Introduction
Breast cancer is one of the most prevalent types of
malignant tumors in females (1).
Significant advances in the treatment for breast cancer have been
achieved (1,2). However, breast cancer remains one of
the most common causes of cancer-associated mortality in females,
accounting for ~14.3% of cancer-associated mortality worldwide in
2012 (2). The majority of
mortalities due to breast cancer can be attributed to metastasis
(2). Metastases occur when tumor
cells disseminate from the primary tumor site to the surrounding
extracellular matrix, travel through the vasculature, and finally
extravasate at a distant site to form a secondary tumor (3,4). In
total, ~10-15% of patients diagnosed with breast cancer present
with distant metastasis within 3 years (4). Therefore, identification of new
molecular targets for the effective treatment of breast cancer is
important.
MicroRNAs (miRNAs) are a class of small non-coding
RNAs of ~20 nucleotides in length that can regulate gene expression
by directly targeting mRNAs for degradation (5,6). miRNAs
regulate a wide range of physiological activities in the cell,
including cell proliferation, metabolism, apoptosis, invasion and
migration (7). Dysregulated miRNAs
are also involved in the initiation and progression of certain
types of cancer, including breast cancer (8,9). miR-616
may function as an oncogene in different types of cancer, including
gastric cancer, glioma, non-small cell lung cancer, hepatocellular
carcinoma and prostate cancer (10–15).
However, the expression, function and molecular mechanism of
miR-616 in breast cancer remain unclear.
Tissue inhibitor of metalloproteinases 2 (TIMP2) is
a critical inhibitor of matrix metalloproteinases (MMPs) and has
been largely studied in various types of human tumor. In addition,
it was demonstrated that TIMP2 is associated with the invasive and
metastatic abilities of various types of tumor cell including
breast cancer cells (16). Previous
studies reported that certain miRNAs, including miR-130a and
miR-552, regulate TIMP2 expression (17,18);
however, the association between TIMP2 and miRNAs in breast cancer
remains unknown.
The results from the present study demonstrated that
miR-616 was upregulated in breast cancer tissues and cell lines.
In vitro functional assays indicated that miR-616 could
promote breast cancer cell proliferation and metastasis. In
addition, this study demonstrated that TIMP2 could be a direct
target of miR-616. These results may help understanding the
underlying mechanism of miR-616 in breast cancer.
Materials and methods
Tissue collection
The present study was approved by the Medical Ethics
Committee of Dezhou No. 2 People's Hospital (Dezhou, China). All
patients included in the current study provided written informed
consent. In total, 30 paired breast tumor tissue and non-tumor
breast tissue samples (>5 cm distant from tumor tissue) were
obtained from 30 female patients (age range, 35–77 years; mean age,
62 years) who underwent surgical resection at Dezhou No. 2 People's
Hospital (Dezhou, China) between January 2016 and July 2017.
Patients who underwent chemotherapy or radiotherapy prior to
surgery were excluded from the study.
Cell culture and transfection
The breast cancer cell lines MDA-MB-231, MCF-7,
BT474 and MDA-MB-468, the immortal mammary epithelial cell line
MCF-10A and the 293 cell line were obtained from the Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China).
All cell lines were incubated in a 95% humidified chamber with 5%
CO2 at 37°C. MCF-7, BT474, MDA-MB-231, MDA-MB-468 and
293 cell lines were maintained in Dulbecco's modified Eagle's
medium (DMEM; Gibco; Thermo Fisher Scientific, Inc.) supplemented
with 10% fetal bovine serum (FBS; GE Healthcare Life Sciences).
MCF-10A cells were maintained in DMEM/F12 medium (GE Healthcare
Life Sciences) containing 5% horse serum (Gibco; Thermo Fisher
Scientific, Inc.) and 1% penicillin/streptomycin (GE Healthcare
Life Sciences). The miR-616 mimic, negative control (miR-NC),
inhibitor and inhibitor negative control (anti-NC) were purchased
from Guangzhou RiboBio, Co., Ltd. The sequences were as follows:
miR-616 mimics, 5′-AGUCAUUGGAGGGUUUGAGCAG-3′; miR-NC,
5′-ACUACUGAGUGACAGUAGA-3′; miR-616 inhibitor,
5′-GAGUAUCCCGUUGCCAACGAGA-3′; and anti-NC,
5′-UUCUCCGAACGUGUCACGUTT-3′. Small interfering RNA (siRNA)
targeting human TIMP2 (siTIMP2) and si-control were obtained from
Santa Cruz Biotechnology, Inc. The sequences were as follows:
siTIMP2, 5′-CTCTGATTTGGTCGTATTGGG-3′ and si-control,
5′-CAGUACUUUUGUGUAGUACAA-3′. MCF-7 and MDA-MB-231 cells (5,000)
were seeded into 6-well plates and incubated at 37°C until they
reached 70–80% confluence. A total of 50 nM miR-616 mimics or
miR-616 inhibitors or/and 50 nM siRNA were transfected into MCF-7
and MDA-MB-231 cells using Lipofectamine™ 2000 (Invitrogen; Thermo
Fisher Scientific, Inc.) according to the manufacturer's protocols.
Following 24 h transfection, cells were collected for subsequent
experiments.
Cell Counting Kit-8 (CCK-8) assay
Cell viability was analyzed using a CCK-8 kit,
according to the manufacturer's protocols (Beyotime Institute of
Biotechnology). Briefly, MDA-MB-231 or MCF-7 cells were plated in
96-well plates at a density of 1,000 cells/well at 24 h
post-transfection. Following incubation for 0, 24, 48 and 72 h at
37°C, 10 µl CCK-8 reagent was added to each well. The cells were
incubated at 37°C in an atmosphere containing 5% CO2 for
2 h. Absorbance was determined at a wavelength of 450 nm using an
ELx808 absorbance reader (BioTek Instruments, Inc.).
