onto polyvinylidene difluoride membranes (ΕΜD
Millipore, Billerica, MA, USA). Membranes were blocked with 5%
skimmed milk dissolved inTris-buffered saline supplemented with 1%
Tween-20 at room temperature for 1 h, and incubated with primary
antibodies against NSPc1 [1:5,000, prepared in our laboratory
(27)] and GAPDH (1:6,000; cat. no.
AP0063; Bioworld, Inc., St. Louis Park, MN, USA) at 4°C overnight.
Membranes were then incubated with the horseradish
peroxidase-conjugated goat anti-rabbit immunoglobulin (Ig) G
secondary antibody (1:4,000; cat. no. sc-2004; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA) for 30 min at room
temperature. Bands were visualized with enhanced chemiluminescence
reagent (Merck KGaA). Data were analysed via densitometry using
ImageJ (version 1.47v; National Institute of Health, Bethesda, MD,
USA) and normalized to expression of GAPDH. All the densitometry
analyses are based on four-five repeats and the blots presented are
representative of the technical repeats.
RNA binding protein
immunoprecipitation (RIP) assay
RIP assays were performed using U87 cells using the
Magna RIP kit (Merck KGaA), according to the manufacturer's
protocol. lncRNAs-NSPc1 immunoprecipitation was performed using
anti-NSPc1 monoclonal antibody (m-NSPc1) (2 µg) developed in our
laboratory (14), anti-enhancer of
zeste 2 polycomb repressive complex 2 subunit (EZH2) antibody (1
µg; cat. no. 07689; EMD Millipore) as a positive control and IgG
beads (Merck KGaA) as a negative control. In addition, purified
RNAs isolated by RIP were detected by RT-qPCR.
Cell transfection
U87 cells were cultured to 60–70% confluence and
transfected with either pCDEF-NSPc1 (in order to overexpress NSPc1;
pCDEF-empty as control vector), pSE-NSPc1 (in order to knockdown
the NSPc1 gene; pSE-empty as control vector) (27) or small interfering (si)RNAs against
MALAT1, SOX2OT and ANRIL. A universal siRNA with no homology with
mammalian gene sequence was used as negative control. Sequences
were designed as follows: MALAT1, forward
5′-GGGCUUCUCUUAACAUUUAUU-3′, reverse 5′-UAAAUGUUAAGAGAAGCCCUU)-3′
(50 nM) (27,28); SOX2OT, forward
5′-GGAGAUUGUGACCUGGCUUTT-3′, reverse 5′-AAGCCAGGUCACAAUCUCCTT)-3′
(50 nM) (29); ANRIL, forward
5′-GGUCAUCUCAUUGCUCUAUTT-3′, reverse 5′-CCAGUAGAGUAACGAGAUATT)-3′
(25 nM) (24,30); and universal siRNA negative control,
forward 5′-UUCUCCGAACGUGUCACGUTT-3′ and reverse
5′-ACGUGACACGUUCGGAGAATT-3′ (50 nM). Transfections were performed
using Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.) at room temperature for 25 min and at 37°C in an
incubator containing 5% CO2 for 6 h. Plasmids were
constructed and extracted in our laboratory and siRNAs were
synthesized by Shanghai GenePharma Co., Ltd. (Shanghai, China).
Flow cytometry
Fluorescein isothiocyanate-AnnexinV (BD Biosciences,
Franklin Lakes, NJ, USA) and propidium iodide (Sigma-Aldrich; Merck
KGaA) were used to assess cell apoptosis according to the
manufacturer's protocol. U87 cells were cultured to 60–70%
confluence and transfected with siMALAT1, siSOX2OT, siANRIL and the
negative control in 6-well plate. Apoptosis of transfected U87
cells were detected using a flow cytometer (BD FACS Calibur™, BD
Biosciences) equipped with CellQuest software version 5.1 (BD
Biosciences).
MTT assay
U87 cells were cultured to 30–50% confluence and
transfected with siMALAT1, siSOX2OT, siANRIL, pSE-NSPc1 and the
negative control in a 24-well plate. MTT assay (Sigma-Aldrich;
Merck KGaA) was used to measure the proliferation of the
transfected U87 cells. Following the manufacturer's protocol, the
absorbance values were detected at optical density 490 nm, 24, 48,
72 and 96 h subsequent to transfection (independently).
Statistical methods
All results were expressed as the means ± standard
deviation. Statistical analyses were performed using SPSS software
(version 13.0; SPSS, Inc., Chicago, IL, USA). Comparisons between
two groups were analyzed by a Student's t-test. One-way analysis of
variance was applied for multiple comparisons with a post-hoc
Student-Newman-Keuls test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Expression of endogenous lncRNAs and
NSPc1 in glioma cells
mRNA and protein expression levels of NSPc1 were
associated with the degree of malignancy of glioma cell lines
(Fig. 1A and B). Furthermore, the
expression levels of MALAT1 and APTR were higher in glioma U87
cells, H4 cells and U251 cells, whereas HOTAIR, H19 and ADAMTS9-AS2
expression was lower in glioma cells. ANRIL expression levels were
relatively high in H4 cells and U251 cells compared with U87 cells.
However, the expression levels of MEG3 were relatively higher in
U251 and U87 cells compared with H4 cells (Fig. 1C).
