Introduction
Acute myeloid leukemia (AML) originates from
hematopoietic stem/progenitor cells characterized by a high degree
of heterogeneity. The diagnostic criteria for AML has shifted from
the French-American-British (FAB) criteria for morphological
diagnosis to the World Health Organization (WHO) criteria, which
includes alterations in cytogenetic and molecular biological
characterization (1,2), demonstrating the importance of
cytogenetic and molecular biological characteristics as important
prognostic factors of AML. Therefore, identification of specific
genes involved in leukemogenesis may improve the diagnosis and
treatment of AML.
In a previous study, the genomes of 24 patients with
acute monocytic leukemia were scanned by high-resolution array
comparative genome hybridization (aCGH) technology. A total of two
patients presented with a deletion of chromosome 14 involving the
forkhead box protein N (FOXN)3 gene (3). FOXN3 was first identified by Pati et
al (4) in a yeast cell with
multiple checkpoint mutations. It is a subtype of the forkhead box
protein (FOX) transcription factor family, and is referred to as
checkpoint suppressor 1 (5,6). FOXN3 possesses an important role in the
development of cells and tissues (7,8). It is
crucial for the development of brain cartilage and indirectly
influences the development of muscle morphology (7). During cellular DNA damage, FOXN3 may
prolong cell survival by inducing cell quiescence (9,10). As a
DNA damage response protein, FOXN3 restored cell cycle arrest
(S-phase) in the mutant fruit fly (11). Previous studies demonstrated the
ability of FOXN3 to decrease the malignancy of tumors, including in
liver cancer, lung cancer, colon cancer and certain hematological
malignancies (12–15). However, the role of FOXN3 in AML is
not yet understood, to the best of the authors' knowledge. It is
hypothesized that FOXN3 is abnormally expressed in patients with
AML and may serve as a tumor suppressor gene contributing to the
transformation of leukemia.
In the present study, FOXN3 expression and its
association with clinicopathological features of AML were
investigated in patients with AML. The role of FOXN3 in promoting
an AML phenotype was further studied in AML cell lines in
vitro.
Materials and methods
Patient samples and cell lines
A total of 53 patients with recently diagnosed AML
(47 patients were confirmed with AML at diagnosis and 6 patients
were confirmed at relapse), who were admitted to The First
Affiliated Hospital of China Medical University (Shenyang, China)
between March 2012 and May 2013 and fulfilled the diagnostic
criteria of FAB and WHO (1,2), were included in the present study.
Patients who were ≤14 years old or patients with secondary AML
(transformed from myelodysplastic syndrome or myeloproliferative
disease) were excluded from the present study. The clinical
characteristics of patients are summarized in Table I. The treatment response was
evaluated as previously described (3). The definition of overall survival (OS)
refers to the time between randomization and mortality.
Relapse-free survival (RFS) refers to the time between complete
remission (CR) and disease recurrence. The present study was
approved by The Ethics Committee of The First Affiliated Hospital
of China Medical University. Prior to data collection, written
informed consent for participation in the present study and for
publishing the present results was obtained from each participant.
Bone marrow samples were collected from 14 normal donors between
May and August 2017 to serve as healthy controls. The average age
was 45 years old with 8 male and 6 female participants.
 | Table I.Clinicopathological characteristics
of patients with acute myeloid leukemia. |
Table I.
Clinicopathological characteristics
of patients with acute myeloid leukemia.
| Clinicopathological
characteristics | Number of
patients | % |
|---|
| Sex |
|
|
|
Male | 27 | 50.94 |
|
Female | 26 | 49.06 |
| Age, years |
|
|
|
≤60 | 40 | 75.47 |
|
>60 | 13 | 24.53 |
| White blood cell
count, 109/l |
|
<30 | 39 | 73.58 |
|
≥30 | 14 | 26.42 |
| Hemoglobin,
g/l |
|
|
|
≤80 | 37 | 69.81 |
|
>80 | 16 | 30.19 |
| Platelet,
109/l |
|
|
|
≤50 | 36 | 67.92 |
|
>50 | 17 | 32.08 |
| Blasts, % |
|
|
|
≤70 | 27 | 50.94 |
|
>70 | 26 | 49.06 |
| FAB subtype |
|
|
| M2 | 13 | 24.53 |
| M3 | 8 | 15.09 |
| M5 | 25 | 47.17 |
| Other
type | 7 | 13.21 |
| Karyotype |
|
|
|
Normal | 17 | 32.08 |
|
t(15;17) | 8 | 15.09 |
|
t(8;21) | 2 |
3.77 |
|
Complex | 11 | 20.76 |
| Not
available | 15 | 28.30 |
Human normal 293T cells and AML cell lines OCI-AML3
(OA3), NB4, U937, JURKAT and K562 were purchased from The American
Type Culture Collection (Manassas, VA, USA), and KASUMI-1 and
THP-AML (THP-1) cell lines were purchased from The Type Culture
Collection of the Chinese Academy of Medical Sciences (Shanghai,
China). Cells were cultured with RPMI-1640 medium (Thermo Fisher
Scientific, Inc., Waltham, MA, USA) containing 10% fetal bovine
serum (Gibco; Thermo Fisher Scientific, Inc.) at 37°C in a
humidified incubator with 5% CO2.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from bone marrow specimens
of patients with AML and AML cell lines using TRIzol®
(Invitrogen; Thermo Fisher Scientific, Inc.). RT was performed at
37°C for 15 min and then 85°C for 15 min using a genomic DNA Eraser
RT kit (Takara Bio, Inc., Otsu, Japan). The relative FOXN3
expression levels were calculated and normalized using the
comparative quantification cycle 2−ΔΔCq method relative
to an endogenous control GAPDH according to a previous study
(16). RT-qPCR was performed as
previously described by Zhang et al (3). The primer sequences are listed in
Table II.
| Table II.Primer sequences in reverse
transcription-quantitative polymerase chain reaction. |
Table II.
Primer sequences in reverse
transcription-quantitative polymerase chain reaction.
| Gene name | Sequence,
5′-3′ |
|---|
| FOXN3 | F:
TGCCAATCACTCCCATTGGG |
|
| R:
CCGCATCCGGCAGCTGG |
| GAPDH | F:
CGGATTTGGTCGTATTGGG |
|
| R:
CTGGAAGATGGTGATGGGATT |
Immunohistochemistry
To assess the FOXN3 protein expression levels in
leukemia blasts, immunohistochemical staining was performed using
the immunoperoxidase technique and an avidin-biotin-peroxidase
complex as previously described (17). Cytospin smears of bone marrow cells
from AMLs with a blast ratio >80% and normal controls (bone
marrow transplant donor) were fixed in formalin acetone (3%) at
room temperature for 3 min. The specimens were incubated with
peroxidase (reagent 1) from a streptavidin-peroxidase (SP) link
detection kit [Biotin-Streptavidin horseradish peroxidase (HRP)
Detection Systems; cat. no. SP9001; OriGene Technologies, Inc.,
Rockville, MD, USA) blocking enzyme for 20 min and then incubated
with normal goat serum (reagent 2 from the kit) for blocking at
room temperature for 15 min, followed by incubation with rabbit
anti-human antibodies against FOXN3 protein (cat. no. HPA059209;
1:50 in PBS; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C
for 60 min. Biotinylated goat anti-rabbit immunoglobulin G (reagent
3 from the kit) was used as the secondary antibody, incubated at
room temperature for 15 min, and detected using the SP complex
(reagent 4 from the kit). The specimens were counterstained with
hematoxylin for 20 sec at room temperature. Antibody reactivity was
detected with vector brown avidin-biotin-peroxidase substrate using
an Olympus BX51 optical microscope (magnification, ×1000).
