Introduction
Pancreatic cancer is one of the top five causes of
cancer-associated mortality with a fatality rate of 95%; the
majority of patients with pancreatic cancer have a delayed
diagnosis, as early diagnosis is difficult (1). Recent data showed that pancreatic
cancer has the lowest 5-year relative survival rate (9%) among all
cancer types (2). Pancreatic ductal
adenocarcinoma (PDAC) accounts for a majority of pancreatic cancer
cases and is an aggressive and difficult malignancy to treat
(3). Despite advancing knowledge of
the mechanisms of carcinogenicity and improvement in diagnosis and
management, the prognosis remains poor for patients with PDAC
(3). Therefore, new mechanisms and
biomarkers are needed to improve the prognosis and detection of
PDAC in patients.
MicroRNAs (miRNAs) are a class of small non-coding
RNAs (ncRNAs), 18–25 nucleotides long, that negatively control gene
expression at the mRNA and protein level (4). Accumulating evidence has suggested that
miRNAs are dysregulated in various types of cancers, including PDAC
(5). As the absence of symptoms in
early disease results in a late diagnosis of PDAC, numerous miRNAs,
such as miR-21, miR-155, miR-196 and miR-210, have been identified
as biomarkers for PDAC (5).
Understanding the mechanisms contributing to miRNA dysregulation
may help to improve PDAC diagnosis and treatment.
Another class of ncRNAs are circular RNAs
(circRNAs), which function as miRNA sponges, thus serving as
competitive inhibitors and suppressing the ability of the miRNA to
bind its mRNA targets (6,7). Although circRNAs were discovered in RNA
viruses in the 1970s, these molecules were considered as molecular
flukes or products of aberrant RNA splicing (7). Recent evidence indicates that circRNAs
may serve important roles in cancers, including bladder cancer
(8), esophageal squamous cell
carcinoma (9), breast cancer
(10), basal cell carcinoma
(11), cutaneous squamous cell
carcinoma (12), colorectal cancer
(13) and PDAC (14). In the present study, microarray data
downloaded from Gene Expression Omnibus (GEO) were used to explore
the roles of circRNAs and miRNAs in PDAC and to provide novel
insights into PDAC biology.
Materials and methods
Microarray data
Raw gene expression profile datasets GSE69362,
GSE79634 and GSE43796 were downloaded from the GEO database
(http://www.ncbi.nlm.nih.gov/geo). The
datasets GSE69362 and GSE79634 were analyzed on the GPL19978
Agilent-069978 Arraystar Human CircRNA microarray V1 platform,
which analyzes 5,396 human circRNAs at the same time. The dataset
GSE43796 was analyzed on the GPL15159 Agilent-031181 Unrestricted
Human miRNA V16.0 Microarray platform, which can detect 1,205
mature human miRNAs.
The GSE69362 dataset accessed the expression of
circRNAs in six pairs of PDAC and adjacent normal tissues. The
GSE79634 dataset accessed the expression of circRNAs in 20 pairs of
samples. GSE43796 detected the expression of miRNAs in
solid-pseudopapillary neoplasm of pancreas (n=14), ductal
adenocarcinoma (n=6), neuroendocrine tumor (n=6) and non-neoplastic
pancreas (n=5). The six PDAC tissues and the five non-neoplastic
pancreases from GSE43796 were included in the present study.
Data analysis
Differentially expressed genes were identified using
the limma package in the R software (version 3.3.2; http://www.r-project.org) (15). Raw data was read and background was
corrected and normalized. Probes that were ≥10% brighter compared
with the negative controls on at least one-half of arrays were
averaged for each gene. The array weights were estimated and used
in the linear model analysis. Paired or unpaired Student's t-test
was used to identify differentially expressed genes between tumor
tissues and normal tissues. The Benjamini-Hochberg method was used
to correct for multiple comparisons (16). Differentially expressed genes between
tumor tissues and normal tissues were identified, using fold-change
>2 and adjusted P<0.05 as selection criteria.
miRNA prediction
As certain sequences of circRNAs in circBase may
differ from the mature sequence of circRNAs (17), sequences of the circRNAs were
extracted using circPrimer (version 1.2; http://www.bioinf.com.cn) (18), which was developed by our laboratory.
As exon-derived circRNAs have been demonstrated to serve as miRNA
sponges, the 256 differentially expressed circRNAs were annotated
using circPrimer and only 214 exon-derived circRNAs were used for
further analysis.
The sequences of the 214 circRNAs and 20
differentially expressed miRNAs were saved as FASTA format.
circRNA/miRNA interactions were predicted using circMir (version
1.0; http://www.bioinf.com.cn), which used
the miRanda (19) and RNAhybrid
(20) algorithms to predict miRNAs
that bind to spliced junctions of circRNAs. As the prediction
algorithm often suffers from high false positive rates, only the
circRNA/miRNA interactions predicted by the two independent
algorithms were used. Cytoscape software version 3.1.1 (http://cytoscape.org) was utilized to construct a
possible functional network of the differentially expressed
circRNAs and miRNAs.
Pathway analysis of circRNA gene
symbols
circRNA host genes were submitted to the web-based
tool DAVID (http://david.abcc.ncifcrf.gov) to perform a Kyoto
Encyclopedia of Genes and Genomes (KEGG) pathway enrichment
analysis. The following default thresholds were used: Count=2 and
EASE (a modified Fisher's Exact test P-value)=0.1. The enriched
pathways and gene symbols were imported into Cytoscape software
version 3.1.1 to construct a network.
