Introduction
Ovarian cancer has the highest mortality of
reproductive system tumors; most patients have already progressed
to a late stage when diagnosed due to the high invasiveness and
recurrence of ovarian tumors (1–4).
Epithelial-mesenchymal transition (EMT) is the biological process
during which epithelial cells acquire abnormal motor forces due to
specific environmental changes or pathological factors, and
transform from an epithelial phenotype to an interstitial phenotype
(5,6). A previous study has demonstrated that
90% of ovarian cancers are epithelial and that EMT is associated
with the metastasis and recurrence of malignant tumors (7). Therefore, understanding EMT is crucial
for elucidating the mechanisms that induce ovarian cancer
metastasis.
Celecoxib is a non-steroidal anti-inflammatory drug
commonly used for the treatment of fever, pain, stiffness and
swelling (8). Previous studies have
confirmed that celecoxib regulates the cell cycle, promotes
apoptosis, inhibits angiogenesis and induces EMT in epithelial
cancers, such as lung cancer, thus affecting the development of
tumors (9,10). Artemisinin and its derivatives
(collectively termed as artemisinins) are among the most important
and effective antimalarial drugs. In addition to their antimalarial
effects, artemisinins have also been demonstrated to possess
selective anticancer properties. These effects appear to be
mediated by artemisinin-induced changes in multiple signaling
pathways, interfering simultaneously with multiple hallmarks of
cancer (11). Artemisinin exhibits
strong antitumor activity, although it is unclear whether
artemisinin can reverse EMT (11,12). In
the present study, an EMT model of ovarian cancer cell line SKOV3
cells induced by celecoxib was established in vitro;
subsequently, mesenchymal-epithelial transition (MET) was induced
by artemisinin. The effects of artemisinin on molecular biological
characteristics of the two models and EMT were analyzed. The
present study has laid a foundation for the study of genes and
signal transduction pathways that are affected by artemisinin
during the development and progression of ovarian cancer and is of
great clinical significance for future research of artemisinin as
an anti-ovarian cancer drug.
Materials and methods
Cell culture
The human ovarian epithelial adenocarcinoma cell
line SKOV3 was purchased from the Type Culture Collection of the
Chinese Academy of Sciences. SKOV3 cells were cultured in RPMI 1640
medium (HyClone; GE Healthcare Life Sciences) containing 10% FBS
(Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.), 50
µg/ml penicillin (HyClone; GE Healthcare Life Sciences), and 50
µg/ml streptomycin (HyClone; GE Healthcare Life Sciences) in a
humidified atmosphere at 37°C and 5% CO2, and the
culture medium was changed every three days.
Establishment of cell models
SKOV3 cells were suspended and cultured in 2 ml of
culture medium at the final concentration of 5×105
cells/ml. After the cells were stably anchored overnight, the
medium was discarded and 2 ml complete medium containing 10 µM
celecoxib (Shanghai Macklin Biochemical Co., Ltd.) was added to the
flask (10). Following incubation
with celecoxib for 12, 24, 36, 48 or 72 h at 37°C, an EMT model of
SKOV3 cells was established. Untreated SKOV3 cells were used as
controls. The EMT cell model with 48-h incubation period (highest
proliferation rate confirmed by cell viability assay) was selected
as the ‘control’ EMT model for subsequent experiments. Cells (i.e.
EMT model) were resuspended at a final concentration of
5×105 cells/ml in 6 ml, of which 2 ml were inoculated
into a new flask resulting in a total of three flasks. The cells
were incubated overnight, the culture medium was discarded and 20,
40 and 80 µM artemisinin (Melone Pharmaceutical Co., Ltd.) were
added to the culture flask with a total volume of 2 ml. A MET cell
model was established following 48-h incubation with
artemisinin.
Cell viability assay
Cell viability was assessed using a Cell Counting
Kit-8(CCK-8; Biosharp). Untreated SKOV3 control cells, cells
treated with 10 µM celecoxib for 48 h (EMT model) and further
treated with 20, 40 and 80 µM artemisinin for 48 h (MET cell model)
were harvested and resuspended at a final concentration of
5×103 cells/ml and seeded into 96-well plates.
Subsequently, 10 µl CCK-8 solution was added to each well, and the
plates were incubated at 37°C for 2 h. Optical density (OD) values
were measured at 450 nm using a microplate reader (Biotex, Inc.).