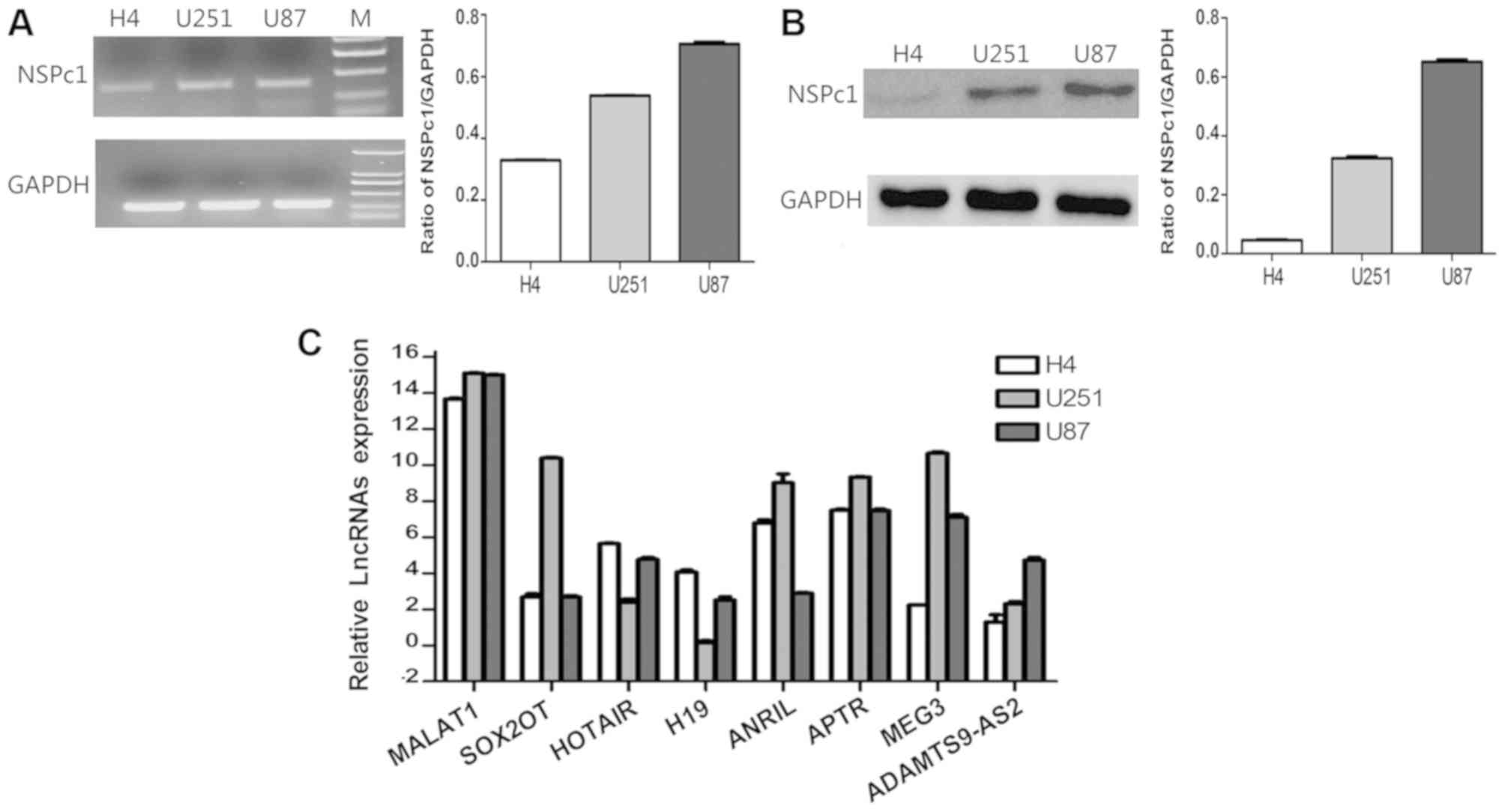 | Figure 1.Expression levels of NSPc1 and
lncRNAs in H4, U251 and U87 glioma cell lines. (A) mRNA expression
levels of NSPc1 as detected by RT-qPCR in glioma cells normalized
to GAPDH and quantified using the 2−ΔΔCq method (right
panel). RT-qPCR products were also removed for electrophoresis and
visualized using a UV Transilluminator (left panel). (B) Protein
expression levels of NSPc1 in glioma cells as detected by western
blotting with GAPDH as an internal control. (C) Expression levels
of lncRNAs in the glioma cell lines. lncRNA candidates' expression
analysis was performed using RT-qPCR. Expression of each lncRNA
candidate was quantified using the 2−ΔΔCq method with
the expression of GAPDH used as the internal control. NSPc1,
nervous system polycomb-1; lncRNAs, long non-coding RNAs; RT-qPCR,
reverse transcription-quantitative polymerase chain reaction;
MALAT1, metastasis associated lung adenocarcinoma transcript 1;
ANRIL, antisense non-coding RNA in the INK4 locus; SOX2OT,
sex-determining region of the Y chromosome-box 2 overlapping
transcript; HOTAIR, homeobox protein transcript antisense RNA;
APTR, alu-mediated CDKN1A/P21 transcriptional regulator; MEG3,
maternally expressed 3; ADAMTS9-AS2, ADAMTS9 antisense RNA 2. |
MALAT1 expression is inversely
associated with NSPc1 expression in U87 cells
To investigate whether the variability of NSPc1
expression influenced the expression of any of the eight lncRNAs,
the overexpression vector, pCDEF-NSPc1, and the knockdown vector,
pSE-NSPc1, were used. The expression of MALAT1 decreased following
upregulation of NSPc1, and it increased when expression of NSPc1
was knocked down (Fig. 2). The
expression of SOX2OT and ANRIL decreased, irrespective of NSPc1
expression levels in U87 cells. The remaining lncRNA candidates
exhibited a significant difference in expression (P<0.01) only
following NSPc1 overexpression or knockdown, whereas SOX2OT and
ANRIL expressions were both significantly decreased (P<0.01)
following NSPc1 overexpression and knockdown (Fig. 2C).