Results
Overview of the datasets
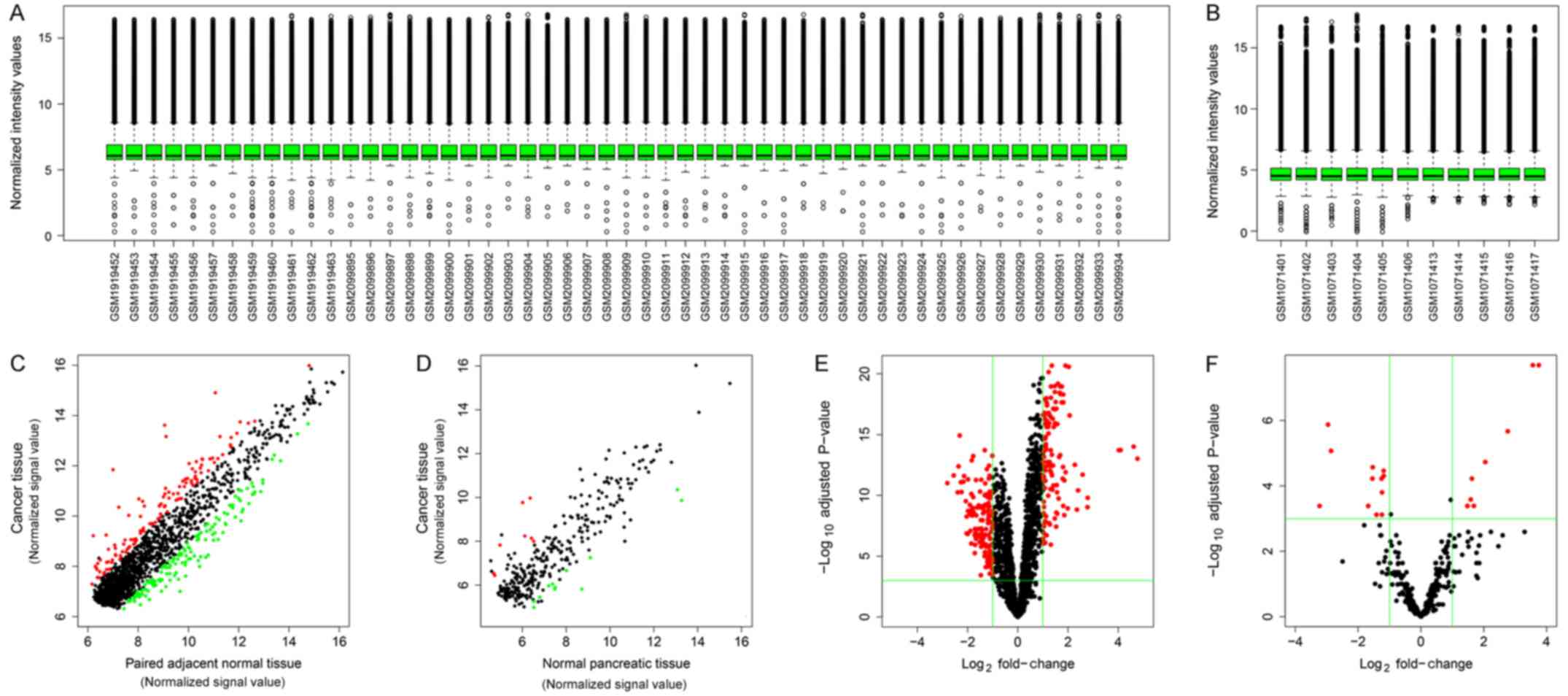
The distribution of normalized signal intensities of
the datasets indicated that microarray quality for circRNA and
miRNA was good (Fig. 1A and B,
respectively). The expression levels of the majority of genes were
correlated between PDAC and normal tissues, although there were a
number of differentially expressed circRNAs and miRNAs (Fig. 1C and D, respectively). Volcano plots
were generated to visualize the relationship of fold-change and
statistical significance for circRNAs and miRNAs (Fig. 1E and F, respectively).
Identification of differentially
expressed genes
Compared with adjacent normal tissues, there were
256 differentially expressed circRNAs in PDAC tissues, including
115 upregulated circRNAs and 141 downregulated circRNAs. The top 40
differentially expressed circRNAs in PDAC are presented in Table I, and the full list is presented in
Table SI. Eight upregulated miRNAs
and 12 downregulated miRNAs were identified in PDAC (Table II).
 | Table I.Top 40 differentially expressed
circRNAs in pancreatic ductal adenocarcinoma. |
Table I.
Top 40 differentially expressed
circRNAs in pancreatic ductal adenocarcinoma.
