Introduction
Osteosarcoma (OS) is the most common type of
malignant primary bone neoplasm of mesenchymal origin. This
high-grade tumor has a high incidence within adolescents. The
metaphyseal regions of the long bones are the most common regions
for the development of OS; ~42% of cases of occurrence are in the
femur, 19% in the tibia, 10% in the humerus, and ~8% in the skull
and mandibula (1). Metastatic
disease is the major cause of cancer-associated mortality; the
lungs are the main sites of OS-related metastasis (2), followed by the bones. Unfortunately,
the 5-year survival rate of patients suffering from OS has not
improved in the last 20 years (3).
Therefore, the development of more effective therapeutic targets is
required for improving the survival rate of patients with OS.
However, the molecular mechanism underlying the progression and
metastasis of OS remains to be fully elucidated.
Quaking (QKI) is a member of the STAR family of
proteins and may regulate mRNA expression at the transcription
level (4–7). Increasing evidence shows that QKI may
act as a tumor suppressor gene, suppressing the occurrence and
progression of various types of tumor, including human astrocytic
tumors, glioblastoma multiforme, oral squamous cell carcinoma,
gastric cancer and colon cancer (8–15).
However, studies investigating the effects and mechanisms
underlying QKI in the progression and metastasis of OS are
required. MicroRNA (miR)-20a, one of the most well-characterized
oncomirs, is a member of the miR-17-92 cluster (16) and is upregulated in multiple types of
human tumor, including gastric cancer, non-small cell lung cancer,
multiple myeloma, colorectal cancer, prostate cancer and
gallbladder carcinoma (17–23). At present, few studies investigating
miR-20a in relation to OS have been performed. Namløs et al
(24) identified that members of the
miR-17-92 cluster, including miR-20a, are expressed at high levels
in OS cell lines relative to normal bone. Evidence indicates that
higher expression levels of miR-20a are significantly associated
with systemic spread and the OS of patients (25). However, the corresponding mechanisms
by which miR-20a functions in OS remain to be elucidated. In the
present study, the expression levels and effects of QKI2 in OS were
elucidated, and it was shown that miR-20a directly targets QKI2 in
OS.
Materials and methods
OS samples and cell lines
Primary OS samples and normal bone samples were
collected from 10 patients (6 males and 4 females) via biopsy at
Harbin Medical University Cancer Hospital (Harbin, China) between
June 2014 and December 2016. The mean age of the patients was 19.3
years (range, 9–67 years). All patients provided written informed
consent, and the study was approved by the Ethics Committee of
Harbin Medical University. The 143B human osteosarcoma cell line,
and human SV40-transfected hFOB 1.19 osteoblasts and 293TN cells
were obtained from the Cell Bank of Type Culture Collection of the
Chinese Academy of Sciences (Shanghai, China). The 143B cells were
cultured in EMEM (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 0.015 mg/ml 5-bromo-2′-deoxyuridine
(Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) and 10% FBS (Gibco;
Thermo Fisher Scientific, Inc.) at 37°C in a 5% CO2
atmosphere. The hFOB 1.19 cells were cultured in D-MEM/F-12 (Gibco;
Thermo Fisher Scientific, Inc.) supplemented with 0.3 mg/ml G418
(Gibco; Thermo Fisher Scientific, Inc.) and 10% FBS (Gibco; Thermo
Fisher Scientific, Inc.) at 34°C in a 5% CO2 atmosphere.
The 293TN cells were cultured in EMEM (Gibco; Thermo Fisher
Scientific, Inc.) supplemented with 10% FBS (Gibco; Thermo Fisher
Scientific, Inc.) at 37°C in a 5% CO2 atmosphere.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from samples and cells by
applying TRIzol reagent (Invitrogen; Thermo Fisher Scientific,
Inc.). The TransScript® One-Step gDNA Removal and cDNA
Synthesis SuperMix kit was purchased from Beijing Transgen Biotech
Co., Ltd. (Beijing, China). Then total RNA was converted into cDNA
through RT according to the manufacturer's protocol. Following RT,
qPCR was performed using the TransScript® Tip Green qPCR
SuperMix kit according to the manufacturer's protocol (Transgen
Biotech Co., Ltd.) with an ABI 7900HT Real-Time PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The following forward
and reverse primers were used for RT-qPCR: miR-20a, forward
5′-TAAAGTGCTTATAGTGCAG-3′ and reverse 5′-TGCGTGTCGTGGAGTC-3′ (LNA);
U6, forward 5′-CTCGCTTCGGCAGCACATATACT-3′ and reverse
5′-ACGCTTCACGAATTTGCGTGTC-3′. The PCR reaction mixtures contained
2× TransScript® Tip Green qPCR SuperMix (10 µl), 4 µM
primers (2 µl), 1 µl cDNA and 7 µl ddH2O in a total
volume of 20 µl. The PCR thermocycling conditions were as follows:
94°C for 30 sec, and 40 cycles of 5 sec at 94°C and 30 sec at 60°C.
The comparative 2−ΔΔCq method (26) was used for relative quantification
and statistical analysis.
Cell transfection
To establish 143B cells stably overexpressing QKI2,
pWPXL lentiviral vectors were used (Addgene, Inc., Cambridge, MA,
USA). To construct the plasmid overexpressing QKI2, the coding
sequence of QKI2 was subcloned into the pWPXL lentiviral vector.