Colony formation assay
For the assessment of colony formation, transfected
breast cancer cells were seeded in 6-well plates at a density of
500 cells/well in triplicate and incubated for 1 week at 37°C.
Subsequently, the plates were washed with PBS and stained with 0.5%
crystal violet at room temperature for 20 min. After washing three
times, colonies with >50 cells were counted and analyzed under a
light microscope (magnification, ×100).
Migration and invasion assays
The ability of migration and invasion was assessed
using Transwell chambers (Corning Inc.). Cells were re-suspended in
serum-free DMEM at a concentration of 1×105/ml and then
200 µl cell suspension was seeded onto the upper well of 8-µm pore
Transwell inserts with or without Matrigel (Sigma Aldrich; Merck
KGaA). Matrigel was only used for invasion assays. DMEM containing
10% FBS was added to the lower chamber. After 24 h incubation,
cells in the upper chambers were removed with a cotton swab. The
migrated and invaded cells were then stained with 0.1% crystal
violet for 20 min at room temperature. Images from five different
fields were captured and counted under a light microscope
(magnification, ×100).
Western blot analysis
Cells were lysed in cold radioimmunoprecipitation
assay buffer (Thermo Fisher Scientific, Inc.) and the protein
concentration was determined using a bicinchoninic acid protein
assay kit (Pierce; Thermo Fisher Scientific, Inc.). Proteins (30
µg) were separated by 10% SDS-PAGE and then transferred to a
polyvinylidene difluoride (PVDF) membrane (Thermo Fisher
Scientific, Inc.) The PVDF membrane was blocked with 5% non-fat
milk in PBS containing 0.1% Tween-20 (Sigma-Aldrich; Merck KGaA) at
room temperature for 3 h. Subsequently, the PVDF membrane was
incubated with TIMP2 antibody (cat. no. sc-21735; 1:200; Santa Cruz
Biotechnology, Inc.), MMP2 antibody (cat. no. sc-13594; 1:200;
Santa Cruz Biotechnology, Inc.), MMP9 antibody (cat. no. sc-21733;
1:200; Santa Cruz Biotechnology, Inc.) and GAPDH antibody (cat. no.
AF0006; 1:500; Beyotime Institute of Biotechnology) at room
temperature for 3 h. Following washing with PBS for 10 min, the
PVDF membrane was incubated with a goat anti-mouse secondary
antibody (cat. no. ab64255; 1:1,000; Abcam) at room temperature for
1 h. Following further washing with PBS for 10 min, the protein
bands were detected using an Enhanced Chemiluminescence Western
Blotting kit (Pierce; Thermo Fisher Scientific, Inc.), according to
the manufacturer's protocols.
Reverse transcription-quantitative PCR
(RT-qPCR)
Total RNA from the breast cancer cell lines was
extracted using TRIzol® reagent (Thermo Fisher
Scientific, Inc.) and 1 µg total RNA was reverse transcribed using
the Reverse Transcription System kit (Promega Corporation). qPCR
was performed with a Power SYBR-Green PCR Master mix (Thermo Fisher
Scientific, Inc.) with human GAPDH used as an internal control. For
miRNA analysis, qPCR was performed using an all-in-one miRNA
RT-qPCR Detection kit (GeneCopoeia, Inc.) and U6 small nuclear RNA
as an endogenous control. For the miRNA and mRNA amplifications,
the PCR thermocycling conditions were as follows: One cycle at 95°C
for 3 min, followed by 40 cycles at 95°C for 12 sec and 62°C for 35
sec, and a final extension step at 95°C for 15 sec. The relative
expression levels were calculated using the 2−ΔΔCq
method (19). The primer sequences
used for amplification were as follows: miR-616, forward,
5′-ACACTCCAGCTGGGAGTCATTGGAGGGTTT-3′, reverse,
5′-TGGTGTCGTGGAGTCG′3′; TIMP2, forward,
5′-CTCTGATTTGGTCGTATTGGG-3′, reverse, 5′-TGGAAGATGGTGATGGGATT-3′;
MMP2, forward, 5′-AAGTCTGAAGAGCGTGAAGTTTGGA-3′, reverse,
5′-TGAGGGTTGGTGGGATTGGAG-3′; MMP9, forward,
5′-AGTCCACCCTTGTGCTCTTCCC-3′, reverse,
5′-TCTGCCACCCGAGTGTAACCAT-3′; U6, forward, 5′-CTCGCTTCGGCAGCACA-3′,
reverse, 5′-AACGCTTCACGAATTTGCGT-3′; and GAPDH, forward,
5′-CGGAGTCAACGGATTTGGTCGTAT-3′, reverse,
5′-AGCCTTCTCCATGGTGGTGAAGAC-3′.
Dual-luciferase reporter assay
The TIMP2 3′-untranslated region (3′-UTR),
containing target sequences complementary to the miR-616 seed
sequence, was cloned downstream of the firefly luciferase gene in
the psiCHECK-2™ luciferase vector (Promega Corporation). Mutated
TIMP2 3′-UTR sequences were cloned into the same luciferase vector
(TIMP2-mut). The indicated reporter constructs and miR-616 mimic or
inhibitor were co-transfected with the phRGTK Renilla
luciferase internal control plasmid (Promega Corporation) into 293
cell line using Lipofectamine™ 2000 (Invitrogen; Thermo Fisher
Scientific, Inc.). The luciferase activity was analyzed following
24 h transfection using a dual-luciferase reporter assay system
(Promega Corporation), according to the manufacturer's
protocols.
