Introduction
Although a variety of treatments are available for
gastric cancer, the 5-year survival rate remains low due to
recurrence and metastasis (1). Tumor
recurrence caused by metastasis accounts for the majority of
gastric cancer mortalities (1).
Growing tumor cells require a new blood supply for survival and
metastasis, which is obtained via angiogenesis involving vascular
endothelial cells (2,3). Consequently, anti-angiogenic drugs have
been incorporated into standard anticancer treatment regimens
(2,3). However, aggressive tumor cells develop
resistance via the process of vasculogenic mimicry (VM), in which
tumor cells create their own blood-delivery channels without the
involvement of endothelial cells (4). VM is facilitated by the plasticity of
cancer cells that form de novo vascular networks for the
perfusion of rapidly growing tumors (5). VM is associated with poor prognosis in
patients with gastric adenocarcinoma (6). Therefore, a greater understanding of VM
formation is vital for the development of novel anticancer
therapies.
Erythropoietin-producing human hepatocellular
receptor A2 (EphA2), a transmembrane receptor tyrosine kinase of
the Eph family, has been implicated in tumorigenesis and cancer
development in a number of different types of solid tumor,
including gastric cancer (7,8). Overexpression of EphA2 and its ligand
ephrinA1 is an independent prognostic factor for postoperative
gastric adenocarcinoma (9). EphA2
may also serve a crucial role in the expression of vascular
endothelial growth factor (VEGF) and in the development of tumor
angiogenesis by interacting with the tumor microenvironment
(10–12). The tumor microenvironment is composed
of malignant cancer cells and the surrounding stroma, which
includes fibroblasts, vascular endothelial cells, immune cells and
the extracellular matrix (13).
Activated fibroblasts, the primary components of the stroma, are
termed cancer-associated fibroblasts (CAFs). In a previous study,
it was observed that CAFs may promote gastric tumorigenesis through
EphA2 (14).
Although CAFs are key determinants in the malignant
progression of cancer, their functional contribution to VM
formation in gastric cancer remains unclear. The present study
hypothesized that CAFs may enhance VM formation in gastric cancer
cells by activating the EphA2 signaling pathway. To test this
hypothesis, the role of EphA2 signaling in the formation of VM
channels was investigated using the indirect co-culture method.
Materials and methods
Primary tumor samples and
patients
Human gastric cancer samples and adjacent
non-cancerous samples (distance, 5–20 cm) were obtained from 12
patients with gastric adenocarcinoma, who underwent total or
subtotal curative gastrectomy at the Department of Surgery at Asan
Medical Center, University of Ulsan College of Medicine (Seoul,
Korea) between May 2015 and June 2016. Of the 12 patients, 10 were
male and 2 were female, 2 had Tumor-Node-Metastasis (TNM) stage IA,
2 had stage IB, 1 had stage IIA, 3 had stage IIB, 2 had stage IIIA
and 2 had stage IIIC tumors. The patients' mean age was 64 years
(range, 39–81 years). All samples were histologically evaluated
according to the World Health Organization criteria (15). Each tumor was classified using the
TNM system recommended by the International Union against Cancer
(16). None of the patients had
received anticancer therapy prior to sample collection; patients
with papillary, mucinous and unclassified adenocarcinomas were
excluded from the study. The present study was approved by the
Institutional Review Board (approval no. 2015-0370) of the Asan
Medical Center, and was conducted in accordance with the
Declaration of Helsinki. Written informed consent was obtained from
all patients.
Isolation and culture of stromal
fibroblasts
CAFs were extracted from the gastric tumor tissues,
while normal gastric fibroblasts (NFs) were obtained from
non-cancerous tissue samples. To isolate stromal fibroblasts,
2–3-mm3 tissue fragments were digested with collagenase
(1 mg/ml) at 37°C for 30 min, and plated in Dulbecco's modified
Eagle's medium (DMEM) with 10% fetal bovine serum (HyClone; Thermo
Fisher Scientific, Inc.), sodium bicarbonate (Sigma-Aldrich; Merck
KGaA), sodium pyruvate (Gibco; Thermo Fisher Scientific, Inc.) and
antibiotics (50 U/ml penicillin and 50 µg/ml streptomycin; Gibco;
Thermo Fisher Scientific, Inc.). After two passages, epithelial
cells were absent from the culture, and fast-growing fibroblasts
were enriched. Isolated fibroblasts were transferred to new culture
dishes and serial passage was performed every 4–7 days. Fibroblasts
between passages 3 and 10 were used, and the majority were used at
passage 5. Activated fibroblasts were confirmed by microscopic
assessment of cell morphology and immunohistochemical staining for
α smooth muscle actin (α-SMA; 1:1,000; catalog no. ab5694; Abcam)
and vimentin (1:1,000; catalog no. V6389; Sigma-Aldrich; Merck
KGaA). Cells cultured on sterilized cover slips were fixed for 30
min at room temperature with 4% paraformaldehyde in phosphate
buffer (PB; 77.4 ml 1 M Na2HPO4 + 22.6 ml 1 M
Na2H2PO4 in 900 ml distilled
water), permeabilized for 5 min with Triton X-100 (Sigma-Aldrich;
Merck KGaA) diluted to 0.5% in PBS and blocked by incubation with
2% normal goat serum (catalog no. ab7481; Abcam) in PBS for 1 h at
room temperature. Following overnight incubation at 4°C with the
primary antibodies, cells were incubated with anti-rabbit
Cy3®-conjugated secondary antibody (1:1,000; catalog no.
ab6939; Abcam) for a-SMA or FITC-conjugated anti-mouse secondary
antibody (1:1,000; catalog no. ab6785; Abcam) for vimentin for 2 h
at room temperature. Cells were rinsed several times with phosphate
buffer and mounted on glass slides for fluorescent microscopic
examination at ×200 magnification (5 fields per slide) and analysis
using NIS-Elements software (version 4.00; Nikon Instruments,
Inc.).
Preparation of conditioned medium
(CM)
For the CM, CAFs and NFs (1×105 cells/ml)
were cultured in DMEM containing 10% FBS. Upon reaching 80%
confluence, fibroblasts were washed with serum-free medium and
incubated for 3 days at 37°C in a humidified atmosphere containing
5% CO2 in DMEM without serum. The supernatant was
collected, centrifuged to remove cellular debris (300 × g at 4°C
for 10 min) and clarified using a 0.45-µm filter. Aliquots were
frozen and stored at −20°C until use.
