Introduction
Liver cancer is the fifth most common type of cancer
and the third leading cause of tumor-associated mortality
worldwide, being particularly prevalent in Asia (1). The most effective treatment for liver
cancer is currently surgery (2).
However, the characteristics of liver cancer, including multifocal
development or distant metastasis, preclude surgical treatment for
the patients from being curative (3). Therefore, in order to determine the
biology of liver cancer, it is important to gain a thorough
understanding of the underlying molecular mechanisms of tumor
growth and metastasis.
Mature microRNAs (miRNAs) are a group of short
non-coding RNAs containing between 18 and 26 nucleotides, which, at
the post-transcriptional level, regulate target gene expression by
targeting the 5′ or 3′-untranslated regions (UTRs) of mRNA
(4). miRNAs regulate a number of
biological processes, and their dysregulation has been demonstrated
to be associated with development, proliferation, stress
resistance, metastasis and apoptosis of cancer cells by regulating
oncogenes or tumor suppressors (5),
including in liver cancer. For example, microRNA (miR)-224 promotes
liver cancer proliferation and metastasis (6); conversely, miR-335 (7) and miR-424 (8) suppress liver cancer proliferation and
metastasis. Furthermore, miR-330-3p has been identified to be
deregulated in malignant liver cancer (9). However, the association of miR-330-3p
with liver cancer and its function in liver cancer remains
unclear.
In the present study, the function of miR-330-3p in
liver cancer was investigated, with the aim of providing new
insights into the pathology of liver cancer, which may have value
in the development of diagnostics and therapeutics for liver
cancer.
Materials and methods
Patient samples
In total, 30 cases of liver cancer samples were
collected between May 2012 and April 2015 at The First Hospital of
Jilin University (Changchun, China). Prior to surgery, none of the
30 patients received chemotherapy or radiation therapy. Prior to
use, the tissue samples were stored in liquid nitrogen. The present
study was approved by the ethical committee of the First Affiliated
Hospital of Jilin University. Table
I presents information concerning patient age, sex, and cancer
stage were obtained from patient records.
 | Table I.Clinicopathological characteristics of
patients with liver cancer and healthy volunteers. |
Table I.
Clinicopathological characteristics of
patients with liver cancer and healthy volunteers.
|
| Patients with liver
cancer (n=30) |
|
|---|
|
|
|
|
|---|
| Characteristic | BCLC 0 (n=3) | BCLC A (n=12) | BCLC B (n=5) | BCLC C+D (n=10) | Normal (n=20) |
|---|
| Clinical factors |
| Age,
years | 61.2±9.7 | 56.2±8.8 | 56.9±8.4 | 52.5±11.2 | 54.4±12.4 |
| Sex
(male/female) | 2/1 | 10/2 | 1/4 | 8/2 | 14/6 |
| AST,
IU/l | 48.6±35.7 | 56.1±78.2 | 54.5±44.6 | 67.8±49.3 | 11.6±5.4 |
| ALT,
IU/l | 51.8±47.2 | 54.8±83.3 | 49.4±33.1 | 72.7±58.4 | 15.4±8.7 |
|
Child-pugh classification | 3/0 | 11/1 | 4/1 | 6/4 | 20/0 |
| (A/B) (23) |
| Tumor-associated
factors |
| AFP,
ng/ml | 148.0±378.3 | 256.5±426.6 | 254.4±405.2 | 789.5±628.7 | 6.39±5.85 |
| Tumor number |
|
(single/multiple) | 3/0 | 11/1 | 1/4 | 4/6 | – |
| Tumor
size, cm3 | 2.1±0.5 | 3.4±1.2 | 6.1±2.3 | 7.5±2.7 | – |
| Vascular
invasion (−/+) | 3/0 | 12/0 | 5/0 | 3/7 | – |
| Edmondson
grade (I/II/III) (24) | 0/1/2 | 1/9/2 | 1/2/2 | 1/7/2 | – |
Cell culture and treatment
The human liver cancer cell line HepG2 were obtained
from the American Type Culture Collection (Manassas, VA, USA).
HepG2 cells were maintained in Dulbecco's modified Eagle's medium
(DMEM; Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) supplemented with 10% fetal bovine serum (FBS; HyClone; GE
Healthcare, Logan, UT, USA) at 37°C in a humidified atmosphere
containing 5% CO2. miR-330-3p mimic
(5′-CTGCAGAGAGGCAGCGCTGT-3′) and negative control (NC) (termed as
NC, 5′-ACUACUGAGUGACAGUAGA-3′) oligonucleotides were purchased from
Guangzhou RiboBio Co., Ltd. (Guangzhou, China). Transfection of
cells with 50 nM oligonucleotides was carried out using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.), according to the manufacturer's protocol.