Cell viability was calculated as follows: Cell viability (%) = (OD
of experimental group - OD of blank group) / (OD of control
group-OD of blank group) × 100. The experiments were repeated at
least five times.
Wound healing assay
Cells from all models were plated in 6-well plates
at a density of 5×105 cells/well. When cell confluence
reached 90%, vertical scratches were formed in the cell layer by
using a 10 µl pipette tip. Floating cells were removed by washing
with PBS. Culture medium without FBS was added to the cells, and
cells were incubated at 37°C with 5% CO2 and saturated
humidity for 48 h. Cell migration was observed at 0 and 48 h. The
scratch-healing areas were calculated using ImageJ 1.8.0 (National
Institutes of Health) and the following formula: Scratch-healing
(%) = (initial scratch area-final scratch area)/initial scratch
area × 100. The experiments were repeated at least three times.
Protein extraction and western blot
analysis
The flasks with cells were washed three times with
ice cold PBS and total protein was harvested in RIPA lysis buffer
(Beyotime Institute of Biotechnology) containing PMSF (1:100). A
bicinchoninic acid protein assay kit (Beyotime Institute of
Biotechnology) was used to determine protein concentrations.
Proteins (30 µg) were separated by SDS-PAGE on 10% gels and
transferred to a PVDF membrane (Beyotime Institute of
Biotechnology). The membranes were blocked with 5% skimmed milk in
TBS containing Tween-20 (TBST) for 1 h at 37°C and incubated with
primary antibodies against vimentin (polyclonal; cat. no. ABP52699;
1:1,000; Abbkine Scientific Co., Ltd.) and E-cadherin (polyclonal;
cat. no. ABP51221; 1:1,000; Abbkine Scientific Co., Ltd.) overnight
at 4°C. Following washing three times with TBST, the membrane was
incubated with horseradish peroxidase-conjugated goat anti-rabbit
IgG antibody (cat. no. BL003A; 1:8,000; Biosharp; Beijing Lanjieke
Technology Co., Ltd.) at 37°C for 1 h. E-cadherin and vimentin were
visualized using an electrochemiluminescence chromogenic kit
(Beyotime Institute of Biotechnology) in a dark room. Densitometry
was performed using ImageJ 1.8.0. The experiments were repeated at
least three times.
Flow cytometric analysis
SKOV3 cells were plated in 6-well plates at a
density of 5×105 cells/well and allowed to attach
overnight prior to treatment (i.e. induction into EMT and MET
models). Cells were harvested, fixed with 4% paraformaldehyde
(Sigma-Aldrich; Merck KGaA) for 30 min at 25°C, and permeabilized
with 0.1% Triton X-100 (Sigma-Aldrich; Merck KGaA) in PBS for 20
min at room temperature. The cells were blocked with 5% goat serum
(Hangzhou Sijiqing Biological Engineering Materials Co., Ltd.) and
0.3% Triton X-100 in PBS for 30 min at 37°C, incubated with the
primary antibodies against vimentin and E-cadherin (polyclonal;
1:1,000; AbbkineScientific Co., Ltd.) for 30 min at 37°C and
further incubated with FITC-labeled goat anti-rabbit IgG (cat. no.
BL033A; 1:50; Biosharp; Beijing Lanjieke Technology Co., Ltd.) for
30 min at 37°C. Labeled cells were detected using a Cytomics FC 500
flow cytometer (BD Biosciences). The average fluorescence intensity
was analyzed using FlowJo 7.6 software (BD Biosciences). The
experiments were repeated at least three times.
Immunofluorescence staining and
confocal imaging
SKOV3 cells were plated onto coverslips in 6-well
plates at a density of 1×105 cells/well and allowed to
attach overnight prior to treatment. The coverslips were fixed with
4% paraformaldehyde for 30 min at room temperature and
permeabilized with 0.1% Triton X-100 in PBS at room temperature for
20 min. The coverslips were blocked in 5% goat serum and 0.3%
Triton X-100 in PBS at 37°C for 30 min, incubated with the primary
antibodies against vimentin (polyclonal; cat. no. ABP52699;
1:1,000; AbbkineScientific Co., Ltd.) and E-cadherin (polyclonal;
cat. no. ABP51221; 1:1,000; Abbkine Scientific Co., Ltd.) at 37°C
for 30 min and further incubated with FITC-labeled goat anti-rabbit
IgG (cat. no. BL033A; 1:50; Biosharp; Beijing Lanjieke Technology
Co., Ltd.) at 37°C for 30 min. DAPI (Beyotime Institute of
Biotechnology) was added to the coverslips and incubated in the
dark at room temperature for 10 min. The coverslips were washed
with PBS and mounted onto slides with glycerol:water (1:1). An
Olympus FV-1200MPE SHARE confocal microscope (Olympus Corporation)
was used to visualize the cells. Images were processed and
quantified using ImageJ. The experiments were repeated at least
three times.