Cell culture and reagents
Human gastric carcinoma cell lines SNU-1 (KCLB
no.00001.1), SNU-216, (KCLB no.00216), SNU-601 (KCLB no.00601),
MKN-45 (KCLB no. 80103), and AGS (KCLB no.21739) were obtained from
the Korean Cell Line Bank (KCLB) of Seoul National University
(Seoul, South Korea). Mycoplasma testing was performed for
all cell lines used, and mycoplasma were eliminated using
MC-210 (WakenBtech Co., Ltd). Cells were maintained in DMEM (Thermo
Fisher Scientific, Inc.) containing 10% FBS, sodium bicarbonate
(Sigma-Aldrich; Merck KGaA), sodium pyruvate (Gibco; Thermo Fisher
Scientific, Inc.), and antibiotics (50 U/ml penicillin and 50 µg/ml
streptomycin; Gibco; Thermo Fisher Scientific, Inc.) in a 5%
CO2 humidified incubator at 37°C. The culture medium was
refreshed every 2–3 days. Mitogen-activated protein kinase (MAPK)
inhibitor PD98059 and phosphoinositide 3-kinase (PI3K) inhibitor
LY294002 were obtained from Sigma-Aldrich; Merck KGaA. To block the
EphA2 function of gastric cancer cells, a novel EphA2 receptor
inhibitor ALW-II-41-27 (MedChem Express) was used. For all in
vitro studies, ALW-II-41-27 was dissolved in 0.01% DMSO and
then diluted to a final concentration of 1 µM.
Immunoblot analyses
Cells were lysed using SDS-PAGE sample buffer (62.5
mM Tris-HCl, pH 6.8, 2% SDS, 7.8% glycerol, 4.5% mercaptoethanol
and 0.1% bromophenol blue) and boiled for 5 min at 100°C. The
protein lysates were clarified by centrifugation (8,000 × g at 4°C
for 10 min), and the concentrations of the supernatants were
determined relative to a bovine serum albumin (BSA; MP Biomedicals,
Santa Ana, California, USA) standard. The cell lysate (30 µg/well)
was separated by SDS-PAGE (4% stacking gel and 10% polyacrylamide
separating gel) for 70 min at 130 V. Protein extracts were
transferred onto nitrocellulose membranes with a Bio-Rad
Laboratories transfer unit for 120 min at 200 mA. The membranes
were incubated in blocking buffer (2% BSA in Tween 20/TBS) for 1 h
on a rotating platform at room temperature. The membranes were
sequentially incubated overnight at 4°C with the following primary
antibodies: Rabbit monoclonal anti-EphA2 (1:1,000; catalog no.
6997), rabbit monoclonal anti-pEphA2-ser (1:1,000; catalog no.
6347), rabbit polyclonal anti-matrix metalloproteinase 2 (MMP2;
1:1,000; catalog no. 4022s), β-actin (1:5,000; catalog no. 4967)
(all from Cell Signaling Technology, Inc.), rabbit polyclonal
anti-VEGF-A (1:1,000; catalog no. ab46154) and rabbit polyclonal
anti-VE-cadherin (1:1,000; ab33168) (both from Abcam). Following
incubation for 1 h at room temperature with a horseradish
peroxidase-conjugated secondary antibody (1:1,000, goat anti-rabbit
immunoglobulin G; catalog no. ab6721; Abcam), immuno-reactive bands
were detected using chemiluminescence reagents (Pierce; Thermo
Fisher Scientific, Inc.). The optical density of the bands was
analyzed using GS-670 densitometry software (ver. 1.4, Bio-Rad
Laboratories, Inc.). Each experiment was performed in
triplicate.
VM tube formation assay
A 24-well culture plate was coated with 0.1 ml (50
µl/cm2) growth factor-reduced Matrigel
(Geltrex® LDEV-Free reduced growth factor basement
membrane matrix), which was allowed to polymerize for 30 min at
37°C. AGS cells (1×105 cells/well) were seeded on the
solid gel, treated with CM with or without inhibitors, and
incubated at 37°C for 24 or 48 h. The number of tubules and
intersections in 5–7 random fields were photographed at ×100
magnification by an inverted fluorescence microscope (Nikon Eclipse
Ti; Nikon Instruments, Inc.) and counted using Image J software
(version 1.50i; National Institutes of Health), and the mean values
were used for analysis.
EphA2 gene knockdown using small
interfering (si)RNA
RNA interference was used to knock down the EphA2
gene in gastric cancer cells. siRNA for EphA2 was obtained from
Santa Cruz Biotechnology. The target sequence was
5′-AATGACATGCCGATCTACATG-3′ (EphA2) (17), and the non-silencing siRNA sequence
5′-AATTCTCCGAACGTGTCACGT-3′ was used as the negative control. AGS
cancer cells (5×105 cells/well) were transfected with
siRNA at a final concentration of 20–100 nM using Lipofectamine™
2000 (Invitrogen; Thermo Fisher Scientific, Inc.). After 8 h of
transfection, the medium was replaced with fresh DMEM medium
containing 10% fetal bovine serum (HyClone; GE Healthcare Life
Sciences). After 48 h siRNA transfection, suppression of EphA2
expression was confirmed by western blotting performed as
aforementioned.
Statistical analysis
The data were analyzed using one-way analysis of
variance and Bonferroni post-hoc test for multiple comparisons. The
results were obtained from ≥3 independent sets of experiments, and
the data are expressed as the mean ± standard deviation. P<0.001
was considered to indicate a statistically significant difference.
Statistical analyses were performed using SPSS software (version
13.0; SPSS Inc.).