Target gene prediction and
verification
The target genes of miR-330-3p were predicted using
three bioinformatics algorithms: PicTar (https://pictar.mdc-berlin.de), miRbase (http://www.mirbase.org) and TargetScan (http://www.targetscan.org). Mitogen-activated protein
kinase kinase 1 (MAP2K1) was identified to be downregulated by
miR-330-3p, therefore, from human genomic DNA, the full-length
3′UTR of MAP2K1 was amplified and cloned into pMIR-GLOTM luciferase
vector (Promega Corporation, Madison, WI, USA) to serve as the
wild-type. The mutations were achieved using site-directed
mutagenesis QuickChange™ Multi Site-Directed Mutagenesis kit
(Clontech Laboratories, Inc., Mountainview, CA, USA). A total of
1×104 HepG2 cells was plated in 48-well plates, and
co-transfection was performed using 200 ng wild-type, mutant or
pMIR-GLOTM empty vector, and either 80 ng miR-330-3p expression
vector or 80 ng pcDNA3.1(+) empty vector. At 48 h after
transfection, cells were harvested and were assayed for firefly and
Renilla luciferase activity using a Dual-Luciferase Glow
assay kit (Promega Corporation) according to the manufacturer's
protocol. Overexpression of MAP2K1 was achieved using a pCMV-MAP2K1
expression plasmid. All transfection experiments were conducted in
triplicate.
Wound healing assay
For the wound-healing assay, HepG2 cells were
collected and trypsinized, followed by seeding equally in 6-well
cell culture plates. After 24 h, the cells reached confluence.
Using a sterile 100 µl pipette tip, artificial homogeneous wounds
in the monolayer were created. The cell culture plate was washed
with serum-free DMEM. The wound distance was determined at 0 and 24
h under a reflected-light microscope (magnification, ×40).
Cell migration assay
To determine cell invasion capability, a Transwell
assay was used. Prior to the experiment, HepG2 cells were
transfected with miR-330-3p mimic or NC oligonucleotides. After 16
h, transfected cells were typsinized and resuspended in DMEM with
10% FBS (Life Technologies; Thermo Fisher Scientific, Inc.), and
1.0×104 cells were placed into the upper chambers (8-µm
pore size; EMD Millipore, Billerica, MA, USA) with 200 µl RPMI-1640
medium. The lower chambers were filled with 600 µl DMEM
supplemented with 10% FBS. Following incubation at 37°C for 48 h,
non-invading cells were removed from the top of the chamber. The
migratory cells in the lower chamber were fixed with 90% methyl
alcohol at room temperature for 30 min and stained with 0.1%
crystal violet. Stained cells in five randomly selected fields were
counted under a microscope (magnification, ×200).
To verify the function of MAP2K1 in the suppression
of liver cancer cell migration by miR-330-3p, HepG2 cells were
transfected with miR-330-3p together with the MAP2K1 expression
vector pCMV-MAP2K1. For lentivirus-mediated expression of SNF2H,
FLAG-tagged full-length SNF2H ORF was cloned in pWPI.1 (plasmid no.
12254; Addgene, Inc., Cambridge, MA, USA). For RNA interference of
SNF2H expression, DNA fragments encoding the hairpin precursors for
shSNF2H#1 (5′-CGTCGAATTAAGGCTGATGTT-3′) and shSNF2H#2
(5′-CGACTGCTGATGTAGTAATTT-3′) were inserted into the pLKO.1-TRC
cloning vector. A scrambled small interfering RNA precursor (Scr)
of similar GC-content to shSNF2H#1 and shSNF2H#2, but without
sequence identity to SNF2H cDNA, was used as the control. The
MAP2K1 expression vector pCMV-MAP2K1 and miR-330-3p mimic were
co-transfected into HepG2 cell using Lipofectamine 2000 (Thermo
Fisher Scientific, Inc.). After 48 h, HepG2 cells were collected
for the following experiments.
Western blot analysis
HepG2 cells were transfected with miR-330-3p mimic
or NC oligonucleotides and incubated at 37°C for 24 h. Total
protein was isolated from cell pellets with
radioimmunoprecipitation lysis buffer supplemented with Complete
protease and phosphatase inhibitor cocktail (Roche Diagnostics,
Basel, Switzerland). Proteins (20 µg) were separated by 12%
SDS-PAGE. Electrophoretically separated proteins were transferred
onto polyvinylidene difluoride membranes (Thermo Fisher Scientific,
Inc.). Membranes were blocked with 5% non-fat milk at room
temperature for 2 h. Blots were probed for MAP2K1 (1:10,000, cat.
no. ab32134; Abcam) and β-actin loading control (1:5,000 cat. no.
ab8227; Abcam). Antibodies bound to target proteins were visualized
using the Enhanced Chemiluminescence Western Blotting Detection
Reagent (Santa Cruz Biotechnology, Dallas, TX, USA). Antibodies
bound to target proteins were visualized using the Enhanced
Chemiluminescence Western Blotting Detection Reagent (Santa Cruz
Biotechnology, Dallas, TX, USA). The images were quantified using
the LAS3000 imaging system (Fujifilm Corporation, Tokyo,
Japan).