Construction of the lentiviral vector
and cell transfection
The coding sequence of FOXN3 was amplified by
RT-qPCR using oligo to design the primers (http://www.genepharma.com/) for the human FOXN3 gene.
The primers were synthesized by Shanghai GenePharma Co., Ltd.
(Shanghai, China) and were as follows: Forward
5′-TCTACTAGAGGATCCACTAGTGCCACCAGGGTCCAGTCATGCCTCCCAGTAAGAAG-3′ and
Reverse 3′-GTCGGGCCCTCTAGACYCGAGTTAATTTTTTGTGGTTTCCTTTTGCTCTTT-5′.
A PLEX vector (Shanghai GenePharma Co., Ltd.) was digested with
SpeI and XhoI (Fermentas; Thermo Fisher Scientific,
Inc.) and the FOXN3 gene was cloned into the vector. Positive
clones were screened using ampicillin as a selection pressure
(Sigma-Aldrich; Merck KGaA) and identified by Sanger sequencing on
recombinant plasmid. For cells in suspension, a six-well plate was
coated with fibronectin (ProSpec-Tany TechnoGene Ltd., East
Brunswick, NJ, USA) and incubated at 37°C on a plate for at least
30 min. A total of 5×105 cells/well were plated in a
final volume of 1 ml culture media. For infection, cells were
plated onto 6-well plates at 1×105 cells/well and
infected with lentiviral stocks at a multiplicity of infection of
100 in the presence of polybrene (8 µg/ml; Sigma-Aldrich; Merck
KGaA). Puromycin at a final concentration of 10 µg/ml
(Sigma-Aldrich; Merck KGaA) was added to the media 72 h after
transfection to select for stably transfected cells. The FOXN3
expression levels in stably transfected cell lines were examined by
RT-qPCR and western blot analysis (18).
Western blot analysis
The cell lines were lysed using a whole protein
extraction kit (Nanjing KeyGen Biotech, Co., Ltd., Nanjing, China).
The total protein concentration was determined using a
bicinchoninic acid protein kit, and 30 μg protein was loaded per
lane. The proteins were separated by 10% SDS-PAGE, transferred to a
polyvinylidene fluoride membrane and blocked with 5% (m/v) fat-free
milk for 60 min at room temperature. The membrane was incubated at
4°C overnight with the following primary antibodies: Anti-FOXN3
(1:2,500; cat. no. ab129453; Abcam, Cambridge, UK) and anti-GAPDH
(1:1,000; cat. no. 5174; Cell Signaling Technology, Inc., Danvers,
MA, USA). Specific bands were detected with an anti-rabbit
immunoglobulin G, HRP-conjugated secondary antibody (1:2,000; cat.
no. 7074; Cell Signaling Technology, Inc.) for 60 min at room
temperature, and visualized using an Electro-Chemiluminescence plus
chemiluminescence kit (cat. no. WBKLS0500; EMD Millipore,
Billerica, MA, USA).
Cell growth analysis
The cells were counted, and the cell suspension was
diluted to a final concentration of 2.5×104/ml. The cell
suspension was seeded into a 96-well plate (100 µl/well).
Subsequent to adding 10 µl Cell Counting Kit-8 (CCK-8; Dojindo,
Molecular Technologies, Inc., Kumamoto, Japan) solution to each
well, the plates were incubated for 1 h in the incubator and the
absorbance at 450 nm was measured with a microplate reader after
24, 48 and 72 h. Cell proliferation curves were plotted based on
the absorbance values.
Flow cytometry
Apoptosis assays were performed using a flow
cytometer with an Annexin V phycoerythrin/7-amino-actinomycin D
cell apoptosis kit (cat. no. 550825; BD Biosciences, San Jose, CA,
USA). The cell cycle was assessed using a cell cycle kit (cat. no.
559763; BD Biosciences) with propidium iodide. Specific steps for
flow cytometry were as previously described (19). FlowJo software version 10 (TreeStar,
Inc., Ashland, OR, USA) was used for flow cytometry analysis.
Statistical analysis
All statistical analyses were performed using SPSS
22.0 (IBM Corp., Armonk, NY, USA) and GraphPad Prism 6.0 software
(GraphPad Software, Inc., La Jolla, CA, USA). The mRNA expression
of FOXN3 was repeated three times and is presented as the mean ±
standard deviation. A Student's t-test was used to compare FOXN3
expression levels between patients with AML and healthy controls.
Associations between FOXN3 expression levels and
clinicopathological parameters were analyzed using non-parametric
tests (Mann-Whitney U test between two groups or Kruskal-Wallis
test between three or more groups). A Spearman's rank correlation
coefficient analysis was used to determine whether there was a
correlation between FOXN3 expression levels and white blood cell
(WBC) count. CR rates were compared between groups using a
χ2 test. OS and the 2-year RFS curves were plotted using
Kaplan-Meier curves, and the difference was tested using a log-rank
test. Analysis of variance with a Tukey's post-hoc test was used to
compare the mRNA expression difference between AML lines and 293T
cell line, as well as the difference in cell proliferation, cell
cycle and apoptosis between the control and FOXN3 overexpression
cell lines. P<0.05 was considered to indicate a statistically
significant difference.