Bioinformatics analysis
The prediction of TIMP2 3′-UTR as a miR-616 binding
target was determined using TargetScan 7.1 software (www.targetscan.org).
Statistical analysis
All experiments were performed three times and data
were analyzed using GraphPad Prism 5 (GraphPad Software Inc.).
Differences between two groups were assessed using a two-tailed
Student's t-test. Data of >2 groups were analyzed using one-way
analysis of variance with a post hoc Tukey's test. The correlation
between miR-616 levels and TIMP2 mRNA expression levels in human
breast cancer tissues was determined using Spearman's rank
correlation coefficient. P<0.05 was considered to indicate a
statistically significant difference.
Results
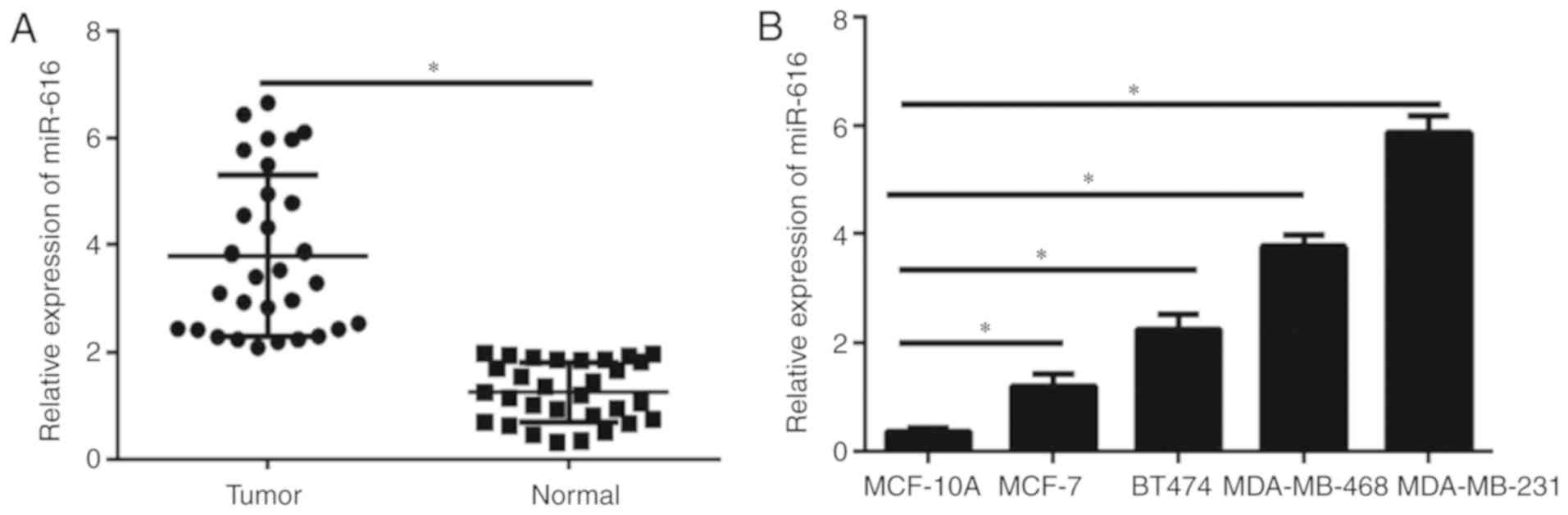
miR-616 expression is upregulated in
breast cancer tissues and cell lines
The expression levels of miR-616 in breast cancer
tissues and cell lines were detected by RT-qPCR. The results
demonstrated that miR-616 levels were significantly upregulated in
breast cancer samples compared with normal tissues (P<0.05;
Fig. 1A). Furthermore, the miR-616
levels were evaluated in the breast cancer cell lines MCF-7, BT474,
MDA-MB-231 and MDA-MB-468, and the immortal mammary epithelial cell
line MCF-10A. The miR-616 expression levels were significantly
increased in all breast cancer cell lines compared with MCF-10A
cells (P<0.05; Fig. 1B), although
the difference varied across the cell lines. These data indicated
that miR-616 may serve an important role in the progression of
breast cancer. The expression levels of miR-616 was the highest in
MDA-MB-231 cells and the lowest in MCF-7 cells; therefore, these
two cell lines were selected for further experiments.
