Introduction
Colorectal cancer (CRC) is one of the most common
malignant tumors worldwide (1). At
present, the main treatments for CRC are surgery, radio- and
chemotherapy (2). In China, CRC is a
serious health problem due to its high rate of mortality, which
represented the third highest cancer-associated mortality rate
nationally in 2012 (3).
Additionally, the mortality rate of CRC in China was 4–8 times
higher than that in Europe and America in 2012 (4). Associated treatments and the prognosis
for patients with CRC are poor. Advanced CRC is usually associated
with invasion and metastasis of tumor cells and distant organ
metastases, making diagnosis more difficult and greatly increasing
the difficulty of treatment (5–7).
microRNAs (miRNAs/miRs) are a widely studied type of
non-coding RNA and have been found to be the closely associated
with the pathogenesis and efficacy of treatment of cancer (8). Numerous miRNAs have been demonstrated
to serve important roles in the growth, differentiation and
apoptosis of cancer cells (9).
Alterations in the expression levels and functions of specific
miRNAs may serve a key role in the genesis of diverse cancer types
(10–14). Our previous study demonstrated that
miR-27a-3p promoted gastric cancer cell proliferation in
vitro as well as tumor growth in vivo (15). miR-27a has further been identified to
act as an oncogene in MGC803 cells and knockdown of miR-27a
inhibits cell growth and was determined to be dose-dependent
(16). Certain studies have
demonstrated that overexpression of miR-27a-3p significantly
promotes growth of cancer cells in glioma (17), hepatocellular carcinoma (18), esophageal cancer (19), renal cell carcinoma (20) and nasopharyngeal carcinoma (21). However, the role of miR-27a-3p in CRC
and the underlying mechanisms are not well defined.
B-cell translocation gene (BTG)1, BTG2, BTG3, BTG4,
transducer of ERBB2 and transducer of ERBB2 2 belong to the BTG
family. As tumor suppressors, these proteins suppress cell
proliferation and cell cycle progression, and induce
differentiation (22,23). In particular, BTG1 has been reported
to regulate cell cycle progression in a variety of cells, including
breast cancer (24) and renal cell
carcinoma cells (25) and has been
suggested to be a potential therapeutic target (26–30).
BTG1 expression is highest in the G0/G1
phases of the cell cycle and suppresses the progression of cells
through G1 phase (31).
While BTG1 exhibits nuclear localization, associated signals enable
it to undergo nucleo-cytoplasmic shuttling (32). Notably, BTG1 has been reported to
increase and enhance antisense Bcl-2-induced cytotoxicity in MCF-7
and MDA-MB-231 breast cancer cells, and leukemia cell lines
(33,34). It has been reported previously that
BTG1 inhibits the proliferation, migration and invasion of gastric
cancer cells (35,36), and is positively associated with
increased expression of cyclin D1 and Bax, also known as anti-tumor
protein (37). Overexpression of
BTG1 serves an important role in CRC. Specifically, BTG1 expression
reverses the aggressive phenotype and may be a candidate for gene
therapy in CRC (38).
In the present study, miR-27a-3p was demonstrated to
be overexpressed in human CRC tissues and colon cancer cell lines.
Furthermore, the anti-proliferative gene BTG1 was predicted to be a
direct target of miR-27a-3p. Therefore, the present study aimed to
explore the association between miR-27a-3p and tumor growth,
apoptosis, cell cycle distribution and the Ras/mitogen-activated
extracellular signal-regulated kinase (MEK)/ERK signaling pathway.
In summary, the miR-27a-3p/BTG1 axis could have potential
implications for diagnostic and therapeutic approaches in CRC.
Materials and methods
Tissues
A total of 20 paired samples of human CRC and
matched normal tissues were collected at Minhang Hospital
(Affiliated to Fudan University) between December 2016 and February
2017. There were 13 males and 7 females, aged 38–62 years, included
in the present study. The surgical procedures performed to obtain
the tissues were laparoscopic radical resection of colorectal
cancer. The lesion was considered to be normal tissue at a margin
>5 cm from the edge of the tumor. The samples were stored in
liquid nitrogen following collection during surgery and were
subsequently stored at −80°C. The use of these tissues was approved
by the Institutional Review Board of Minhang Branch, Zhongshan
Hospital and Fudan University Shanghai Cancer Center, and signed
informed consent was obtained from all participants.
Plasmid construction
The homo sapiens-miR-27a (hsa-miR-27a) expression
vector pEGFP-C1-miR-27a (+), the hsa-miR-27a competitive inhibitor
vector pEGFP-C1-miR-27a (−) and the vector pEGFP-C1 were obtained
from the State Key Laboratory of Bioreactor Engineering and
Shanghai Key Laboratory of New Drug Design, School of Pharmacy,
East China University of Science and Technology. The hsa-miR-27a
expression vector pEGFP-C1-miR-27a (+) contains primary-miR-27a and
some of its flanking sequences (33). The sequences of the has-miR-27a were:
Forward, 5′-CCGCTCGAGACTGGCTGCTAGGAAGGTG-3′ and reverse,
5′-GCGAATTCTTGCTGTAGCCTCCTTGTC-3′. The hsa-miR-27a competitive
inhibitor vector pEGFP-C1-miR-27a (−) was designed as a sponge of
miR-27a with repeated binding sites complementary cloned into the
pEGFP-C1 vector. The sequences of the miR-27a sponge were: Forward,
5′-CCCAAGCTTACTGTGAAACTGTGAAACGTGAAACTGTGAAACTGTGAAACTGTGAATCTAGAGC-3′
and reverse,
5′-GCTCTAGATTTCACAGTTTCACAGTTTCACAGTTTCACAGTTTCACAGTAAGCTTGGG-3′.
The construction of plasmids and respective sequences were
performed as described previously (14). DNA was extracted from HCT-116 cells
using TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
and the coding region of BTG1 cDNA was amplified by PCR using 2X
Hieff™ PCR Master mix (Yeasen Biotech Co., Ltd.)
according to the manufacturer's protocol. The thermocycling
conditions were: 95°C for 1 min; followed by 35 cycles of 62°C for
50 sec, 94°C for 30 sec, 60°C for 50 sec and 72°C for 35 sec; and
final extension at 72°C for 10 min. PCR products were purified from
the agarose gel using a gel purification kit (Promega Corporation)
and were ligated to empty pcDNA3.1 vector at a 3:1 ratio at 4°C for
16 h. Recombinant plasmids were then transferred to E. coli
JM109 competent cells (cat. no. 9052; Takara Bio, Inc.). These
cells were normally stored at −80°C. Prior to use, the E.
coli JM109 competent cells were thawed on ice, and the pcDNA3.1
plasmid was mixed with the competent state at a ratio of 1:100,
placed on ice for 30 min, then placed at 42°C for 60–90 sec,
incubated for 2–3 min and diluted in LB medium (Hangzhou Baisi
Biotechnology Co., Ltd.). An appropriate amount of mixed solution
was applied, and subsequently E.coli were cultured overnight
in LB medium-containing plates with 1% ampicillin (Beijing Solarbio
Science & Technology Co., Ltd.) for amplification in a normal
environment at 37°C. Specific primers were designed by
PrimerPremier 6.0 (Premier Biosoft International). The primer
sequences were: Forward, 5′-GGAATTCATGCATCCCTTCTACACCCGG-3′ and
reverse, 5′-CGACGCGTTTAACCTGATACAGTCATCAT-3′. Purified pcDNA-BTG1
recombinant plasmids were treated with EcoRI + XhoI
restriction enzymes (Promega Corporation) at 37°C for 4 h. Digested
products were separated using 1.5% agarose gel electrophoresis with
ethidium bromide. DNA bands were identified by UV transilluminator
(FR-200A; Shanghai Furi Science & Technology Co., Ltd.).
Purified recombinant plasmids (1 µg/µl; A260/A280=1.8) were used to
transfect cultured cells.