Results
FOXN3 expression levels are decreased
in patients with AML
FOXN3 expression levels were detected in 53 bone
marrow specimens of patients with AML present in the initial
diagnosis and 14 normal healthy controls using RT-qPCR. FOXN3
expression levels were significantly decreased in patients with AML
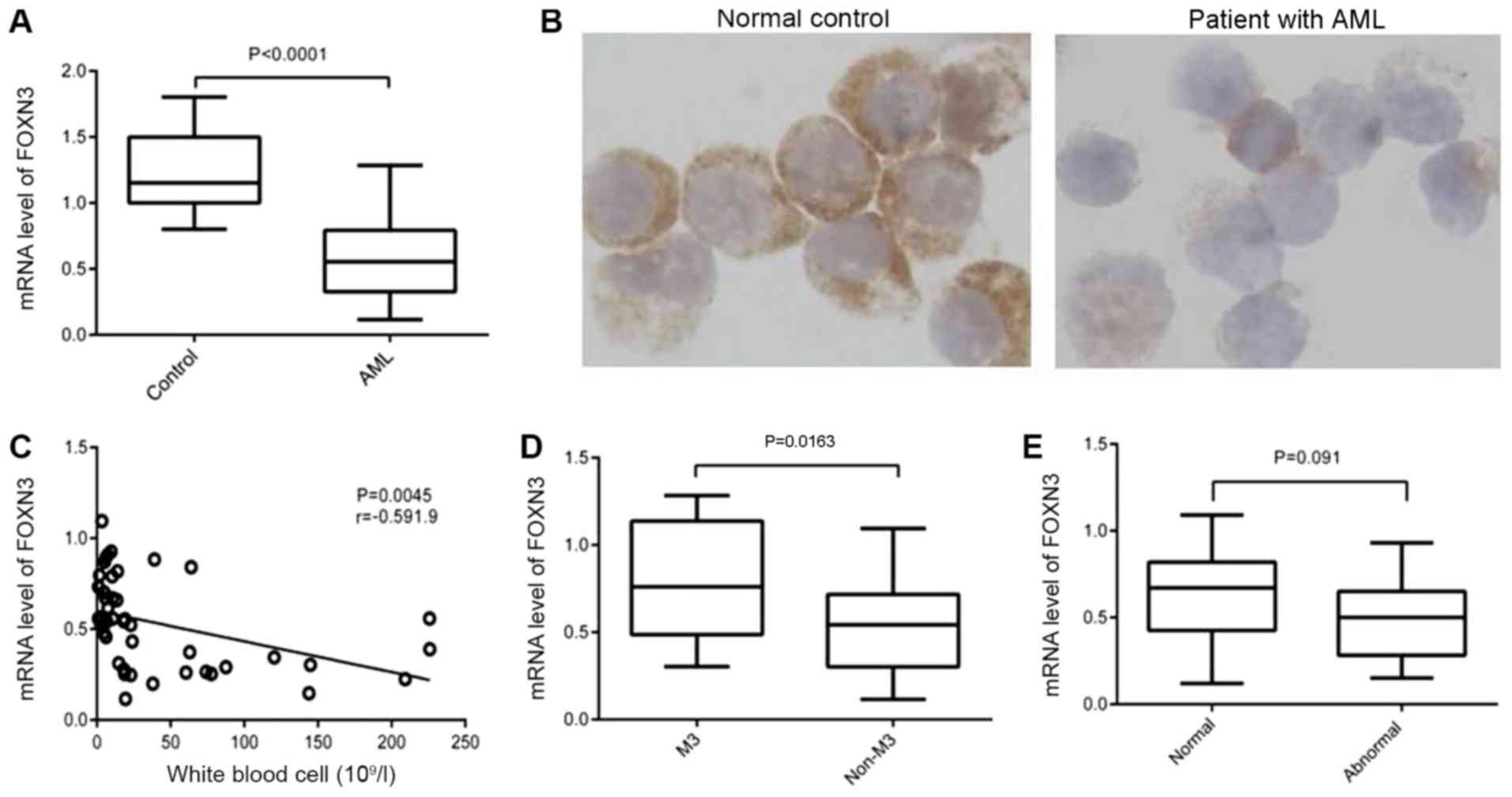
(0.5764±0.03774) compared with the controls (1.229±0.07865;
P<0.0001; Fig. 1A). Decreased
protein expression levels of FOXN3 were observed in six patients
with decreased FOXN3 mRNA expression levels (representative example
in Fig. 1B). The correlation between
FOXN3 mRNA expression levels and the clinicopathological features
of the 53 patients were assessed to further examine the
significance of decreased FOXN3 expression levels (Table III). FOXN3 expression levels were
significantly decreased in patients with WBC counts
>30×109/l, and the expression level of FOXN3 was
negatively correlated with the WBC count (r=−0.5919; P=0.0045;
Fig. 1C). However, FOXN3 expression
levels were not significantly decreased in patients with an
increased blast ratio in bone marrow (P=0.845; Table III). Although no significant
differences were observed in patients with different FAB subtypes
(P=0.146; Table III), FOXN3
expression levels in patients with FAB non-M3-subtype
(0.5387±0.03645) were significantly decreased compared with
patients with FAB M3-subtype (0.7884±0.3529; P=0.0163; Fig. 1D). Furthermore, no statistically
significant differences were identified in FOXN3 expression levels
between normal and abnormal karyotypes in non-M3 patients (P=0.091;
Fig. 1E). No significant
associations were identified between FOXN3 and other clinical
parameters, including age, sex, hemoglobin or platelet count.
| Table III.Association between FOXN3 expression
levels and the clinicopathological features of the patients. |
Table III.
Association between FOXN3 expression
levels and the clinicopathological features of the patients.
| Clinicopathological
characteristics | FOXN3
mRNAa | P-value |
|---|
| Sex |
| 0.078 |
|
Male | 0.5070±0.2212 |
|
|
Female | 0.6431±0.2748 |
|
| Age, years |
| 0.582 |
| Age
≤60 | 0.5998±0.2850 |
|
| Age
>60 | 0.5520±0.2671 |
|
| Peripheral blood
count |
| White
blood cell, 109/l |
| 0.005 |
|
<30 | 0.6859±0.1838 |
|
|
≥30 | 0.5151±0.2996 |
|
| Hemoglobin,
g/l |
| 0.669 |
|
≤80 | 0.5970±0.2911 |
|
|
>80 | 0.5549±0.2607 |
|
| Platelet,
109/l |
| 0.790 |
|
≤50 | 0.5689±0.2660 |
|
|
>50 | 0.5921±0.3003 |
|
| Bone marrow, % |
| 0.845 |
| Blast
≤70 | 0.5868±0.2773 |
|
| Blast
>70 | 0.5655±0.2772 |
|
| FAB subtype |
| 0.146 |
| M2 | 0.5209±0.2529 |
|
| M3 | 0.7884±0.3529 |
|
| M5 | 0.5687±0.2560 |
|
|
Other | 0.4642±0.1934 |
|
| Karyotype |
| 0.122 |
|
Normal | 0.6178±0.2644 |
|
|
t(15;17) | 0.7884±0.3528 |
|
|
t(8;21) | 0.6073±0.0743 |
|
|
Complex | 0.5132±0.2390 |
|
| Not
available | 0.4584±0.2261 |
|
Association of FOXN3 expression levels
with the survival of patients with AML
The association of FOXN3 expression levels with the
treatment outcome of AML was investigated in a cohort of patients
with the non-M3-subtype. A total of 45 patients were divided into a
sub-group with decreased FOXN3 expression levels (n=22) and a
sub-group with increased FOXN3 expression levels (n=23) according
to the median level of FOXN3 expression (0.505; data not shown).