RNA isolation and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
HepG2 cells transfected with miR-330-3p mimic or NC
oligonucleotides were incubated for 24 h and collected using the
mirVana Kit (Ambion Inc., Austin, TX, USA), total RNA was isolated.
cDNA was generated using a PrimeScript™ RT-qPCR kit from
total RNA and amplified according to the manufacturer's protocol
(Takara Biotechnology Co., Ltd., Dalian, China). Amplification was
performed with the following thermocycling conditions: 5 min at
95°C, followed by 40 cycles of 95°C for 30 sec and 65°C for 45 sec.
Primer sequences are presented in Table
II. The expression of the target gene or miRNA was normalized
to GAPDH or U6, respectively. Analysis of relative gene expression
data was performed using the real-time quantitative PCR and the
2−ΔΔCq method (10).
| Table II.Primer sequence for reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primer sequence for reverse
transcription-quantitative polymerase chain reaction.
| Gene | Sense primer
(5′-3′) | Antisense primer
(5′-3′) |
|---|
| U6 |
TGCGGGTGCTCGCTTCGGCA |
CCAGTGCAGGGTCCGAGGT |
| β-actin |
CTCCATCCTGGCCTCGCTGT |
GCTGTCACCTTCACCGTTCC |
| miR-330-3p |
CAACTGCCTCTCTGGGCCTG |
CTGCAGAGAGGCAGCGCTG |
| MAP2K1 |
CAAGAAGAAGCCGACGCCCAT |
GACGCCAGCAGCATGGGTTG |
Statistical analysis
Results are expressed as the mean ± standard
deviation. Each experiment was repeated at least three times. SPSS
(version 13.0; SPSS, Inc., Chicago, IL, USA) was used for
statistical analysis. The statistical significance of the
difference between groups was determined by analysis of variance
(two-way ANOVA) with Duncan's post-hoc test) or an unpaired
Student's t-test. P≤0.05 was considered to indicate a statistically
significant difference.
Results
miR-330-3p is downregulated in
patients with liver cancer
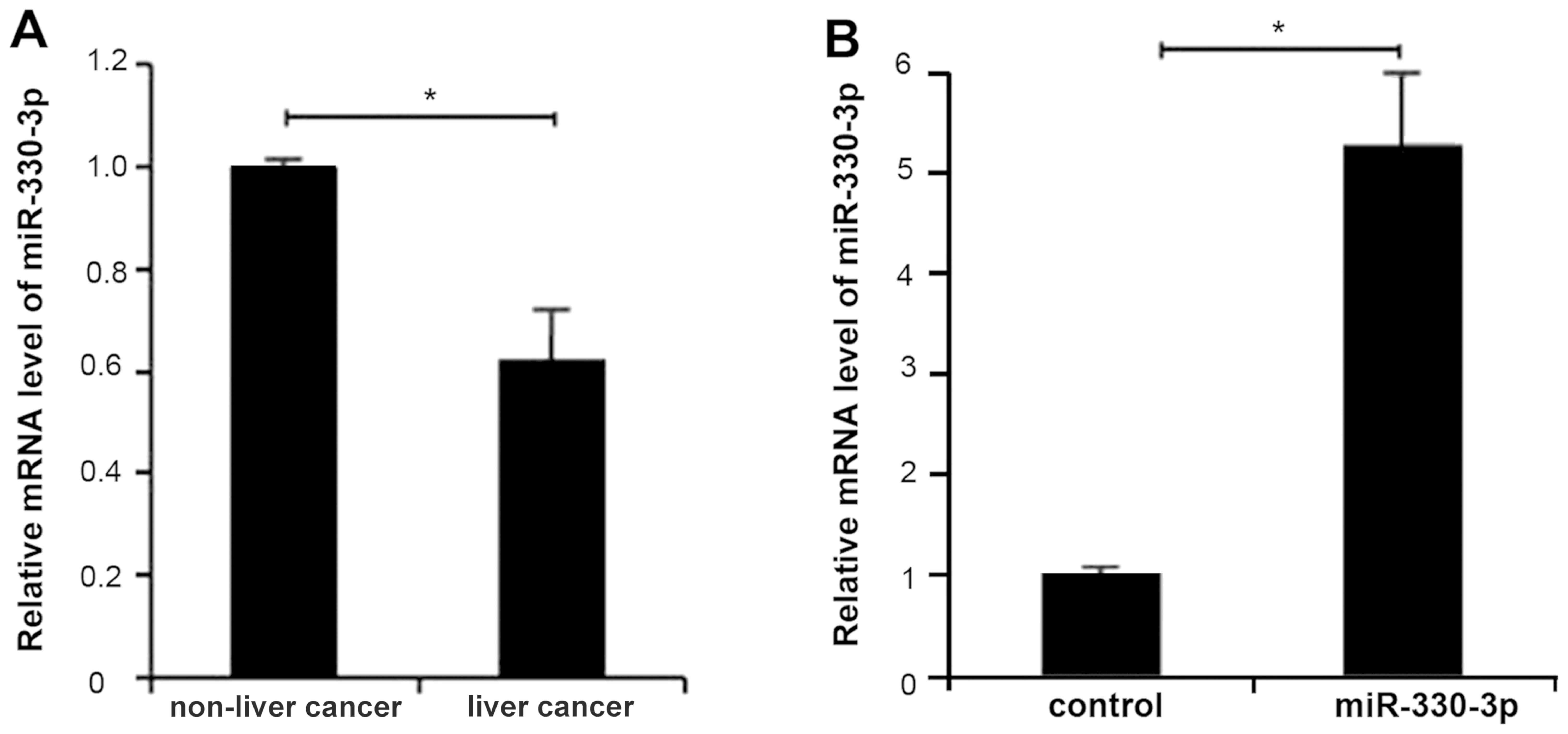
In order to determine the expression level of
miR-330-3p in liver cancer tissue, mRNAs were extracted from liver
cancer tissues and analyzed using RT-qPCR. Compared with normal
tissues, the expression of miR-330-3p in liver cancer tissues was
significantly decreased (P<0.05; Fig.