Statistical analysis
Data are presented as the mean ± standard deviation.
SPSS software (version 16.0 for Windows; SPSS, Inc.) was used for
statistical analysis. Data were analyzed with one-way analysis of
variance and Student's t-test. The least significant difference
test was used as a post hoc test for comparison between multiple
groups. P<0.05 was considered to indicate a statistically
significant difference.
Results
SKOV3 cell proliferation was promoted
by celecoxib and suppressed by artemisinin
To assess the effect of celecoxib and artemisinin on
SKOV3 cell proliferation, cell viability was tested using CCK-8.
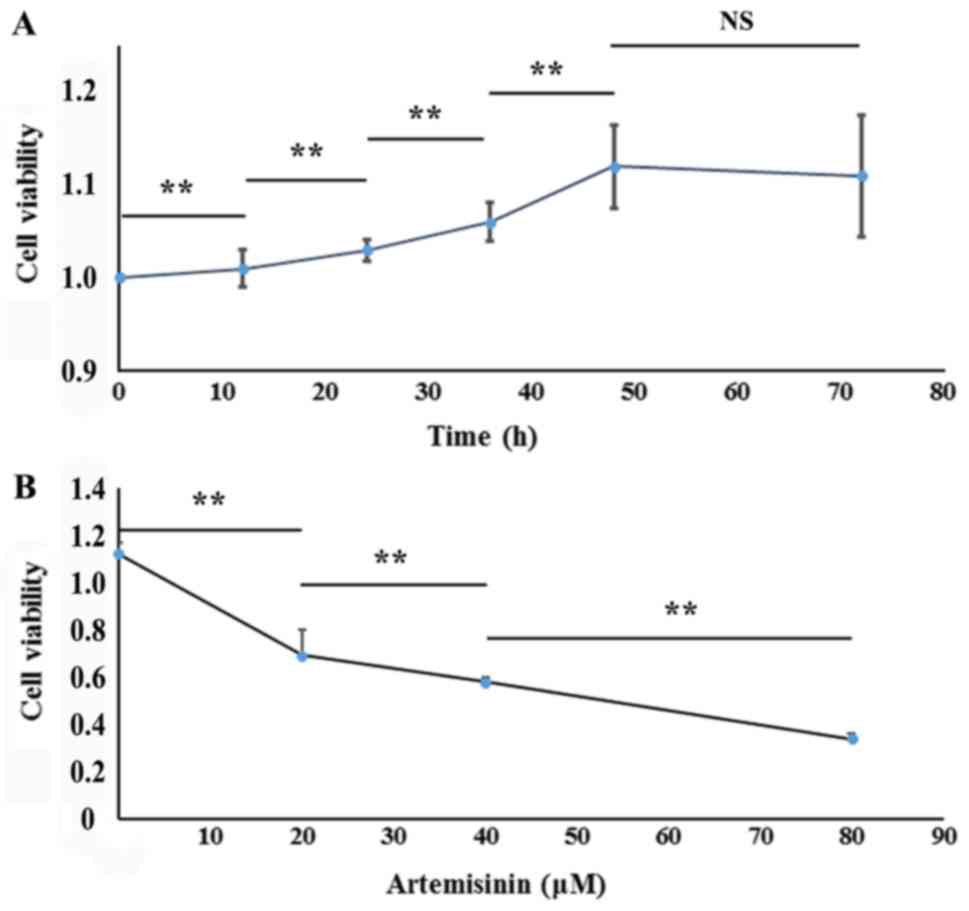
SKOV3 cells were incubated with 10 µM celecoxib for 0, 12, 24, 36,
48 and 72 h, which resulted in cell viability values of
1,1.01±0.02, 1.03±0.01, 1.06±0.02, 1.12±0.05, and 1.11±0.07,
respectively (Fig. 1A). SKOV3 cells
incubated with celecoxib for 48 h exhibited the highest viability;
thus, the 48 h incubation period was chosen as optimal for the EMT
cell model for subsequent experiments. The cell viability values of
the EMT and MET cell models (20, 40 and 80 µM artemisinin; 48 h)
were 1.12±0.05, 0.69±0.11, 0.58±0.39, 0.34±0.58, respectively
(P<0.01; Fig. 1B). These results
indicated that celecoxib promoted cell viability in a
time-dependent manner, whereas artemisinin inhibited cell viability
in a dose-dependent manner.