The pWPXL lentiviral plasmid overexpressing QKI2 and the packaging
mix (pSPAX2 and pMD.2G) were cotransfected into 293TN cells. After
48 h, the virus-containing supernatant was harvested for the
infection of 143B cells. The miR-20a mimic
(5′-UAAAGUGCUUAUAGUGCAGGUAG-3′), inhibitor
(5′-CUACCUGCACUAUAAGCACACUUUA-3′), miR-negative control (miR-NC,
scrambled sequence; 5′-UCACAACCUCCUAGAAAGAGUAGA-3′), small
interfering (si)RNA-QKI2 (5′-GAAUUCAAGAACGGUCUUAAUU-3′) and siRNA
negative control (5′-AAUUUCUUCACUUCUUCAACUGCUC-3) were obtained
from Invitrogen; Thermo Fisher Scientific, Inc. and were
transfected into 143B cells by applying Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.).
Cell growth assay
To determine the effect of miR-20a and QKI2 on the
proliferative ability of the 143B cells, an MTT assay was
performed. The 143B cells (2×103 cells/well) were seeded
in 96-well plates and incubated in complete growth medium for 1, 2
and 3 days. Subsequently, 10 µl MTT solution was mixed into the
samples and then incubated for 4 h. Following removal of the
culture medium, 150 µl DMSO was added to samples. The cell
proliferative capacity was determined at a wavelength of 570
nm.
Transwell migration and invasion
assays
The transfected 143B cells (2×104) in 200
µl of FBS-free EMEM were seeded in the upper part of a Transwell
chamber (Costar; Corning Incorporated, Corning, NY, USA) with or
without Matrigel. Subsequently, the lower part of the Transwell
chamber was filled with complete growth medium. The 143B cells were
then incubated and allowed to migrate or invade, respectively for
24 h; the 143B cells that migrated or invaded into the lower part
were stained with 1% crystal violet for 30 min. Images of the
migrated or invaded 143B cells were captured under a light
microscope and counted using ImageJ 2.0 software (National
Institutes of Health, Bethesda, MD, USA).
Western blotting
The transfected 143B cells were lysed using RIPA
buffer, and the total protein was isolated and quantified by
applying a BCA protein assay kit (Beyotime Institute of
Biotechnology, Haimen, China). The proteins (30 µg per lane) were
resolved via 12% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and transferred onto PVDF membranes (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). Membranes were blocked with
5% non-fat dried milk in 1×TBS and 0.1% Tween-20 (TBST;
Sigma-Aldrich; Merck KGa) for 3 h at room temperature. The PVDF
membranes were incubated with rabbit polyclonal anti-human QKI
(1:5,000; catalog no. ab78518; Abcam, Cambridge, MA, USA) and mouse
monoclonal anti-human β-tubulin (1:5,000; catalog no. ab7751;
Abcam) antibodies overnight at 4°C. The membranes were then washed
with TBST, and horseradish peroxidase-conjugated secondary goat
anti-rabbit IgG (1:5,000; catalog no. ab6721; Abcam) or goat
anti-mouse IgG (1:5,000; catalog no. ab6789; Abcam) were applied
for 60 min at room temperature. The protein expression of QKI2 was
assessed using an ECL kit (Beyotime Institute of
Biotechnology).
miR-20a target prediction
The potential target genes of miR-20a were predicted
using miRanda (http://www.microrna.org/microrna/; August 2010
release) and TargetScan 7.2 (http://www.TargetScan.org). These databases search the
presence of conserved seven-mer and six-mer sites on the 3′-UTRs of
messenger RNA that match the seed region of miR-20a.
Dual-luciferase reporter assay
The wild-type (WT) and mutant (MUT) 3′-UTR fragment
of QKI2 were synthesized directly and separately inserted into the
pmiR-report plasmids. The 293TN cells were co-transfected with a
pmirGlo Dual-Luciferase miRNA Target Expression Vector (Promega
Corporation, Madison, WI, USA) containing the WT or MUT 3′-UTR
fragment of QKI2, and miR-20a mimic. The luciferase activities of
Firefly and Renilla were measured using a Dual-Luciferase
Reporter assay system (Promega Corporation).
Statistical analysis
Statistical analyses were performed using SPSS
version 17 (SPSS, Inc., Chicago, IL, USA). All statistical data are
reported as the mean ± standard deviation. Significant differences
between groups were assessed using Student's t-test (two-tailed).
P<0.05 was considered to indicate a statistically significant
difference.