The CR rate in the FOXN3 high-expression group (n=19) was 88%,
whereas, the CR rate in the low-expression group (n=17) was 77.2%.
No significant difference was observed in the CR rate between the
two groups (P=0.608; data not shown).
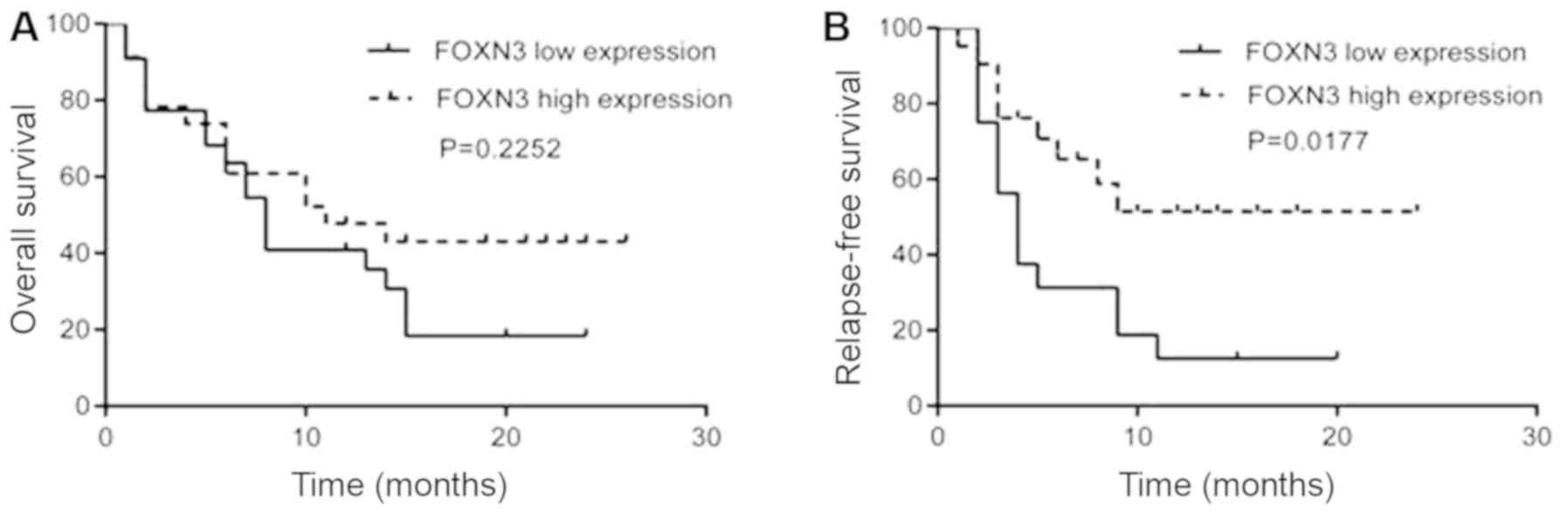
A 2-year follow-up was conducted on the 45 patients
with the non-M3-subtype to study the effect of FOXN3 expression
levels on the prognosis of patients with AML. The median survival
time for patients was 13 months. There was no significant
difference in the OS between patients with higher and lower FOXN3
expression (P=0.2252; Fig. 2A).
However, a significantly decreased 2-year RFS was observed in
patients with lower FOXN3 expression (P=0.0177; Fig. 2B).
AML cell lines exhibit decreased FOXN3
expression levels
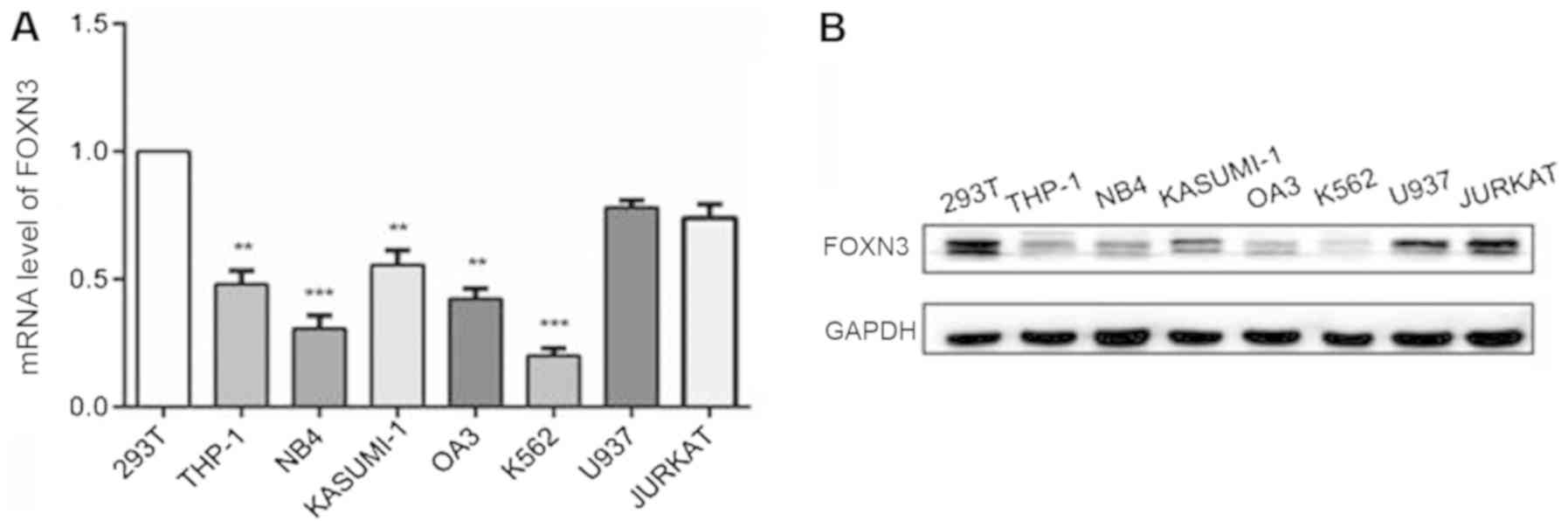
The mRNA and protein expression levels of FOXN3 in
leukemia cell lines, including THP-1, NB4, KASUMI-1, OA3, K562,
U937 and JURKAT, and the control 293T cell line were detected using
RT-qPCR and western blot analysis. Significantly decreased mRNA
expression levels of FOXN3 were observed in all the AML cell lines
tested, with the exception of the JURKAT and U937 cells, compared
with the control 293T cells (P<0.01; Fig. 3A). Correspondingly, decreased protein
expression levels of FOXN3 were observed in the AML cells compared
with the control 293T cells (Fig.
3B).