Cell culture and transfection
The human colon cancer cell lines HCT-116, HCT8,
SW480, HT29, LOVO and Caco2 and the normal colorectal epithelial
NCM460 cell line were purchased from the Type Culture Collection of
the Chinese Academy of Sciences. All cell lines were cultured in
DMEM (Gibco; Thermo Fisher Scientific, Inc.) supplemented with 10%
FBS (Gibco; Thermo Fisher Scientific, Inc.) containing 100 U/ml
penicillin and 100 U/ml streptomycin (Thermo Fisher Scientific,
Inc.) with 5% CO2 at 37°C in a humidified environment.
Cell transfection was performed using Lipofectamine®
3000 reagent (Invitrogen; Thermo Fisher Scientific, Inc.) according
to the manufacturer's protocol. In brief, cultured cells were
seeded into 6-well plates (5×105 cells/well). Then, 5 µg
miR-27 mimic or 5 µg miR-27 inhibitor and 10 ml P3000™
(Invitrogen; Thermo Fisher Scientific, Inc.) reagent were dissolved
in 120 µl DMEM each. Following incubation at room temperature for
20 min, both miRNA and P3000™ reagent were slowly added to cultured
HCT-116 cells. After 24 h of incubation at 37°C, the culture medium
was changed. The control (no miRNA transfection) and empty
(transfected with empty pEGFP-C1) group transfections were
performed in parallel. Transfection efficiency was determined by
reverse transcription-quantitative PCR (RT-qPCR) after 48 h.
Additional subsequent experiments were performed 48 h after the
cells were transfected.
RNA extraction and RT-qPCR
Total RNA was extracted from HCT-116 cells using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.).
Isolated miRNAs were reverse transcribed using TransScript miRNA
First-Strand cDNA Synthesis SuperMix (Beijing Transgen Biotech Co.,
Ltd.). Briefly, 5 µl total RNA, 1 µl TransScript® miRNA
RT Enzyme mix (Beijing Transgen Biotech Co., Ltd.), 10 µl 2X TS
miRNA Reaction mix (Beijing Transgen Biotech Co., Ltd.) and 4 µl
RNase-free water were mixed according to the manufacturer's
instructions and incubated for 1 h at 37°C, followed by incubation
for 5 sec at 85°C. Subsequently, 0.2 µM forward primer and 10 µl 2X
TransScript® Tip/Top Green qPCR SuperMix (Beijing
Transgen Biotech Co., Ltd.) were mixed. The thermocycling
conditions were: 94°C for 30 sec; followed by 45 cycles of 94°C for
5 sec, 60°C for 15 sec and 72°C for 15 sec; dissociation stage. The
primer sequence for miR-27a-3p was 5′-TTCACAGTGGCTAAGTTCCGC-3′.
mRNA was reverse transcribed with TransScript One-Step gDNA Removal
and cDNA Synthesis SuperMix (Beijing Transgen Biotech Co., Ltd.).
qPCR was performed according to the manufacturer's protocol of the
TransScript Top Green qPCR SuperMix (Beijing Transgen Biotech Co.,
Ltd.) using an iCycler thermal cycler (Bio-Rad Laboratories, Inc.).
The thermocycling conditions were: 94°C for 30 sec; followed by 40
cycles of 94°C for 5 sec, 60°C for 15 sec and 72°C for 10 sec;
return to room temperature. The comparative cycle threshold method
(2−ΔΔCq) (39) was used
to conduct the relative quantification of target genes and miRNAs
(10). Levels of miRNA and mRNA were
normalized against U6 snRNA and GAPDH, respectively. The primer
sequences were: BTG1 forward, 5′-AGCTGAACCTGTATCTGCGG-3′ and
reverse, 5′-GAATTCCTGGTGCCAAAGGC-3′; U6 snRNA forward,
5′-ATTGGAACGATACAGAGAAGATT-3′ and reverse,
5′-GGAACGCTTCACGAATTTG-3′; and GAPDH forward,
5′-GGTGAAGGTCGGAGTCAACG-3′ and reverse, 5′-CAAAGTTGTCATGGA-3′.
Transfection efficiency was determined by RT-qPCR.
Western blot analysis
RIPA lysis buffer (Beyotime Institute of
Biotechnology) was used for cell lysis, the protein concentration
was determined using a bicinchoninic acid assay (Thermo Fisher
Scientific, Inc.) and 10% SDS-PAGE was used to separate cellular
proteins (50 µg/lane). Subsequently, protein was transferred to a
0.45 µm PVDF filter at 200 V for 2 h. The subsequent western blot
analysis was carried out as described previously (38). Following blocking in 3% BSA (A8020;
Beijing Solarbio Science & Technology) for 2 h at room
temperature, membranes were incubated with appropriate primary
antibodies in dilution buffer (3% BSA; 1:1,000 dilution) overnight
at 4°C. β-actin was used as a loading control. Membranes were
washed with TBS with 0.1% Tween-20 and incubated with horseradish
peroxidase-conjugated secondary antibodies (cat. nos. 111-035-003
and 115-035-003; Jackson ImmunoResearch Laboratories, Inc.) at a
dilution of 1:2,000 in 3% BSA for 3 h at room temperature. Protein
expression was assessed by enhanced chemiluminescence (cat. no.
D3030L1260; Shanghai Life iLab Biological Technology Co., Ltd.) and
exposure to chemiluminescent film. LabWorks™ Image Acquisition
(version 4.0; UVP, LLC) was used.
Antibodies used in the present study, including
anti-MEK (cat. no. 4694), anti-ERK (cat. no. 9102), anti-p-ERK
(cat. no. 4370), anti-Ras (cat. no. 3965), anti-cyclin D1 (cat. no.
2922), anti-cyclin E1 (cat. no. 4129), anti-cleaved caspase-3 (cat.
no. 9661), anti-cleaved poly(ADP-ribose) polymerase 1 (PARP1; cat.
no. 5625) and anti-β-actin (cat. no. 3700), were purchased from
Cell Signaling Technology, Inc. The anti-uncleaved caspase-3 (cat.
no. ab13847) and anti-c-Myc (cat. no. ab32072) antibodies were
purchased from Abcam. The antibody against BTG1 was purchased from
ProteinTech Group, Inc. (cat. no. 14879-1-AP). The anti-p-MEK-1
(cat. no. sc-101733) antibody was purchased from Santa Cruz
Biotechnology, Inc.
Proliferation and clone formation
assays
Using the MTT method, the proliferation of HCT-116
cells transfected with mimics/inhibitor was assessed after 5 days
of transfection. Cultured cells were seed into 96-well plates
(1×104 cells/well), and after 0, 24, 48 and 72 h, 20 µl
MTT solution (5 µg/ml; Sigma-Aldrich; Merck KGaA) was added to each
well and the plate was further incubated at 37°C for 4 h.
Subsequently, the medium was aspirated and the wells were washed
with PBS, allowed to dry for ~2 h and 200 µl DMSO (Sigma-Aldrich;
Merck KGaA) was added to each well. The optical density was
measured at 492 and 630 nm wavelength. Clone formation assays were
performed as described previously (15).
miR-27a-3p potential target gene
prediction
miRbase (version 20.0; http://www.mirbase.org/) was used to find the miRNA
sequence. Subsequently, TargetMiner (https://www.isical.ac.in/~bioinfo_miu/TargetMiner.html),
PicTar (https://pictar.mdc-berlin.de/cgi-bin/PicTar_vertebrate.cgi)
(40) and TargetScan (version 7.1;
www.targetscan.org) were used to predict the
potential target genes of miR-27a-3p, and the intersection of the
results was the target sequence.
Flow cytometry analysis
A Cell Cycle Assay kit (cat. no. FXP0211-200; 4A
Biotech Co., Ltd.) was used according to the manufacturer's
protocols and cell cycle distribution was analyzed by flow
cytometry using a Fluorescence-activated Cell Sorting Vantage
cytometer (BD Biosciences). In brief, the cells were harvested and
fixed in ice-cold 70% (v/v) ethanol for 24 h at 4°C. The cell
pellet was collected by centrifugation at 500 × g for 10 min at
4°C, resuspended in PBS, and stained with a mixture of RNase (10
µg/ml) and propidium iodide (50 µg/ml) in sodium citrate containing
0.5% Triton X-100 for 20 min in the dark at room temperature. The
percentages of cells in the different cell cycle phases were
analyzed using WinMDI software (version 2.8; K. Trotter, The
Scripps Research Institute, La Jolla, CA, USA).