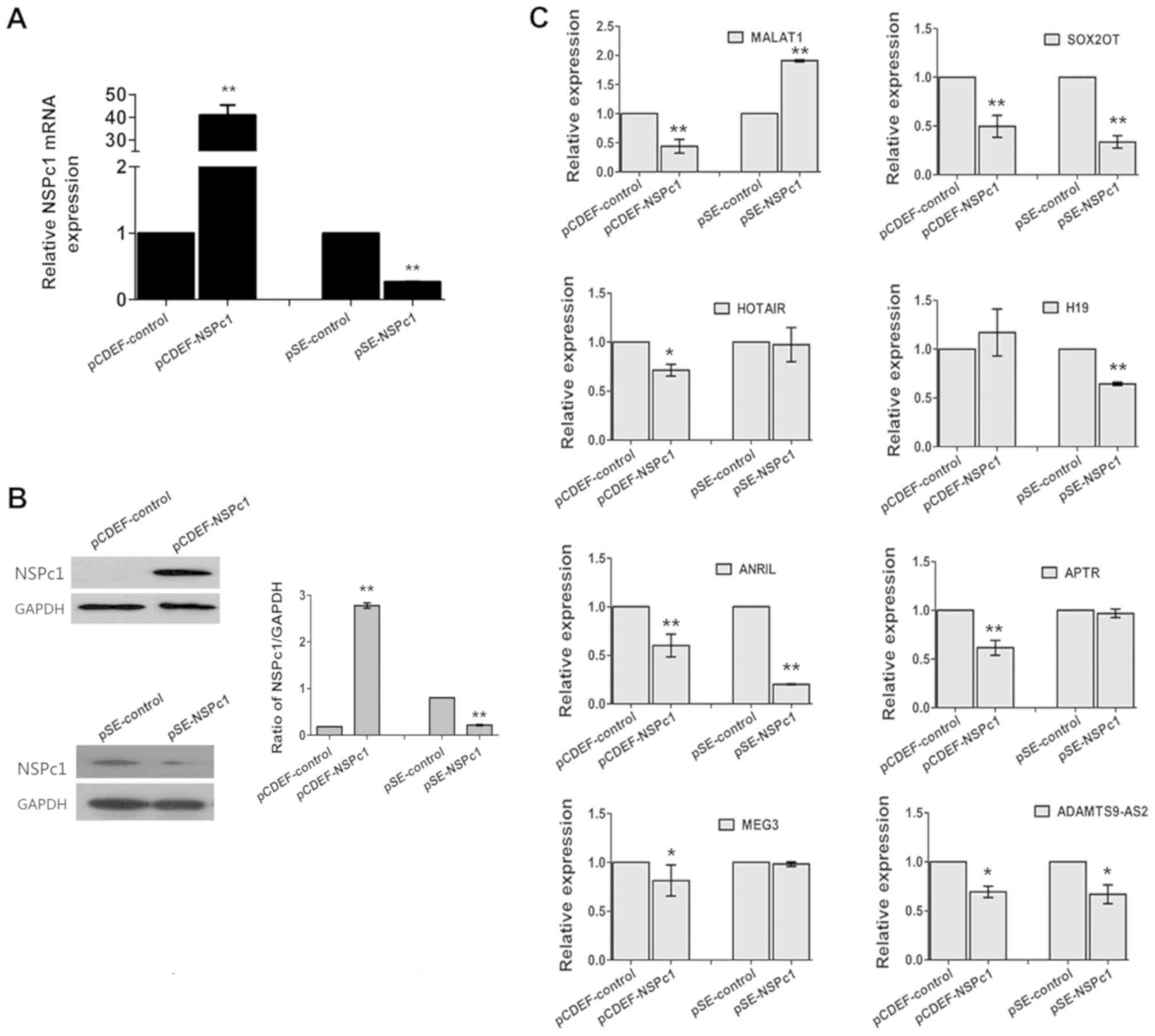 | Figure 2.The expression of NSPc1 and lncRNAs
in U87 cells subsequent to increasing or decreasing the expression
of NSPc1. U87 cells were transfected with pCDEF-NSPc1
(overexpression plasmid) or pSE-NSPc1 (knockdown plasmid). (A)
NSPc1 expression levels were determined using reverse
transcription-quantitative polymerase chain reaction. GAPDH was
used as the internal control. Results were analyzed using the
2−ΔΔCq method. (B) Protein expression levels of NSPc1
following transfection or knockdown of NSPc1 in U87 cells. GAPDH
was used as an internal control. (C) lncRNA expression levels
subsequent to regulation of NSPc1 expression with GAPDH as an
internal control. Results were analyzed using the 2−ΔΔCq
method. *P<0.05, **P<0.01 vs. control group. NSPc1, nervous
system polycomb-1; lncRNAs, long non-coding RNAs; lncRNAs, long
non-coding RNAs; MALAT1, metastasis associated lung adenocarcinoma
transcript 1; ANRIL, antisense non-coding RNA in the INK4 locus;
SOX2OT, sex-determining region of the Y chromosome-box 2
overlapping transcript; HOTAIR, homeobox protein transcript
antisense RNA; APTR, alu-mediated CDKN1A/P21 transcriptional
regulator; MEG3, maternally expressed 3; ADAMTS9-AS2, ADAMTS9
antisense RNA 2. |
lncRNA candidate expression levels are
variable following ATRA mediated induction of differentiation in
U87 glioblastoma cells
Following induction of differentiation using ATRA,
alterations in expression of NSPc1 and lncRNAs during the
differentiation process were examined. mRNA and protein expression
levels of NSPc1 decreased at the beginning and reached the lowest
expression levels measured by the 6th day of differentiation.
However, the expression levels increased subsequent to the 6th day
till the 10th day of differentiation. By the 10th day, the
expression levels of NSPc1 were higher compared with the 1st day
(Fig. 3A and B). MALAT1 expression
decreased initially, but increased by the 10th day, H19 increased
for the duration of the experiment, and MEG3 decreased
significantly by the 10th day (Fig.
3C). The expression changes of the remaining lncRNA candidates
largely followed a similar pattern; a slight increase in expression
was seen on day 1 after which expression decreased (Fig. 3C).