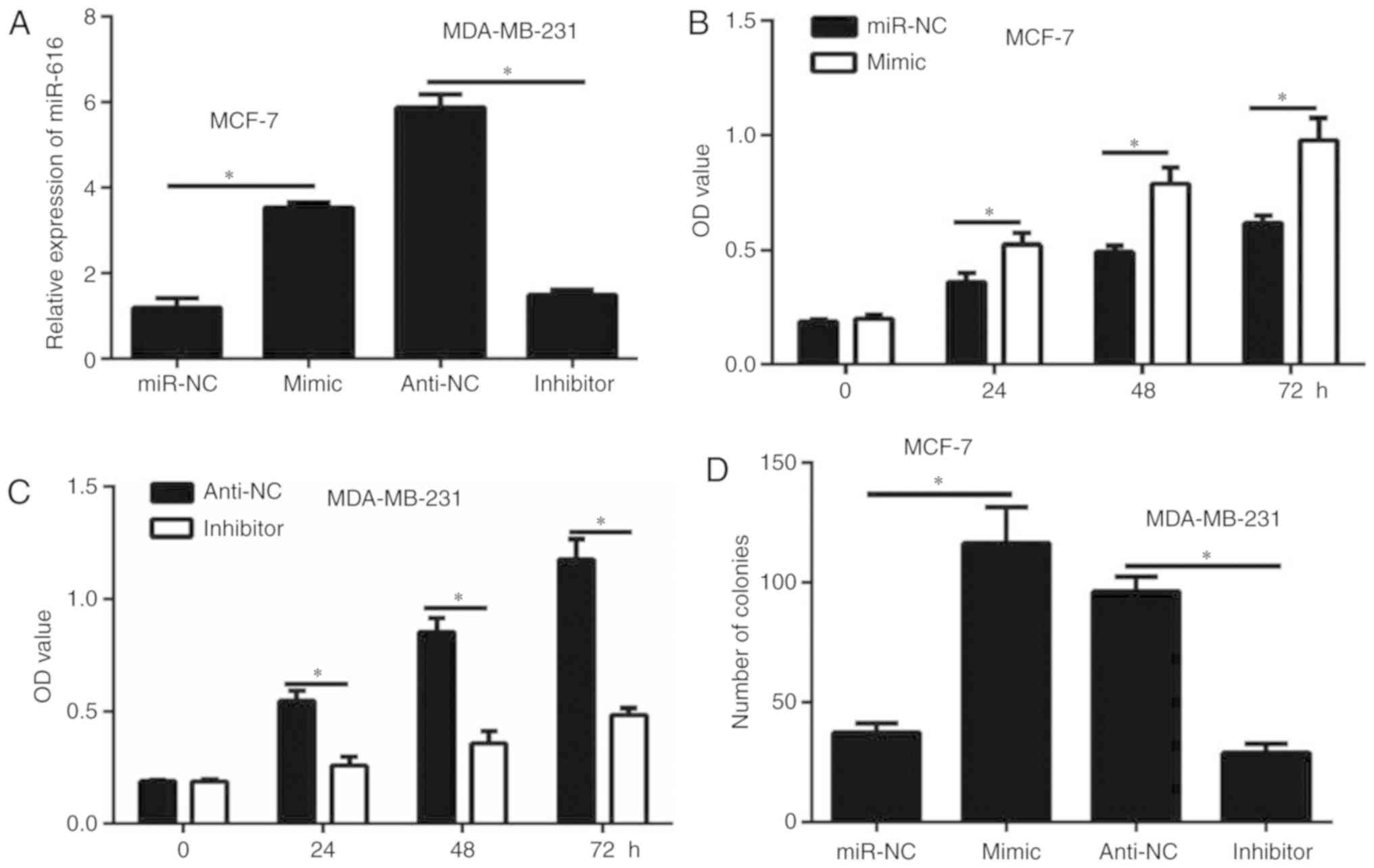
miR-616 enhances cell growth in breast
cancer cells
Overexpression of miR-616 was achieved by
transfection of MCF-7 cells with miR-616 mimic. By contrast,
miR-616 silencing was achieved by transfection of MDA-MB-231 cells
with miR-616 inhibitor. Transfection efficiency for both the
overexpression and the silencing experiments was confirmed by
RT-qPCR (Fig. 2A). The results of
CCK-8 assay demonstrated that miR-616 overexpression significantly
enhanced the proliferation of MCF-7 cells (P<0.05; Fig. 2B). By contrast, miR-616 silencing
significantly inhibited the proliferation of MDA-MB-231 cells
(P<0.05; Fig. 2C). Similar
results were observed with a colony formation assay (P<0.05;
Fig. 2D). These data suggested that
miR-616 enhanced breast cancer cell growth in vitro.
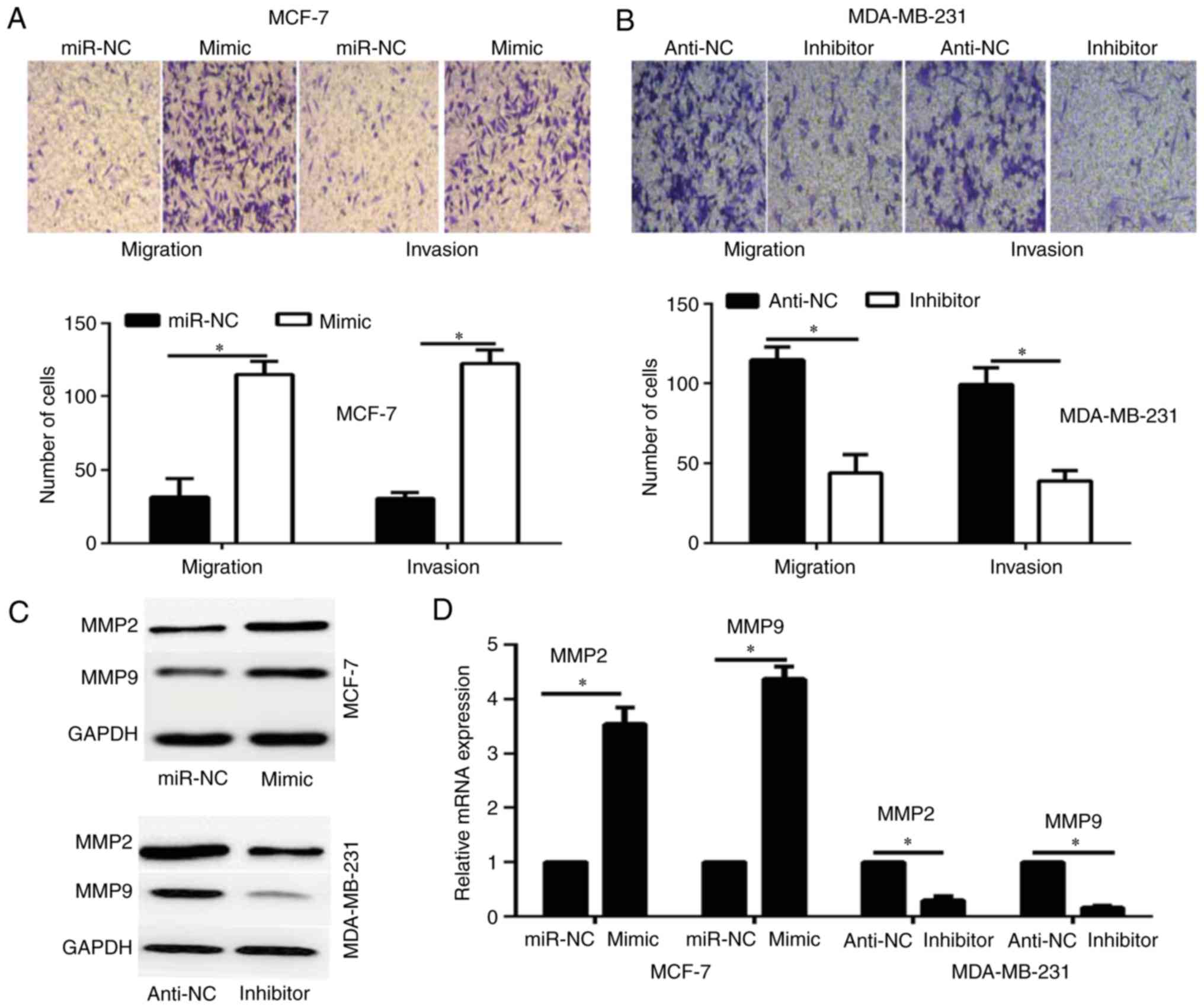
miR-616 enhances breast cancer cell
invasion in vitro
To investigate whether the overexpression of miR-616
affects the migration and invasion of breast cancer cells,
Transwell assays were performed. The results revealed that
overexpression of miR-616 significantly enhanced the migration and
invasion potential of MCF-7 cells compared with the miR-NC group,
whereas miR-616 silencing significantly inhibited the migration and
invasion capacities of MDA-MB-231 cells compared with the anti-NC
group (P<0.05; Fig. 3A and
B).
A previous study has reported that MMP2 and MMP9
serve a critical role in the migration and invasion of breast
cancer cells (20). Therefore, the
current study investigated whether miR-616 affects the expression
of MMP2 and MMP9. The results of RT-qPCR and western blot analysis
demonstrated that overexpression of miR-616 enhanced the expression
levels of MMP2 and MMP9, while miR-616 silencing significantly
reduced their expression levels (P<0.05; Fig. 3C and D).
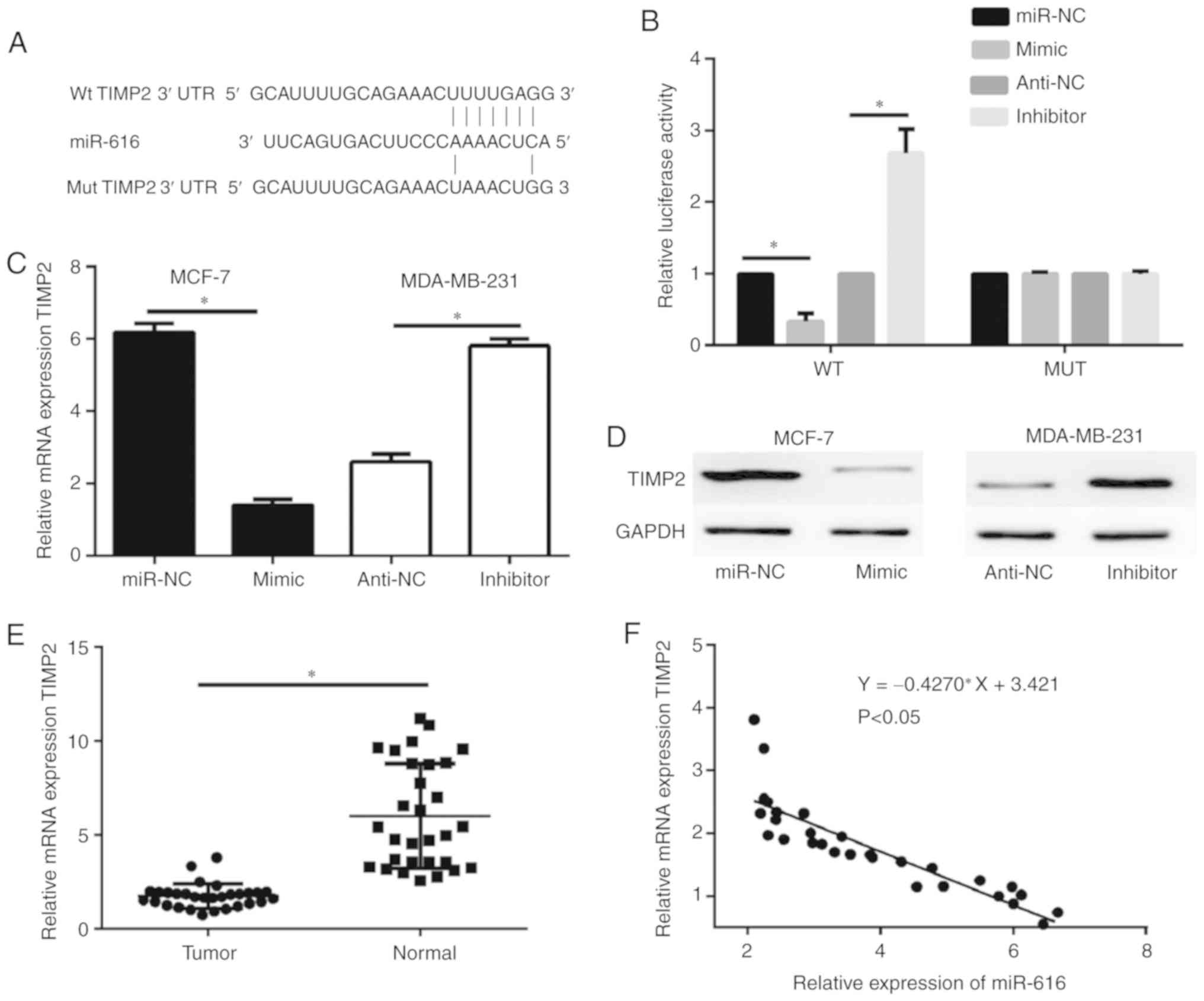
TIMP2 is a direct target of
miR-616
The potential targets of miR-616 were predicted by
bioinformatics analysis. Previous studies have demonstrated that
TIMP2 acts as a tumor suppressor and downregulates the expression