1A). The results suggested that deregulation of miR-330-3p was
associated with the progression of liver cancer.
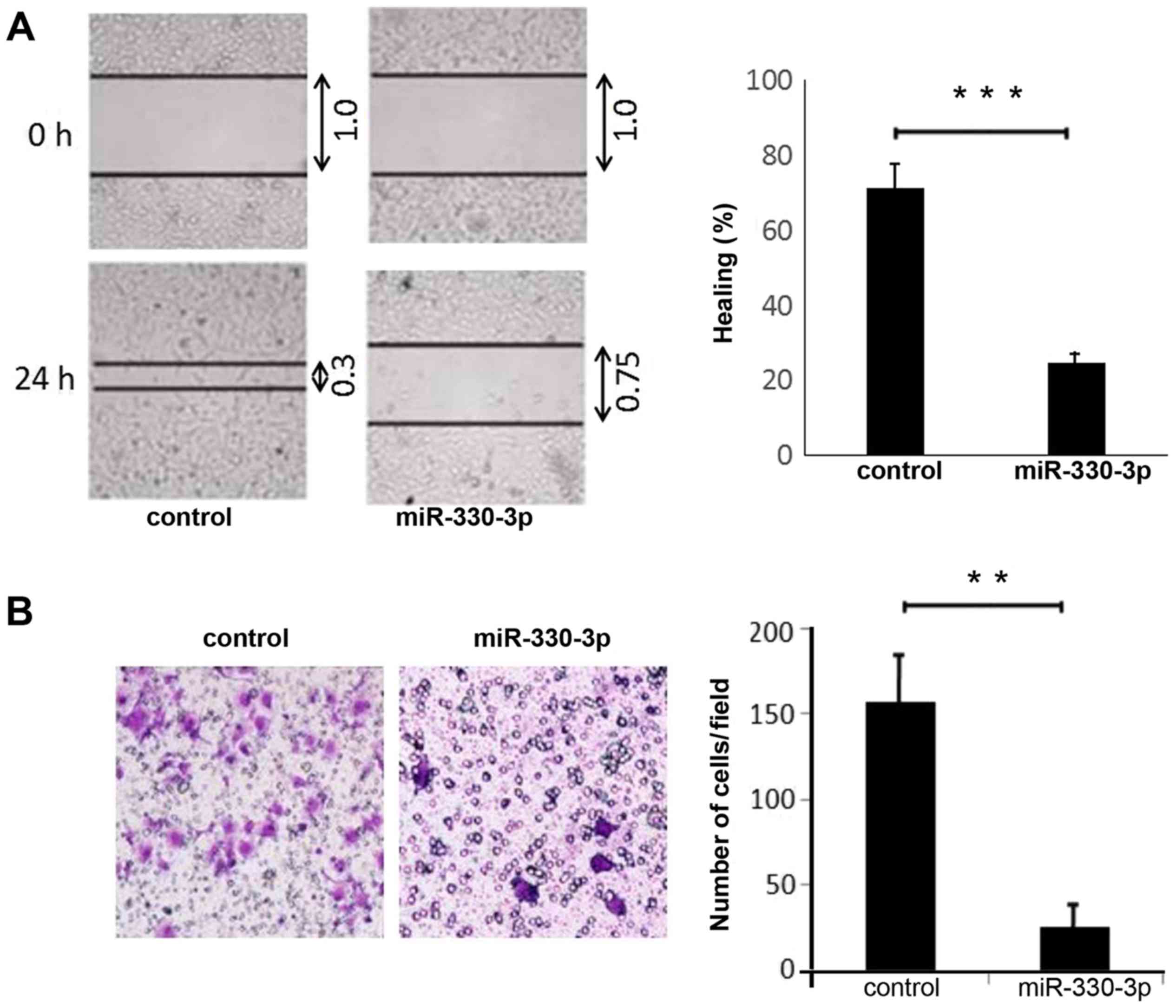
miR-330-3p suppresses migration of
liver cancer cells
miR-330-3p expression was significantly upregulated
in HepG2 cells by transfection of miR-330-3p mimic (P<0.05;
Fig. 1B). As observed in wound
healing assays, cell migration was inhibited by the expression of
miR-330-3p in HepG2 cells (P<0.01; Fig. 2A). Transwell assays confirmed that
overexpression of miR-330-3p significantly inhibited liver cancer
cell migration (P<0.05; Fig.