Cells (1×105) were stained with the FITC
Annexin-V Apoptosis Detection kit (R&D Systems, Inc.) according
to the manufacturer's protocol. Briefly, cells from cultures were
collected, washed with cold PBS, and then stained with
Annexin-V-FITC (0.25 µg/ml) and propidium iodide for 15 min at room
temperature in the dark. The stained cells were then analyzed by
flow cytometry, acquiring 1×105 events gated according
to a large gate established on cell forward and side scatters
within 30 min of staining. The apoptotic cell ratio was analyzed by
flow cytometry using a fluorescence-activated cell sorting vantage
cytometer (BD Biosciences). FlowJo version 10.5.0 software (FlowJo
LLC) was used for analysis.
TUNEL analysis
Cells (1×105) were plated in 6-well
culture plates containing gelatin-coated cover slips. Adherent
cells were transfected with mimics or inhibitor of miR-27a-3p and
pcDNA-BTG1 for 48 h. Cells in the control group were not
transfected. Treated cells and control cells were stained using a
TUNEL FITC Apoptosis Detection kit (cat. no. C1088; Beyotime
Institute of Biotechnology) according to the manufacturer's
protocols. In brief, 1×105 cells were seeded on 6-well
culture plates. Cells were washed with PBS for 5 min three times
and fixed in 4% paraformaldehyde (cat. no. AR1069; Wuhan Boster
Biological Technology, Ltd.) at 4°C for 20 min. The cells were
incubated with the TUNEL enzyme for 60 min at 3°C. Finally, the
fluorescent reaction was counterstained with DAPI (1:1,000 in PBS;
Sigma-Aldrich; Merck KGaA) to dye the nucleus for 10 min at room
temperature. Antifade mounting medium (cat. no. P0126; Beyotime
Institute of Biotechnology) was used. Images from four fields of
view were captured using a fluorescence microscope (magnification,
×20; DMI3000B; Leica Microsystems, Inc.).
Statistical analysis
Data are presented as the mean ± SD. A
Kruskal-Wallis-Test was performed to evaluate the significance of
differences between the clinical tumor and normal samples, using a
Steel Dwass post hoc test. Other data were analyzed using one-way
ANOVA with a Tukey's HSD post hoc test. P<0.05 was considered to
indicate a statistically significant difference, and P<0.01 was
considered to indicate a statistically highly significant
difference. Analysis was performed using Student's t-test. All data
were representative of an average of three independent experiments.
All statistical analyses were performed using GraphPad Prism 5.0
software (GraphPad Software, Inc.).
Results
miR-27a-3p is overexpressed in CRC
tissues and colon cancer cells
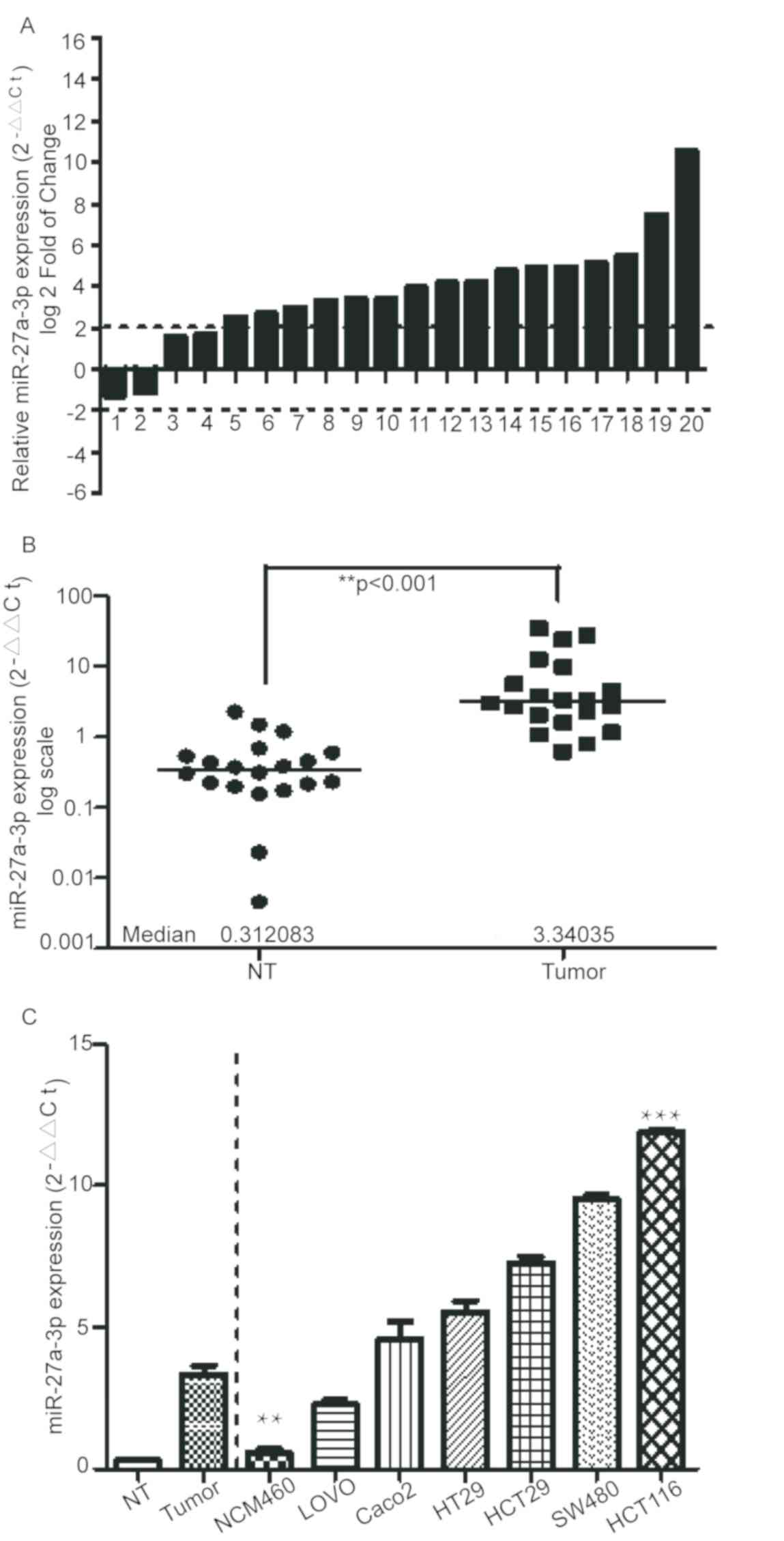
To determine the expression levels of miR-27a-3p,
expression levels in 20 paired CRC and normal tissues were assessed
by RT-qPCR. Compared with in the corresponding non-tumor samples,
the expression of miR-27a-3p was markedly increased in the CRC
tissues, with a fold-change >2.0 (Fig. 1A and B). In addition, consistent with
the results for the clinical CRC samples, expression levels of
miR-27a-3p were identified to be markedly upregulated in colon
cancer cell lines (HCT-116, HCT8, SW480, HT29, LOVO and Caco2)
compared with in the normal colorectal cell line NCM460 (Fig. 1C). Overall, these data indicated that
expression of miR-27a-3p was increased in CRC tissues and colon
cancer cell lines.