of MMP2 and MMP9 in tumor cells (20,21);
therefore, out of the predicted potential targets for miR-616,
TIMP2 was selected for further analysis in the current study. A
schematic of the target sequence in the 3′UTR of TIMP2 is shown in
Fig. 4A. Dual-luciferase reporter
assays were performed to evaluate whether miR-616 directly targets
TIMP2. The results revealed that miR-616 mimic significantly
decreased the luciferase activity of the wild-type TIMP2 and
miR-616 silencing significantly increased the luciferase activity
of the wild-type TIMP2; however, no significant difference was
observed in the luciferase activity of the mutant TIMP2 (P<0.05;
Fig. 4B). Furthermore, RT-qPCR and
western blot analysis demonstrated that the mRNA and protein
expression levels of TIMP2 were negatively regulated by miR-616
expression in MCF-7 and MDA-MB-231 cells (Fig. 4C and D). When examining the tissue
samples from the patient cohort, RT-qPCR results revealed that the
mRNA expression levels of TIMP2 were significantly downregulated in
breast cancer tissue samples compared with normal tissue samples
(P<0.05; Fig. 4E). Notably, the
mRNA expression levels of TIMP2 were negatively correlated with
miR-616 levels in the breast cancer tissues (P<0.05; Fig. 4F).
TIMP2 silencing reverses the effects
of miR-616 in breast cancer cells
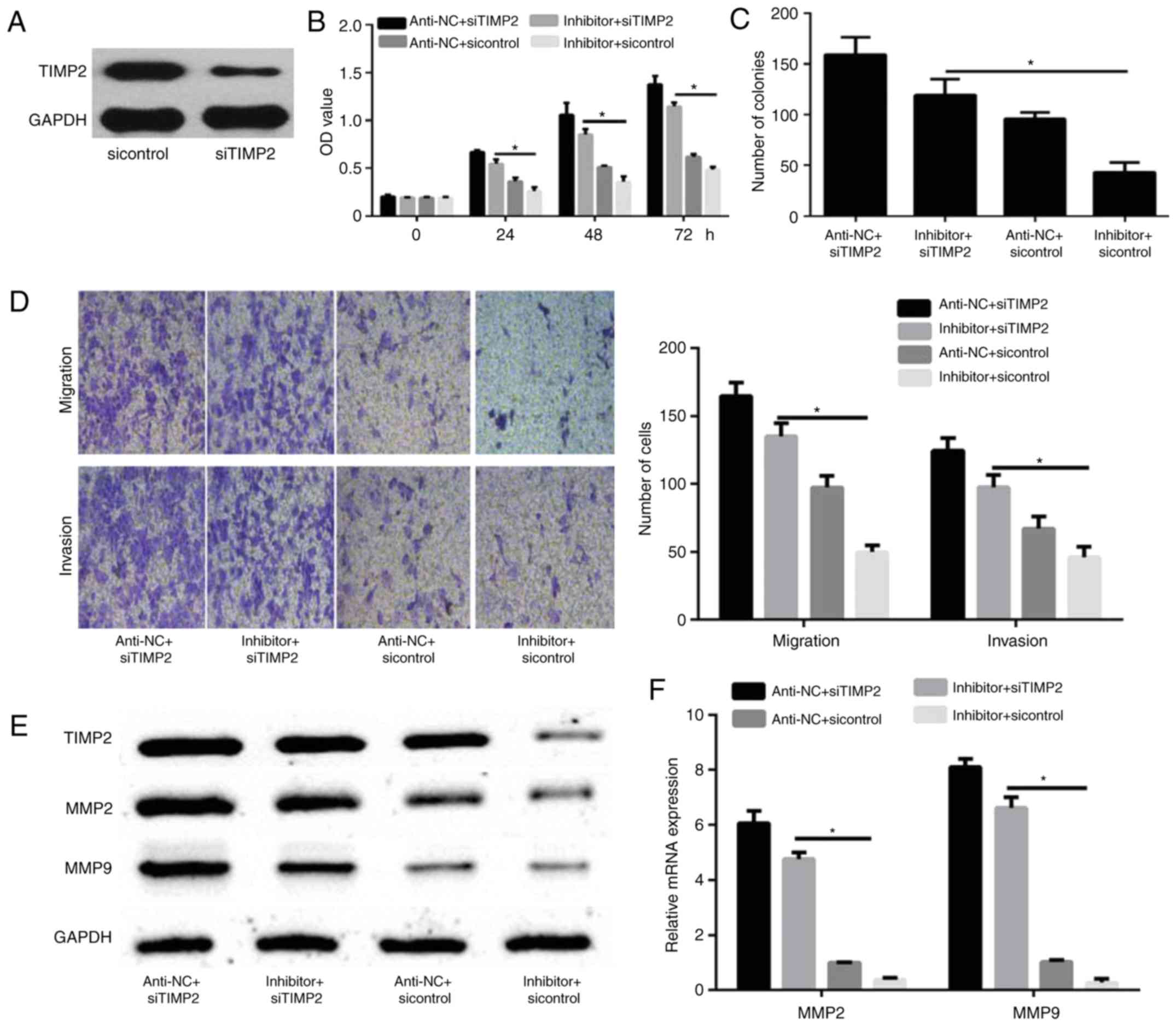
To confirm that TIMP2 is a functional target of
miR-616, MDA-MB-231 cells were transfected with sicontrol or
siTIMP2. Western blot analysis revealed that the expression of
TIMP2 was decreased in siTIMP2-transfected MDA-MB-231 cells
compared with sicontrol-transfected MDA-MB-231 cells (Fig. 5A). MDA-MB-231 cells were then
transfected with miR-616 inhibitor and/or siTIMP2 and their
corresponding controls. CCK-8 and colony formation assays
demonstrated that TIMP2 knockdown significantly promoted cell
proliferation and colony formation, which were initially reduced by
miR-616 silencing (P<0.05; Fig. 5B
and C). Furthermore, a Transwell assay demonstrated that TIMP2