| A, Upregulated
circRNAs |
|---|
|
|---|
| circRNA | Host gene | Fold change | P-value | Adjusted
P-valuea |
|---|
|
hsa_circ_0000977 | NOL10 | 27.155 |
1.498×10−7 |
2.223×10−6 |
|
hsa_circ_0006220 | TADA2A | 24.448 |
3.806×10−8 |
8.269×10−7 |
|
hsa_circ_0001666 | FAM120B | 17.041 |
5.381×10−8 |
1.096×10−6 |
|
hsa_circ_0043278 | TADA2A | 16.234 |
5.970×10−8 |
1.110×10−6 |
|
hsa_circ_0078297 | MTHFD1L | 6.870 |
9.875×10−6 |
5.566×10−5 |
|
hsa_circ_0013912 | POLR3C | 6.834 |
2.484×10−5 |
1.198×10−4 |
|
hsa_circ_0003600 | SOX13 | 5.984 |
8.372×10−7 |
8.265×10−6 |
|
hsa_circ_0044436 | KAT7 | 5.272 |
4.677×10−6 |
3.019×10−5 |
|
hsa_circ_0018909 | VDAC2 | 5.108 |
3.209×10−5 |
1.463×10−4 |
|
hsa_circ_0029634 | ZMYM2 | 4.829 |
2.919×10−7 |
3.679×10−6 |
|
hsa_circ_0001907 | LOC100507412 | 4.182 |
1.461×10−9 |
6.333×10−8 |
|
hsa_circ_0066147 | SFMBT1 | 4.079 |
2.067×10−12 |
1.165×10−9 |
|
hsa_circ_0092314 | RANBP1 | 3.967 |
2.826×10−5 |
1.317×10−4 |
|
hsa_circ_0050102 | PGPEP1 | 3.741 |
6.531×10−7 |
6.735×10−6 |
|
hsa_circ_0006117 | PTPRA | 3.722 |
1.245×10−12 |
1.053×10−9 |
|
hsa_circ_0000912 | FCHO1 | 3.613 |
1.603×10−6 |
1.327×10−5 |
|
hsa_circ_0080210 | GRB10 | 3.607 |
5.627×10−5 |
2.310×10−4 |
|
hsa_circ_0030292 | FAM124A | 3.515 |
3.366×10−10 |
2.159×10−8 |
|
hsa_circ_0082452 | EXOC4 | 3.503 |
1.274×10−10 |
1.134×10−8 |
|
hsa_circ_0005105 | SEC24A | 3.484 |
3.878×10−7 |
4.401×10−6 |
|
| B, Downregulated
circRNAs |
|
| circRNA | Host
gene | Fold
change | P-value | Adjusted
P-valuea |
|
|
hsa_circ_0000518 | RPPH1 | 6.986 |
2.148×10−6 |
1.649×10−5 |
|
hsa_circ_0005556 | NBAS | 5.853 |
9.288×10−7 |
8.823×10−6 |
|
hsa_circ_0005918 | FCHSD2 | 5.249 |
5.710×10−6 |
3.537×10−5 |
|
hsa_circ_0081188 | SLC25A13 | 5.129 |
3.563×10−7 |
4.156×10−6 |
|
hsa_circ_0013587 | LRIG2 | 4.969 |
1.231×10−8 |
3.304×10−7 |
|
hsa_circ_0006110 | USP34 | 4.813 |
1.952×10−6 |
1.536×10−5 |
|
hsa_circ_0005273 | PTK2 | 4.716 |
5.497×10−6 |
3.455×10−5 |
|
hsa_circ_0003930 | GGNBP2 | 4.478 |
1.848×10−6 |
1.474×10−5 |
|
hsa_circ_0000511 | RPPH1 | 4.398 |
9.923×10−7 |
9.271×10−6 |
|
hsa_circ_0000517 | RPPH1 | 4.193 |
1.349×10−5 |
7.288×10−5 |
|
hsa_circ_0005394 | ZC3H7A | 4.102 |
6.545×10−5 |
2.594×10−4 |
|
hsa_circ_0092367 | SNORD116-14 | 4.099 |
3.443×10−4 |
1.036×10−3 |
|
hsa_circ_0000520 | RPPH1 | 3.981 |
3.628×10−4 |
1.082×10−3 |
|
hsa_circ_0003831 | PPP2R5C | 3.950 |
1.410×10−6 |
1.210×10−5 |
|
hsa_circ_0006944 | GTF2I | 3.746 |
5.175×10−5 |
2.172×10−4 |
|
hsa_circ_0061749 | BRWD1 | 3.745 |
1.933×10−5 |
9.955×10−5 |
|
hsa_circ_0000958 | PPP1R12C | 3.704 |
3.575×10−5 |
1.603×10−4 |
|
hsa_circ_0000662 | AXIN1 | 3.700 |
2.699×10−4 |
8.420×10−4 |
|
hsa_circ_0079385 | ZDHHC4 | 3.605 |
9.123×10−4 |
2.374×10−3 |
|
hsa_circ_0001288 | SETD2 | 3.600 |
6.598×10−6 |
4.043×10−5 |
| Table II.Differentially expressed miRNAs in
pancreatic ductal adenocarcinoma. |
Table II.
Differentially expressed miRNAs in
pancreatic ductal adenocarcinoma.