Results
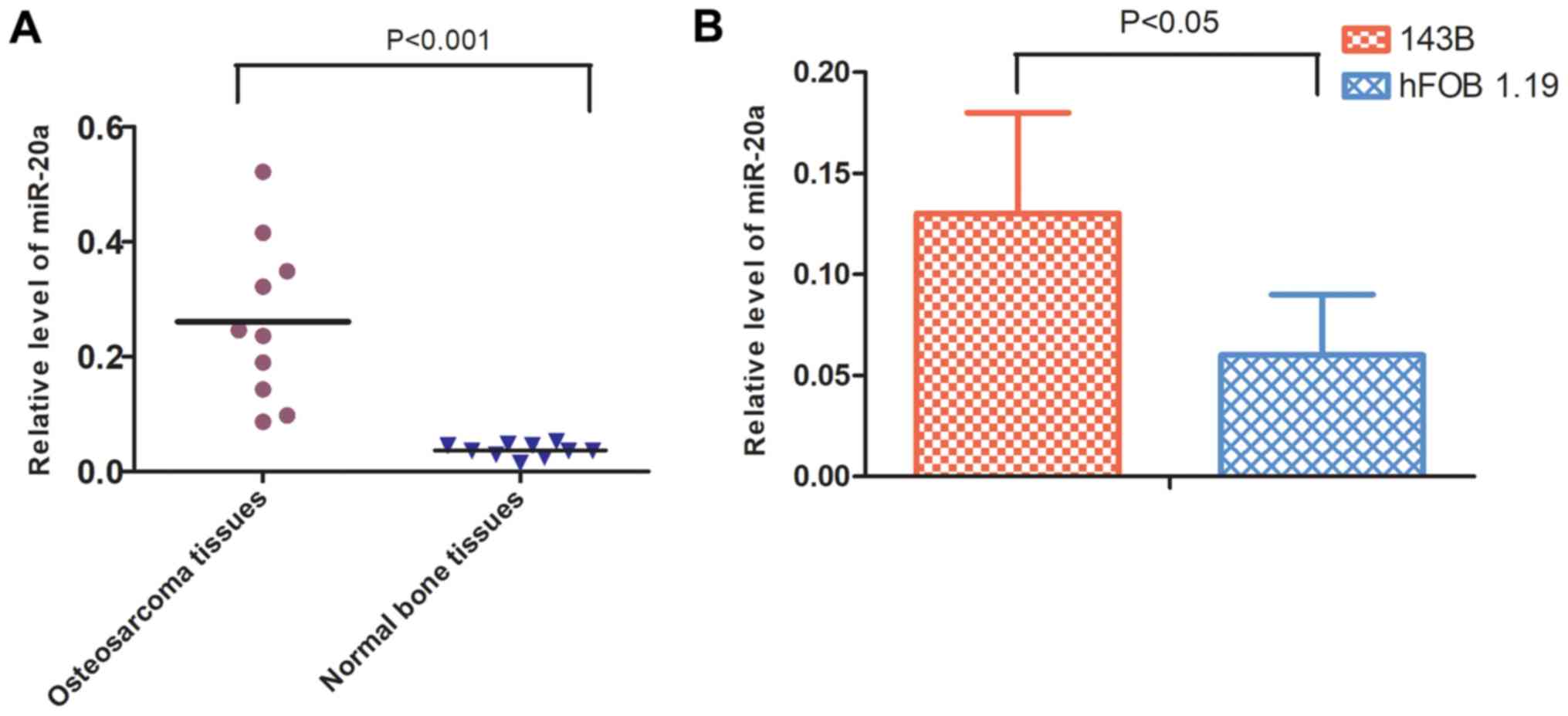
Expression levels of miR-20a in
OS
The expression levels of miR-20a in 10 primary OS
and normal bone biopsy samples were initially quantified. The
results of the RT-qPCR analysis showed that the expression levels
of miR-20a were higher in OS tissues compared with those in the
control group (Fig. 1A). Similarly,
the expression levels of miR-20a were higher in 143B cells compared
with those in hFOB 1.19 cells, also demonstrated by RT-qPCR
analysis (Fig. 1B).
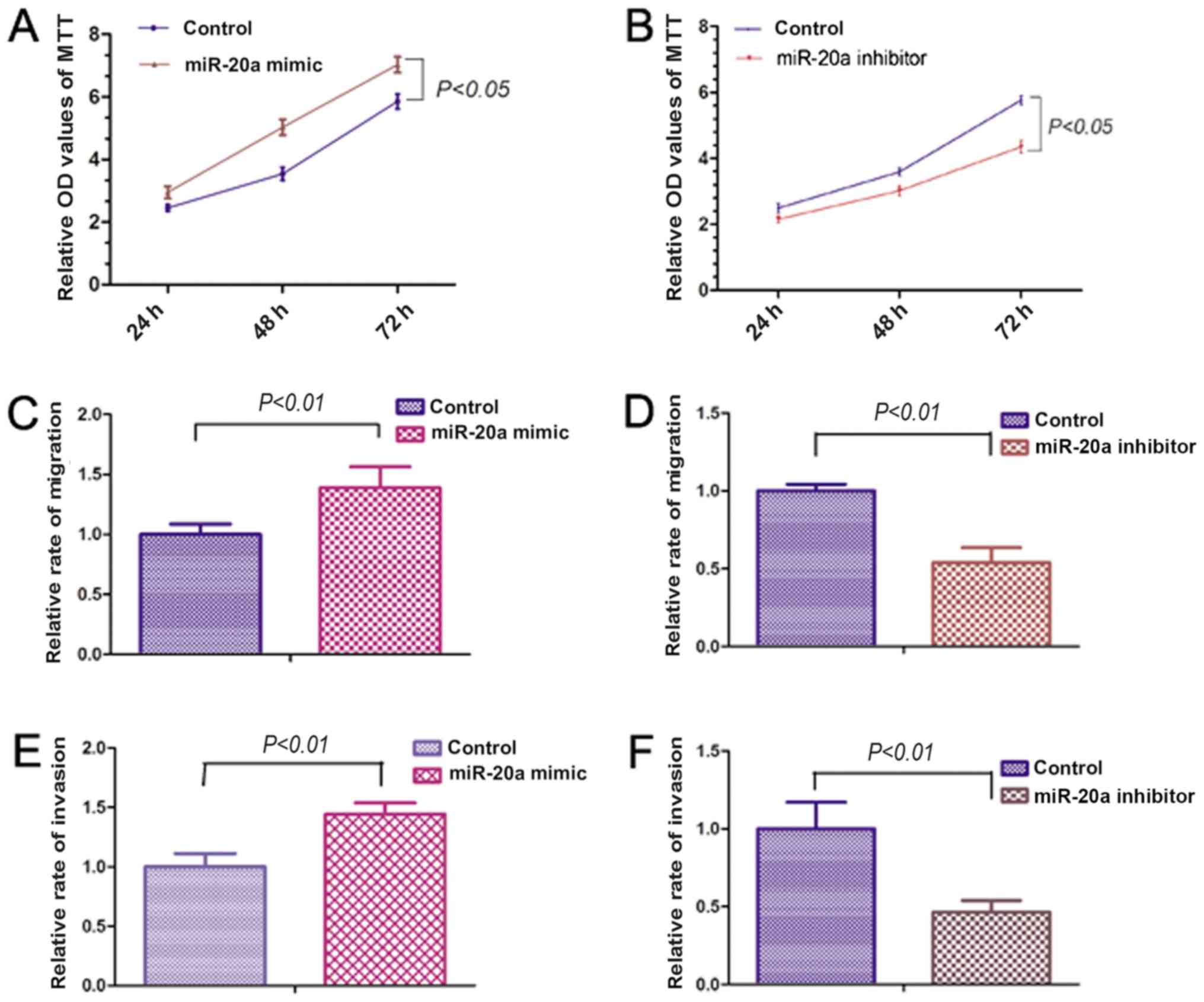
miR-20a promotes OS cell
proliferation, migration and invasion
In order to examine the effects of miR-20a on OS
cell proliferation, migration and invasion, 143B cells were
transfected with miR-20a mimic or inhibitor. The MTT assay showed
that the miR-20a mimic significantly increased the proliferation of
143B cells (Fig. 2A). However,
miR-20a inhibitor significantly reduced the proliferative capacity
of the 143B cells (Fig. 2B).
Migration and invasion assays were performed to investigate the
effects of miR-20a on the migration and invasive abilities of OS
cells. The miR-20a mimic significantly promoted the migration and