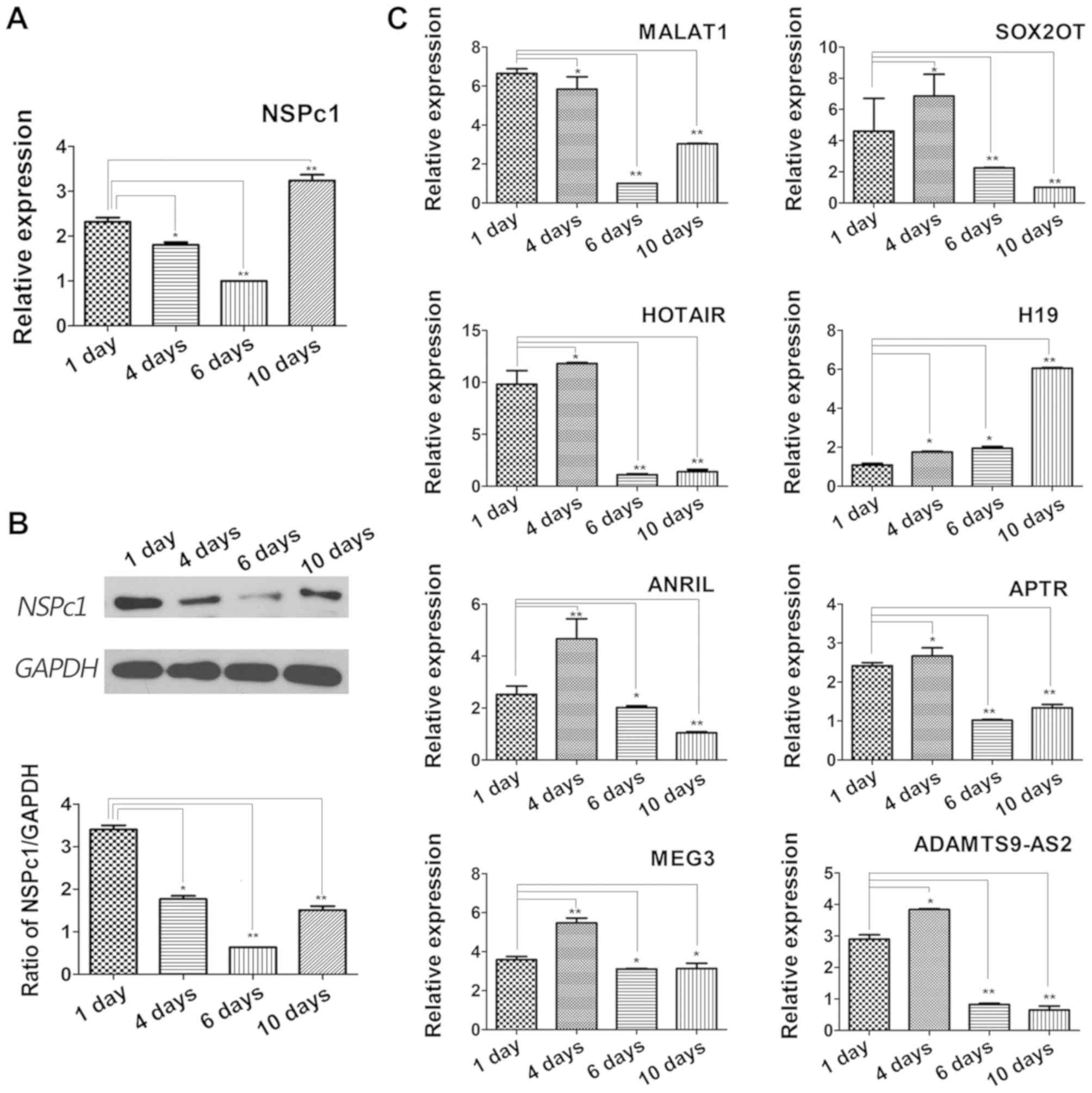 | Figure 3.Expression of NSPc1 and lncRNAs in
U87 cells following ATRA-induced differentiation. (A) Measurement
of the expression levels of NSPc1 by reverse
transcription-quantitative polymerase chain reaction following
ATRA-induced differentiation of U87 cells. GAPDH was used as an
internal control. Results were analyzed using the 2−ΔΔCq
method. (B) Protein expression levels of NSPc1 following
ATRA-induced differentiation of U87 cell by western blotting. GAPDH
was used as an internal control. (C) Expression of lncRNAs
following ATRA-induced differentiation of U87 cells. GAPDH was used
as the internal control. Results were analyzed using the
2−ΔΔCq method. *P<0.05, **P<0.01. NSPc1, nervous
system polycomb-1; lncRNAs, long non-coding RNAs; ATRA, all-trans
retinoic acid; MALAT1, metastasis associated lung adenocarcinoma
transcript 1; ANRIL, antisense non-coding RNA in the INK4 locus;
SOX2OT, sex-determining region of the Y chromosome-box 2
overlapping transcript; HOTAIR, homeobox protein transcript
antisense RNA; APTR, alu-mediated CDKN1A/P21 transcriptional
regulator; MEG3, maternally expressed 3; ADAMTS9-AS2, ADAMTS9
antisense RNA 2. |
Evaluation of lncRNA candidates
interacting with NSPc1 during ATRA-induced U87 differentiation
To investigate a potential interaction between
endogenous NSPc1 and lncRNA candidates, RIP assays were used.
According to the aforementioned data, MALAT1, SOX2OT and ANRIL were
selected for further studies (as presented in Fig. 2). From the RIP assays on U87 cells
(Fig. 4A-C), the enrichment of
immunoprecipitation of MALAT1 and SOX2OT decreased throughout the
whole differentiation process and the enrichment of SOX2OT was
almost absent by the 10th day (Fig.
4C; left and middle panels). The enrichment of
immunoprecipitation of ANRIL decreased until the 6th day of
differentiation, following which the expression levels increased
until the 10th day of differentiation (Fig. 4C; right panel).