| A, Upregulated
miRNAs |
|---|
|
|---|
| miRNA | Fold change | P-value | Adjusted
P-valuea |
|---|
| hsa-miR-210-3p | 13.480 |
2.649×10−6 |
4.584×10−4 |
|
hsa-miR-135b-5p | 11.830 |
2.289×10−6 |
4.584×10−4 |
|
hsa-miR-203a-3p | 6.798 |
3.989×10−5 |
3.450×10−3 |
| hsa-miR-222-3p | 4.139 |
1.787×10−4 |
8.834×10−3 |
|
hsa-miR-196b-5p | 3.216 |
1.923×10−3 |
3.379×10−2 |
|
hsa-miR-200a-5p | 3.080 |
5.337×10−4 |
1.467×10−2 |
| hsa-miR-221-3p | 3.003 |
1.204×10−3 |
2.777×10−2 |
|
hsa-miR-181b-5p | 2.791 |
1.953×10−3 |
3.379×10−2 |
|
| B, Downregulated
miRNAs |
|
| miRNA | Fold
change | P-value | Adjusted
P-valuea |
|
|
hsa-miR-148a-3p | 9.347 |
1.815×10−3 |
3.379×10−2 |
| hsa-miR-144-3p | 7.768 |
2.443×10−5 |
2.818×10−3 |
| hsa-miR-451a | 7.255 |
1.090×10−4 |
6.285×10−3 |
|
hsa-miR-376c-3p | 3.198 |
1.902×10−3 |
3.379×10−2 |
| hsa-miR-204-5p | 2.916 |
4.908×10−4 |
1.467×10−2 |
| hsa-miR-154-5p | 2.897 |
2.396×10−4 |
1.036×10−2 |
| hsa-miR-381-3p | 2.665 |
2.865×10−3 |
4.425×10−2 |
| hsa-miR-654-3p | 2.382 |
5.511×10−4 |
1.467×10−2 |
| hsa-miR-377-3p | 2.365 |
2.942×10−3 |
4.425×10−2 |
| hsa-miR-136-3p | 2.359 |
9.045×10−4 |
2.235×10−2 |
| hsa-miR-557 | 2.275 |
3.002×10−4 |
1.154×10−2 |
|
hsa-miR-487b-3p | 2.271 |
3.903×10−4 |
1.350×10−2 |
Prediction of circRNA/miRNA
interaction
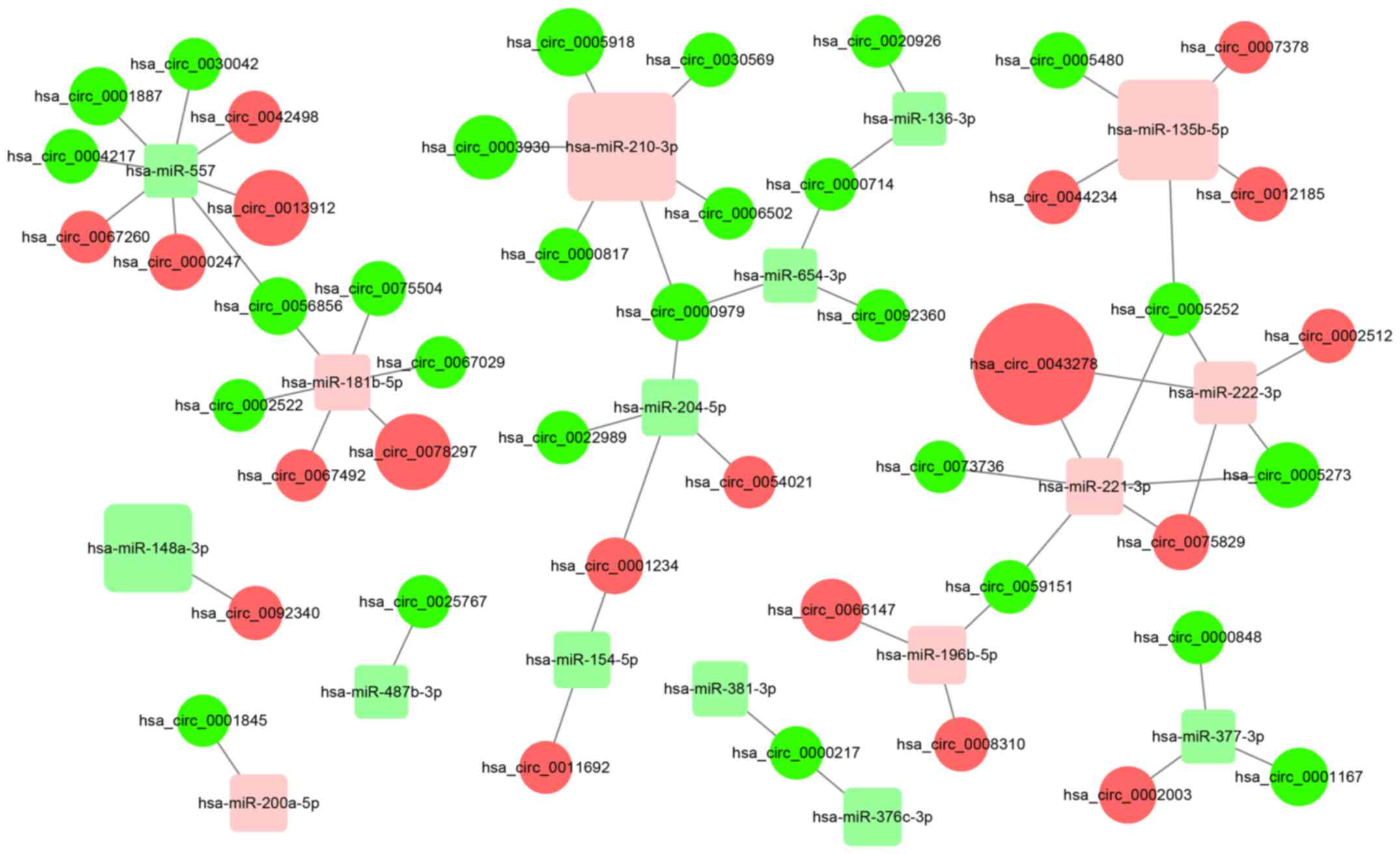
circRNAs may upregulate the expression of target
genes of miRNAs by sequestering the miRNAs. In the present study, a
total of 51 circRNA/miRNA interactions were identified and a
possible functional network of the differentially expressed
circRNAs and miRNAs was generated (Fig.
2). The detailed results including miRNA binding sites are
presented in Table SII.