Results
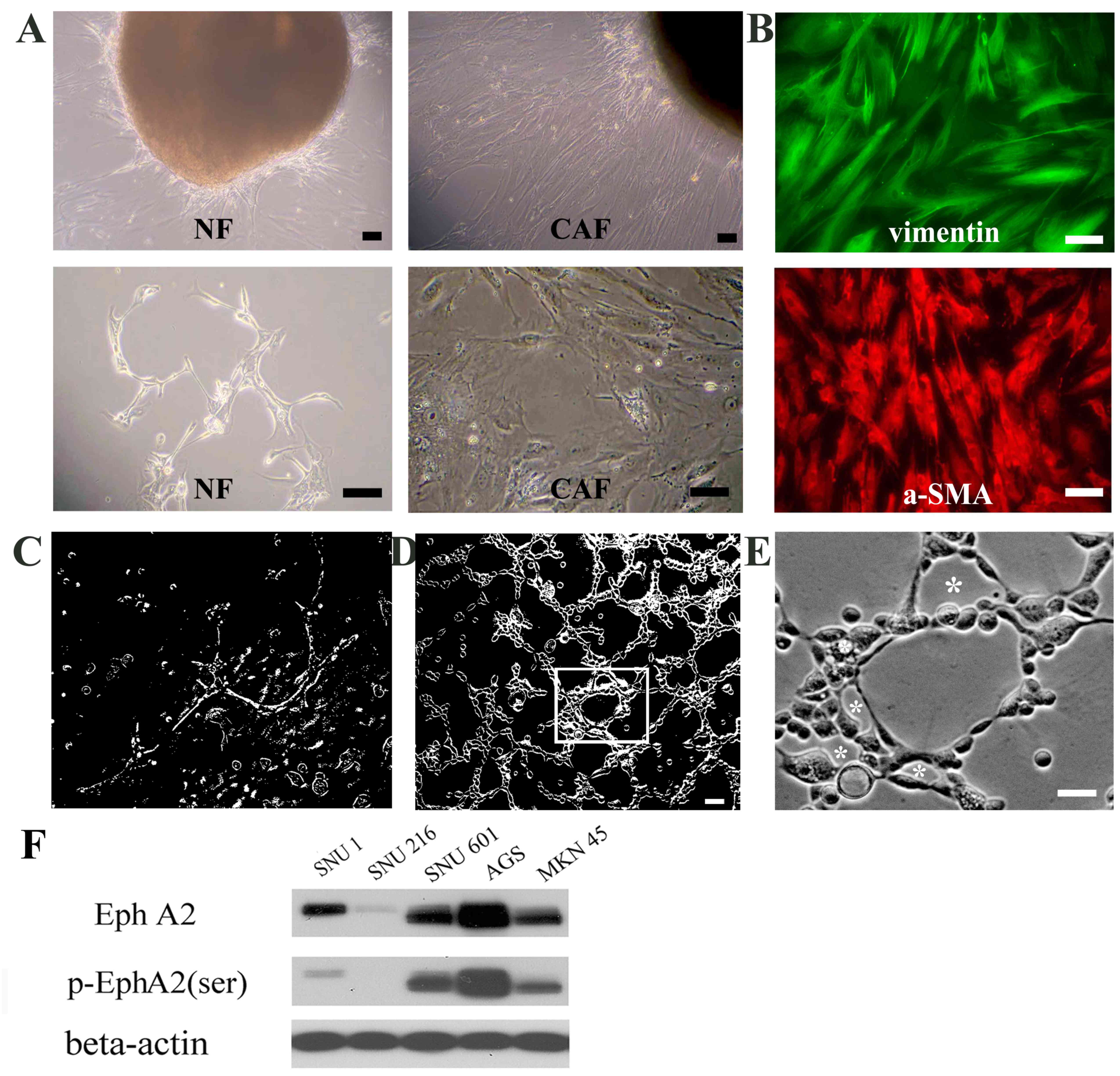
In order to elucidate the effect of CAFs on VM
formation, CAFs and adjacent NFs were isolated from resected
gastric cancer tissue fragments (Fig.
1A). Almost all CAFs exhibited positive staining for vimentin
(a mesenchymal marker) and α-SMA (an activated tumor fibroblast
marker; Fig. 1B). Following
identification of CAFs, CAF-CM was prepared.
EphA2 expression is important for
CAF-CM-induced VM formation in gastric cancer cell lines
First, EphA2 expression and phosphorylation levels
were assessed in the five gastric carcinoma lines. The highest
levels of EphA2 and phosphorylated EphA2 (serine-residue) were
observed in AGS cells, whereas the expression of EphA2 and
pEphA2-ser were not detected in the SNU216 cell line (Fig. 1F). To investigate the involvement of
EphA2 in VM formation, upon interaction with CAFs, two of the
cancerous cell lines (SNU216 expressing low-EphA2 and AGS
expressing high-EphA2) were treated with CAF-CM (50%) on a Matrigel
matrix for VM-tube formation. SNU216 cells exhibited defective VM
channel formation (Fig. 1C). AGS
cells, which possessed high EphA2 activity, produced a prominent
tubular network (Fig. 1D) consisting
of tubular structures of various sizes, including micro vessel-like
channels (Fig. 1E). This result
indicated that EphA2 signaling may serve a key role in initiating
VM formation in gastric cancer cells stimulated by CAF-CM.
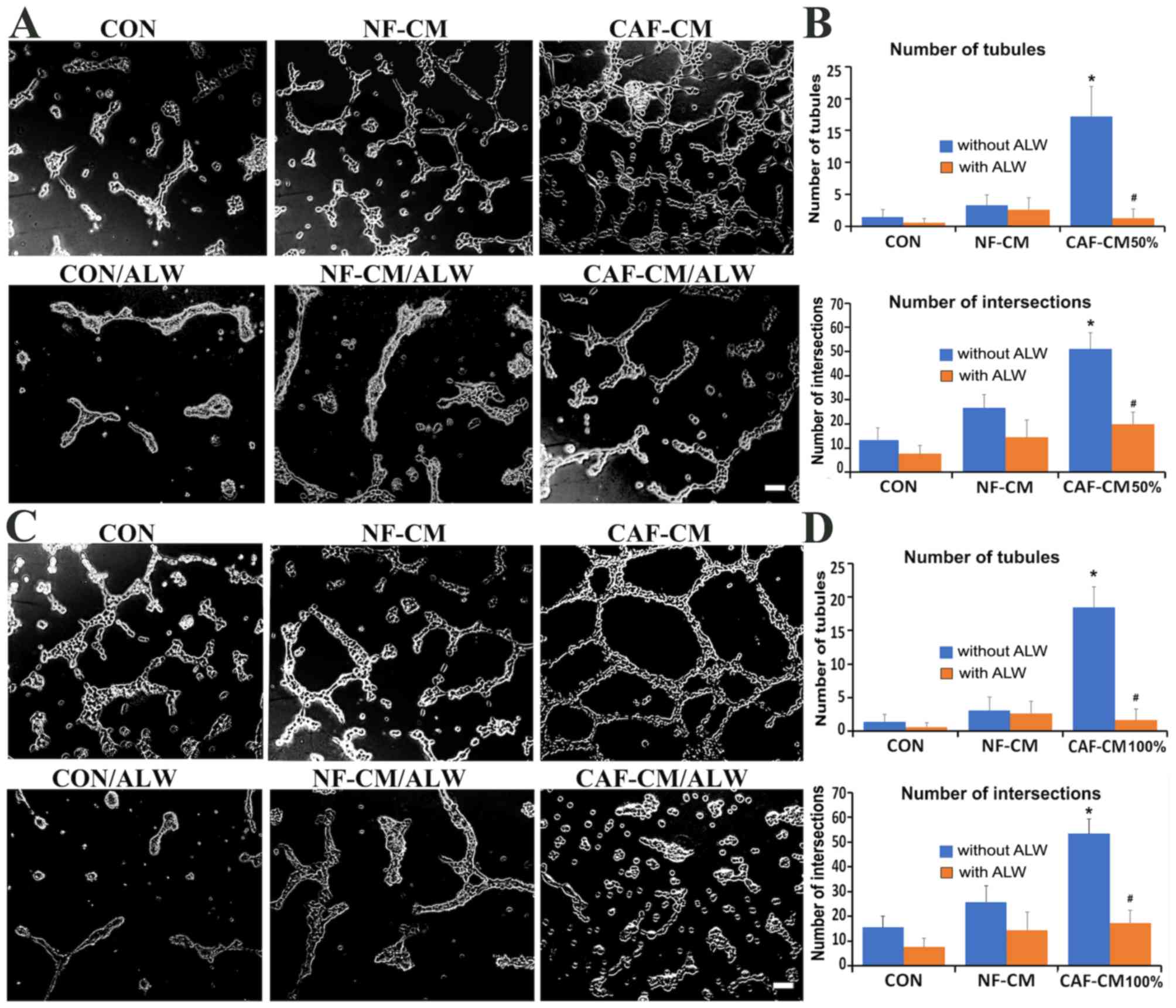
EphA2-inhibition abrogates VM
formation promoted by CAF-CM in gastric cancer cells
In order to elucidate the role of EphA2 in VM
formation promoted by CAF-CM, AGS cells were treated with DMEM
(control), NF-CM and CAF-CM (50 or 100%) with or without
ALW-II-41-27, a novel inhibitor of EphA2 receptor tyrosine kinase
(18,19). Cells treated with DMEM (CON) or NF-CM
exhibited cellular rearrangement and a few scattered tubules
(Fig. 2A and C). Both CAF-CM (50%)
and CAF-CM (100%) enhanced the ability of AGS cells to generate VM
channels (Fig. 2A and C), and
significantly increased the number of tubules and intersections