SKOV3 cell migratory ability was
promoted by celecoxib and inhibited by artemisinin
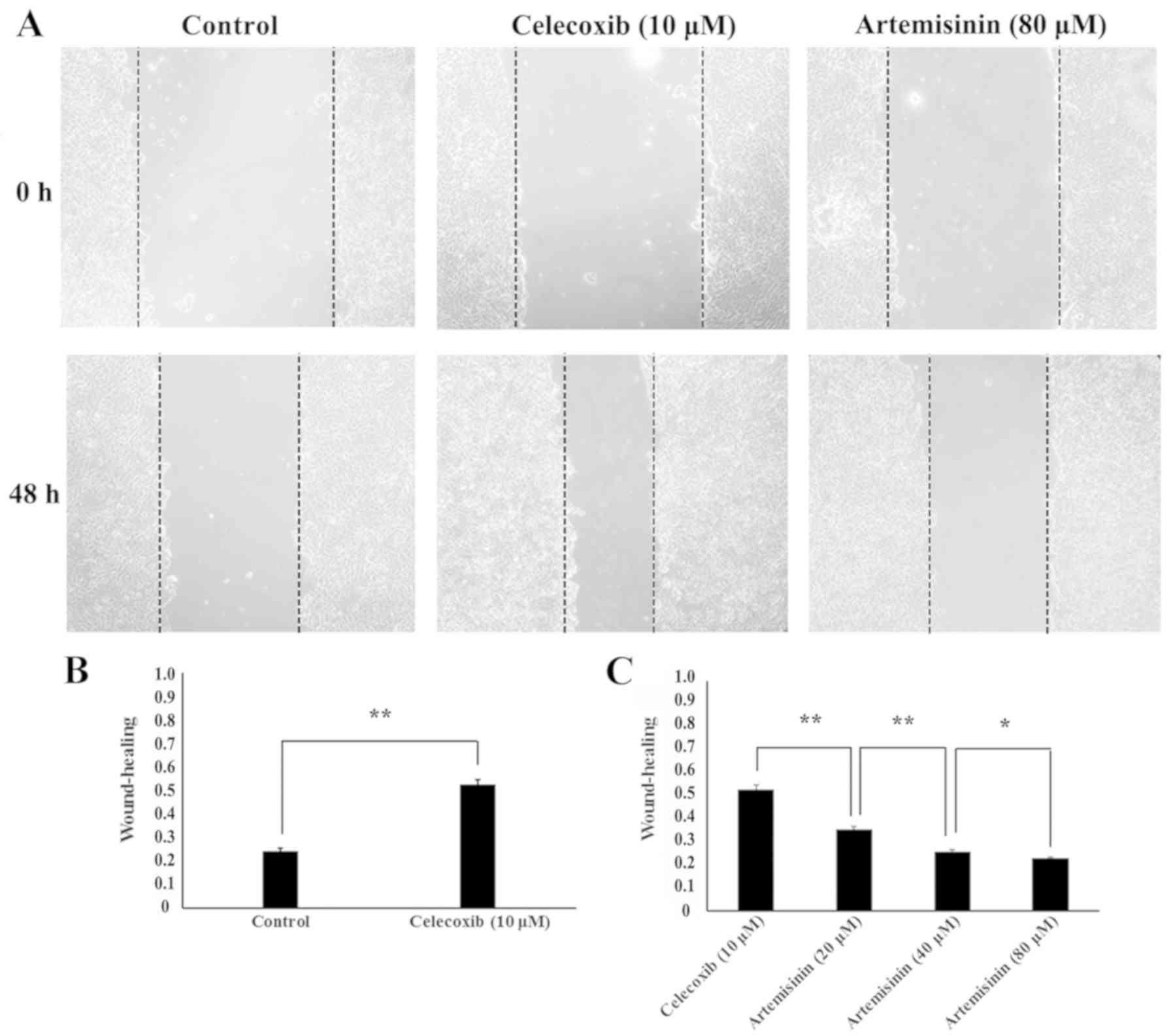
To evaluate the potential role of celecoxib and
artemisinin on the migratory ability of SKOV3 cells, wound healing
assay was performed. The scratch-healing rates were 0.25±0.01 in
the control group and 0.53±0.02 following 48-h 10 µM celecoxib
treatment, which indicated that cell migration was significantly
increased in the EMT model (P<0.01; Fig. 2B). Cell migration was significantly
decreased in the MET model (10 µM celecoxib for 48 h and 20, 40 and
80 µM artemisinin for 48 h) compared with the EMT model (P<0.05)
with scratch-healing rates of 0.36±0.01,0.26±0.01, and 0.23±0.01,
respectively (Fig. 2C). These
results indicated that, in SKOV3 cells, celecoxib may promote
migratory ability, whereas artemisinin may inhibit migratory
ability.