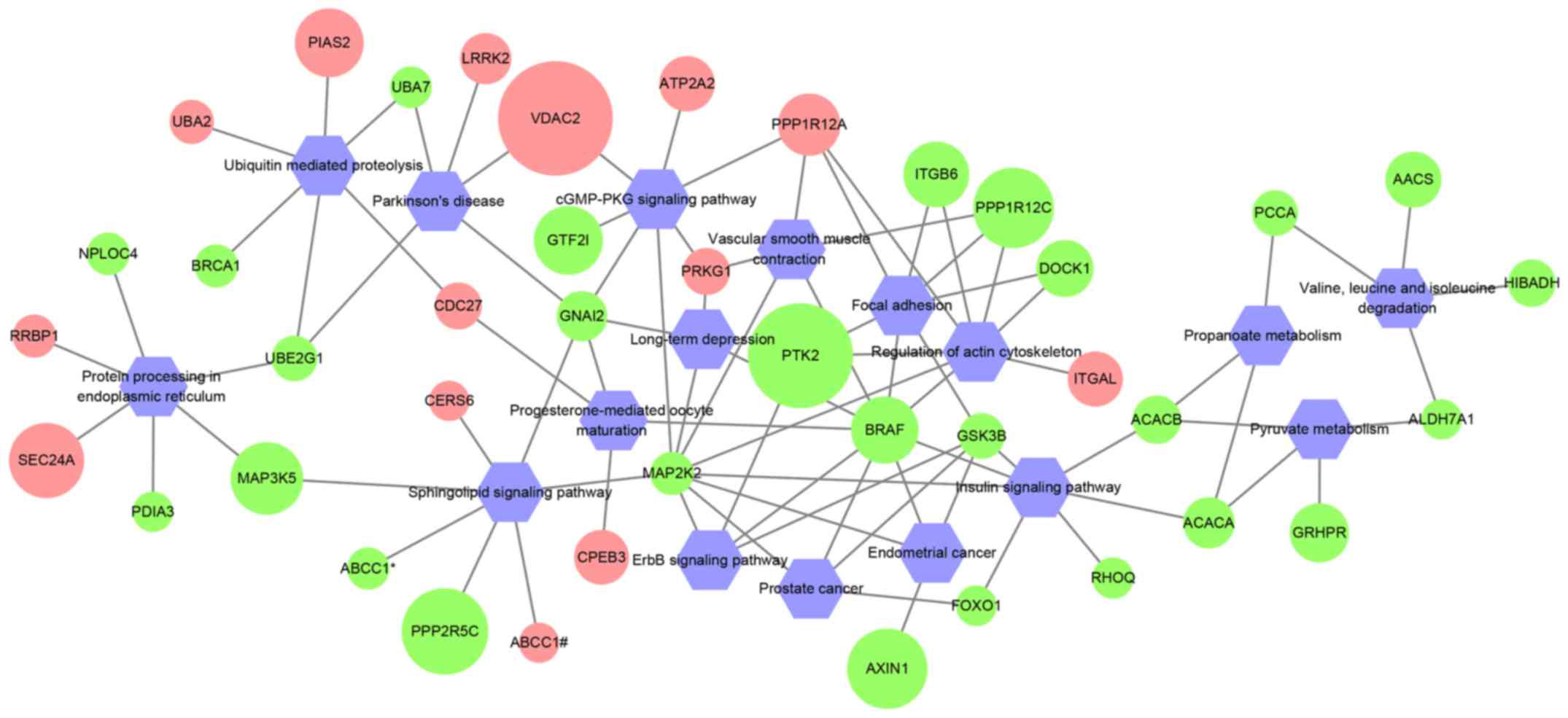
KEGG pathway analysis
Previous studies have shown that circRNAs modulate
the expression of their host genes (6,7). Thus,
KEGG pathway enrichment analysis was performed using host genes of
differentially expressed circRNAs. A total of 41 host genes were
enriched in 17 pathways (Fig. 3;
Table SIII). B-Raf proto-oncogene,
serine/threonine kinase (BRAF) and mitogen-activated protein kinase
kinase 2 (MAP2K2) interacted with the highest number of pathways,
which suggested that these two genes may serve important roles in
PDAC.
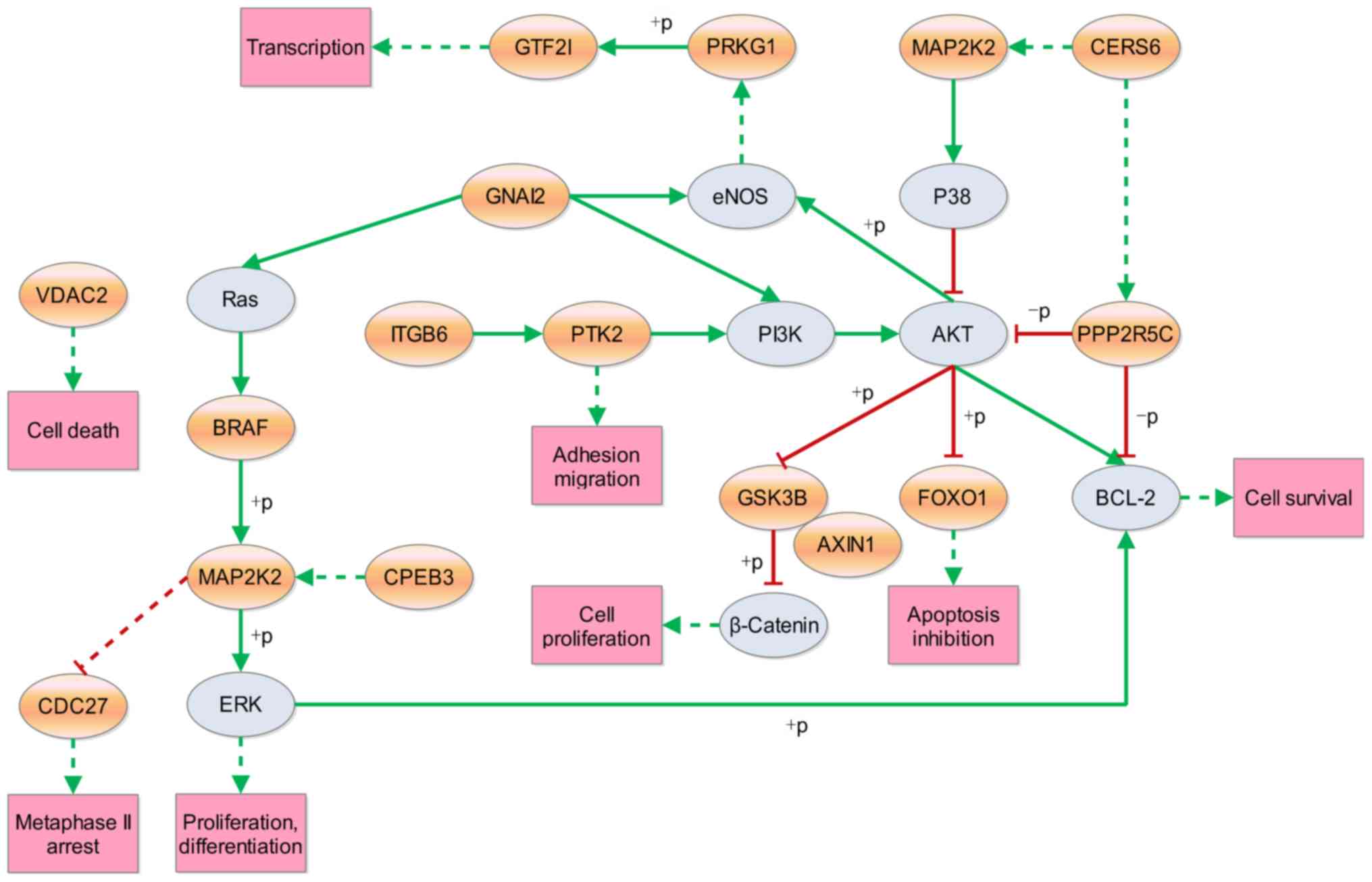
Since apoptosis and proliferation are associated
with cancer development and progression, the subnetworks of these
pathways were extracted from the 17 enriched KEGG pathways.
Subsequently, a new network was constructed with these extracted
subnetworks, which included 16 host genes that potentially affect
cell survival directly or indirectly (Fig. 4). Thus, target circRNAs were obtained
for future studies.
Discussion
Two former studies from one research group have
profiled the expression of circRNAs in PDAC (14,21).
However, only six pairs of PDAC and adjacent normal tissues were
profiled and the sample size was relatively small. In the present
study, comprehensive analysis with 26 samples from GEO was
performed to explore the putative roles of circRNAs in PDAC and to
provide novel insights into PDAC biology. The regulatory roles of
circRNAs in cancer are attracting increasing attention. Li et
al (14) have identified a
number of dysregulated circRNAs in PDAC. Another study further
investigated the expression level of one dysregulated circRNA
termed circ-LDLRAD3, which was upregulated in pancreatic cancer
cell lines as well as tumors and plasma from patients with
pancreatic cancer, and circ-LDLRAD3 was demonstrated to be
associated with invasion and metastasis of pancreatic cancer
(22). circRNA exerts its regulatory
roles through sequestering cancer-related miRNAs (13,23–26).
Therefore, differentially expressed circRNAs and miRNAs in PDAC
were used to predict circRNA/miRNA interactions. circRNAs may
upregulate the expression of target genes of miRNAs by sequestering
the miRNAs. In addition, if circRNAs and their host genes share the
same miRNA binding sites, the circRNAs may upregulate the
expression of their host genes. In the present study, a total of 51