(P<0.001 vs. DMEM-treated AGS cells; Fig. 2B and D). Furthermore, treatment with
1 µM ALW-II-41-27 significantly suppressed the stimulatory effects
of CAF-CM on VM formation (P<0.001 vs. CAF-CM; Fig. 2B and D). This result suggested that
the EphA2 pathway may facilitate CAF-CM-induced VM formation.
CAF-CM-induced VM formation is blocked
by PI3K/AKT-inhibition in AGS cells
In order to investigate whether the PI3K/AKT pathway
is required for VM formation by CAF-CM, AGS cells were treated with
the PI3K/Akt inhibitor, LY294002 (LY). PD98059 (PD), a specific
inhibitor of MAPK-extracellular signal-regulated kinase (ERK), was
used to compare the effects of the two inhibitors. VM formation was
observed at 24 and 48 h following incubation on a Matrigel matrix.
Blocking PI3K/AKT signaling with LY treatment significantly
inhibited VM formation induced by CAF-CM, while PD treatment did
not affect the formation of tubule-like VM structures by AGS cells
incubated in CAF-CM for 24 h (Fig.
3A; 24 h). Similar results were observed in the 48-h incubation
group, though the VM channels and tubule walls possessed a greater
number of cells (Fig. 3A; 48 h).
Quantitative analysis indicated that the number of tubules and
intersections in CAF-CM of LY-treated AGS cells was significantly
less than those in the CAF-CM-treated cells (#P<0.001 vs.
CAF-CM; Fig. 3B), suggesting that
PI3K/AKT signaling may be important in VM structure acquisition by
CAF-CM in AGS gastric cancer cells.
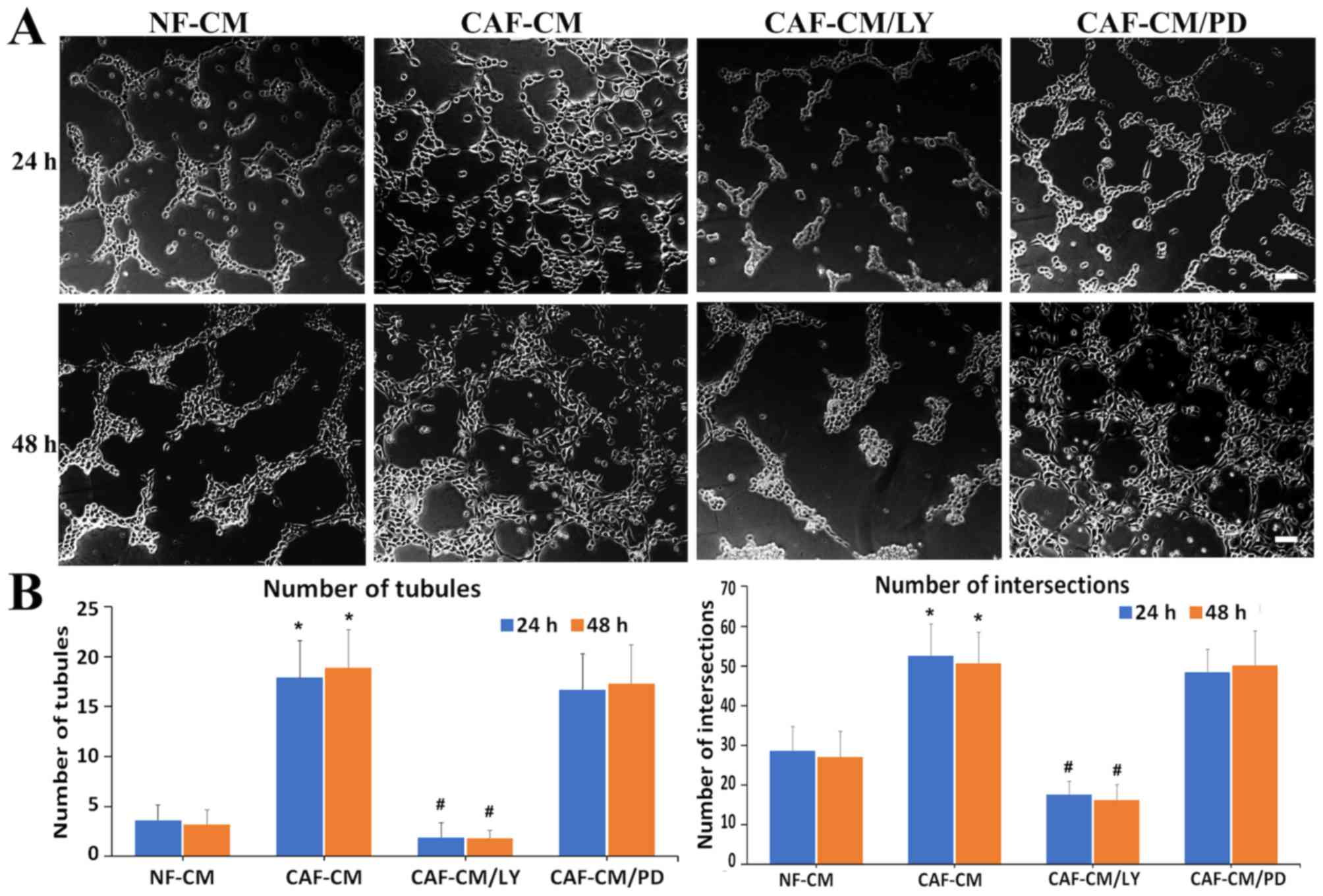 | Figure 3.CAF-CM-induced vasculogenic mimicry
formation is blocked by a PI3K/AKT-inhibitor in gastric cancer
cells. (A) AGS gastric cancer cells were treated with CAF-CM (50%)
with or without inhibitors of PI3K/AKT signaling (LY294002, 20 µM)
and MAPK-ERK kinase (PD98059, 20 µM) for 24 and 48 h. Scale bars,
100 µm. (B) Graphs present the number of tubules and intersections.
Values are expressed as the mean ± standard deviation of three
independent experiments. *P<0.001 vs. control;
#P<0.001 vs. CAF-CM. CAF, cancer-associated