FOXN3 overexpression inhibits
proliferation and promotes apoptosis
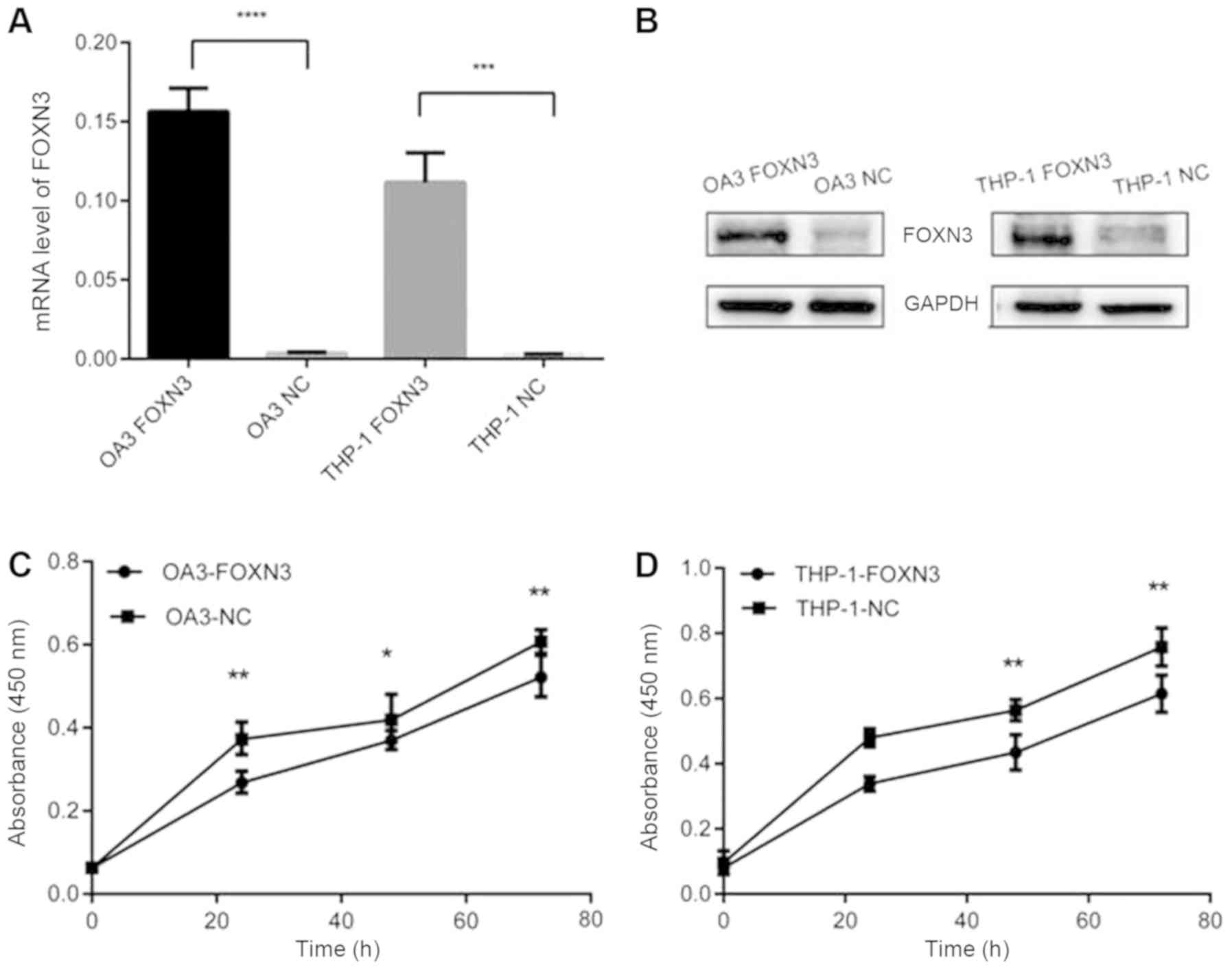
mRNA and protein expression levels of FOXN3 were
increased in the FOXN3-transfected OA3 and THP-1 cells compared
with the negative controls to study the role of FOXN3 in cell
growth (Fig. 4A and B). Cell
proliferation, cell cycle and apoptosis were measured by CCK-8 and
flow cytometry. FOXN3 overexpression significantly inhibited the
proliferation of leukemia cells after 48 and 72 h compared with the
control group (P<0.05; Fig. 4C and
D). In addition, the total apoptosis (including early and late
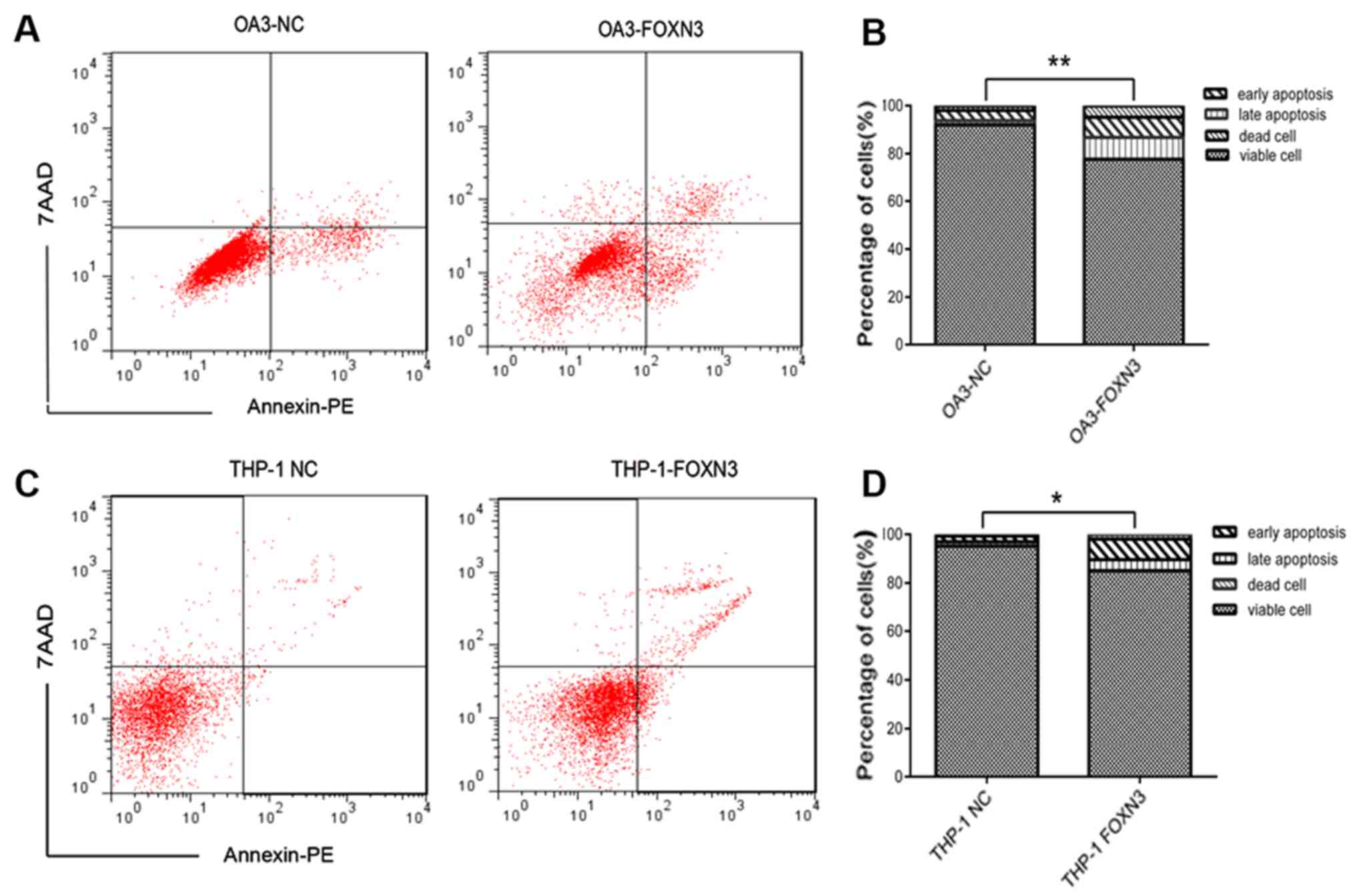
apoptosis) of leukemia cells increased following FOXN3
overexpression in OA3 cells (17.46±6.92 vs. 6.07±2.11%; P<0.01;
Fig. 5A and B) and THP-1 cells
(12.99±1.19 vs. 4.12±1.41%; P<0.05; Fig. 5C and D). The proportion of S-phase
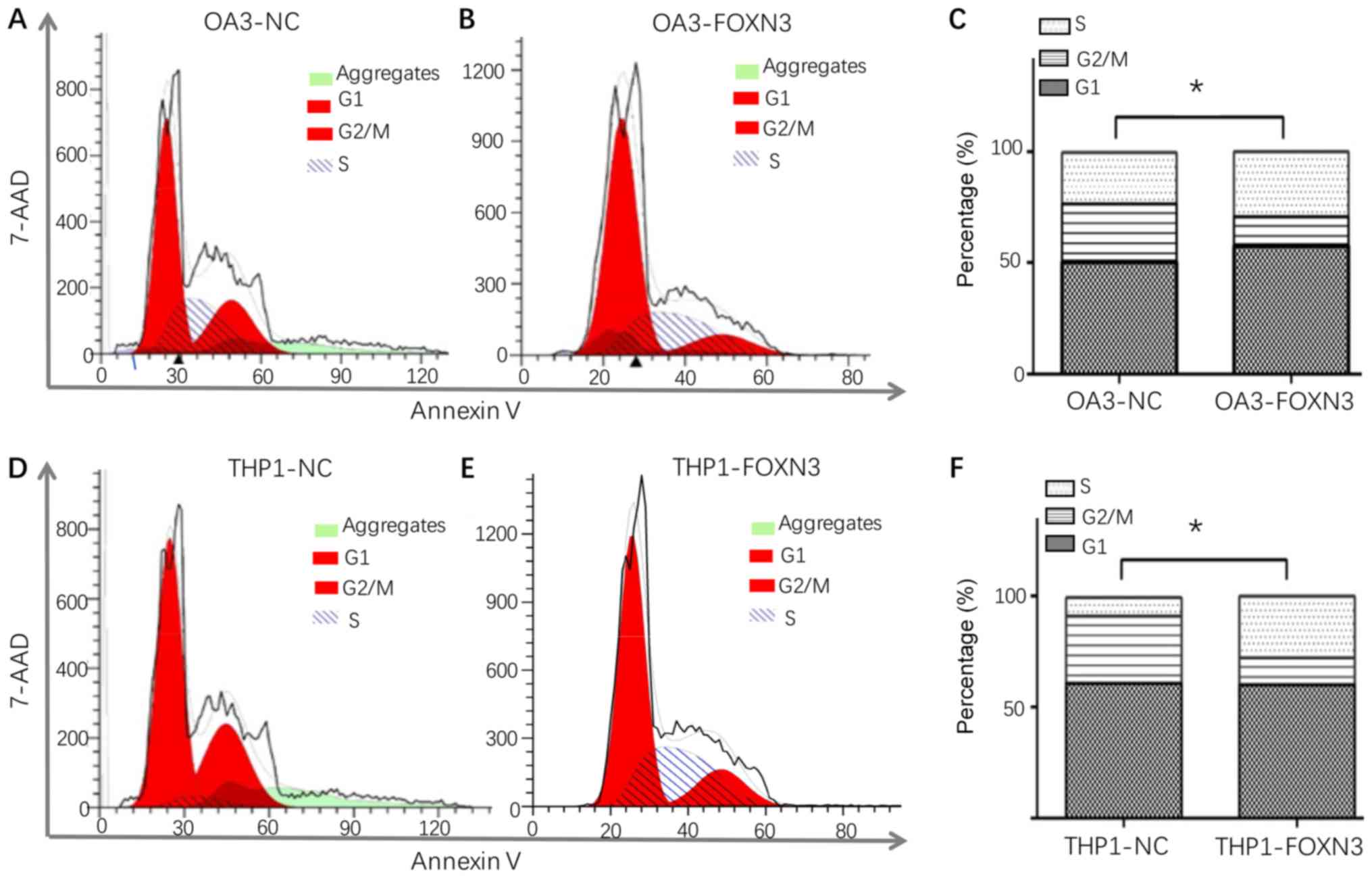
cells increased significantly following FOXN3 overexpression in OA3
cells (25.77±0.45 vs. 16.76±0.37%; P<0.05; Fig. 6A-C) and THP-1 cells (29.33±0.47 vs.
13.42±0.47%; P<0.05; Fig.
6D-F).
Discussion
A large number of FOX transcription factors were
demonstrated to serve important roles in tumorigenesis, including
forkhead box protein R1, forkhead box protein R2 and forkhead box
protein D1 (20–22). As a member of the FOXN subclass,
FOXN3 is abnormally expressed in a variety of malignancies. FOXN3
may be a direct target of microRNA-574-5p, as it was downregulated
by microRNA-574-5p to promote the progression of human lung cancer
(23). A recent study on colon
cancer demonstrated that the downregulation of FOXN3 activated the
β-catenin/Wnt signaling pathway, thereby promoting colon cancer
tumor cell growth, migration and invasion (13). In Hodgkin's lymphoma, FOXN3 may
activate the Janus kinase-signal transducer and activator of
transcription signaling pathways in transcriptional regulation
(14). In addition, FOXN3 expression
levels are decreased in a series of lymphomas, including primary
exudative lymphoma, diffuse large B-cell lymphoma and hairy cell
leukemia (14). The loss of FOXN3
led to an increase in the expression levels of the target gene
serine/threonine-protein kinase Pim-2 (PIM2), thereby decreasing
the effect of FOXN3 on tumor growth (24). However, the function of FOXN3 in AML
has not yet been fully elucidated.