circRNA/miRNA interactions were identified using 256 differentially
expressed circRNAs and 20 differentially expressed miRNAs and a
network of the circRNA/miRNA interactions was produced to
illustrate the relationships of these genes. The network contained
41 circRNA nodes, 14 miRNA nodes and 51 edges. The number of
connections indicates gene involvement in various pathways in PDAC;
therefore, the nodes connecting multiple genes may be worth further
investigation using experimental methods.
circRNAs are encoded in exons and/or introns of
their parental genes (27). Previous
studies have shown that circRNAs modulate the expression of their
parental genes (6,7,28,29). To
explore the potential roles of circRNAs, the predominant pathways
of the gene symbols of the differentially expressed circRNAs were
identified by a pathway-mapping tool. BRAF and MAP2K2 interacted
with the highest number of pathways. BRAF and MAP2K2 are two
important members of MAPK signaling pathway, which is associated
with PDAC (30,31). The results of the present study
indicated that the majority of the inspected genes achieved their
effects through the PI3K/AKT signaling pathway. Pancreatic
acinar-to-ductal metaplasia is an initiating event that is induced
through the PI3K/AKT signaling pathway and can progress to PDAC
(32). Another pathway involving
cell proliferation is the Wnt/β-catenin signaling pathway. Sox15
exerts its tumor-suppressive effects in pancreatic cancer by
suppressing the Wnt/β-catenin signaling pathway (33). Recent data has suggested that the
WNT/β-catenin signaling pathway upregulated the expression of
cellular communication network factor 1 and consequently enhanced
pancreatic cancer development and malignancy (34). In the present study, the aim was to
identify circRNA-associated pathways that are involved in the
development of PDAC.
As well as acting as miRNA sponges, circRNAs may
exert their function through protein binding (35), protein coding (36,37),
modulating transcriptional activity of RNA polymerase II (38) and competing with linear splicing
(39). Cellular localization has
been linked with physiological function and functional mechanism of
circRNAs (40,41). Most circRNAs comprise of exonic
sequences and are located in the cytoplasm, whereas a small number
of intron-derived circRNAs are located in the nucleus (42). Therefore, it is necessary to confirm
the cellular localization of a circRNA before studying its
roles.