fibroblast; CM, conditioned-medium; PI3K, phosphoinositide
3-kinase; AKT, protein kinase B; MAPK, mitogen activated protein
kinase; ERK, extracellular signal-regulated kinases; NF, normal
gastric fibroblasts; LY, LY294002; PD, PD98059. |
Silencing of the EphA2 gene suppresses
the VM-forming ability of gastric cancer cells
In order to investigate the role of EphA2 expression
in the induction of VM formation by CAF-CM, EphA2 was
knocked down in AGS cells using RNA interference. EphA2 siRNA
efficiently silenced EphA2 expression (Fig. 4C). AGS cells were then transfected
with non-silencing siRNA (siCON) and siRNA EphA2 (siEphA2), and
subsequently treated with NF-CM or CAF-CM (50%) on a Matrigel
matrix for 24 or 48 h. In both the NF-CM and CAF-CM-treated groups,
the VM formation capacity of siEphA2 cells was reduced compared
with siCON cells (Fig. 4A).
Similarly, fewer tubule structures were observed in siEphA2 cells
of the 48-h group (Fig. 4A). Plots
presenting the number of tubules and intersections demonstrated
that siRNA knockdown of EphA2 significantly impaired the ability of
AGS cells to develop vascular-like channels (P<0.001 vs. siCON)
(Fig. 4B). These data suggest that
in AGS gastric cancer cells, CAF-CM induced VM formation via the
EphA2 signaling pathway. Silencing of EphA2 affected the expression
levels of proteins associated with VM formation, such as MMP2,
VEGF-A and VE-cadherin (Fig. 4C). In
siRNA EphA2 (siEphA2)-treated AGS cells, the expression levels of
MMP2, VEGF-A and VE-cadherin were significantly decreased compared
with those of the non-silencing siRNA (siCON) treated cells
(P<0.001 vs. siCON; Fig. 4D).
This result suggests that EphA2 expression may be significantly
associated with the expression of VM-associated proteins.
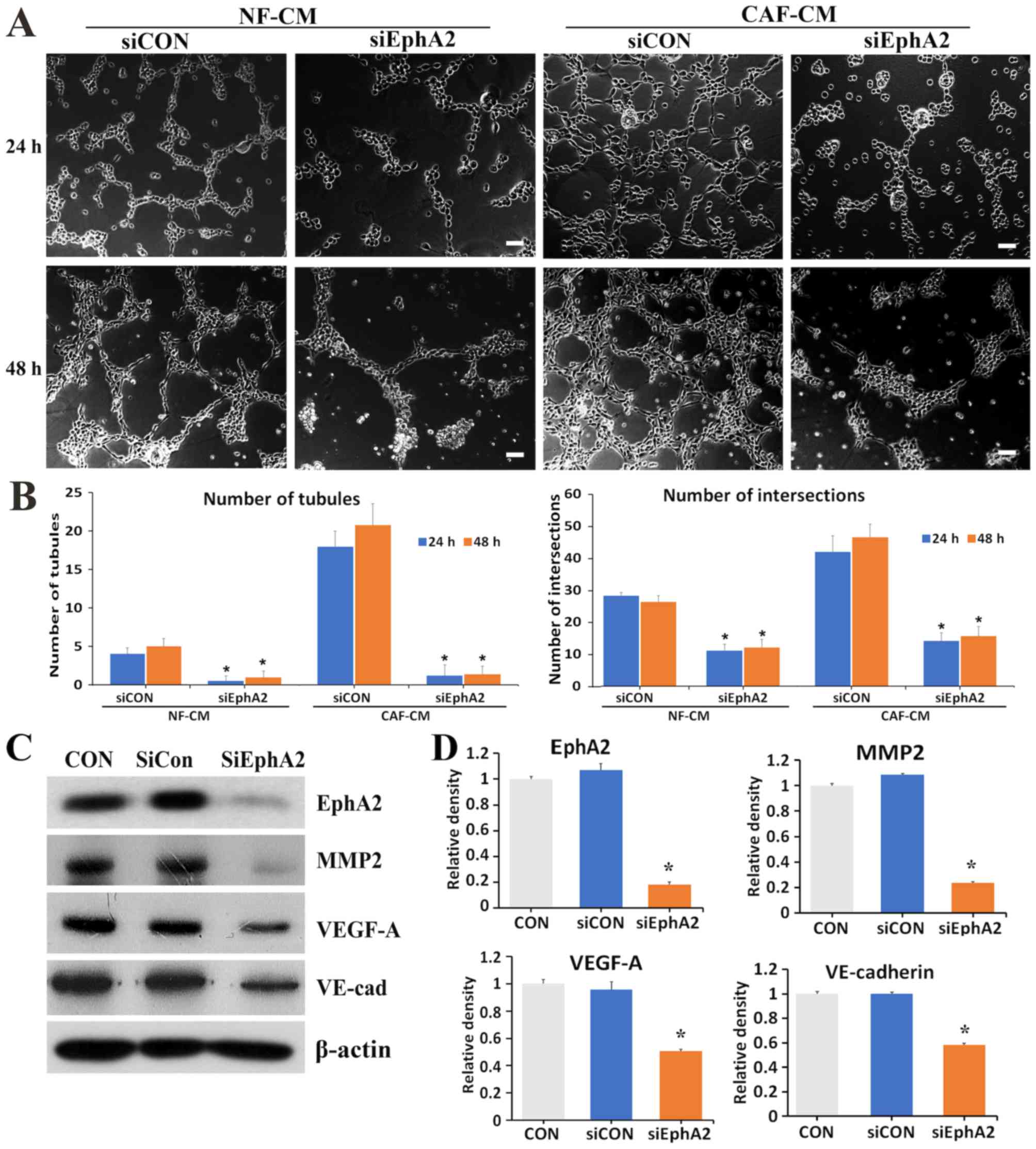 | Figure 4.Silencing of the EphA2 gene
suppresses the VM forming ability of gastric cancer cells. (A)
Representative images of VM formation in AGS cells transfected with
siCON or siEphA2 for 24 and 48 h, in either NF-CM or CAF-CM (50%).