2B).
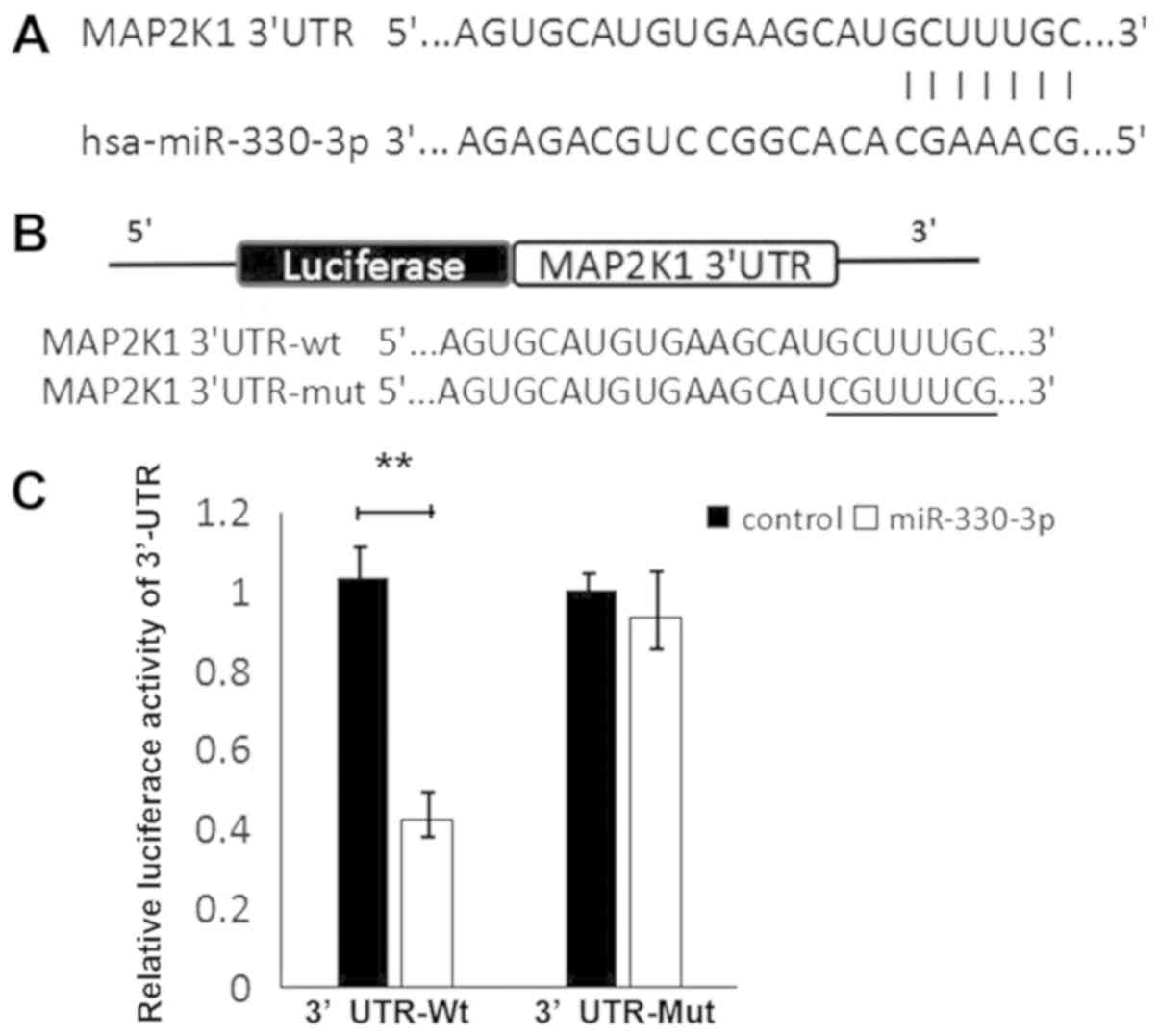
MAP2K1 is post-transcriptionally
downregulated by miR-330-3p
To investigate the underlying mechanism by which
miR-330-3p inhibits liver cancer cell migration, potential targets
of miR-330-3p were identified. As predicted using TargetScan,
PicTar and miRanda, MAP2K1 was selected as the candidate for
further research owing to its vital functions in tumorigenesis and
cancer progression. Wild-type and mutant MAP2K1 3′UTRs were
constructed and it was identified that miR-330-3p significantly
decreased luciferase activity in HepG2 cells transfected with
wild-type MAP2K1 3′UTR (P<0.01; Fig.