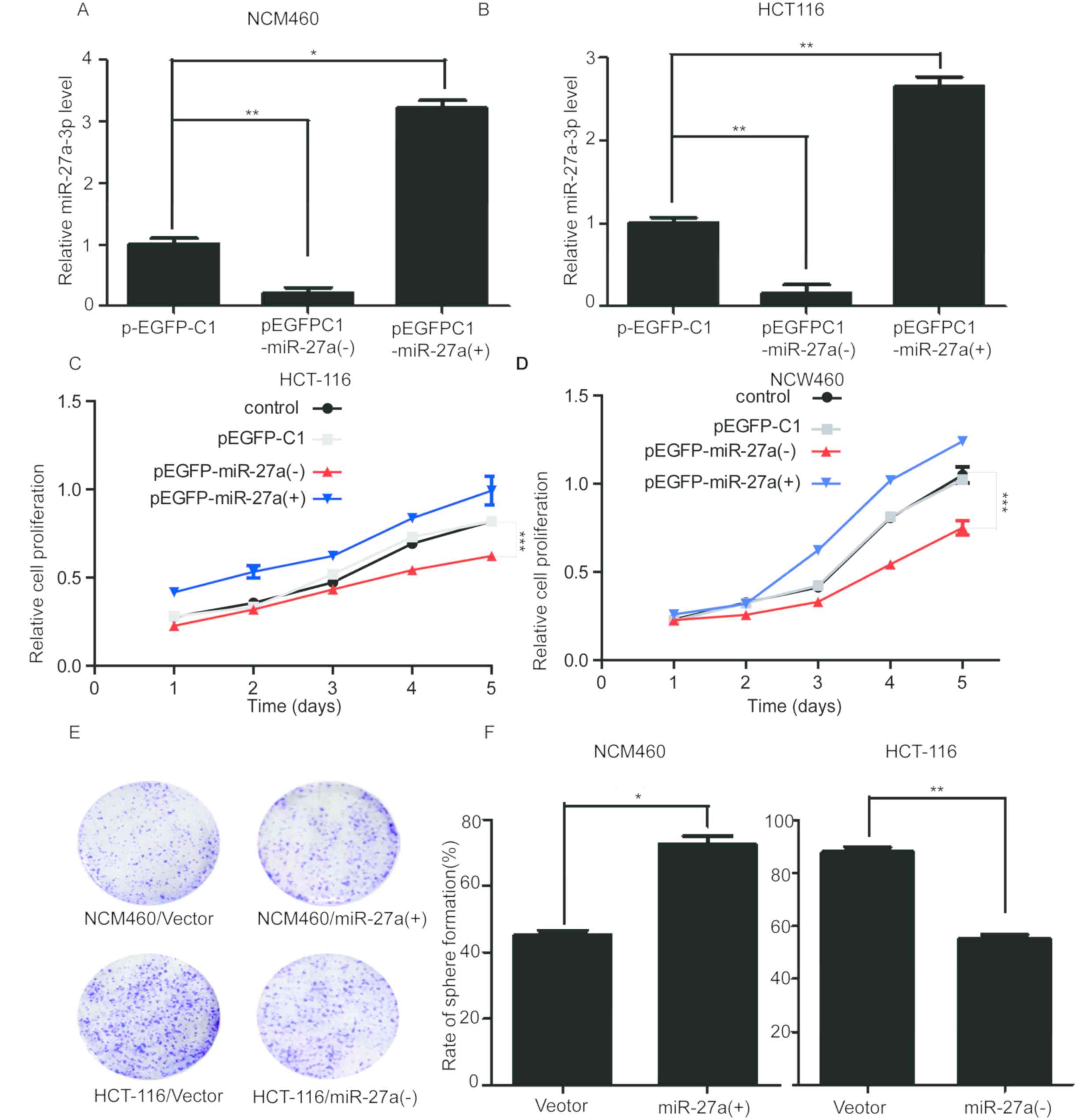
miR-27a-3p promotes colon cancer cell
proliferation
To evaluate the role of miR-27a-3p in colon cancer
cells, the miR-27a overexpression vector pEGFP-C1-miR-27a (+) was
established by inserting a miR-27a precursor containing flanking
sequences into the pEGFP-C1 vector. Subsequently, the miR-27a
expression vector or a competitive inhibitor plasmid,
pEGFP-C1-miR-27a (−), were transfected into HCT-116 and NCM460
cells. RT-qPCR was used to verify the transfection efficiency.
Results revealed that in pEGFP-C1-miR-27a (+) transfected cells,
expression of miR-27a-3p was significantly upregulated, while in
pEGFP-C1-miR-27a (−) transfected cells it was significantly
decreased compared with the pEGFP-C1 transfected cells (Fig. 2A and B). The results of cell
proliferation assays indicated that inhibiting miR-27a-3p
expression reduced the growth rate of HCT-116 cells, whereas
miR-27a-3p overexpression had no obvious effect on cell
proliferation (Fig. 2C).
Additionally, cell proliferation assays were used to detect the
effect on NCW460 cells. It was revealed that miR-27a-3p
overexpression promoted cell proliferation, although this effect
was not observed to be statistically significant, while inhibiting
miR-27a-3p expression had a significant effect on cell
proliferation (Fig. 2D). Clone
formation assays revealed the effect of inhibiting miR-27a-3p
expression to inhibit the growth in HCT-116 cells and
overexpression of miR-27a-3p could promote the growth of NCM460
cells (Fig. 2E and F). Since
miR-27a-3p was overexpressed in CRC tissues and cells, it could
promote the proliferation of cancer cells. The MTT assay revealed
that miR-27a-3p overexpression could promote the proliferation of
NCW460 cells, indicating that overexpression of miR-27a-3p could
promote the proliferation of normal cells and cancer cells.
Inhibiting miR-27a-3p expression could reduce the proliferation of
HCT-116 cells. The role of miR-27a-3p in normal and cancer cells
was investigated by overexpression and knockdown, respectively.
Since miR-27a-3p overexpression had no effect on the growth of
HCT-116 cells in the MTT assay, inhibition of miR-27a-3p also had
no effect on the NCM460 cells in the MTT assay. Therefore, the
clone formation assay with significant influence was selected.
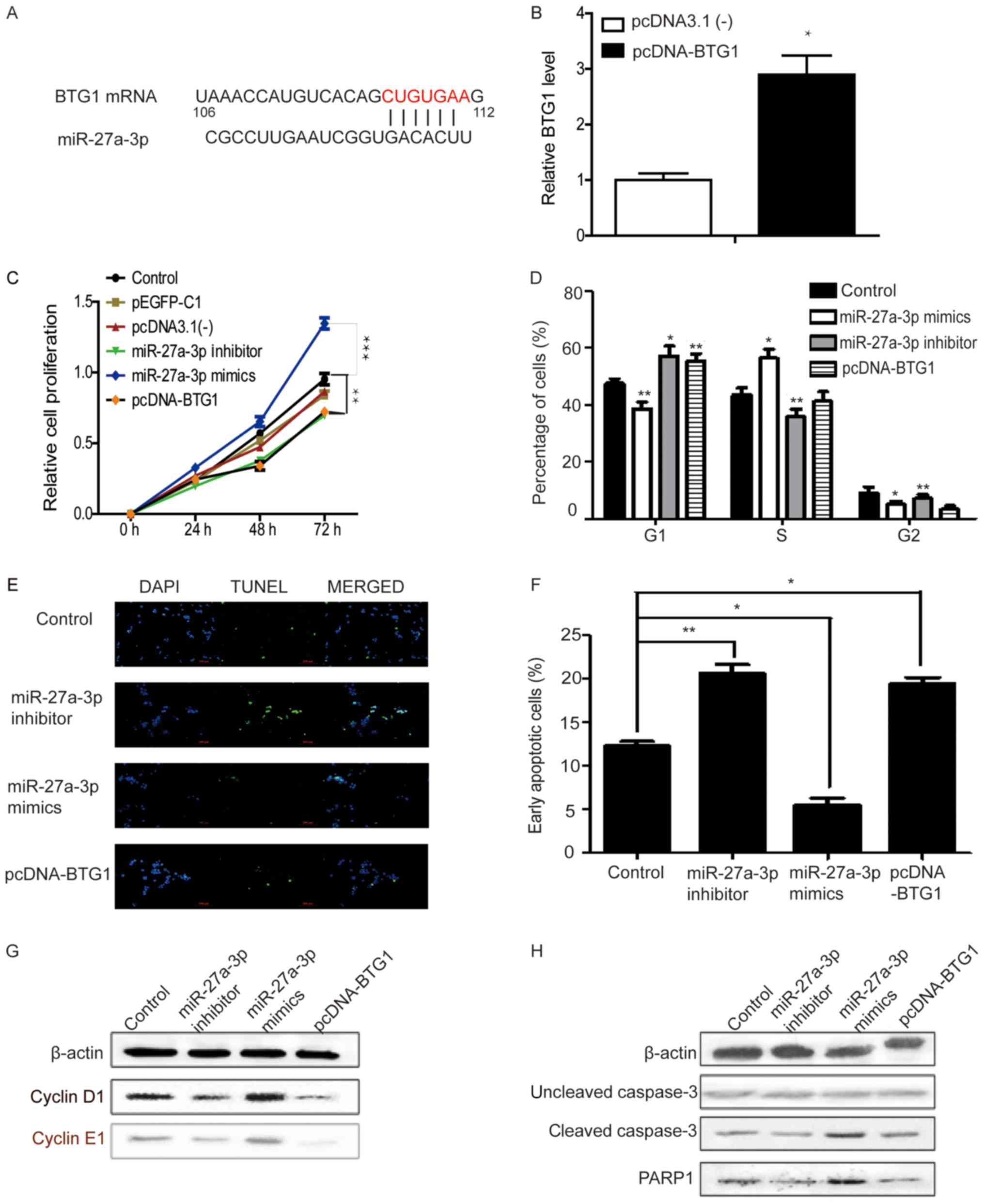
miR-27a-3p and BTG1 regulate cell
cycle progression and apoptosis in colon cancer cells
It has been reported that BTG1 acts as a tumor
suppressor in several human malignant tumors, including CRC
(38). It is also known to be a gene
that induces apoptosis and inhibits proliferation (23). In order to improve the understanding
of the functional mechanism of miR-27a-3p in colorectal
tumorigenesis, direct targets of miR-27a-3p that may have
biological functions need to be identified. Therefore, possible
targets of miR-27a-3p were predicted using the target prediction
programs TargetMiner (41), miRbase,
PicTar (40) and TargetScan
(42). BTG1, which is involved in
cell proliferation, was indicated to be associated with the
biological function of miR-27a-3p among hundreds of potential
candidates, since its 3′-untranslated region contains a putative
target sequence for miR-27a-3p (Fig.