Scale bars, 100 µm. (B) Graphs present the number of tubules and
intersections in each experimental group. (C) Western blot images
of EphA2, MMP2, VEGF-A and VE-cadherin expression in siCON and
siEphA2 AGS cells. (D) Graphs presenting the protein expression
levels of EphA2, MMP2, VEGF-A and VE-cadherin. Mean ± standard
deviation from three independent experiments. *P<0.001 vs.
siCON. EphA2, erythropoietin-producing human hepatocellular
receptor A2; VM, vasculogenic mimicry; CON, non-treated AGS cells;
siRNA, small interfering RNA; siCON, non-silencing siRNA; siEphA2,
siRNA-EphA2; VE-cad, VE-cadherin; NF, normal gastric fibroblasts;
CM, conditioned medium; CAF, cancer-associated fibroblasts; MMP2,
matrix metalloproteinase 2; VEGF-A, vascular endothelial growth
factor A. |
Discussion
CAFs in the cancer stroma have been reported to be
key modulators of tumor growth and metastasis (20). Crosstalk between cancer cells and
CAFs in the tumor microenvironment may be associated with the
development and progression of gastric cancer (21–23).
Previously, it was demonstrated that CAF-CM induced
epithelial-mesenchymal transition (EMT) and promoted gastric cancer
cell migration and invasion (14).
The present study further revealed that CAF-CM from human gastric
adenocarcinoma tissues promoted VM formation in gastric cancer AGS
cells. A number of studies have indicated that CAFs in the
microenvironment act as key factors in VM formation via
EMT-associated processes (24,25).
The present study demonstrated that AGS cells with
high EphA2-expression levels formed VM structures on a Matrigel
matrix, whereas SNU216 cells with poor EphA2-expression did not
form VM networks under CAF-CM treatment conditions. Furthermore,
treatment with a selective EphA2 inhibitor (ALW-II-41-27), or
EphA2-targeted siRNA, markedly decreased CAF-CM-induced VM
formation, indicating that CAFs induce VM formation by activating
the EphA2 pathway in gastric cancer cells. The results of the
present study support previous indications that EphA2 serves a
critical role in the formation of matrix-rich tubular networks by
aggressive melanoma cells (26). In
addition, EphA2 receptor tyrosine kinase was significantly
associated with VM formation in gastric cancer cells, which
supported previous results suggesting that EphA2 acted as a key
regulator in VM formation in head and neck squamous cell carcinoma
(12), and that downregulated EphA2
expression inhibited VM formation in gallbladder cancer cells
(27).
In gastric cancer, EphA2 overexpression has been
demonstrated to be associated with a higher disease stage,
decreased survival rate and metastasis (9,28,29).
VEGF, a key factor in angiogenesis, and EphA2 are associated with
endothelial cell migration, paracellular permeability and tumor
neovascularization (30,31). Reportedly, EphA2 is involved in
VEGF-induced vascular assembly and tumor angiogenesis, which are
indicators of tumor recurrence and progression (32,33).
Previously, it was revealed that EphA2 activation in gastric cancer
cells was triggered by VEGF release upon treatment with CAF-CM
(14). Furthermore, the
phosphorylated forms of ERK and VEGFR2 are important downstream
factors of VEGF, and, thus, their protein expression levels would
be required to delineate the exact role of VEGF in VM
formation.
The results of the present study indicate that
CAF-CM enhanced VM formation in high EphA2-expressing AGS cells via
activation of the PI3K signaling pathway. The role of the PI3K
signaling pathway in VEGF-mediated cell migration and invasion was
evident in cholangiocarcinoma (34).
A recent study (13) demonstrated
that treatment with recombinant human hepatocyte growth factor
(HGF) and CAF-CM increased the formation of VM and mosaic vessels,
comprising human umbilical vein endothelial cells (HUVECs) and
gastric cancer cells. It was argued that CAF-derived HGF promoted
angiogenesis, VM and mosaic vessel formation via PI3K/AKT and
ERK1/2 signaling in gastric cancer (13). Furthermore, the contributions of PI3K
and EphA2 signaling to tumor growth and VM formation were
demonstrated using gall bladder carcinoma and prostate cancer
tissues (35,36). The results from these studies were
consistent with those of the present study.
MMP2 and VE-cadherin are key mediators of invasion,
metastasis and matrix remodeling during VM formation in the tumor
microenvironment. MMP2 and VE-cadherin expression were
significantly associated with VM formation in glioma and pancreatic
cancer (37,38). The results of the present study
demonstrated that EphA2 silencing decreased the expression levels
of MMP2 and VE-cadherin, and suppressed the VM-forming ability of
gastric cancer cells, which is largely in accordance with previous
studies (37,38).