invasive abilities of 143B cells. By contrast, miR-20a inhibitor
reduced the migration and invasive abilities of the 143B cells
(Fig. 2C-F). The above results
indicate that miR-20a may be involved in the proliferation,
migration and invasion of OS cells.
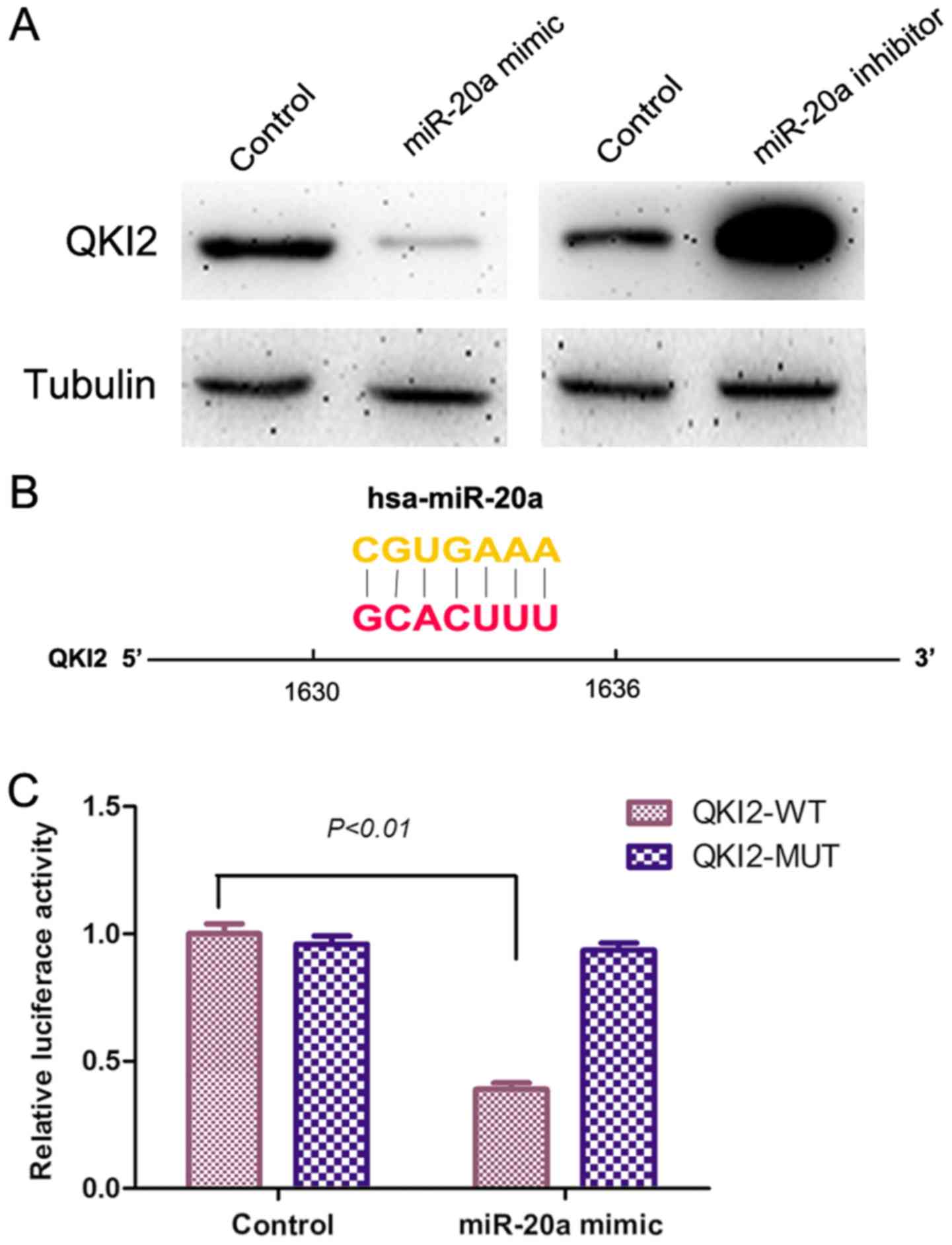
QKI2 is a direct target of
miR-20a
Using prediction tools, including TargetScan, QKI2
was hypothesized to be a direct target of miR-20a. In the present
study, no difference in the mRNA expression of QKI2 was observed
between the osteosarcoma tissue and normal bone tissue, or between
the 143B cells and hFOB 1.19 cells, determined using RT-qPCR
analysis. The protein expression of QK12 was decreased
significantly in the OS tissue and 143B cells, as determined using
western blot analysis. In order to further examine the association
between QKI2 and miR-20a in OS, the miR-20a mimic and inhibitor
were used for the transfection of 143B cells, following which the
protein expression levels of QKI2 in the transfected 143B cells
were detected using western blot analysis. The miR-20a mimic
reduced the protein expression of QKI2 in 143B cells compared with