3A). Transfection efficiency for the BTG1 overexpression system
was determined by RT-qPCR illustrating that BTG1 expression was
significantly increased compared with the empty vector (Fig. 3B). To further investigate the effects
of miR-27a-3p and BTG1 on HCT-116 cells, miR-27a-3p
inhibitor/mimics or BTG1 plasmid were transfected into the cells to
regulate the respective expression. In MTT assays at 48 and 72 h,
miR-27a-3p inhibitor and BTG1 markedly suppressed the proliferation
of HCT-116 cells. There was no difference identified between the
vector pEGFP-C1, pcDNA3.1(−) and control groups in terms of cell
growth. The miR-27a-3p mimics significantly promoted cells growth
compared with the control group (Fig.
3C). Additionally, following transfection with miR-27a-3p
inhibitor or BTG1, the proportion of cells in the G1
phase increased. miR-27a-3p mimic transfection had the opposite
effects (Fig. 3D). Transfection with
miR-27a-3p inhibitor or pcDNA-BTG1 significantly induced apoptosis
in HCT-116 cells and miR-27a-3p mimics had the opposite effect
(Fig. 3E and F). Fig. 3G and H shows the results of the
western blotting analysis of cell cycle and apoptosis-associated
proteins. The present study identified a negative association
between cyclin D1 and cyclin E1 and BTG1. BTG1, similar to the
miR-27a-3p inhibitor, could reduce the protein expression of cyclin
D1 and cyclin E1 (Fig. 3G).
Overexpression of miR-27a-3p could increase cyclin D1 and cyclin E1
expression. Similarly, BTG1 overexpression and miR-27a-3p
inhibition increased the percentage of cells in G1
phase, and miR-27a-3p mimics decreased the percentage of cells in
G1 phase (Fig. 3D). The
protein expression levels of cleaved-caspase 3 were observed in
miR-27a-3p inhibitor/mimics or BTG1 plasmid treated HCT-116 cells
(Fig. 3H), levels of cleaved-caspase
3 were higher in HCT-116 cells treated with miR-27a-3p mimic
compared with the untreated control cells. Collectively, these
results indicated that inhibition of miR-27a-3p or overexpression
of BTG1 reduced proliferation and promoted apoptosis in HCT-116
cells.
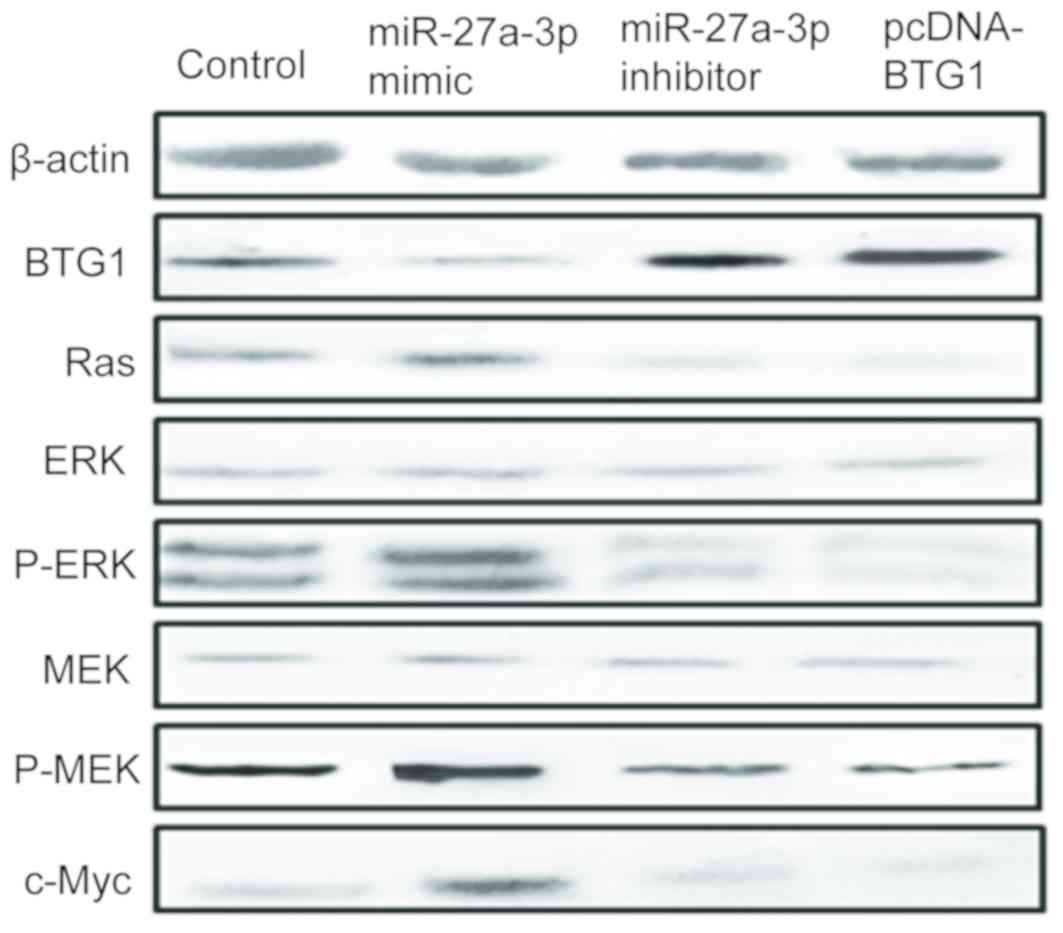
miR-27a-3p affects cell proliferation
and apoptosis via the ERK/MEK signaling pathway
In order to further investigate how miR-27a-3p
affected cell proliferation and apoptosis, the downstream signaling
pathway mediated by BTG1 was studied. A previous study demonstrated
that the ERK/MEK signaling pathway is a downstream pathway of BTG2
in gastric carcinoma (15). This was
verified by western blot analysis in the present study. The results
revealed that inhibition of miR-27a-3p and overexpression of BTG1
decreased the levels of p-ERK and p-MEK-1 but had no effect on ERK
or MEK expression. Inhibition of miR-27a-3p and overexpression of
BTG1 could increase BTG1 expression but decreased Ras and c-Myc
expression (Fig. 4).