3), which was not observed following transfection with mutant
3′UTR.
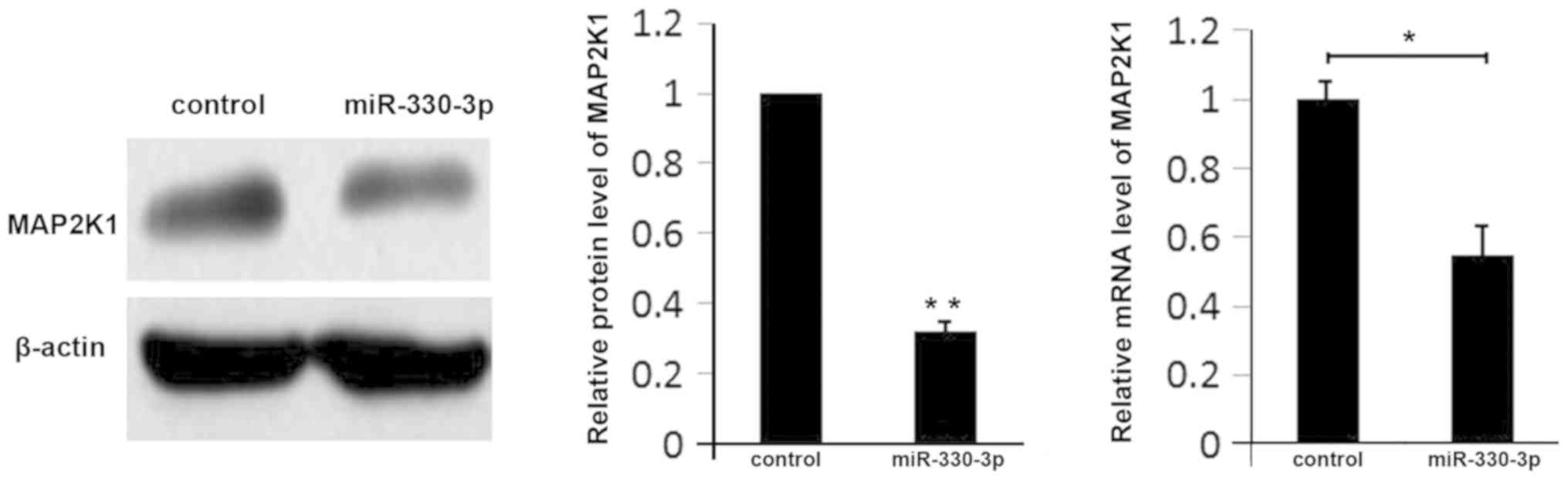
Conversely, overexpression of miR-330-3p in liver
cancer cells inhibited the expression of MAP2K1 at the mRNA and
protein levels (Fig. 4). These
results suggested that miR-330-3p regulates the expression of
MAP2K1 in liver cancer.
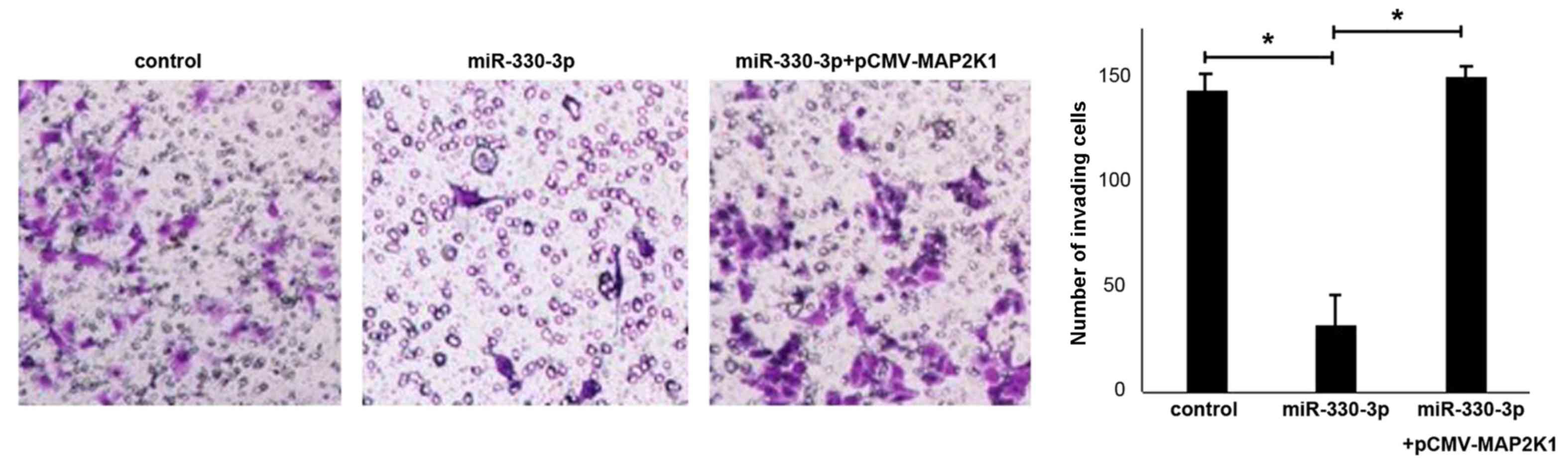
miR-330-3p inhibits liver cancer cell
migration by downregulating MAP2K1
To verify the function of MAP2K1 in the suppression
of liver cancer cell migration by miR-330-3p, HepG2 cells were
transfected with miR-330-3p together with the MAP2K1 expression
vector pCMV-MAP2K1. Compared with the control group, an increased
number of liver cancer cells transfected with miR-330-3p lost the
ability of migration, while cells co-transfected with pCMV-MAP2K1
exhibited increased migration abilities in the Transwell assay
(Fig. 5). The results suggested that
the increased expression of MAP2K1 contributed to the migratory
ability of HepG2 cells.