The potential limitations of the present study need
to be considered when interpreting the results. Firstly, the
results were not verified in clinical samples; differentially
expressed circRNAs and miRNAs may be verified using reverse
transcription-quantitative PCR in larger clinical samples.
Secondly, the circRNA/miRNA interactions were predicted using a
bioinformatics method and should be confirmed by the luciferase
reporter assay. Thirdly, host gene symbols of circRNAs were used to
carry out the KEGG pathway enrichment analysis; whether these
circRNAs exert an effect in the enriched pathways should be
confirmed using experimental methods.
In conclusion, the potential roles of circRNAs in
PDAC were explored using bioinformatics. In total, 256
differentially expressed circRNAs and 20 differentially expressed
miRNAs were identified in PDAC tissues compared with normal
tissues. The roles of circRNAs in PDAC were explored by prediction
of circRNA/miRNA interactions and KEGG pathway enrichment analysis.
The circRNA-associated pathways presented in present study could
help uncover the effect of circRNAs on PDAC development. Further
study confirming the effects of these circRNAs on PDAC development
are required in order to improve PDAC diagnosis and therapy.
Supplementary Material
Supporting Data
Acknowledgements
Not applicable
Funding
This study was funded by The National Natural
Science Foundation of China (grant no. 81602551) and the young
talents program of Jiangsu Cancer Hospital (grant no.
2017YQL-10).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
QZ and JYW interpreted the data and wrote the paper.
SLZ, SYZ and SJY collected the data and conducted the analysis. SLZ
designed this study and revised the manuscript. All authors read
and approved the final manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Wachsmann MB, Pop LM and Vitetta ES:
Pancreatic ductal adenocarcinoma: A review of immunologic aspects.
J Investig Med. 60:643–663. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2019. CA Cancer J Clin. 69:7–34. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Stark A and Eibl G: Pancreatic ductal
adenocarcinoma. Pancreapedia: Exocrine Pancreas Knowledge Base,
Version 1.0. 2015.https://www.pancreapedia.org/reviews/pancreatic-ductal-adenocarcinomaMay
23–2015
|
|
4
|
He L and Hannon GJ: MicroRNAs: Small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hernandez YG and Lucas AL: MicroRNA in
pancreatic ductal adenocarcinoma and its precursor lesions. World J
Gastrointest Oncol. 8:18–29. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li J, Yang J, Zhou P, Le Y, Zhou C, Wang
S, Xu D, Lin HK and Gong Z: Circular RNAs in cancer: Novel insights
into origins, properties, functions and implications. Am J Cancer
Res. 5:472–480. 2015.PubMed/NCBI
|
|
7
|
Qu S, Yang X, Li X, Wang J, Gao Y, Shang
R, Sun W, Dou K and Li H: Circular RNA: A new star of noncoding
RNAs. Cancer Lett. 365:141–148. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhong Z, Lv M and Chen J: Screening
differential circular RNA expression profiles reveals the
regulatory role of circTCF25-miR-103a-3p/miR-107-CDK6 pathway in
bladder carcinoma. Sci Rep. 6:309192016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xia W, Qiu M, Chen R, Wang S, Leng X, Wang
J, Xu Y, Hu J, Dong G, Xu PL and Yin R: Circular RNA
has_circ_0067934 is upregulated in esophageal squamous cell
carcinoma and promoted proliferation. Sci Rep. 6:355762016.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nair AA, Niu N, Tang X, Thompson KJ, Wang
L, Kocher JP, Subramanian S and Kalari KR: Circular RNAs and their
associations with breast cancer subtypes. Oncotarget.
7:80967–80979. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sand M, Bechara FG, Sand D, Gambichler T,
Hahn SA, Bromba M, Stockfleth E and Hessam S: Circular RNA
expression in basal cell carcinoma. Epigenomics. 8:619–632. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Sand M, Bechara FG, Gambichler T, Sand D,
Bromba M, Hahn SA, Stockfleth E and Hessam S: Circular RNA
expression in cutaneous squamous cell carcinoma. J Dermatol Sci.
83:210–218. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Xie H, Ren X, Xin S, Lan X, Lu G, Lin Y,
Yang S, Zeng Z, Liao W, Ding YQ and Liang L: Emerging roles of
circRNA_001569 targeting miR-145 in the proliferation and invasion
of colorectal cancer. Oncotarget. 7:26680–26691. 2016.PubMed/NCBI
|
|
14
|
Li H, Hao X, Wang H, Liu Z, He Y, Pu M,
Zhang H, Yu H, Duan J and Qu S: Circular RNA expression profile of
pancreatic ductal adenocarcinoma revealed by microarray. Cell
Physiol Biochem. 40:1334–1344. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Ritchie ME, Phipson B, Wu D, Hu Y, Law CW,
Shi W and Smyth GK: Limma powers differential expression analyses
for RNA-sequencing and microarray studies. Nucleic Acids Res.
43:e472015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Benjamini Y, Drai D, Elmer G, Kafkafi N
and Golani I: Controlling the false discovery rate in behavior
genetics research. Behav Brain Res. 125:279–284. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yang Y, Gao X, Zhang M, Yan S, Sun C, Xiao
F, Huang N, Yang X, Zhao K, Zhou H, et al: Novel role of FBXW7
circular RNA in repressing glioma tumorigenesis. J Natl Cancer
Inst. 110:2018. View Article : Google Scholar
|
|
18
|
Zhong S, Wang J, Zhang Q, Xu H and Feng J:
CircPrimer: A software for annotating circRNAs and determining the
specificity of circRNA primers. BMC Bioinformatics. 19:2922018.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Enright AJ, John B, Gaul U, Tuschl T,
Sander C and Marks DS: MicroRNA targets in Drosophila.