Discussion
CRC, a type of malignant tumor, is the second
leading cause of cancer-associated mortality in Asia (43). It is also the third most commonly
diagnosed type of cancer in men and the second in women worldwide
(44). Despite advances in treatment
modalities, the prognosis for patients with CRC has not
significantly improved (45).
miR-27a-3p (has-miR-27a-3p) has been identified as an onco-miRNA in
several solid tumors, including breast (46), ovarian (47), pancreatic (48) and gastric (16) cancer. miR-27a has also been reported
to be a key oncogenic component in CRC and miR-27a is overexpressed
in CRC (49). miR-27a-3p may
accelerate tumorigenesis by targeting several tumor suppressors,
including BTG2 (15), F-box and WD
repeat domain containing 7 (19),
Yes associated protein 1 (50) and
Wnt family member 3A (51).
Regarding BTG1, it has been reported that the tumor suppressor
enhances Hoxb9-mediated transcription and inhibits HeLa cell
proliferation (52). Overexpression
of BTG1 has been detected in apoptotic cells exhibiting DNA
fragmentation and nuclear condensation (53). Additionally, expression levels of
BTG1 are decreased in kidney cancer and are associated with poor
prognosis (54).
Our previous study (15) confirmed that overexpression of
miR-27a-3p promoted gastric cancer cell proliferation in
vitro as well as tumor growth in vivo and that BTG2 was
a direct target of miR-27a-3p in gastric cancer. Zhao et al
(35) reported that BTG1
overexpression suppresses proliferation and cell cycle progression,
and induces apoptosis, autophagy and differentiation in CRC cells.
Therefore, the present study aimed to examine the effects of
miR-27a-3p in CRC and verify BTG1 as a putative target of
miR-27a-3p. The findings of the present study revealed that
miR-27a-3p served as an oncogene in CRC. The expression levels of
miR-27a-3p were significantly increased in CRC tissues compared
with in the paired non-tumor tissues, and miR-27a-3p overexpression
promoted the proliferation of HCT-116 cells. Furthermore, in cell
proliferation experiments, the empty vectors had no effect on cell
proliferation. miR-27a-3p mimic could reduce the proportion of
cells in G1 phase. Additionally, transfection with
miR-27a-3p mimic induced early apoptosis in HCT-116 cells.
Cyclin E and D activate cyclin dependent kinases
(CDKs) during the cell cycle and serve an important regulatory role
in the transition between G1 and S phases. Increased p21
and p27 binding to cyclin and CDKs results in G1 arrest
(55). Cyclin B1-CDK1 is involved in
the early events of mitosis, and CDC25B activates CDC2 to initiate
mitosis (56). In the present study,
increased expression of miR-27a-3p was indicated to result in a
lowered ratio of HCT-116 cells in the G1 phase.
Additionally, the protein expression levels of cyclin E1 and D1
were evaluated.
Zhao et al (35), assessed levels of apoptosis
regulators, including Bcl-2, Bcl-xL, Bax, survivin, X-linked
inhibitor of apoptosis, Akt1 and tumor suppressor p53 in HCT-15
cells overexpressing BTG1. Overexpression of BTG1 decreases Bcl-2
and Bax protein levels and upregulates p53 expression in the human
esophageal cancer cell line ECA-109 (57), human thyroid cancer cell line FTC-133
(58), human breast tumors and
breast cancer cells (MCF-7 and MDA-MB-231) (24), ovarian carcinoma cells OVCAR3
(59) and hematological malignancies
cells (60). Furthermore,
overexpression of BTG1 induces apoptosis of HCT-15 and HCT-116
cells, and decreases mitochondrial potential and increases
senescence only in HCT-116 cells, suggesting differences in the
mechanisms of apoptosis in these two cell lines (35). The induction of cell death requires
activation of one or more members of the well-conserved caspases, a
family of cysteinyl proteases (61,62).
Following proteolytic activation from their proenzyme forms,
caspases cleave various protein substrates, including lamins and
PARP (63). PARP-1 is a member of
the PARP family that is involved in differentiation, proliferation
and tumor transformation (64). The
present study investigated how the expression of miR-27a-3p
affected colon cancer cell apoptosis. The current study suggested
that BTG1 was a downstream target of miR-27a-3p. Different from
previous studies, the present study investigated the effects of
miR-27a-3p on proliferation and apoptosis of CRC cells by BTG1
overexpression. Transfection of HCT-116 cells with miR-27a-3p
mimics revealed the decrease of BTG1 expression and the elevation
of cleaved-caspase3 and PARP1 protein expression. The downregulated
expression of apoptosis-associated genes and their encoded proteins
may be the molecular basis of the difference in the apoptotic
mechanism of BTG1-overexpressing cancer cells. To confirm this, the
mechanism of BTG1-induced apoptosis will be investigated in future
studies.
In human cancers, aberrant activation of the
RAF/MEK/ERK signaling pathway is frequently observed. There has
been increasing evidence revealing the importance of the
RAF/MEK/ERK signaling pathway in tumorigenesis (65–67). The
relatively high frequency of activating mutations of RAS (~20% of
all human types of cancer), an upstream activator of the
well-established RAF/MEK/ERK signaling cascade, as well as frequent
activating mutations in the BRAF kinase gene (~7% of all human
types of cancer) have been observed (68). Therefore, RAF and MEK kinases have
received great attention among the numerous kinases (69,70).
miRNAs perform biological functions by suppressing
their target genes (71). To explore
the functional mechanism of miR-27a-3p in CRC, TargetMiner,
miRBase, PicTar and TargetScan were used to predict possible
targets of miR-27a-3p. From various potential candidates, BTG1 was
selected, since it is accepted as a tumor suppressor and closely
involved in cell proliferation (70,72). The
present study revealed that transfection with miR-27a-3p inhibitor
markedly upregulated the protein expression of BTG1 in HCT-116
cells, while transfection with miR-27a-3p mimic had the opposite
effect. The present study also demonstrated that transfection with
BTG1 had similar effects as transfection with the miR-27a-3p
inhibitor, causing cell cycle arrest at G1 phase and
inducing apoptosis in HCT-116 cells. Furthermore, inhibition of
miR-27a-3p and overexpression of BTG1 decreased the level of p-ERK
and p-MEK-1 in HCT-116 cells. All these results indicated that BTG1
may be a novel target of miR-27a-3p and may serve a tumor
suppressor role in CRC. These findings may provide novel insights
into the mechanisms underlying CRC and have potential applications
in diagnosis and therapy.
In conclusion, the present study showed that
downregulation of miR-27a-3p significantly inhibited proliferation
and enhanced apoptosis in HCT-116 cells. miR-27a-3p may serve an
important role in CRC cells by directly targeting BTG1.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Natural
Science Research Project of Minhang District (grant no.
2017MHZ75).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
CS and YOC designed the study and prepared the
figures. DPH and WYL collated the data and carried out data
analyses. JWL drafted the manuscript and revised it critically for
important intellectual content, in addition they contributed to the
interpretation of data. All authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The study was approved by the Associated
Institutional Review Board of Minhang Branch, Zhongshan Hospital,
Fudan University and signed informed consent was obtained from all
participants.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
BTG1
|
B-cell translocation gene 1
|
|
CRC
|
colorectal cancer
|
|
MEK
|
mitogen-activated extracellular
signal-regulated kinase
|
|
miR-27a-3p
|
microRNA 27a-3p
|
References
|
1
|
Cunningham D, Atkin W, Lenz HJ, Lynch HT,
Minsky B, Nordlinger B and Starling N: Colorectal cancer. Lancet.
375:1030–1047. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics. 2012. CA
Cancer J Clin. 65:872015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhang Y, Shi J, Huang H, Ren J, Li N and
Dai M: Burden of colorectal cancer in China. Zhonghua Liu Xing Bing
Xue Za Zhi. 36:709–714. 2015.(In Chinese). PubMed/NCBI
|
|
4
|
Carrie P: Declines in death from
colorectal cancer in europe deemed major success. Cancer. 124:2876.