that in the control group. However, the miR-20a inhibitor increased
the protein expression levels of QKI2 (Fig. 3A). These data supported the
hypothesis that QKI2 acts as a direct target of miR-20a.
Additionally, in order to confirm that miR-20a can directly target
QKI2 mRNA, a double fluorescence reporter assay was performed. The
results of this assay showed that the miR-20a mimic significantly
reduced the luciferase activity of the 293TN cells co-transfected
with the plasmid containing the WT 3′-UTR fragment of QKI2, whereas
the miR-20a mimic had no inhibitory effect when with the MUT QKI2
3′-UTR fragment (Fig. 3B and C).
Taken together, the above findings show that miR-20a inhibited the
protein expression of QKI2 by targeting the QKI2 3′-UTR in OS.
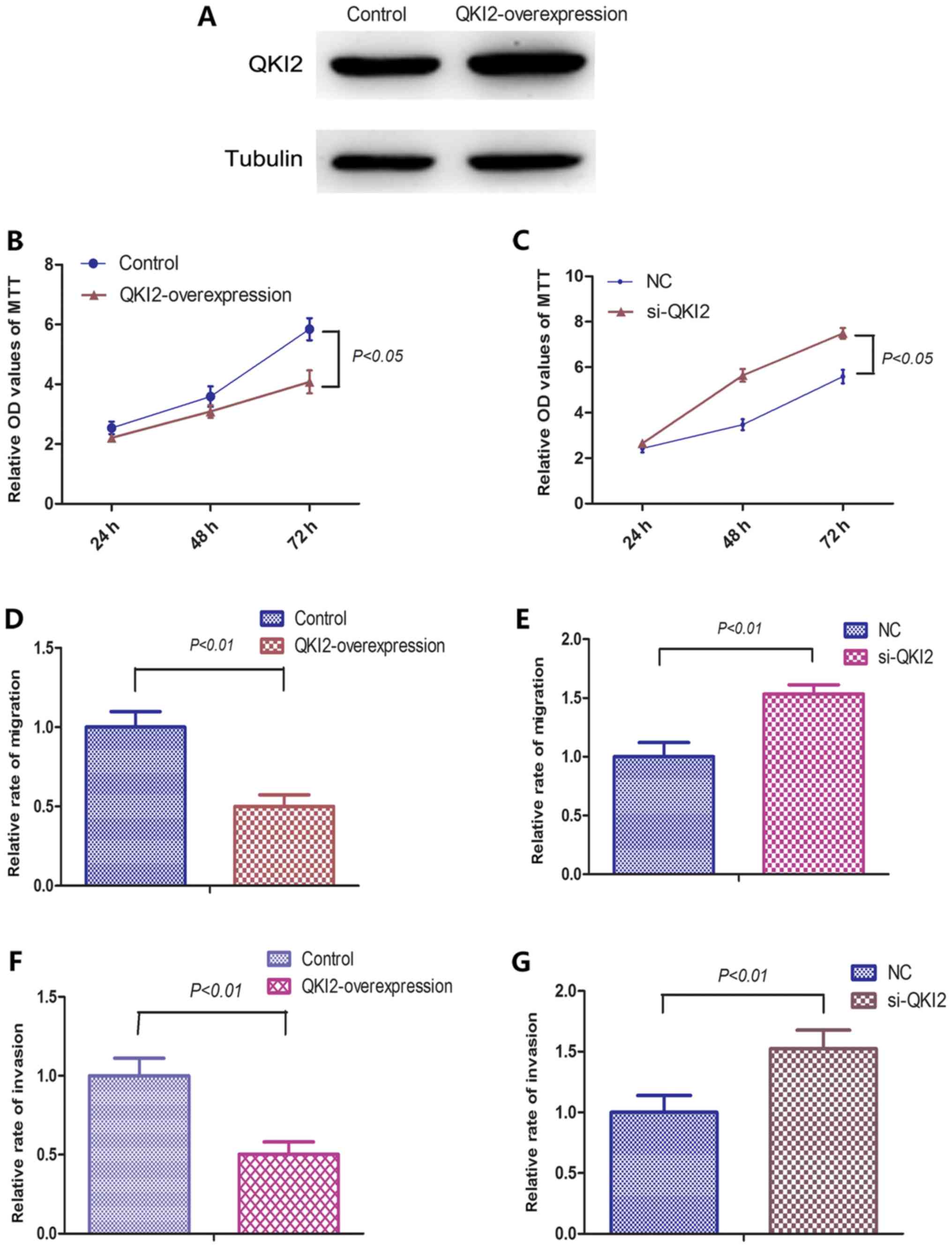
QKI2 inhibits OS cell proliferation,
migration and invasion
In order to further investigate the roles of QKI2 on
OS cells, 143B cells stably overexpressing QKI2 were established
using pWPXL lentiviral vectors. The subsequent western blot
analysis showed that the protein expression levels of QKI2 were
overexpressed in the stable cells (Fig.