The present study was limited in that it used a cell
culture model to demonstrate the role of EphA2-PI3K signaling in VM
formation. However, these results are supported by a previous
publication (14) in which it was
revealed that CAF-CM significantly enhances gastric EMT, as well as
gastric cancer cell migratory and invasive properties via EphA2
signaling in a ligand-independent manner. These results support
those stated in the present study, as EMT and cancer cell motility
are both essential for the development of VM. A positive
correlation between EphA2 expression and VM in patients with
gastric adenocarcinoma has also been reported in a recently
published paper (39). Animal models
should be utilized in order to further investigate the association
between EphA2-PI3K signaling and CAF-CM-induced VM formation, in
which the animals are transplanted with CAF-CM-treated AGS or
siEphA2-AGS cells.
In conclusion, the present study indicated that high
levels of EphA2 were associated with VM formation induced by CAF-CM
in gastric cancer AGS cells. CAF-CM promoted VM formation by
activating the EphA2-PI3K pathway, indicating that CAFs promote
gastric cancer progression by inducing VM formation via the
activation of the EphA2-PI3K pathway.
Acknowledgements
Not applicable.
Funding
The present study was supported by the Asan
Institute for Life Sciences, Seoul, South Korea (grant no.
2016-014).
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author upon reasonable
request.
Authors' contributions
HSK and HNH generated the hypothesis, designed the
experiments and wrote the manuscript. YJW, JHS and HJK performed
the experiments. HNH and BSK interpreted the data, designed and
refined the study and participated in the writing of the
manuscript. HNH also revised the manuscript.
Ethics approval and consent to
participate
The present study was approved by the Institutional
Review Board (approval no. 2015-0370) of the Asan Medical Center
and conducted in accordance with the Declaration of Helsinki.
Written informed consent was obtained from all patients included in
the present study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Lee JH, Kim HI, Kim MG, Ha TK, Jung MS and
Kwon SJ: Recurrence of gastric cancer in patients who are
disease-free for more than 5 years after primary resection.
Surgery. 159:1090–1098. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sitohy B, Nagy JA and Dvorak HF:
Anti-VEGF/VEGFR therapy for cancer: Reassessing the target. Cancer
Res. 72:1909–1914. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Pinto MP, Sotomayor P, Carrasco-Avino G,
Corvalan AH and Owen GI: Escaping antiangiogenic therapy:
Strategies employed by cancer cells. Int J Mol Sci. 17:2016.
View Article : Google Scholar
|
|
4
|
Maniotis AJ, Folberg R, Hess A, Seftor EA,
Gardner LM, Pe'er J, Trent JM, Meltzer PS and Hendrix MJ: Vascular
channel formation by human melanoma cells in vivo and in vitro:
Vasculogenic mimicry. Am J Pathol. 155:739–752. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Seftor RE, Hess AR, Seftor EA, Kirschmann
DA, Hardy KM, Margaryan NV and Hendrix MJ: Tumor cell vasculogenic
mimicry: From controversy to therapeutic promise. Am J Pathol.
181:1115–1125. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Li M, Gu Y, Zhang Z, Zhang S, Zhang D,
Saleem AF, Zhao X and Sun B: Vasculogenic mimicry: A new prognostic
sign of gastric adenocarcinoma. Pathol Oncol Res. 16:259–266. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakamura R, Kataoka H, Sato N, Kanamori M,
Ihara M, Igarashi H, Ravshanov S, Wang YJ, Li ZY, Shimamura T, et
al: EPHA2/EFNA1 expression in human gastric cancer. Cancer Sci.
96:42–47. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wykosky J and Debinski W: The EphA2
receptor and ephrinA1 ligand in solid tumors: Function and
therapeutic targeting. Mol Cancer Res. 6:1795–1806. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Yuan WJ, Ge J, Chen ZK, Wu SB, Shen H,
Yang P, Hu B, Zhang GW and Chen ZH: Over-expression of EphA2 and
EphrinA-1 in human gastric adenocarcinoma and its prognostic value
for postoperative patients. Dig Dis Sci. 54:2410–2417. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Brantley-Sieders DM, Fang WB, Hwang Y,
Hicks D and Chen J: Ephrin-A1 facilitates mammary tumor metastasis
through an angiogenesis-dependent mechanism mediated by EphA
receptor and vascular endothelial growth factor in mice. Cancer
Res. 66:10315–10324. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cheng N, Brantley DM, Liu H, Lin Q,
Enriquez M, Gale N, Yancopoulos G, Cerretti DP, Daniel TO and Chen
J: Blockade of EphA receptor tyrosine kinase activation inhibits
vascular endothelial cell growth factor-induced angiogenesis. Mol
Cancer Res. 1:2–11. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Wang W, Lin P, Sun B, Zhang S, Cai W, Han
C, Li L, Lu H and Zhao X: Epithelial-mesenchymal transition
regulated by EphA2 contributes to vasculogenic mimicry formation of
head and neck squamous cell carcinoma. Biomed Res Int.
2014:8039142014.PubMed/NCBI
|
|
13
|
Ding X, Xi W, Ji J, Cai Q, Jiang J, Shi M,
Yu Y, Zhu Z and Zhang J: HGF derived from cancer-associated
fibroblasts promotes vascularization in gastric cancer via PI3K/AKT
and ERK1/2 signaling. Oncol Rep. 40:1185–1195. 2018.PubMed/NCBI
|
|
14
|
Hong HN, Won YJ, Shim JH, Kim HJ, Han SH,
Kim BS and Kim HS: Cancer-associated fibroblasts promote gastric
tumorigenesis through EphA2 activation in a ligand-independent
manner. J Cancer Res Clin Oncol. 144:1649–1663. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Japanese classification of gastric
carcinoma: 3rd English edition. Gastric cancer: Official journal of
the International Gastric Cancer Association and the Japanese
Gastric Cancer Association, . 14:101–112. 2011.