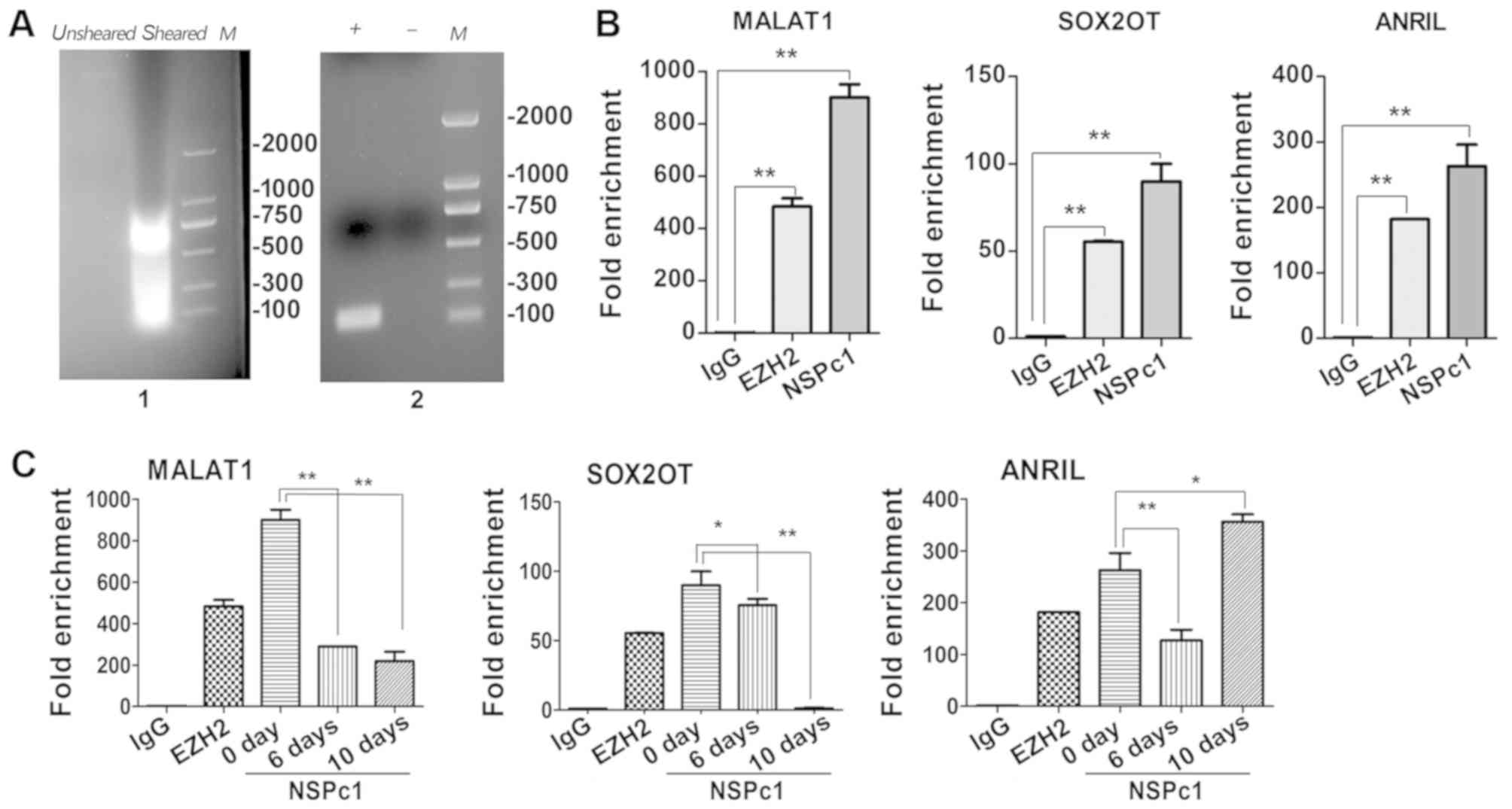 | Figure 4.Identification of lncRNAs
interactions with NSPc1 in U87 cells. (A) Identification of nucleic
acid shear fragments by 2% agarose gel electrophoresis. The nucleic
acid clip was between ~200 and 1,000 bp (left panel). Efficiency
test of immunoprecipitation enrichment of positive and negative
control analysed by 2% agarose gel electrophoresis (right panel).
(B) Purified RNA immunoprecipitated by antibodies and analysed by
RT-qPCR in U87 cells. Verification of RIP enrichment was performed
by applying the comparative 2−ΔΔCq method with
anti-NSPc1 binding lncRNA compared with positive control
(anti-EZH2) binding lncRNA and negative control (IgG) binding
lncRNA. (C) The relative enrichment folds of three lncRNA
candidates in the all-trans retinoic assay differentiated cells
were calculated using the 2−ΔΔCq methodfor each group.
All data are presented as the mean ± standard deviation of three
independent repeats. *P<0.05, **P<0.01. EZH2, enhancer of
zeste 2 polycomb repressive complex 2 subunit; NSPc1, nervous
system polycom-1; RIP, RNA immunoprecipitation; MALAT1, metastasis
associated lung adenocarcinoma transcript 1; ANRIL, antisense
non-coding RNA in the INK4 locus; SOX2OT, sex-determining region of
the Y chromosome-box 2 overlapping transcript; IgG, immunoglobulin
G; M, ladder; +, positive control; -, negative control. |
Knocking down expression of lncRNA
candidates reduces proliferation and apoptosis in U87 cells
The proliferation and apoptosis of U87 cells
following siRNA transfection are presented in Fig. 5. The knockdown efficiency of MALAT1,
SOX2OT and ANRIL in U87 glioma cells was examined with increasing
concentrations of siRNAs using RT-qPCR (Fig. 5A). The MTT assay demonstrated that
knocking down the expression of MALAT1, SOX2OT and ANRIL using
their respective siRNAs, reduced proliferation of U87 cells, and
the reduction in proliferation was more prominent when NSPc1 and
ANRIL were knocked down together (Fig.
5B). Furthermore, using flow cytometry, it was determined that
the rates of apoptosis was significantly increased following
downregulation of MALAT1, SOX2OT and ANRIL expression compared with
the control group. Similarly, the apoptotic rate was further
reduced when NSPc1 and ANRIL expression were knocked down together
(Fig. 5C).