In a previous study, high-resolution aCGH technology
identified the presence of a FOXN3 gene deletion on chromosome 14
in two patients classified with M5-subtype AML (3), thus, the present study aimed to examine
the potential role of FOXN3 in AML. M3 is a subtype of AML with
aberrant t(15;17) chromosome, serving as a treatment target. It is
possible to cure the AML subtype using retinoic acid and arsenic
containing compounds (25). At
present, 95% of patients with M3 may achieve complete remission and
long-term survival. The classification of AML as a M3 or
non-M3-subtype determines the treatment regimen. FOXN3 expression
levels were significantly decreased in patients with AML,
particularly in patients classified with non-M3-subtype AML.
Furthermore, the downregulation of FOXN3 was significantly
correlated with the peripheral WBC count in patients, which is a
high-risk prognostic marker of AML (26). This suggested that FOXN3 may be
involved in the proliferation of leukemia cells. It was
subsequently demonstrated that FOXN3 overexpression inhibited the
malignant transformation of AML cells by decreasing cell
proliferation, promoting apoptosis and enhancing cell cycle arrest.
The prognostic significance of FOXN3 requires further
investigation, as the downregulation of FOXN3 was associated with a
decreased RFS, with no significant effect on OS. A possible
explanation may be the regulatory effect of FOXN3 on cell
proliferation, cell apoptosis and the cell cycle, suggesting the
role of FOXN3 as a tumor suppressor in AML. Unlike RFS, the OS of
AML is impacted by numerous factors, in addition to the relapse of
leukemia. One possible explanation for the discrepancy was the
association between lower FOXN3 expression levels and an increased
WBC count, as the latter is considered a high-risk marker for
relapse of leukemia (26).
However, the cellular signaling pathway regulated by
FOXN3 in AML remains unknown. Scott and Plon (27) demonstrated that FOXN3 protein
negatively regulated transcription through the COOH end of
Ski-interacting protein (SKIP). SKIP inhibited tumor formation by
competing with Ski oncogene to activate the transforming growth
factor-β (TGF-β)/mothers against decapentaplegic homolog (Smad)
signaling pathway (28). Previous
studies suggested that the TGF-β/Smad cell signaling pathway may
regulate the proliferation and differentiation of hematopoietic
stem cells, and is closely associated with the occurrence of
leukemia (29–31). However, whether FOXN3 mediates
hematopoiesis through the TGF-β/Smad signaling pathway has not yet
been determined (32,33). It is hypothesized that the
downregulation of FOXN3 may lead to the activation of the
TGF-β/Smad signaling pathway and promote cell proliferation. This
may result in hematopoietic stem cell proliferation and
differentiation imbalances, which may induce leukemia. In addition,
as a transcription inhibitor, FOXN3 binds directly to the promoter
of PIM2 and decreases PIM2 expression (24), which is an important target of
fms-like tyrosine kinase-3 (FLT3)-internal tandem duplication (ITD)
induced transformation (34). PIM2
may complement the FLT3/wild-type (WT) signaling pathway and induce
transformation similar to FLT3-ITD in myeloid cells (35). Furthermore, PIM2 and protein
biosynthesis are principal targets of the antiproliferative effects
of FOXN3 (24). Therefore, the
downregulation of FOXN3 may increase PIM2 expression levels and
promote leukemia transformation by activating FLT3/WT
signaling.
The sample size was relatively small in the present
study. Therefore, more patients are required for future studies to
identify the effect of FOXN3 downregulation on the progression of
AML. Furthermore, the use of in vivo experimental models is
necessary to verify the tumor suppressive role of FOXN3 in AML. As
a normal monocyte or granulocyte cell line was unavailable for the
present study, the use of 293T cells as a control cell line was
additionally a limitation.
In summary, FOXN3 was downregulated in AML. It may
be a biomarker of high-risk AML, as the expression levels of FOXN3
were negatively correlated with peripheral WBC count and negatively
associated with RFS in patients with AML. The contribution of FOXN3
to leukemogenesis may be through its regulatory effect on cell
proliferation, apoptosis and the cell cycle. Whether FOXN3 affects
any cellular signaling pathways, including the TGF-β/Smad and
FLT3/WT pathways, in AML requires further study. The results of the
present study suggested that FOXN3 may be a novel therapeutic
target as FOXN3 may be implicated in multiple signaling pathways
associated with AML.
Acknowledgements
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of China (grant no. 81600117).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the author on reasonable request.
Authors' contributions
HH collected the bone marrow samples of the
patients, and analyzed and interpreted the data. JZ conducted the
immunohistochemical study and follow-up of the patients. YQ
conducted the cell experiments. YW performed the polymerase chain
reaction assays. YZ was involved in the cell experiments. XY
contributed to the data analysis. YL designed the research. RZ
designed the study and was a major contributor in writing the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
The present study was approved by The Ethics
Committee of The First Affiliated Hospital of China Medical
University (Shenyang, China). Prior to data collection, written
informed consent for participation in the present study was
obtained from each participant.
Patient consent for publication
Written informed consent for publishing the present
study was obtained from each participant.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Smith M, Barnett M, Bassan R, Gatta G,
Tondini C and Kern W: Adult acute myeloid leukaemia. Crit Rev Oncol
Hematol. 50:197–222. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
Statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang R, Lee JY, Wang X, Xu W, Hu X, Lu X,
Niu Y, Tang R, Li S and Li Y: Identification of novel genomic
aberrations in AML-M5 in a level of array CGH. PLoS One.
9:e876372014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pati D, Keller C, Groudine M and Plon SE:
Reconstitution of a MEC1-independent checkpoint in yeast by
expression of a novel human fork head cDNA. Mol Cell Biol.