Genome Biol. 5:R12003. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Krüger J and Rehmsmeier M: RNAhybrid:
microRNA target prediction easy, fast and flexible. Nucleic Acids
Res 34 (Web Server Issue). W451–W454. 2006. View Article : Google Scholar
|
|
21
|
Qu S, Song W, Yang X, Wang J, Zhang R,
Zhang Z, Zhang H and Li H: Microarray expression profile of
circular RNAs in human pancreatic ductal adenocarcinoma. Genom
Data. 5:385–387. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Yang F, Liu DY, Guo JT, Ge N, Zhu P, Liu
X, Wang S, Wang GX and Sun SY: Circular RNA circ-LDLRAD3 as a
biomarker in diagnosis of pancreatic cancer. World J Gastroenterol.
23:8345–8354. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zheng Q, Bao C, Guo W, Li S, Chen J, Chen
B, Luo Y, Lyu D, Li Y, Shi G, et al: Circular RNA profiling reveals
an abundant circHIPK3 that regulates cell growth by sponging
multiple miRNAs. Nat Commun. 7:112152016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xu H, Guo S, Li W and Yu P: The circular
RNA Cdr1as, via miR-7 and its targets, regulates insulin
transcription and secretion in islet cells. Sci Rep. 5:124532015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Nan A, Chen L, Zhang N, Liu Z, Yang T,
Wang Z, Yang C and Jiang Y: A novel regulatory network among
LncRpa, CircRar1, MiR-671 and apoptotic genes promotes lead-induced
neuronal cell apoptosis. Arch Toxicol. 91:1671–1684. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hansen TB, Jensen TI, Clausen BH, Bramsen
JB, Finsen B, Damgaard CK and Kjems J: Natural RNA circles function
as efficient microRNA sponges. Nature. 495:384–388. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen LL and Yang L: Regulation of circRNA
biogenesis. RNA Biol. 12:381–388. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
He J, Xie Q, Xu H, Li J and Li Y: Circular
RNAs and cancer. Cancer Lett. 396:138–144. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Li Z, Huang C, Bao C, Chen L, Lin M, Wang
X, Zhong G, Yu B, Hu W, Dai L, et al: Exon-intron circular RNAs
regulate transcription in the nucleus. Nat Struct Mol Biol.
22:256–264. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Papademetrio DL, Lompardía SL, Simunovich
T, Costantino S, Mihalez CY, Cavaliere V and Álvarez É: Inhibition
of survival pathways MAPK and NF-κB triggers apoptosis in
pancreatic ductal adenocarcinoma cells via suppression of
autophagy. Target Oncol. 11:183–195. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Herner A, Sauliunaite D, Michalski CW,
Erkan M, De Oliveira T, Abiatari I, Kong B, Esposito I, Friess H
and Kleeff J: Glutamate increases pancreatic cancer cell invasion
and migration via AMPA receptor activation and Kras-MAPK signaling.
Int J Cancer. 129:2349–2359. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen Y, Bai X, Zhang Q, Wen L, Su W, Fu Q,
Sun X, Lou Y, Yang J, Zhang J, et al: The hepatitis B virus X
protein promotes pancreatic cancer through modulation of the
PI3K/AKT signaling pathway. Cancer Lett. 380:98–105. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Thu KL, Radulovich N, Becker-Santos DD,
Pikor LA, Pusic A, Lockwood WW, Lam WL and Tsao MS: SOX15 is a
candidate tumor suppressor in pancreatic cancer with a potential
role in Wnt/β-catenin signaling. Oncogene. 33:279–288. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Sano M, Driscoll DR, DeJesus-Monge WE,
Quattrochi B, Appleman VA, Ou J, Zhu LJ, Yoshida N, Yamazaki S,
Takayama T, et al: Activation of WNT/β-catenin signaling enhances
pancreatic cancer development and the malignant potential Via
Up-regulation of Cyr61. Neoplasia. 785–794. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Du WW, Yang W, Liu E, Yang Z, Dhaliwal P
and Yang BB: Foxo3 circular RNA retards cell cycle progression via
forming ternary complexes with p21 and CDK2. Nucleic Acids Res.
44:2846–2858. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Legnini I, Di Timoteo G, Rossi F, Morlando
M, Briganti F, Sthandier O, Fatica A, Santini T, Andronache A, Wade
M, et al: Circ-ZNF609 Is a circular RNA that can be translated and
functions in myogenesis. Mol Cell. 66:22–37.e9. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Yang Y, Fan X, Mao M, Song X, Wu P, Zhang
Y, Jin Y, Yang Y, Chen LL, Wang Y, et al: Extensive translation of
circular RNAs driven by N6-methyladenosine. Cell Res.
27:626–641. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang Y, Zhang XO, Chen T, Xiang JF, Yin
QF, Xing YH, Zhu S, Yang L and Chen LL: Circular intronic long
noncoding RNAs. Mol Cell. 51:792–806. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ashwal-Fluss R, Meyer M, Pamudurti NR,
Ivanov A, Bartok O, Hanan M, Evantal N, Memczak S, Rajewsky N and
Kadener S: circRNA biogenesis competes with pre-mRNA splicing. Mol
Cell. 56:55–66. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen L and Shan G: Circular RNAs remain
peculiarly unclear in biogenesis and function. Sci China Life Sci.
58:616–618. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chen L, Huang C, Wang X and Shan G:
Circular RNAs in eukaryotic cells. Curr Genomics. 16:312–318. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Chen S and Zhao Y: Circular RNAs:
Characteristics, function, and role in human cancer. Histol
Histopathol. 33:887–893. 2018.PubMed/NCBI
|