silencing significantly increased the migration and invasion
capabilities of MDA-MB-231 cells, which were initially reduced by
miR-616 inhibitor (P<0.05; Fig.
5D). Finally, it was demonstrated that TIMP2 knockdown enhanced
the protein and mRNA expression levels of MMP2 and MMP9, which were
reduced by miR-616 inhibitor alone (P<0.05; Fig. 5E and F).
Discussion
Investigation of the molecular mechanisms involved
in tumor progression facilitates the identification of molecules
that may act as new targets for the treatment of breast cancer
(22,23). miRNAs serve important roles in
tumorigenic processes, including cell viability, proliferation,
apoptosis, migration and invasion (22,23).
miRNAs may serve as oncogenes or tumor suppressors, depending on
the type of tissue and the context of expression (5). However, the underlying molecular
mechanisms of miRNAs in cancer progression remain unclear.
Certain types of miRNAs, including miR-155 and
miR-21, serve critical roles in the progression of breast cancer
(8,24). miR-616 is a novel miRNA that serves
as an oncogene in glioma, gastric cancer, prostate cancer,
non-small cell lung cancer and hepatocellular carcinoma (10–15). In
breast cancer, to the best of our knowledge, the expression levels,
biological function and molecular mechanism of miR-616 have not
been fully elucidated. In the present study, the expression levels
of miR-616 were significantly upregulated in breast cancer cell
lines and tissues. Functional in vitro experiments
demonstrated that miR-616 promoted the proliferation, migration and
invasion of breast cancer cells. According to these data, miR-616
may act as an oncogene and may be a potential biomarker for breast
cancer.
miRNAs exert their biological roles in cancer by
regulating their target genes (5).
Therefore, identifying the association of a miRNA and its target
gene is important for elucidating the molecular mechanism
underlying its action in cancer. miR-616 can regulate numerous
genes, including phosphatase and tensin homolog, SRY-box 7, tissue
factor pathway inhibitor 2 and glycogen synthase kinase 3β