17:3037–3046. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ostrow AZ, Kalhor R, Gan Y, Villwock SK,
Linke C, Barberis M, Chen L and Aparicio OM: Conserved forkhead
dimerization motif controls DNA replication timing and spatial
organization of chromosomes in S. cerevisiae. Proc Natl Acad Sci
USA. 114:E2411–E2419. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kaestner KH, Knochel W and Martinez DE:
Unified nomenclature for the winged helix/forkhead transcription
factors. Genes Dev. 14:142–146. 2000.PubMed/NCBI
|
|
7
|
Schmidt J, Piekarski N and Olsson L:
Cranial muscles in amphibians: Development, novelties and the role
of cranial neural crest cells. J Anat. 222:134–146. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schmidt J, Schuff M and Olsson L: A role
for FoxN3 in the development of cranial cartilages and muscles in
Xenopus laevis (Amphibia: Anura: Pipidae) with special emphasis on
the novel rostral cartilages. J Anat. 218:226–242. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Robert T, Vanoli F, Chiolo I, Shubassi G,
Bernstein KA, Rothstein R, Botrugno OA, Parazzoli D, Oldani A,
Minucci S, et al: HDACs link the DNA damage response, processing of
double-strand breaks and autophagy. Nature. 471:74–79. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Cirillo LA and Zaret KS: Specific
interactions of the wing domains of FOXA1 transcription factor with
DNA. J Mol Biol. 366:720–724. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Busygina V, Kottemann MC, Scott KL, Plon
SE and Bale AE: Multiple endocrine neoplasia type 1 interacts with
forkhead transcription factor CHES1 in DNA damage response. Cancer
Res. 66:8397–8403. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sun J, Li H, Huo Q, Cui M, Ge C, Zhao F,
Tian H, Chen T, Yao M and Li J: The transcription factor FOXN3
inhibits cell proliferation by downregulating E2F5 expression in
hepatocellular carcinoma cells. Oncotarget. 7:43534–43545.
2016.PubMed/NCBI
|
|
13
|
Dai Y, Wang M, Wu H, Xiao M, Liu H and
Zhang D: Loss of FOXN3 in colon cancer activates beta-catenin/TCF
signaling and promotes the growth and migration of cancer cells.
Oncotarget. 8:9783–9793. 2017.PubMed/NCBI
|
|
14
|
Nagel S, Meyer C, Kaufmann M, Drexler HG
and MacLeod RA: Deregulated FOX genes in Hodgkin lymphoma. Genes
Chromosomes Cancer. 53:917–933. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mudduluru G, Abba M, Batliner J, Patil N,
Scharp M, Lunavat TR, Leupold JH, Oleksiuk O, Juraeva D, Thiele W,
et al: A Systematic Approach to Defining the microRNA Landscape in
Metastasis. Cancer Res. 75:3010–3019. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2-ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hsu SM, Raine L and Fanger H: Use of
avidin-biotin-peroxidase complex (ABC) in immunoperoxidase
techniques: A comparison between ABC and unlabeled antibody (PAP)
procedures. J Histochem Cytochem. 29:577–580. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhu Y, Dai B, Zhang H, Shi G, Shen Y and
Ye D: Long non-coding RNA LOC572558 inhibits bladder cancer cell
proliferation and tumor growth by regulating the AKT-MDM2-p53
signaling axis. Cancer Lett. 380:369–374. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhu Z, Yang C, Wang J, Feng Q, Chen Q and
Yang P: Altered chemokine receptor expression in the peripheral
blood lymphocytes in polymyositis and dermatomyositis. Cytokine.
99:316–321. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pan F, Li M and Chen W: FOXD1 predicts
prognosis of colorectal cancer patients and promotes colorectal
cancer progression via the ERK 1/2 pathway. Am J Transl Res.
10:1522–1530. 2018.PubMed/NCBI
|
|
21
|
Katoh M, Igarashi M, Fukuda H, Nakagama H
and Katoh M: Cancer genetics and genomics of human FOX family
genes. Cancer Lett. 328:198–206. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang X, He B, Gao Y and Li Y: FOXR2
contributes to cell proliferation and malignancy in human
hepatocellular carcinoma. Tumour Biol. 37:10459–10467. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li Q, Li X, Guo Z, Xu F, Xia J, Liu Z and
Ren T: MicroRNA-574-5p was pivotal for TLR9 signaling enhanced
tumor progression via down-regulating checkpoint suppressor 1 in
human lung cancer. PLoS One. 7:e482782012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Huot G, Vernier M, Bourdeau V, Doucet L,
Saint-Germain E, Gaumont-Leclerc MF, Moro A and Ferbeyre G:
CHES1/FOXN3 regulates cell proliferation by repressing PIM2 and
protein biosynthesis. Mol Biol Cell. 25:554–565. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lo-Coco F, Avvisati G, Vignetti M, Thiede
C, Orlando SM, Iacobelli S, Ferrara F, Fazi P, Cicconi L, Di Bona
E, et al Gruppo Italiano Malattie Ematologiche dell'Adulto;
German-Austrian Acute Myeloid Leukemia Study Group; Study Alliance
Leukemia, : Retinoic acid and arsenic trioxide for acute
promyelocytic leukemia. N Engl J Med. 369:111–121. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mamez AC, Raffoux E, Chevret S, Lemiale V,
Boissel N, Canet E, Schlemmer B, Dombret H, Azoulay E and Lengliné
E: Pre-treatment with oral hydroxyurea prior to intensive
chemotherapy improves early survival of patients with high
hyperleukocytosis in acute myeloid leukemia. Leuk Lymphoma.
57:2281–2288. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Scott KL and Plon SE: CHES1/FOXN3
interacts with Ski-interacting protein and acts as a
transcriptional repressor. Gene. 359:119–126. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Leong GM, Subramaniam N, Figueroa J,
Flanagan JL, Hayman MJ, Eisman JA and Kouzmenko AP: Ski-interacting
protein interacts with Smad proteins to augment transforming growth
factor-beta-dependent transcription. J Biol Chem. 276:18243–18248.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tabe Y, Shi YX, Zeng Z, Jin L, Shikami M,
Hatanaka Y, Miida T, Hsu FJ, Andreeff M and Konopleva M:
TGF-β-Neutralizing Antibody 1D11 Enhances Cytarabine-Induced
Apoptosis in AML Cells in the Bone Marrow Microenvironment. PLoS
One. 8:e627852013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Su E, Han X and Jiang G: The transforming
growth factor beta 1/SMAD signaling pathway involved in human
chronic myeloid leukemia. Tumori. 96:659–666. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kim SJ and Letterio J: Transforming growth
factor-beta signaling in normal and malignant hematopoiesis.
Leukemia. 17:1731–1737. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhou S, Fujimuro M, Hsieh JJ, Chen L,
Miyamoto A, Weinmaster G and Hayward SD: SKIP, a CBF1-associated
protein, interacts with the ankyrin repeat domain of NotchIC To
facilitate NotchIC function. Mol Cell Biol. 20:2400–2410. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vaidya A and Kale VP: TGF-β signaling and
its role in the regulation of hematopoietic stem cells. Syst Synth
Biol. 9:1–10. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Hospital MA, Jacquel A, Mazed F, Saland E,
Larrue C, Mondesir J, Birsen R, Green AS, Lambert M, Sujobert P, et
al: RSK2 is a new Pim2 target with pro-survival functions in
FLT3-ITD-positive acute myeloid leukemia. Leukemia. 32:597–605.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Agrawal S, Koschmieder S, Bäumer N, Reddy
NG, Berdel WE, Müller-Tidow C and Serve H: Pim2 complements Flt3
wild-type receptor in hematopoietic progenitor cell transformation.
Leukemia. 22:78–86. 2008. View Article : Google Scholar : PubMed/NCBI
|