Discussion
Invasion typically leads to metastasis and these
events are primary reasons for the poor prognosis of liver cancer
(11). For liver cancer, surgical
resection is the most effective therapy, but only between 10 and
15% of patients are suitable for surgery; the majority of the
remaining patients exhibit distant metastases at diagnosis, which
renders curing liver cancer difficult (11,12).
Therefore, it is important to identifyfactors vital in invasion and
metastasis and the underlying mechanisms. Investigation into miRNAs
has led to novel studies to identify the molecular mechanism of
cancer cell metastasis (13,14), and therefore the development of
potential therapeutics in the treatment of human cancer.
Previous results support a fundamental function of
microRNAs in tumor invasion and metastasis; these biological
functions are associated with the deregulation of microRNAs in
various types of cancer (14).
Previous studies have suggested that miR-330-3pis deregulated in
the tissues and blood of patients with non-small-cell lung cancer
(15). In the present study, it was
identified that miR-330-3p was downregulated in patients with liver
cancer. Furthermore, miR-330-3p regulates the proliferation and
migration of prostate cancer cells (16). These results prompted the
investigation of whether miR-330-3p functioned in a similar manner
in liver cancer. Therefore, miR-330-3p was restored in liver cancer
cells by high efficiency transfection. The results revealed that
the migration of liver cancer cells was inhibited by miR-330-3p. By
combining the 3′UTRs of target genes, miRNAs were able to regulate
many genes. Studies to identify miRNA target genes have identified
several novel cancer invasion and metastasis suppressor genes.
Identification of new targets involved in the metastasis of liver
cancer may lead to novel diagnostic strategies or drug
development.
Mitogen-activated protein
kinase/extracellular-signal-regulated kinase activation has been
reported to be associated with the malignant progression of liver
cancer. Overexpression of activated MAP2K1 enhances cancer cell
proliferation and confers drug resistance on the cells (17). Using bioinformatic analysis, MAP2K1
was identified as one of the targets of miR-330-3p. It was also
observed that upregulation of miR-330-3p in liver cancer cells led
to downregulation of MAP2K1 at the RNA and protein levels. The
negative regulation of MAP2K1 by miR-330-3p was demonstrated using
a luciferase reporter assay. Furthermore, when MAP2K1 and
miR-330-3p were knocked down, the migratory ability of HepG2 cells
was also inhibited. These results suggested that the function of
miR-330-3p in suppressing migration of liver cancer cells was
mediated by MAP2K1.
As a number of miRNAs have been reported to be
associated with the progression of liver cancer (18–20),
those miRNAs may have synergistic or expanded effect on biological
behavior of tumor cells. Compared with pure blocking reagent,
miRNAs may have an advantage on influencing the tumor
microenvironment (21,22). In this regard, the results of the
present study provide novel insights into the suppressor function
of miR-330-3p and network of miRNAs in liver cancer. The focus of
future studies will be on the effect of miR-330-3p in
vivo.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contribution
ZJ designed the study, analyzed the data and wrote
the manuscript. BJ and LT performed the experiments and prepared
the figures. LT and YL collected the clinical specimens and
analyzed the clinical data. All authors reviewed the
manuscript.
Ethics approval and consent to
participate
All protocols dealing with the patients conformed to
the ethical guidelines of the Helsinki Declaration and were
approved by the ethics committee of First affiliated Hospital of
Jilin University. Patients provided written informed consent.
Patient consent for publication
Patients provided consent for the publication of any
data and associated images.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Forner A, Hessheimer AJ, Isabel Real M and
Bruix J: Treatment of hepatocellular carcinoma. Crit Rev Oncol
Hematol. 60:89–98. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jeng KS and Ching HJ: The role of surgery
in the management of unusual complications of transcatheter
arterial embolization for hepatocellular carcinoma. World J Surg.