2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Al-Sukhni E and Gallinger S: Treatment of
colorectal cancer. Springer Netherlands. 809–812. 2010.
|
|
6
|
Coppedè F, Lopomo A, Spisni R and Migliore
L: Genetic and epigenetic biomarkers for diagnosis, prognosis and
treatment of colorectal cancer. World J Gastroenterol. 20:943–956.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ciombor KK, Wu C and Goldberg RM: Recent
therapeutic advances in the treatment of colorectal cancer. Annu
Rev Med. 66:83–95. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ju J: miRNAs as biomarkers in colorectal
cancer diagnosis and prognosis. Bioanalysis. 2:901–906. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen X, Shi K, Wang Y, Song M, Zhou W, Tu
H and Lin Z: Clinical value of integrated-signature miRNAs in
colorectal cancer: miRNA expression profiling analysis and
experimental validation. Oncotarget. 6:37544–37556. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Yang Q, Jie Z, Ye S, Li Z, Han Z, Wu J,
Yang C and Jiang Y: Genetic variations in miR-27a gene decrease
mature miR-27a level and reduce gastric cancer susceptibility.
Oncogene. 33:193–202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Calin GA, Sevignani C, Dumitru CD, Hyslop
T, Noch E, Yendamuri S, Shimizu M, Rattan S, Bullrich F, Negrini M
and Croce CM: Human microRNA genes are frequently located at
fragile sites and genomic regions involved in cancers. Proc Natl
Acad Sci USA. 10:2999–3004. 2004. View Article : Google Scholar
|
|
12
|
Lu J, Getz G, Miska EA, Alvarez-Saavedra
E, Lamb J, Peck D, Sweet-Cordero A, Ebert BL, Mak RH, Ferrando AA,
et al: MicroRNA expression profiles classify human cancers. Nature.
43:834–838. 2005. View Article : Google Scholar
|
|
13
|
Calin GA and Croce CM: MicroRNA signatures
in human cancers. Nat Rev Cancer. 6:857–866. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Calin GA and Croce CM: MicroRNA-cancer
connection: The beginning of a new tale. Cancer Res. 66:7390–7394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhou L, Liang X, Zhang L, Yang L, Nagao N,
Wu H, Liu C, Lin S, Cai G and Liu J: MiR-27a-3p functions as an
oncogene in gastric cancer by targeting BTG2. Oncotarget.
7:51943–51954. 2016.PubMed/NCBI
|
|
16
|
Liu T, Tang H, Lang Y, Liu M and Li X:
MicroRNA-27a functions as an oncogene in gastric adenocarcinoma by
targeting prohibitin. Cancer Lett. 273:233–242. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Xu W, Liu M, Peng X, Zhou P, Zhou J, Xu K,
Xu H and Jiang S: miR-24-3p and miR-27a-3p promote cell
proliferation in glioma cells via cooperative regulation of MXI1.
Int J Oncol. 42:757–766. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhao N, Sun H, Sun B, Zhu D, Zhao X, Wang
Y, Gu Q, Dong X, Liu F, Zhang Y and Li X: miR-27a-3p suppresses
tumor metastasis and VM by down-regulating VE-cadherin expression
and inhibiting EMT: an essential role for Twist-1 in HCC. Sci Rep.
6:230912016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wu XZ, Wang KP, Song HJ, Xia JH, Jiang Y
and Wang YL: MiR-27a-3p promotes esophageal cancer cell
proliferation via F-box and WD repeat domain-containing 7 (FBXW7)
suppression. Int J Clin Exp Med. 8:15556–15562. 2015.PubMed/NCBI
|
|
20
|
Nakata W, Uemura M, Sato M, Fujita K,
Jingushi K, Ueda Y, Kitae K, Tsujikawa K and Nonomura N: Expression
of miR-27a-3p is an independent predictive factor for recurrence in
clear cell renal cell carcinoma. Oncotarget. 6:21645–21654. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li L and Luo Z: Dysregulated miR-27a-3p
promotes nasopharyngeal carcinoma cell proliferation and migration
by targeting Mapk10. Oncol Rep. 37:2679–2687. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Winkler GS: The mammalian
anti-proliferative BTG/Tob protein family. J Cell Physiol.
222:66–72. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Sasajima H, Nakagawa K and Yokosawa H:
Antiproliferative proteins of the BTG/Tob family are degraded by
the ubiquitin-proteasome system. Eur J Biochem. 269:3596–3604.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhu R, Zou ST, Wan JM, Li W, Li XL and Zhu
W: BTG1 inhibits breast cancer cell growth through induction of
cell cycle arrest and apoptosis. Oncol Rep. 30:2137–2144. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu C, Tao T, Xu B, Lu K, Zhang L, Jiang
L, Chen S, Liu D, Zhang X, Cao N and Chen M: BTG1 potentiates
apoptosis and suppresses proliferation in renal cell carcinoma by
interacting with PRMT1. Oncol Lett. 10:619–624. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
He C, Yu T, Shi Y, Ma C, Yang W, Fang L,
Sun M, Wu W, Xiao F, Guo F, et al: MicroRNA 301A promotes
intestinal inflammation and colitis-associated cancer development
by inhibiting BTG1. Gastroenterology. 152:1434–1448. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee AS, Kranzusch PJ and Cate JH: eIF3
targets cell-proliferation messenger RNAs for translational
activation or repression. Nature. 522:111–114. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Waanders E, Scheijen B, van der Meer LT,
van Reijmersdal SV, van Emst L, Kroeze Y, Sonneveld E, Hoogerbrugge
PM, van Kessel AG, van Leeuwen FN and Kuiper RP: The origin and
nature of tightly clustered BTG1 deletions in precursor B-cell
acute lymphoblastic leukemia support a model of multiclonal
evolution. PLoS Genet. 8:e10025332012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Rouault JP, Rimokh R, Tessa C, Paranhos G
Ffrench M, Duret L, Garoccio M, Germain D, Samarut J and Magaud JP:
BTG1, a member of a new family of antiproliferative genes. Embo J.
11:1663–1670. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Corjay MH, Kearney MA, Munzer DA, Diamond
SM and Stoltenborg JK: Antiproliferative gene BTG1 is highly
expressed in apoptotic cells in macrophage-rich areas of advanced
lesions in Watanabe heritable hyperlipidemic rabbit and human. Lab
Invest. 78:47–58. 1998.PubMed/NCBI
|
|
31
|
Nahta R, Yuan LX, Fiterman DJ, Zhang L,
Symmans WF, Ueno NT and Esteva FJ: B cell translocation gene 1
contributes to antisense Bcl-2-mediated apoptosis in breast cancer
cells. Mol Cancer Ther. 5:93–60. 2006. View Article : Google Scholar
|
|
32
|
Li Y, Choi PS, Casey SC, Dill DL and
Felsher DW: MYC through miR-17-92 suppresses specific target genes
to maintain survival, autonomous proliferation, and a neoplastic
state. Cancer Cell. 26:62–72. 2014. View Article : Google Scholar
|
|
33
|
Zheng HC, Li J, Shen DF, Yang XF, Zhao S,
Wu YZ, Takano Y, Sun HZ, Su RJ, Luo JS and Gou WF: BTG1 expression
correlates with pathogenesis, aggressive behaviors and prognosis of
gastric cancer: A potential target for gene therapy. Oncotarget.
19:685–705. 2015.
|
|
34
|
Williams G and Stoeber K: The cell cycle
and cancer. J Pathol. 226:352–364. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao S, Chen SR, Yang XF, Shen DF, Takano
Y, Su RJ and Zheng HC: BTG1 might be employed as a biomarker for
carcinogenesis and a target for gene therapy in colorectal cancers.
Oncotarget. 8:7502–7520. 2017.PubMed/NCBI
|
|
36
|
Shen K, Liang Q, Xu K, Cui D, Jiang L, Yin
P, Lu Y, Li Q and Liu J: MiR-139 inhibits invasion and metastasis
of colorectal cancer by targeting the type I insulin-like growth
factor receptor. Biochem Pharmacol. 84:320–330. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Zou F, Mao R, Yang L, Lin S, Lei K, Zheng
Y, Ding Y, Zhang P, Cai G, Liang X and Liu J: Targeted deletion of
miR-139-5p activates MAPK, NF-κB and STAT3 signaling and promotes
intestinal inflammation and colorectal cancer. FEBS J.