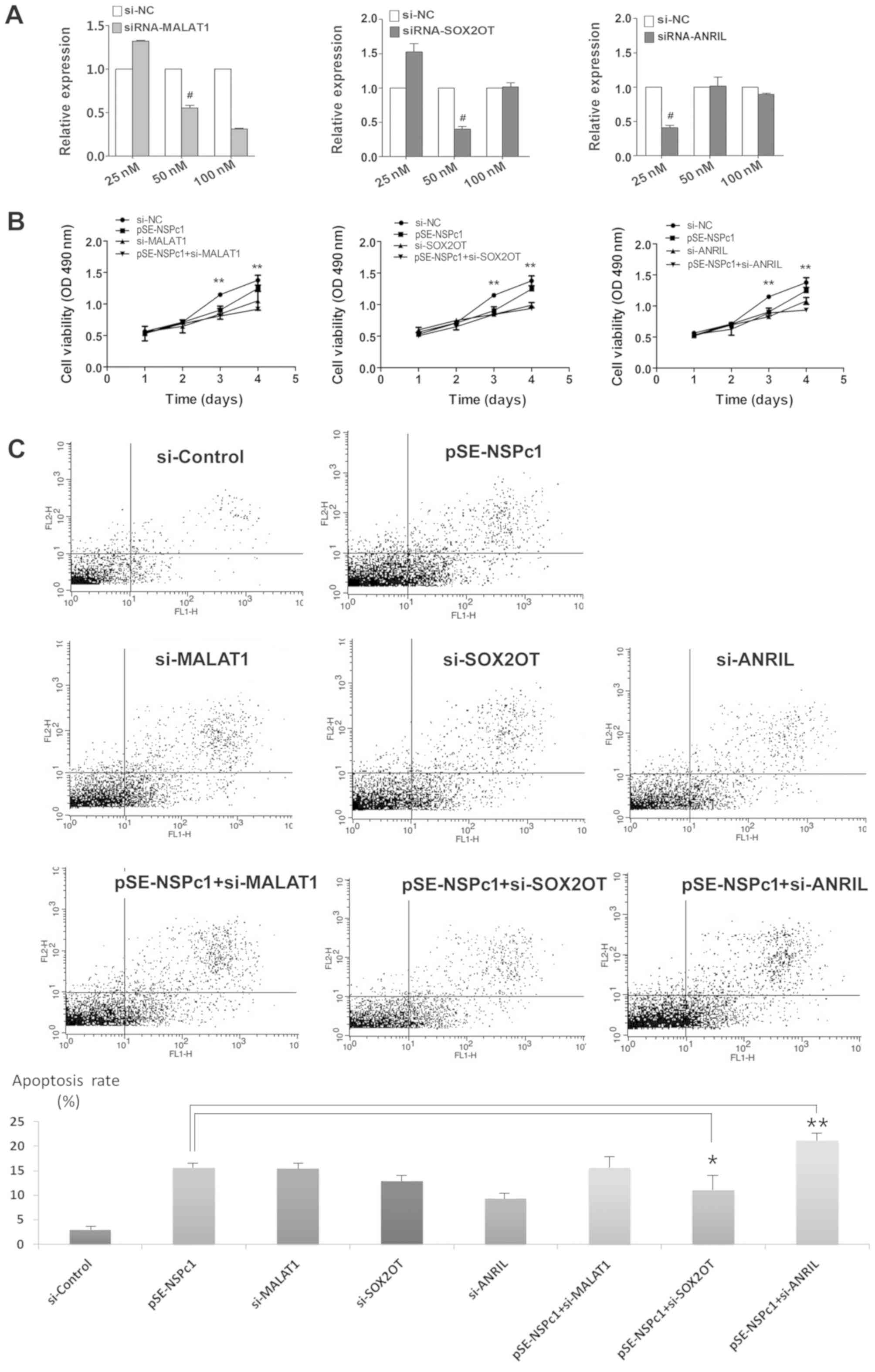 | Figure 5.Proliferative and apoptotic rates of
U87 glioma cells under different NSPc1 and lncRNAs expression
conditions. (A) siRNA knockdown efficiency of MALAT1, SOX2OT and
ANRIL in U87 glioma cells. The knockdown efficiency was determined
using a concentration gradient of each siRNA and reverse
transcription-quantitative polymerase chain reaction with GAPDH as
the loading control. (B) MTT assay was used to determine the
viability of U87 cells following transfection with si-NC,
pSE-NSPc1, si-MALAT1, si-SOX2OT, si-ANRIL, pSE-NSPc1+si-MALAT1,
pSE-NSPc1+ si-SOX2OT, pSE-NSPc1+ si-ANRIL. (C) Flow cytometry
analysis was performed to evaluate the apoptotic rate of U87 cells
following knockdown of MALAT1, SOX2OT and ANRIL, and following the
knockdown of the three candidate lncRNAs combined with knockdown of
NSPc1. *P<0.05, **P<0.01; #siRNA concentration
used for subsequent experiments. si, small-interfering; lncRNAs,
long non-coding RNAs, NSPc1, nervous system polycomb-1; pSE-NSPc1,
NSPc1 knockdown plasmid; MALAT1, metastasis associated lung
adenocarcinoma transcript 1; ANRIL, antisense non-coding RNA in the
INK4 locus; SOX2OT, sex-determining region of the Y chromosome-box
2 overlapping transcript. |
Discussion
Emerging evidence has demonstrated that glioma cells
recruited undifferentiated and differentiated cells, providing
heterogeneity for tumorigenesis (31). NSPc1, as a key member of PRC1, was
associated with the differentiation of tumor cells or cancer stem
cells in (17,18). Certain lncRNAs were identified as
novel epigenetic regulators (32).
Previous studies have hypothesized a link between PcG complexes and
lncRNAs (24). There are a number of
established glioma cell line models, including U251, H4 and U87. In
the present study, only the U87 cell line was used as: i) U251
cells are difficult to transiently transfected; ii) H4 neuroglioma
cells are derived from low-grade malignant glioma and has a
relatively low NSPc1 expression level, thus making it unsuitable
for RIP or RNAi assays; and iii) of the three cell lines, only the
U87 cells have been demonstrated to respond to differentiation
following treatment with ATRA (33),
thus making it a suitable model for the present studies. Therefore,
the association between NSPc1 and a number of lncRNAs in
differentiated U87 glioma cells was examined.
A total of eight lncRNAs were selected, which were
most commonly associated with glioma, and exhibit high correlation
scores with NSPc1 based on bioinformatics analysis (32). The expression of NSPc1 and lncRNA
candidates is associated with the degree of malignancy of different
glioma cells lines. This agrees with a recent study by Hu et
al (18), where it was
demonstrated that NSPc1 was highly expressed in stem cell-like
glioma cells (SLCs).
The expression levels of HOTAIR, APTR and
ADAMTS9-AS2 were decreased following NSPc1 overexpression, and the
levels of MALAT1, SOX2OT and ANRIL were significantly reduced
(P<0.001); therefore, these were investigated further. SOX2OT
and ANRIL were downregulated upon either overexpression or
silencing of NSPc1. NSPc1 is typically considered to be a
transcription repressor (17);
however, a previous study has demonstrated that it may additionally
be an activator of the SOX2 gene (14). The present data raise the possibility
of an, as yet, unidentified indirect regulatory mechanism between
the expression levels of NSPc1 and those of SOX2OT and ANRIL under
certain circumstances.