|
|
16
|
Kwon SJ: Evaluation of the 7th UICC TNM
staging system of gastric cancer. J Gastric Cancer. 11:78–85. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhou Z, Yuan X, Li Z, Tu H, Li D, Qing J,
Wang H and Zhang L: RNA interference targeting EphA2 inhibits
proliferation, induces apoptosis, and cooperates with cytotoxic
drugs in human glioma cells. Surg Neurol. 70:562–568; discussion
568-569. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Moccia M, Liu Q, Guida T, Federico G,
Brescia A, Zhao Z, Choi HG, Deng X, Tan L, Wang J, et al:
Identification of novel small molecule inhibitors of oncogenic RET
kinase. PLoS One. 10:e01283642015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Amato KR, Wang S, Hastings AK, Youngblood
VM, Santapuram PR, Chen H, Cates JM, Colvin DC, Ye F,
Brantley-Sieders DM, et al: Genetic and pharmacologic inhibition of
EPHA2 promotes apoptosis in NSCLC. J Clin Invest. 124:2037–2049.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Whiteside TL: The tumor microenvironment
and its role in promoting tumor growth. Oncogene. 27:5904–5912.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Fuyuhiro Y, Yashiro M, Noda S, Matsuoka J,
Hasegawa T, Kato Y, Sawada T and Hirakawa K: Cancer-associated
orthotopic myofibroblasts stimulates the motility of gastric
carcinoma cells. Cancer Sci. 103:797–805. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhi K, Shen X, Zhang H and Bi J:
Cancer-associated fibroblasts are positively correlated with
metastatic potential of human gastric cancers. J Exp Clin Cancer
Res. 29:662010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yu B, Chen X, Li J, Qu Y, Su L, Peng Y,
Huang J, Yan J, Yu Y, Gu Q, et al: Stromal fibroblasts in the
microenvironment of gastric carcinomas promote tumor metastasis via
upregulating TAGLN expression. BMC Cell Biol. 14:172013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sun T, Zhao N, Zhao XL, Gu Q, Zhang SW,
Che N, Wang XH, Du J, Liu YX and Sun BC: Expression and functional
significance of Twist1 in hepatocellular carcinoma: Its role in
vasculogenic mimicry. Hepatology. 51:545–556. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu K, Sun B, Zhao X, Wang X, Li Y, Qiu Z,
Gu Q, Dong X, Zhang Y, Wang Y and Zhao N: Hypoxia induced
epithelial-mesenchymal transition and vasculogenic mimicry
formation by promoting Bcl-2/Twist1 cooperation. Exp Mol Pathol.
99:383–391. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hess AR, Seftor EA, Gardner LM,
Carles-Kinch K, Schneider GB, Seftor RE, Kinch MS and Hendrix MJ:
Molecular regulation of tumor cell vasculogenic mimicry by tyrosine
phosphorylation: Role of epithelial cell kinase (Eck/EphA2). Cancer
Res. 61:3250–3255. 2001.PubMed/NCBI
|
|
27
|
Wang H, Sun W, Zhang WZ, Ge CY, Zhang JT,
Liu ZY and Fan YZ: Inhibition of tumor vasculogenic mimicry and
prolongation of host survival in highly aggressive gallbladder
cancers by norcantharidin via blocking the ephrin type a receptor
2/focal adhesion kinase/paxillin signaling pathway. PLoS One.
9:e969822014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yuan W and Chen Z, Wu S, Ge J, Chang S,
Wang X, Chen J and Chen Z: Expression of EphA2 and E-cadherin in
gastric cancer: Correlated with tumor progression and lymphogenous
metastasis. Pathol Oncol Res. 15:473–478. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Hou F, Yuan W, Huang J, Qian L and Chen Z,
Ge J, Wu S, Chen J, Wang J and Chen Z: Overexpression of EphA2
correlates with epithelial-mesenchymal transition-related proteins
in gastric cancer and their prognostic importance for postoperative
patients. Med Oncol. 29:2691–2700. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Youngblood V, Wang S, Song W, Walter D,
Hwang Y, Chen J and Brantley-Sieders DM: Elevated Slit2 activity
impairs VEGF-induced angiogenesis and tumor neovascularization in
EphA2-deficient endothelium. Mol Cancer Res. 13:524–537. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Miao Z, Dong Y, Fang W, Shang D, Liu D,
Zhang K, Li B and Chen YH: VEGF increases paracellular permeability
in brain endothelial cells via upregulation of EphA2. Anat Rec
(Hoboken). 297:964–972. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Shen J, Xie B, Hatara CM, Hackett SF and
Campochiaro PA: Vegf or EphA2 antisense polyamide-nucleic acids;
vascular localization and suppression of retinal
neovascularization. Mol Ther. 15:1924–1930. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wu L, Zhang YS, Ye ML, Shen F, Liu W, Hu
HS, Li SW, Wu HW, Chen QH and Zhou WB: Overexpression and
correlation of HIF-2α, VEGFA and EphA2 in residual hepatocellular
carcinoma following high-intensity focused ultrasound treatment:
Implications for tumor recurrence and progression. Exp Ther Med.
13:3529–3534. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Huang M, Huang B, Li G and Zeng S:
Apatinib affect VEGF-mediated cell proliferation, migration,
invasion via blocking VEGFR2/RAF/MEK/ERK and PI3K/AKT pathways in
cholangiocarcinoma cell. BMC Gastroenterol. 18:1692018. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lu XS, Sun W, Ge CY, Zhang WZ and Fan YZ:
Contribution of the PI3K/MMPs/Ln-5γ2 and EphA2/FAK/Paxillin
signaling pathways to tumor growth and vasculogenic mimicry of
gallbladder carcinomas. Int J Oncol. 42:2103–2115. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wang H, Lin H, Pan J, Mo C, Zhang F, Huang
B, Wang Z, Chen X, Zhuang J, Wang D and Qiu S: Vasculogenic mimicry
in prostate cancer: The roles of EphA2 and PI3K. J Cancer.
7:1114–1124. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu Y, Li F, Yang YT, Xu XD, Chen JS, Chen
TL, Chen HJ, Zhu YB, Lin JY, Li Y, et al: IGFBP2 promotes
vasculogenic mimicry formation via regulating CD144 and MMP2
expression in glioma. Oncogene. 38:1815–1831. 2019. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Guo JQ, Zheng QH, Chen H, Chen L, Xu JB,
Chen MY, Lu D, Wang ZH, Tong HF and Lin S: Ginsenoside Rg3
inhibition of vasculogenic mimicry in pancreatic cancer through
downregulation of VE-cadherin/EphA2/MMP9/MMP2 expression. Int J
Oncol. 45:1065–1072. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kim HS, Won YJ, Shim JH, Kim HJ, Kim J,
Hong HN and Kim BS: Morphological characteristics of vasculogenic
mimicry and its correlation with EphA2 expression in gastric
adenocarcinoma. Sci Rep. 9:34142019. View Article : Google Scholar : PubMed/NCBI
|