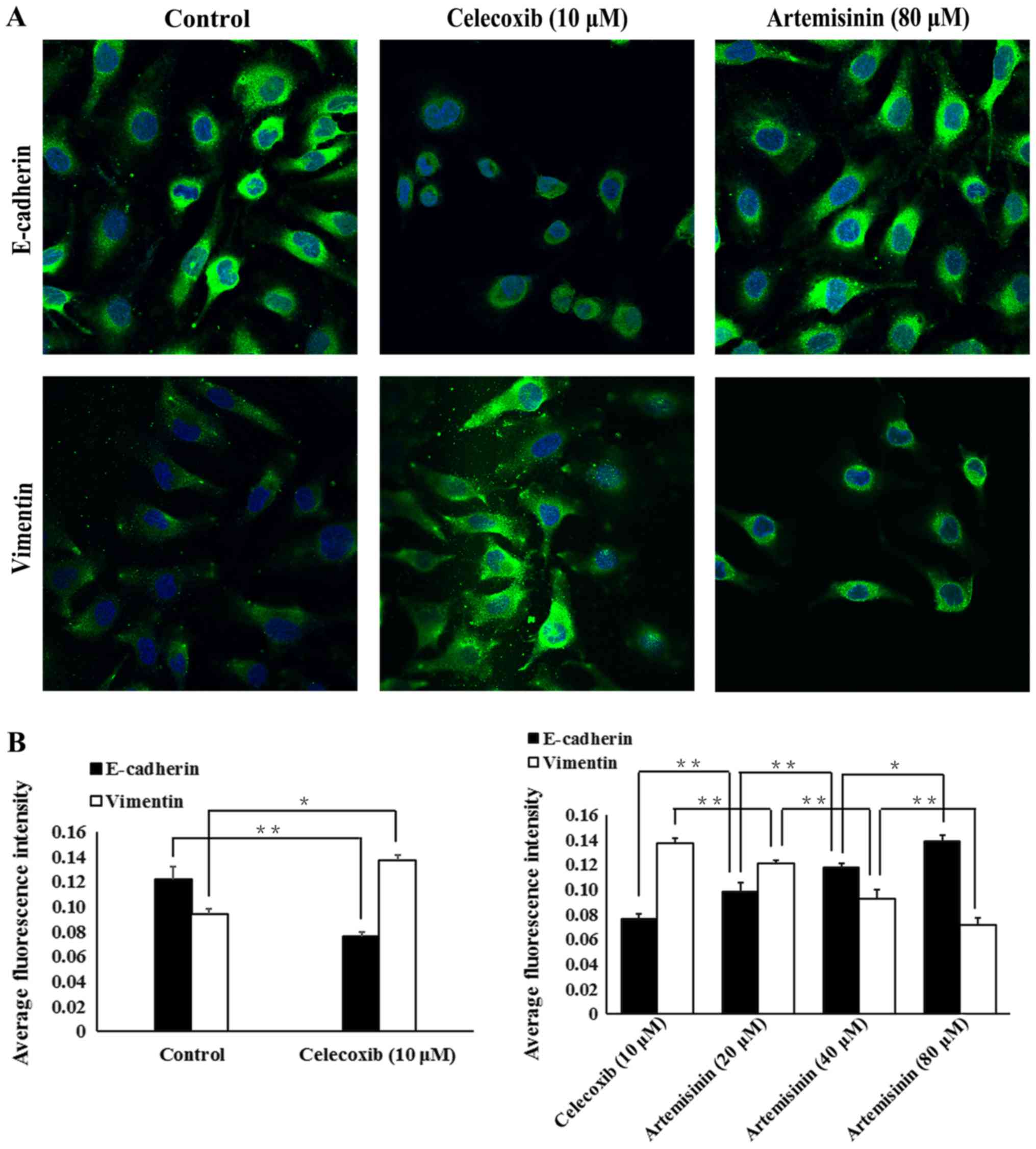
Celecoxib and artemisinin exhibit
opposite effects on E-cadherin and vimentin protein expression in
SKOV3 cells
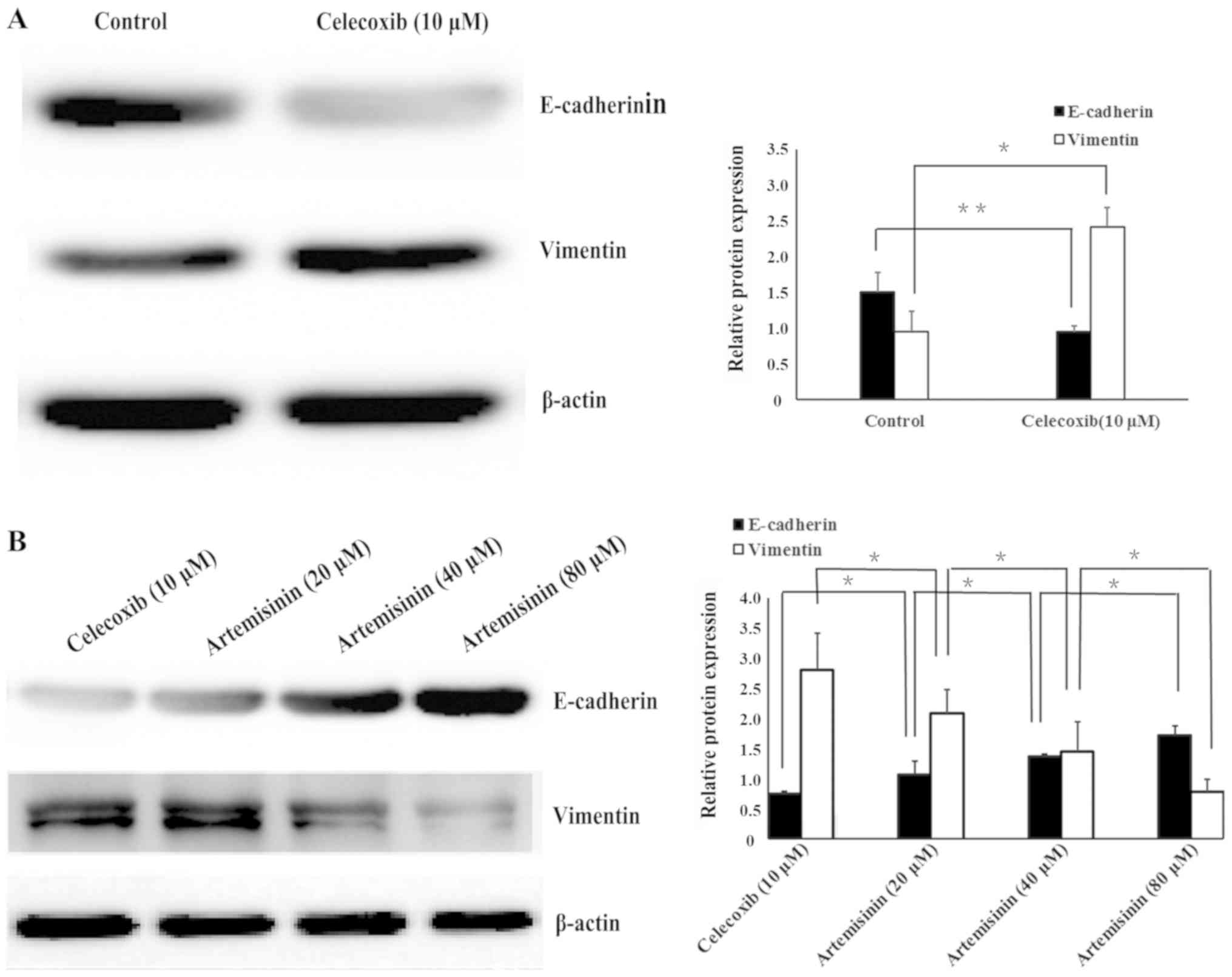
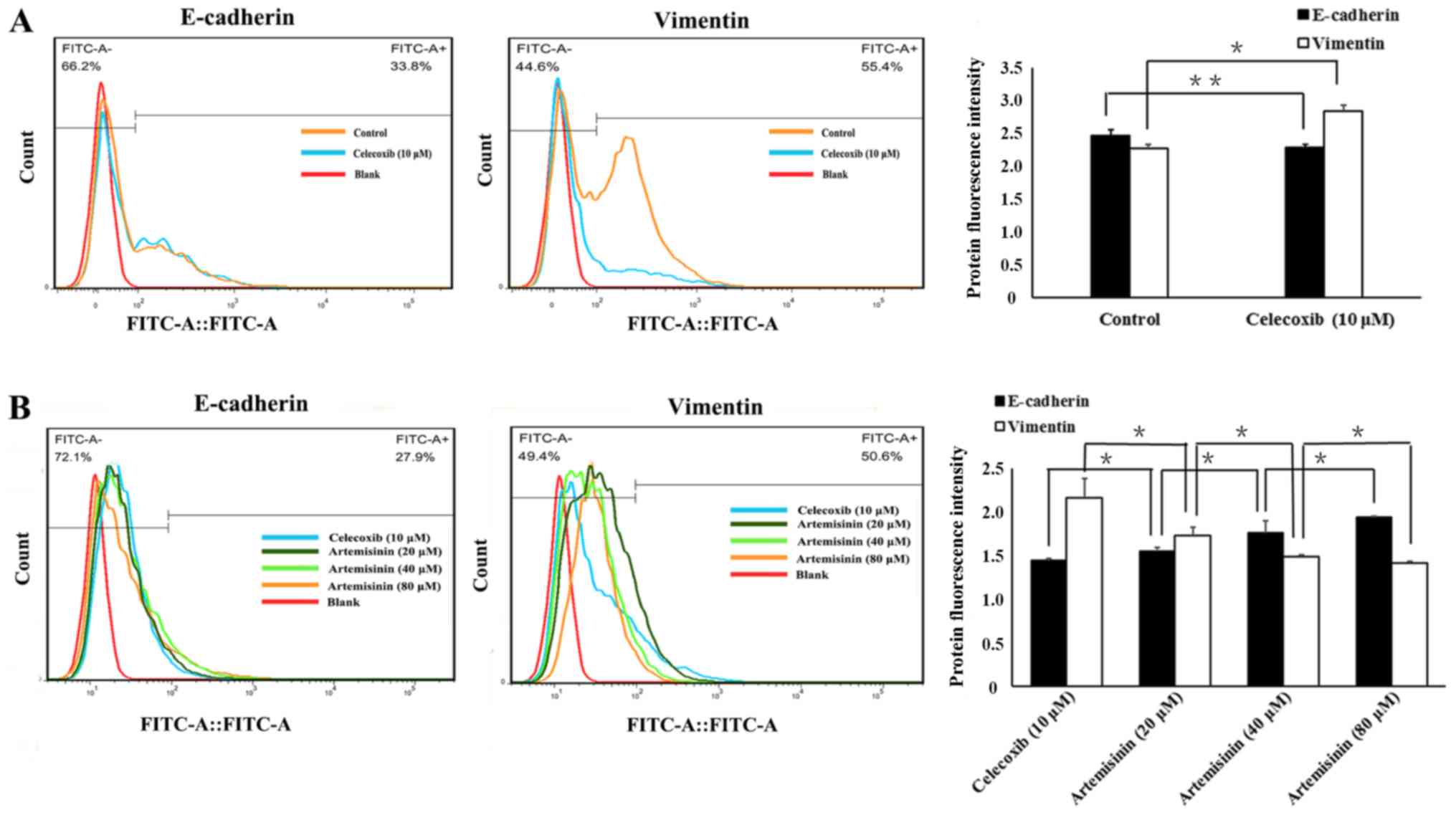
Western blotting (Fig.
3), flow cytometry (Fig. 4) and
immunofluorescence (Fig. 5) were
used to evaluate whether celecoxib and artemisinin affect
E-cadherin and vimentin expression in SKOV3 cells. The results
demonstrated that celecoxib reduced the expression of E-cadherin
and increased the expression of vimentin (Figs. 3A, 4A
and 5A), whereas artemisinin
increased the expression of E-cadherin and reduced the expression
of vimentin (Figs. 3B, 4B and 5B) in
a dose-dependent manner (P<0.05).