Previously, Hu et al (18) demonstrated that ATRA partly reversed
the NSPc1-induced stemness enhancement in SLCs through mechanisms
associated with an ATRA-dependent decrease in the expression of
NSPc1. It was demonstrated that there was a significant change in
MALAT1, SOX2OT and ANRIL with variation of NSPc1 expression levels
in the ATRA mediated differentiation process of U87 malignant
glioma cells. Hu et al (18)
additionally demonstrated that NSPc1 promotes self-renewal of stem
cell-like glioma cells by repressing the synthesis of ATRA by
targeting RDH16, suggesting that NSPc1 expression levels in U87
cells may be crucial for determining whether a cell should
differentiate or proliferate.
In the ATRA-induced differentiation of U87 cells,
results demonstrated that NSPc1 and MALAT1 expression levels were
decreased in a similar manner. Furthermore, the lncRNAs that did
not demonstrate a decrease in expression upon NSPc1 silencing were
downregulated following ATRA-induced differentiation (for example,
HOTAIR and APTR expression), suggesting that the lncRNAs respond to
ATRA-induced differentiation independently of NSPc1 expression
levels. An experiment overexpressing NSPc1 following ATRA-induced
differentiation may help to determine which effects are dependent
on NSPc1 expression levels and which effects are dependent on the
differentiation process itself. However, it is very difficult to
efficiently transfect the cells following induced differentiation
of U87 cells. At present, it is difficult to ascertain which
effects are the result of NSPc1 expression, and which effects are
the results of the differentiation process.
MALAT1 was demonstrated to interact with NSPc1 in
U87 cells using an RIP assay. It was previously demonstrated that
MALAT1 was able to maintain self-renewing ability and increase the
proportion of pancreatic cancer stem cells (CSCs) (34). In U87 cells, ATRA-induced
differentiation activates the same upstream as retinoic acid
signaling pathway of NSPc1 and MALAT1. Additionally, SOX2OT and
ANRIL were also demonstrated to interact with NSPc1 in U87
glioblastoma cells by RIP assay. The expression of SOX2OT is
associated with that of regulators of pluripotency, including SOX2
and OCT4, in the pluripotent cell line NT2 (35). ANRIL binds to PRC2 to promote
proliferation in gastric cancer, resulting in epigenetic
suppression in trans (as transcription factor) (36). Importantly, it was demonstrated that
ANRIL interacted with the PRC2 member at the INK4b/ARF/INL4a locus
(24). Since MALAT1, SOX2OT and
ANRIL interact with NSPc1 in the U87 cells during the
differentiation process, it is hypothesized that these three
lncRNAs are closely associated with NSPc1 in the canonical PRC1
complex in glioma cells.
The downregulated expression of MALAT1, SOX2OT and
ANRIL inhibited the proliferation, and promoted apoptosis of U87
cells. Furthermore, downregulation of NSPc1 combined with ANRIL but
not with MALAT1 or SOX2OT was determined to exhibit a more
significant effect on proliferation and apoptosis of U87 glioma
cells compared with downregulating NSPc1 or ANRIL alone. Therefore,
an in-depth study on the interaction mechanism between ANRIL and
NSPc1 is required to better understand the formation and
development of malignant glioma.
In conclusion, the present data demonstrated that
MALAT1, SOX2OT and ANRIL interacted with NSPc1. In addition, ANRIL
may interact with NSPc1 during ATRA-induced differentiation of U87
cells, and may be involved in regulating proliferation and
apoptosis of U87 cells. Further studies will be required to
determine the role of the ANRIL-NSPc1-PRC1 complex in
tumorigenesis.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Sciences Foundation of China (grant no. 31070929).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
YW, ZL and HL performed experiments. HL, YS and YG
analysed and interpreted the data. ZL, YW and YG were major
contributors in writing the manuscript. All authors read and
approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Stupp R, Mason WP, van den Bent MJ, Weller
M, Fisher B, Taphoorn MJ, Belanger K, Brandes AA, Marosi C, Bogdahn
U, et al: Radiotherapy plus concomitant and adjuvant temozolomide
for glioblastoma. N Engl J Med. 352:987–996. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sathornsumetee S, Rich JN and Reardon DA:
Diagnosis and treatment of high-grade astrocytoma. Neurol Clin.
251111–1139. (x)2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Filippini G, Falcone C, Boiardi A, Broggi
G, Bruzzone MG, Caldiroli D, Farina R, Farinotti M, Fariselli L,
Finocchiaro G, et al: Prognostic factors for survival in 676
consecutive patients with newly diagnosed primary glioblastoma.
Neuro Oncol. 10:79–87. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Barnes MN, Deshane JS, Rosenfeld M, Siegal
GP, Curiel DT and Alvarez RD: Gene therapy and ovarian cancer: A
review. Obstet Gynecol. 89:145–155. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wilson BG, Wang X, Shen X, McKenna ES,
Lemieux ME, Cho YJ, Koellhoffer EC, Pomeroy SL, Orkin SH and
Roberts CW: Epigenetic antagonism between polycomb and SWI/SNF
complexes during oncogenic transformation. Cancer Cell. 18:316–328.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Su Y, Deng B and Xi R: Polycomb group
genes in stem cell self-renewal: A double-edged sword. Epigenetics.
6:16–19. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Morey L and Helin K: Polycomb group
protein-mediated repression of transcription. Trends Biochem Sci.
35:323–332. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Sauvageau M and Sauvageau G: Polycomb
group proteins: Multi-faceted regulators of somatic stem cells and
cancer. Cell Stem Cell. 7:299–313. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Prezioso C and Orlando V: Polycomb
proteins in mammalian cell differentiation and plasticity. FEBS
Lett. 585:2067–2077. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Pasini D, Bracken AP and Helin K: Polycomb
group proteins in cell cycle progression and cancer. Cell Cycle.