283:1438–1452. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bai Y, Qiao L, Xie N, Shi Y, Liu N and
Wang J: Expression and prognosis analyses of the Tob/BTG
antiproliferative (APRO) protein family in human cancers. PLoS One.
12:e01849022017. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Krek A, Grün D, Poy MN, Wolf R, Rosenberg
L, Epstein EJ, MacMenamin P, da Piedade I, Gunsalus KC, Stoffel M
and Rajewsky N: Combinatorial microRNA target predictions. Nat
Genet. 37:495–500. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
41
|
Bandyopadhyay S and Mitra R: TargetMiner:
microRNA target prediction with systematic identification of
tissue-specific negative examples. Bioinformatics. 25:2625–2631.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Xu W, San Lucas A, Wang Z and Liu Y:
Identifying microRNA targets in different gene regions. BMC
Bioinformatics. 15 (Suppl 7):S42014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Ferrer RR, Ramirez M, Beckman LJ, Danao LL
and Ashing-Giwa KT: The impact of cultural characteristics on
colorectal cancer screening adherence among Filipinos in the United
States: A pilot study. Psychooncology. 20:862–870. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Weitz J, Koch M, Debus J, Höhler T, Galle
PR and Büchler MW: Colorectal cancer. Lancet. 365:1066. 2005.
View Article : Google Scholar
|
|
45
|
Wolpin BM and Mayer RJ: Systemic treatment
of colorectal cancer. Gastroenterology. 134:1296–1310. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhou S, Huang Q, Zheng S, Lin K, You J and
Zhang X: miR-27a regulates the sensitivity of breast cancer cells
to cisplatin treatment via BAK-SMAC/DIABLO-XIAP axis. Tumour Biol.
37:6837–6845. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lin-Lin XU, et al: Effects of genistein on
the growth of ovarian cancer cell SKOV3 by regulating miR-27a and
target gene expression. Chin J Clin Pharmacol Ther. 17:1321–1326.
2012.(In Chinese).
|
|
48
|
Ma Y, Yu S, Zhao W, Lu Z and Chen J:
miR-27a regulates the growth, colony formation and migration of
pancreatic cancer cells by targeting Sprouty2. Cancer Lett.
298:150–158. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Choo KB, Soon YL, Nguyen PN, Hiew MS and
Huang CJ: MicroRNA-5p and −3p co-expression and cross-targeting in
colon cancer cells. J Biomed Sci. 21:952014. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zeng G, Xun W, Wei K, Yang Y and Shen H:
MicroRNA-27a-3p regulates epithelial to mesenchymal transition via
targeting YAP1 in oral squamous cell carcinoma cells. Oncol Rep.
36:1475–1482. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Zhao Y, Wang P, Meng J, Ji Y, Xu D, Chen
T, Fan R, Yu X, Yao J and Dong C: MicroRNA-27a-3p inhibits
melanogenesis in mouse skin melanocytes by targeting Wnt3a. Int J
Mol Sci. 16:10921–10933. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Prévôt D, Voeltzel T, Birot AM, Morel AP,
Rostan MC, Magaud JP and Corbo L: The Leukemia-associated Protein
Btg1 and the p53-regulated Protein Btg2 Interact with the
Homeoprotein Hoxb9 and enhance its transcriptional activation. J
Biol Chem. 275:147–153. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Corjay MH, Kearney MA, Munzer DA, Diamond
SM and Stoltenborg JK: Antiproliferative gene BTG1 is highly
expressed in apoptotic cells in macrophage-rich areas of advanced
lesions in Watanabe heritable hyperlipidemic rabbit and human. Lab
Invest. 78:847–858. 1998.PubMed/NCBI
|
|
54
|
Sun G, Liu Q, Cheng Y and Hu W: B cell
translocation gene 1 reduces the biological outcome of kidney
cancer through induction of cell proliferation, cell cycle arrest,
cell apoptosis and cell metastasis. Int J Mol Med. 35:777–8348.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Bretones G, Delgado MD and León J: Myc and
cell cycle control. Biochim Biophys Acta. 1849:506–516. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Sun GG, Wang YD, Cheng YJ and Hu WN: The
expression of BTG1 is downregulated in nasopharyngeal carcinoma and
possibly associated with tumour metastasis. Mol Biol Rep.
41:5979–5988. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Sun GG, Wang YD, Cheng YJ and Hu WN: BTG1
underexpression is an independent prognostic marker in esophageal
squamous cell carcinoma. Tumour Biol. 35:9707–9716. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Lu YF, Sun GG, Liu Q, Yang CR and Cheng
YJ: BTG1 expression in thyroid carcinoma: Diagnostic indicator and
prognostic marker. Int J Oncol. 45:1574–1582. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Zhao Y, Gou WF, Chen S, Takano Y and Zheng
HC: BTG1 expression correlates with pathogenesis and progression of
ovarian carcinomas. Int J Mol Sci. 14:19670–19680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Smolewski P and Robak T: Inhibitors of
apoptosis proteins (IAPs) as potential molecular targets for
therapy of hematological malignancies. Curr Mol Med. 11:633–649.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Nicholson DW and Thornberry NA: Trends
Biochem. Sci. 22:299–306. 1997.
|
|
62
|
Salvesin GS and Dixit VM: Caspases:
Intracellular signaling by proteolysis. Cell. 91:443–446. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shin S, Sung BJ, Cho YS, Kim HJ, Ha NC,
Hwang JI, Chung CW, Jung YK and Oh BH: An anti apoptotic protein
human survivin is a direct inhibitor of caspase-3 and −7.
Biochemistry. 40:1117–1123. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Nossa CW, Jain P, Tamilselvam B, Gupta VR,
Chen LF, Schreiber V, Desnoyers S and Blanke SR: Activation of the
abundant nuclear factor poly(ADP-ribose) polymerase-1 by
Helicobacter pylori. Proc Natl Acad Sci USA. 106:19998–20003. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Li Y, Zhang Y, Xiao S, Kong P, Cheng C,
Shi R, Wang F, Zhang L, Wang J, Jia Z, et al: Mps1 is associated
with the BRAFV600E mutation but does not rely on the classic
RAS/RAF/MEK/ERK signaling pathway in thyroid carcinoma. Oncol Lett.
15:9978–9986. 2018.PubMed/NCBI
|
|
66
|
Cristea S and Sage J: Is the Canonical
RAF/MEK/ERK signaling pathway a therapeutic target in SCLC? J
Thorac Oncol. 11:1233–1241. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Zhou K, Luo X, Wang Y, Cao D and Sun G:
MicroRNA-30a suppresses tumor progression by blocking
Ras/Raf/MEK/ERK signaling pathway in hepatocellular carcinoma.
Biomed Pharmacother. 93:1025–1032. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Rapp UR, Goldsborough MD, Mark GE, Bonner
TI, Groffen J, Reynolds FH Jr and Stephenson JR: Stephenson
Structure and biological activity of v-raf, a unique oncogene
transduced by a retrovirus Proc. Natl Acad Sci. 80:4218–4222. 1983.
View Article : Google Scholar
|
|
69
|
Montagut C and Settleman J: Targeting the
RAF-MEK-ERK pathway in cancer therapy. Cancer Lett. 283:125–134.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Steelman LS, Chappell WH, Abrams SL, Kempf
RC, Long J, Laidler P, Mijatovic S, Maksimovic-Ivanic D, Stivala F,
Mazzarino MC, et al: Roles of the Raf/MEK/ERK and
PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity
to therapy-implications for cancer and aging. Aging (Albany NY).
3:192–222. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Zhang H, Tang J, Li C, Kong J, Wang J, Wu
Y, Xu E and Lai M: MiR-22 regulates 5-FU sensitivity by inhibiting
autophagy and promoting apoptosis in colorectal cancer cells.
Cancer Lett. 356:781–790. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Weng W, Liu N, Toiyama Y, Kusunoki M,
Nagasaka T, Fujiwara T, Wei Q, Qin H, Lin H, Ma Y and Goel A: Novel
evidence for a PIWI-interacting RNA (piRNA) as an oncogenic
mediator of disease progression, and a potential prognostic
biomarker in colorectal cancer. Mol Cancer. 17:162018. View Article : Google Scholar : PubMed/NCBI
|