(10–14). In the present study, it was
demonstrated that miR-616 could positively regulate the expression
levels of MMP2 and MMP9. Using bioinformatics analysis, it was
identified that the 3′-UTR of TIMP2 contained a miR-616 response
element. TIMP2 inhibits cell proliferation and migration in
vitro by inhibiting the function of MMPs (21,25). In
addition, TIMP2 serves as a tumor suppressor in the progression of
breast cancer (26). However, to the
best of our knowledge, the mechanism of TIMP2 in the promotion of
breast cancer remains unclear and the association of TIMP2 with
miRNAs has not been fully investigated in breast cancer.
For the first time, the present study demonstrated
the expression and function of miR-616 in breast cancer. A negative
correlation was identified between TIMP2 and miR-616 in breast
cancer, which was consistent with a previous study regarding
ovarian cancer (15). Notably, the
present study identified that miR-616 promoted the expression of
MMP2 and MMP9 via TIMP2 in breast cancer. However, certain
limitations were present in the current study. Firstly, the
experiments were performed in vitro. Therefore, in
vivo experiments are required to further support the
conclusions of the current study. In addition, associations between
the expression of miR-616 and clinical features were not
investigated; therefore, this is required in future studies.
Finally, other molecular mechanisms of miR-616 in the progression
of breast cancer should be examined in the future.
In summary, the present data demonstrated that
miR-616 enhanced the proliferation, migration and invasion of
breast cancer cells by directly targeting TIMP2. Although clinical
applications should be further investigated, the present study
revealed that the miR-616/TIMP2/MMP axis may serve as role in the
regulation of breast cancer progression and identified miR-616 as a
potential novel therapeutic target for the treatment of breast
cancer.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
All data analyzed during this study are included in
the published article.