4A). siRNAs against QKI2 were transfected into 143B cells to
inhibit the expression of QKI2. An MTT assay, and Transwell
migration and invasion assays were performed to examine the effects
of QKI2 on OS cell proliferation, migration and invasive abilities.
As shown in Figs. 4A-G and 5, the overexpression of QKI2 inhibited the
proliferative, migration and invasive abilities of the 143B cells,
whereas si-QKI2 evidently promoted the proliferation, migration and
invasion of 143B cells. Taken together, the above results suggested
that QKI2 acts as a tumor suppressor against the proliferation,
migration and invasion of OS cells.
Discussion
OS is the most common type of high-grade bone
neoplasm during adolescence. Although neoadjuvant chemotherapy is
administered, patients who develop relapse of disease and/or
metastatic disease have a severely poor prognosis (27). To improve the 5-year survival rate of
patients suffering from OS, further investigation of the cellular,
molecular and signal transduction mechanisms underlying the
initiation and progression of OS is required to develop novel and
specific molecular targeted therapeutic strategies. miRNAs are a
class of small, non-coding, single-stranded RNAs that are ~22
nucleotides in length and post-transcriptionally regulate the
expression of protein-coding genes by inhibiting mRNA translation
and/or directly cleaving target mRNA (28–33). In
previous years, miRNAs have been reported to act as important
regulators in cells (34). An
increasing number of studies have shown that dysregulated miRNAs in
cancer act as tumor suppressor genes or oncogenes (35,36).
Marginal dysregulation in miRNA expression levels can be sufficient
to affect the translation of protein-encoding genes. Consequently,
one issue influencing the analysis of miRNA expression in relation
to OS is that the majority of patients are treated with
chemotherapeutic drugs prior to surgery; as a result, the nucleic
acids required for analysis are degraded. In the present study,
miRNAs were extracted from primary OS biopsy samples collected from
untreated patients.
Several studies have indicated that miR-20a is a
member of the oncogenic miRNAs and that it is overexpressed in
multiple types of human tumor. One previous study demonstrated that
the expression levels of miR-20a in gastric cancer samples were
upregulated (8.9-fold) compared with those in normal adjacent
samples (17). Zhang et al
reported that the expression level of plasma miR-20a was
significantly upregulated in early-stage non-small cell lung cancer
samples when compared with control samples (18). miR-20a dysregulates the dynamic
balance of colonic epithelial cells by interfering with the
transforming growth factor-β-mediated regulation of Myc/p21, which
is necessary for the development of colorectal cancer (21). In the cancer stem cells of glioma,
the expression of miR-20a was reported to be upregulated and the
invasive ability of the cells significantly promoted by targeting
tissue inhibitor of metalloproteinase 2 (37). Another study showed that the
overexpression of miR-20a was associated with the metastases and
prognosis of patients suffering from OS (25). The present study revealed that the
expression of miR-20a was upregulated in OS tissues and OS cells.
To the best of our knowledge, the present study is the first to
show that miR-20a can enhance the proliferative, invasive and
migration abilities of OS cells.
The RNA-binding protein QKI is a member of the STAR
family of proteins, and can act as a tumor suppressor gene in
multiple types of human tumor. A previous study showed that QKI-5
reduces the proliferative ability of clear cell renal cell
carcinoma by post-transcriptionally regulating the
Ras-mitogen-activated protein kinase signaling pathway (38). de Miguel et al showed that
QKI, as a tumor suppressor, is associated with the prognosis of
lung carcinoma by targeting extended synaptotagmin-2 (39); the roles of QKI in OS remain to be
fully elucidated. In the present study, target genes regulated by
miR-20a were we screened using online databases miRanda and
TargetScan. It was shown that QKI2 may interact with miR-20a,
whereas other QKI members may not interact with miR-20a. Therefore,
QKI2 was selected for investigation. To the best of our knowledge,
the present study is among the first to demonstrate that QKI2 can
inhibit OS cell proliferation, invasion and migration. These
results demonstrated that miR-20a can directly target QKI2 mRNA in
OS.