3:396–400. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jacobs JJ and van Lohuizen M: Polycomb
repression: From cellular memory to cellular proliferation and
cancer. Biochim Biophys Acta. 1602:151–161. 2002.PubMed/NCBI
|
|
12
|
Chang CJ and Hung MC: The role of EZH2 in
tumour progression. Br J Cancer. 106:243–247. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nunes M, Blanc I, Maes J, Fellous M,
Robert B and McElreavey K: NSPc1, a novel mammalian Polycomb gene,
is expressed in neural crest-derived structures of the peripheral
nervous system. Mech Dev. 102:219–222. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li H, Fan R, Sun M, Jiang T and Gong Y:
Nspc1 regulates the key pluripotent Oct4-Nanog-Sox2 axis in P19
embryonal carcinoma cells via directly activating Oct4. Biochem
Biophys Res Commun. 440:527–532. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pritsker M, Ford NR, Jenq HT and Lemischka
IR: Genomewide gain-of-function genetic screen identifies
functionally active genes in mouse embryonic stem cells. Proc Natl
Acad Sci USA. 103:6946–6951. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Yan Y, Zhao W, Huang Y, Tong H, Xia Y,
Jiang Q and Qin J: Loss of polycomb group protein Pcgf1 severely
compromises proper differentiation of embryonic stem cells. Sci
Rep. 7:462762017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gong Y, Yue J, Wu X, Wang X, Wen J, Lu L,
Peng X, Qiang B and Yuan J: NSPc1 is a cell growth regulator that
acts as a transcriptional repressor of p21Waf1/Cip1 via the RARE
element. Nucleic Acids Res. 34:6158–6169. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hu PS, Xia QS, Wu F, Li DK, Qi YJ, Hu Y,
Wei ZZ, Li SS, Tian NY, Wei QF, et al: NSPc1 promotes cancer stem
cell self-renewal by repressing the synthesis of all-trans retinoic
acid via targeting RDH16 in malignant glioma. Oncogene.
36:4706–4718. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang Y, Liu N, Wang JP, Wang YQ, Yu XL,
Wang ZB, Cheng XC and Zou Q: Regulatory long non-coding RNA and its
functions. J Physiol Biochem. 68:611–618. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Okazaki Y, Furuno M, Kasukawa T, Adachi J,
Bono H, Kondo S, Nikaido I, Osato N, Saito R, Suzuki H, et al:
Analysis of the mouse transcriptome based on functional annotation
of 60,770 full-length cDNAs. Nature. 420:563–573. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wilusz JE: Long noncoding RNAs: Re-writing
dogmas of RNA processing and stability. Biochim Biophys Acta.
1859:128–138. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tsai MC, Spitale RC and Chang HY: Long
intergenic noncoding RNAs: New links in cancer progression. Cancer
Res. 71:3–7. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsai MC, Manor O, Wan Y, Mosammaparast N,
Wang JK, Lan F, Shi Y, Segal E and Chang HY: Long noncoding RNA as
modular scaffold of histone modification complexes. Science.
329:689–693. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kotake Y, Nakagawa T, Kitagawa K, Suzuki
S, Liu N, Kitagawa M and Xiong Y: Long non-coding RNA ANRIL is
required for the PRC2 recruitment to and silencing of p15(INK4B)
tumor suppressor gene. Oncogene. 30:1956–1962. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu Q, Ren S, Lu M, Zhang Y, Zhu D, Zhang X
and Li T: Computational prediction of associations between long
non-coding RNAs and proteins. BMC Genomics. 14:6512013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu X, Gong Y, Yue J, Qiang B, Yuan J and
Peng X: Cooperation between EZH2, NSPc1-mediated histone H2A
ubiquitination and Dnmt1 in HOX gene silencing. Nucleic Acids Res.
36:3590–3599. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xiang J, Guo S, Jiang S, Xu Y, Li J, Li L
and Xiang J: Silencing of long non-coding RNA MALAT1 promotes
apoptosis of glioma cells. J Korean Med Sci. 31:688–694. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Shahryari A, Rafiee MR, Fouani Y, Oliae
NA, Samaei NM, Shafiee M, Semnani S, Vasei M and Mowla SJ: Two
novel splice variants of SOX2OT, SOX2OT-S1, and SOX2OT-S2 are
coupregulated with SOX2 and OCT4 in esophageal squamous cell
carcinoma. Stem Cells. 32:126–134. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhang D, Ding L, Li Y, Ren J, Shi G, Wang
Y, Zhao S, Ni Y and Hou Y: Midkine derived from cancer-associated
fibroblasts promotes cisplatin-resistance via up-regulation of the
expression of lncRNA ANRIL in tumour cells. Sci Rep. 7:162312017.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Rolle K: miRNA multiplayers in glioma.
From bench to bedside. Acta Biochim Pol. 62:353–365. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Caley DP, Pink RC, Trujillano D and Carter
DR: Long noncoding RNAs, chromatin, and development.
ScientificWorldJournal. 10:90–102. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Ling GQ, Liu YJ, Ke YQ, Chen L, Jiang XD,
Jiang CL and Ye W: All-trans retinoic acid impairs the vasculogenic
mimicry formation ability of U87 stem-like cells through promoting
differentiation. Mol Med Rep. 12:165–172. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jiao F, Hu H, Han T, Yuan C and Wang L,
Jin Z, Guo Z and Wang L: Long noncoding RNA MALAT-1 enhances stem
cell-like phenotypes in pancreatic cancer cells. Int J Mol Sci.
16:6677–6693. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Shahryari A, Rafiee MR, Fouani Y, Oliae
NA, Samaei NM, Shafiee M, Semnani S, Vasei M and Mowla SJ: Two
novel splice variants of SOX2OT, SOX2OT-S1, and SOX2OT-S2 are
coupregulated with SOX2 and OCT4 in esophageal squamous cell
carcinoma. Stem Cells. 32:126–134. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang EB, Kong R, Yin DD, You LH, Sun M,
Han L, Xu TP, Xia R, Yang JS, De W and Chen JF: Long noncoding RNA
ANRIL indicates a poor prognosis of gastric cancer and promotes
tumor growth by epigenetically silencing of miR-99a/miR-449a.
Oncotarget. 5:2276–2292. 2014. View Article : Google Scholar : PubMed/NCBI
|