Authors' contributions
CY conceived, designed and conducted all
experiments, and wrote the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Medical Ethics
Committee of Dezhou No. 2 People's Hospital (Dezhou, China). All
patients included in the current study provided written informed
consent.
Patients consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2017. CA Cancer J Clin. 67:7–30. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ferlay J, Soerjomataram I, Dikshit R, Eser
S, Mathers C, Rebelo M, Parkin DM, Forman D and Bray F: Cancer
incidence and mortality worldwide: Sources, methods and major
patterns in GLOBOCAN 2012. Int J Cancer. 136:E359–E386. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Valastyan S and Weinberg RA: Tumor
metastasis: Molecular insights and evolving paradigms. Cell.
147:275–292. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Weigelt B, Peterse JL and van't Veer LJ:
Breast cancer metastasis: Markers and models. Nat Rev Cancer.
5:591–602. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shen J, Stass SA and Jiang F: MicroRNAs as
potential biomarkers in human solid tumors. Cancer Lett.
329:125–136. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Piasecka D, Braun M, Kordek R, Sadej R and
Romanska H: MicroRNAs in regulation of triple-negative breast
cancer progression. J Cancer Res Clin Oncol. 144:1401–1411. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hu W, Tan C, He Y, Zhang G, Xu Y and Tang
J: Functional miRNAs in breast cancer drug resistance. Onco Targets
Ther. 11:1529–1541. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu ZH, Lin C, Liu CC, Jiang WW, Huang MZ,
Liu X and Guo WJ: MiR-616-3p promotes angiogenesis and EMT in
gastric cancer via the PTEN/AKT/mTOR pathway. Biochem Biophys Res
Commun. 501:1068–1073. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bai QL, Hu CW, Wang XR, Shang JX and Yin
GF: MiR-616 promotes proliferation and inhibits apoptosis in glioma
cells by suppressing expression of SOX7 via the Wnt signaling
pathway. Eur Rev Med Pharmacol Sci. 21:5630–5637. 2017.PubMed/NCBI
|
|
12
|
Wang D, Cao Q, Qu M, Xiao Z, Zhang M and
Di S: MicroRNA-616 promotes the growth and metastasis of non-small
cell lung cancer by targeting SOX7. Oncol Rep. 38:2078–2086. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang D, Zhou P, Wang W, Wang X, Li J, Sun
X and Zhang L: MicroRNA-616 promotes the migration, invasion and
epithelial-mesenchymal transition of HCC by targeting PTEN. Oncol
Rep. 35:366–374. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ma S, Chan YP, Kwan PS, Lee TK, Yan M,
Tang KH, Ling MT, Vielkind JR, Guan XY and Chan KW: MicroRNA-616
induces androgen-independent growth of prostate cancer cells by
suppressing expression of tissue factor pathway inhibitor TFPI-2.
Cancer Res. 71:583–592. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chen Z, Zhu J, Zhu Y and Wang J:
MicroRNA-616 promotes the progression of ovarian cancer by
targeting TIMP2. Oncol Rep. 39:2960–2968. 2018.PubMed/NCBI
|
|
16
|
Chien YC, Liu LC, Ye HY, Wu JY and Yu YL:
EZH2 promotes migration and invasion of triple-negative breast
cancer cells via regulating TIMP2-MMP-2/-9 pathway. Am J Cancer
Res. 8:422–434. 2018.PubMed/NCBI
|
|
17
|
Chao Y, Hu K, Wang X and Wang L:
MicroRNA-552 promotes migration and invasion of osteosarcoma
through targeting TIMP2. Biochem Biophys Res Commun. 511:63–68.
2019. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yin S, Zhang Q, Wang Y, Li S and Hu R:
MicroRNA-130a regulated by HPV18 E6 promotes proliferation and
invasion of cervical cancer cells by targeting TIMP2. Exp Ther Med.
17:2837–2846. 2019.PubMed/NCBI
|
|
19
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ren F, Tang R, Zhang X, Madushi WM, Luo D,
Dang Y, Li Z, Wei K and Chen G: Overexpression of MMP family
members functions as prognostic biomarker for breast cancer
patients: A systematic review and meta-analysis. PLoS One.
10:e01355442015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tjomsland V, Pomianowska E, Aasrum M,
Sandnes D, Verbeke CS and Gladhaug IP: Profile of MMP and TIMP
expression in human pancreatic stellate cells: Regulation by IL-1α
and TGFβ and implications for migration of pancreatic cancer cells.
Neoplasia. 18:447–456. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Iorio MV and Croce CM: MicroRNA
dysregulation in cancer: Diagnostics, monitoring and therapeutics.
A comprehensive review. EMBO Mol Med. 4:143–159. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pasquinelli AE: MicroRNAs and their
targets: Recognition, regulation and an emerging reciprocal
relationship. Nat Rev Genet. 13:271–282. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu L, Mao X, Shi P, He B, Xu K, Zhang S
and Wang J: MicroRNAs in the prognosis of triple-negative breast
cancer: A systematic review and meta-analysis. Medicine
(Baltimore). 96:e70852017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kurzawski M, Kaczmarek M, Kłysz M,
Malinowski D, Kazienko A, Kurzawa R and Droździk M: MMP2, MMP9 and
TIMP2 polymorphisms affect sperm parameters but not fertility in
Polish males. Andrologia. 49:2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim HJ, Park CI, Park BW, Lee HD and Jung
WH: Expression of MT-1 MMP, MMP2, MMP9 and TIMP2 mRNAs in ductal
carcinoma in situ and invasive ductal carcinoma of the breast.
Yonsei Med J. 47:333–342. 2006. View Article : Google Scholar : PubMed/NCBI
|