In conclusion, the present study first identified
that miR-20a can inhibit the expression levels of QKI2 in OS by
targeting QKI2 mRNA, which subsequently promotes the proliferation,
invasion and migration of OS cells. The pathological role of the
‘miR-20a/QKI2’ axis in regulating the metastasis of OS was
demonstrated. These results not only improve our understanding of
the mechanism underlying the metastasis of OS, but also provide a
novel strategy and target for OS treatment. However, at present,
there are no agents available targeting miR-20a or QKI2 in the
clinical setting; further investigation is required to determine
these findings clinically.
The main purpose of the present study was to
investigate pathological role of miR-20a/QKI2 in the invasion and
migration of OS. The 143B cell line is a metastatic subline of HOS
and has the ability to metastasize, therefore the 143B cell line
was used in the present study. However, results obtained only in
the 143B cell line are not fully convincing. In future
investigations, experiments using other OS cell lines, including
human OS U2OS, Saos-2, HOS and MG-63 cell lines, and in vivo
experiments are to be performed.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The data used and analyzed in the present study are
available from the corresponding author on reasonable request.
Authors' contributions
YW conceived and designed the experiments; HY, YL
and ZP performed the experiments and analyzed the data; YW wrote
the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The present study was performed in compliance with
the Helsinki Declaration and approved by the Institutional Review
Board of the First Affiliated Hospital of Harbin Medical University
(IRB no. 2016-029). The data collection and analyses were performed
without disclosing patient identities.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Mirabello L, Troisi RJ and Savage SA:
Osteosarcoma incidence and survival rates from 1973 to 2004: Data
from the surveillance, epidemiology, and end results program.
Cancer. 115:1531–1543. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Longhi A, Errani C, De Paolis M, Mercuri M
and Bacci G: Primary bone osteosarcoma in the pediatric age: State
of the art. Cancer Treat Rev. 32:423–436. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Friebele JC, Peck J, Pan X, Abdel-Rasoul M
and Mayerson JL: Osteosarcoma: A meta-analysis and review of the
literature. Am J Orthop (Belle Mead NJ). 44:547–553.
2015.PubMed/NCBI
|
|
4
|
Kondo T, Furuta T, Mitsunaga K, Ebersole
TA, Shichiri M, Wu J, Artzt K, Yamamura K and Abe K: Genomic
organization and expression analysis of the mouse qkI locus. Mamm
Genome. 10:662–669. 1999.PubMed/NCBI
|
|
5
|
Galarneau A and Richard S: Target RNA
motif and target mRNAs of the Quaking STAR protein. Nat Struct Mol
Biol. 12:691–698. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhang Y, Lu Z, Ku L, Chen Y, Wang H and
Feng Y: Tyrosine phosphorylation of QKI mediates developmental
signals to regulate mRNA metabolism. EMBO J. 22:1801–1810. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yu F, Jin L, Yang G, Ji L, Wang F and Lu
Z: Post-transcriptional repression of FOXO1 by QKI results in low
levels of FOXO1 expression in breast cancer cells. Oncol Rep.
31:1459–1465. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ichimura K, Mungall AJ, Fiegler H, Pearson
DM, Dunham I, Carter NP and Collins VP: Small regions of
overlapping deletions on 6q26 in human astrocytic tumours
identified using chromosome 6 tile path array-CGH. Oncogene.
25:1261–1271. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Li ZZ, Kondo T, Murata T, Ebersole TA,
Nishi T, Tada K, Ushio Y, Yamamura K and Abe K: Expression of Hqk
encoding a KH RNA binding protein is altered in human glioma. Jpn J
Cancer Res. 93:167–177. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mulholland PJ, Fiegler H, Mazzanti C,
Gorman P, Sasieni P, Adams J, Jones TA, Babbage JW, Vatcheva R,
Ichimura K, et al: Genomic profiling identifies discrete deletions
associated with translocations in glioblastoma multiforme. Cell
Cycle. 5:783–791. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yin D, Ogawa S, Kawamata N, Tunici P,
Finocchiaro G, Eoli M, Ruckert C, Huynh T, Liu G, Kato M, et al:
High-resolution genomic copy number profiling of glioblastoma
multiforme by single nucleotide polymorphism DNA microarray. Mol
Cancer Res. 7:665–677. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Fu X and Feng Y: QKI-5 suppresses cyclin
D1 expression and proliferation of oral squamous cell carcinoma
cells via MAPK signalling pathway. Int J Oral Maxillofac Surg.