12:362–368. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ryan BM, Robles AI and Harris CC: Genetic
variation in microRNA networks: The implications for cancer
research. Nat Rev Cancer. 10:389–402. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ma D, Tao X, Gao F, Fan C and Wu D:
miR-224 functions as an onco-miRNA in hepatocellular carcinoma
cells by activating AKT signaling. Oncol Lett. 4:483–488. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu H, Li W, Chen C, Pei Y and Long X:
MiR-335 acts as a potential tumor suppressor miRNA via
downregulating ROCK1 expression in hepatocellular carcinoma. Tumour
Biol. 36:6313–6319. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Y, Li T, Guo P, Kang J, Wei Q, Jia
X, Zhao W, Huai W, Qiu Y, Sun L and Han L: MiR-424-5p reversed
epithelial-mesenchymal transition of anchorage-independent HCC
cells by directly targeting ICAT and suppressed HCC progression.
Sci Rep. 4:62482014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen J, Wu X, Cai B, Su Z, Li L, An Y and
Wang L: Circulating microRNAs as potential biomarkers of HBV
infection persistence. Infect Genet Evol. 54:152–157. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kenneth J and Livak TD: Analysis of
relative gene expression data using rea l—time
quantitative PCR a nd the 2 □ ct method. Method. 2001.
|
|
11
|
Vitali GC, Laurent A, Terraz S, Majno P,
Buchs NC, Rubbia-Brandt L, Luciani A, Calderaro J, Morel P, Azoulay
D and Toso C: Minimally invasive surgery versus percutaneous radio
frequency ablation for the treatment of single small (</=3 cm)
hepatocellular carcinoma: A case-control study. Surg Endosc.
30:2301–2307. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xiong W, Sun LP, Chen XM, Li HY, Huang SA
and Jie SH: Comparison of microRNA expression profiles in
HCC-derived microvesicles and the parental cells and evaluation of
their roles in HCC. J Huazhong Univ Sci Technolog Med Sci.
33:346–352. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Deng S, Calin GA, Croce CM, Coukos G and
Zhang L: Mechanisms of microRNA deregulation in human cancer. Cell
Cycle. 7:2643–2646. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dinkova-Kostova AT, Jenkins SN, Fahey JW,
Ye L, Wehage SL, Liby KT, Stephenson KK, Wade KL and Talalay P:
Protection against UV-light-induced skin carcinogenesis in SKH-1
high-risk mice by sulforaphane-containing broccoli sprout extracts.
Cancer Lett. 240:243–252. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wei CH, Wu G, Cai Q, et al:
MicroRNA-330-3p promotes cell invasion and metastasis in non-small
cell lung cancer through GRIA3 by activating MAPK/ERK signaling
pathway. J Hematol Oncol. 10:1252017. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mao Y, Chen H, Lin Y, Xu X, Hu Z, Zhu Y,
Wu J, Xu X, Zheng X and Xie L: microRNA-330 inhibits cell motility
by downregulating Sp1 in prostate cancer cells. Oncol Rep.
30:327–333. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Huynh H, Nguyen TT, Chow KH, Tan PH, Soo
KC and Tran E: Over-expression of the mitogen-activated protein
kinase (MAPK) kinase (MEK)-MAPK in hepatocellular carcinoma: Its
role in tumor progression and apoptosis. BMC Gastroenterol.
3:192003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liang HH, Wei PL, Hung CS, Wu CT, Wang W,
Huang MT and Chang YJ: MicroRNA-200a/b influenced the therapeutic
effects of curcumin in hepatocellular carcinoma (HCC) cells. Tumour
Biol. 34:3209–3218. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen X, Zhang L, Zhang T, Hao M, Zhang X,
Zhang J, Xie Q, Wang Y, Guo M, Zhuang H and Lu F:
Methylation-mediated repression of microRNA 129-2 enhances
oncogenic SOX4 expression in HCC. Liver Int. 33:476–486. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhang D, Zhou P, Wang W, Wang X, Li J, Sun
X and Zhang L: MicroRNA-616 promotes the migration, invasion and
epithelial-mesenchymal transition of HCC by targeting PTEN. Oncol
Rep. 35:366–374. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Paiva I, Gil da Costa RM, Ribeiro J, Sousa
H, Bastos MM, Faustino-Rocha A, Lopes C, Oliveira PA and Medeiros
R: MicroRNA-21 expression and susceptibility to HPV-induced
carcinogenesis-role of microenvironment in K14-HPV16 mice model.
Life Sci. 128:8–14. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Paiva I, Gil da Costa RM, Ribeiro J, Sousa
H, Bastos M, Faustino-Rocha A, Lopes C, Oliveira PA and Medeiros R:
A role for microRNA-155 expression in microenvironment associated
to HPV-induced carcinogenesis in K14-HPV16 transgenic mice. PLoS
One. 10:e01168682015. View Article : Google Scholar : PubMed/NCBI
|