44:562–567. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bian Y, Wang L, Lu H, Yang G, Zhang Z, Fu
H, Lu X, Wei M, Sun J, Zhao Q, et al: Downregulation of tumor
suppressor QKI in gastric cancer and its implication in cancer
prognosis. Biochem Biophys Res Commun. 422:187–193. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yang G, Fu H, Zhang J, Lu X, Yu F, Jin L,
Bai L, Huang B, Shen L, Feng Y, et al: RNA-binding protein quaking,
a critical regulator of colon epithelial differentiation and a
suppressor of colon cancer. Gastroenterology. 138:231–240.e1-5.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen AJ, Paik JH, Zhang H, Shukla SA,
Mortensen R, Hu J, Ying H, Hu B, Hurt J, Farny N, et al: STAR
RNA-binding protein Quaking suppresses cancer via stabilization of
specific miRNA. Genes Dev. 26:1459–1472. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Mendell JT: miRiad roles for the miR-17-92
cluster in development and disease. Cell. 133:217–222. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Jafarzadeh-Samani Z, Sohrabi S,
Shirmohammadi K, Effatpanah H, Yadegarazari R and Saidijam M:
Evaluation of miR-22 and miR-20a as diagnostic biomarkers for
gastric cancer. Chin Clin Oncol. 6:162017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang H, Mao F, Shen T, Luo Q, Ding Z,
Qian L and Huang J: Plasma miR-145, miR-20a, miR-21 and miR-223 as
novel biomarkers for screening early-stage non-small cell lung
cancer. Oncol Lett. 13:669–676. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Babu KR and Muckenthaler MU: miR-20a
regulates expression of the iron exporter ferroportin in lung
cancer. J Mol Med (Berl). 94:347–359. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peng J, Thakur A, Zhang S, Dong Y, Wang X,
Yuan R, Zhang K and Guo X: Expressions of miR-181a and miR-20a in
RPMI8226 cell line and their potential as biomarkers for multiple
myeloma. Tumour Biol. 36:8545–8552. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Sokolova V, Fiorino A, Zoni E, Crippa E,
Reid JF, Gariboldi M and Pierotti MA: The effects of miR-20a on
p21: Two mechanisms blocking growth arrest in TGF-β-responsive
colon carcinoma. J Cell Physiol. 230:3105–3114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pesta M, Klecka J, Kulda V, Topolcan O,
Hora M, Eret V, Ludvikova M, Babjuk M, Novak K, Stolz J and Holubec
L: Importance of miR-20a expression in prostate cancer tissue.
Anticancer Res. 30:3579–3583. 2010.PubMed/NCBI
|
|
23
|
Chang Y, Liu C, Yang J, Liu G, Feng F,
Tang J, Hu L, Li L, Jiang F, Chen C, et al: MiR-20a triggers
metastasis of gallbladder carcinoma. J Hepatol. 59:518–527. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Namløs HM, Meza-Zepeda LA, Barøy T,
Østensen IH, Kresse SH, Kuijjer ML, Serra M, Bürger H,
Cleton-Jansen AM and Myklebost O: Modulation of the osteosarcoma
expression phenotype by microRNAs. PLoS One. 7:e480862012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Arabi L, Gsponer JR, Smida J, Nathrath M,
Perrina V, Jundt G, Ruiz C, Quagliata L and Baumhoer D:
Upregulation of the miR-17-92 cluster and its two paraloga in
osteosarcoma - reasons and consequences. Genes Cancer. 5:56–63.
2014.PubMed/NCBI
|
|
26
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wu PK, Chen WM, Chen CF, Lee OK, Haung CK
and Chen TH: Primary osteogenic sarcoma with pulmonary metastasis:
Clinical results and prognostic factors in 91 patients. Jpn J Clin
Oncol. 39:514–522. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Costa FF: Non-coding RNAs, epigenetics and
complexity. Gene. 410:9–17. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun J, Liu HP, Deng JE and Zhou M:
Systematic analysis of genomic organization and heterogeneities of
miRNA cluster in vertebrates. Mol Biol Rep. 39:5143–5149. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Wang QH, Zhou M, Sun J, Ning SW, Li Y,
Chen L, Zheng Y and Li X, Lv SL and Li X: Systematic analysis of
human microRNA divergence based on evolutionary emergence. FEBS
Lett. 585:240–248. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhou M, Wang Q, Sun J, Li X, Xu L, Yang H,
Shi H, Ning S, Chen L, Li Y, et al: In silico detection and
characteristics of novel microRNA genes in the Equus
caballus genome using an integrated ab initio and comparative
genomic approach. Genomics. 94:125–131. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hwang HW and Mendell JT: MicroRNAs in cell
proliferation, cell death, and tumorigenesis. Br J Cancer.
94:776–780. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Passetti F, Ferreira CG and Costa FF: The
impact of microRNAs and alternative splicing in pharmacogenomics.
Pharmacogenomics J. 9:1–13. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Chen CZ: MicroRNAs as oncogenes and tumor
suppressors. N Engl J Med. 353:1768–1771. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wang Z, Wang B, Shi Y, Xu C, Xiao HL, Ma
LN, Xu SL, Yang L, Wang QL, Dang WQ, et al: Oncogenic miR-20a and
miR-106a enhance the invasiveness of human glioma stem cells by
directly targeting TIMP-2. Oncogene. 34:1407–1419. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang RL, Yang JP, Peng LX, Zheng LS, Xie
P, Wang MY, Cao Y, Zhang ZL, Zhou FJ, Qian CN and Bao YX:
RNA-binding protein QKI-5 inhibits the proliferation of clear cell
renal cell carcinoma via post-transcriptional stabilization of
RASA1 mRNA. Cell Cycle. 15:3094–3104. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
de Miguel FJ, Pajares MJ, Martínez-Terroba
E, Ajona D, Morales X, Sharma RD, Pardo FJ, Rouzaut A, Rubio A,
Montuenga LM and Pio R: A large-scale analysis of alternative
splicing reveals a key role of QKI in lung cancer. Mol Oncol.
10:1437–1449. 2016. View Article : Google Scholar : PubMed/NCBI
|