Discussion
Ovarian cancer the highest mortality rates amongst
gynecological malignancies. Metastasis, drug resistance, and
recurrence are important factors leading to patient mortality
(1). At present, the clinical
treatment of ovarian cancer is a multidisciplinary comprehensive
process, and cytoreductive surgery combined with platinum-based
chemotherapy is the current standard treatment (13–16).
However, the ovarian tissue of patients with ovarian cancer
gradually increases its tolerance to anticancer drugs, which
results in ineffective treatment (17). The selective cyclooxygenase-2 (COX-2)
inhibitor and non-steroidal anti-inflammatory drug celecoxib can
promote the growth, proliferation and migration of epithelial
cancer cells (10). Celecoxib
induces EMT and increases the cellular invasiveness in epithelial
ovarian cancer cells by activating the PI3K/AKT and
mitogen-activated protein kinase kinase/ERK signaling pathways, in
which zinc finger E-box binding homeobox 1is an important regulator
(10,18). Liu et al (9) demonstrated that 10 µM celecoxib
treatment in ovarian cancer A2780 and SKOV3 cells for 48 h
increased invasiveness compared with untreated ovarian cancer cells
independently of the COX-2 inhibitory activity of celecoxib;
however, the in vitro celecoxib concentration was higher
compared with the maximum blood concentrations in human. Celecoxib
used in the range of 5–15 µM, which is close to physiologically
relevant conditions, did not induce physiological damage to ovarian
cancer cells (9). Based on these
results, 10 µM celecoxib was chosen for use in the present study,
consistent with other references (19,20). In
addition, a clinical study has demonstrated that patients with
ovarian cancer with significantly increased COX-2 expression
exhibit significantly lower E-cadherin expression (21), which suggests that celecoxib may
induce EMT in ovarian cancer cells. In addition to celecoxib,
TGF-β1 is also a potential factor that affects various biological
activities such as proliferation, differentiation, and immune
regulation of cancer cells (19).
TGF-β1 has been identified as the inducing factor of EMT (19,22).
However, TGF-β1 stably induces EMT for a prolonged period of time
(~2 weeks) (23), whereas, celecoxib
induces EMT for only 48 h (19).
Artemisinin is an antimalarial drug that also exhibits anticancer
activity by inhibiting the proliferation, blocking the cell cycle
and promoting apoptosis in cancer cells (24). One study has demonstrated that
artemisinin and its derivatives have inhibitory effects on 68
different solid tumors and 24 blood-derived malignant tumors
(25). Consistent with these
results, the present study demonstrated that the EMT cell model
exhibited increased cell proliferation compared with untreated
SKOV3 cells, and that artemisinin decreased the cell
proliferation.
The invasive and metastatic ability of ovarian
cancer cells and drug resistance are associated with EMT (26–29). EMT
is a common pathological feature of gastric, colorectal and lung
cancer, as well as other malignant tumors (30). EMT affects the morphology of tumor
cells, which exhibit pleomorphism, numerous pseudopods, loss of
cell polarity and enlarged or multiple nuclei; such morphological
features contribute to cell invasion and migration (31). The expression of certain proteins in
tumor cells changes during the process of EMT; E-cadherin is a
molecular marker of epithelial cells that is localized at the cell
membrane, where its extracellular fragments form dimers with
adjacent cells, and its intracellular fragments form complexes with
related proteins to form an adhesion junction between tumor cells
(32,33). Vimentin is marker protein expressed
in interstitial cells that reduces adhesion between tumor cells,
and increases migration and invasiveness (32,33). In
the present study, EMT models exhibited increased vimentin
expression and decreased E-cadherin expression, whereas artemisinin
reduced vimentin expression and increased E-cadherin expression in
a dose-dependent manner. In addition, previous studies have
demonstrated that the artemisinin derivative, artesunate,
upregulates the expression of genes that inhibit invasion and
migration of tumor cells and can, thus, inhibit the invasion and
metastasis of renal cancer (34,35). The
present study used the wound healing assay to analyze the migration
of EMT and MET models and demonstrated that the scratch healing
rate of the EMT model was higher compared with control cells or the
MET model. These results suggested that artemisinin may reverse and
inhibit the EMT process of ovarian cancer cells in a dose-dependent
manner.
As a proprietary Chinese medicine, artemisinin is
abundant and inexpensive and exhibits low toxicity and high
efficiency. Artemisinin not only inhibits the proliferation of
cancer cells, but also reverses EMT of tumor cells, blocking their
activity (11). Although a number of
studies on the antitumor effect of artesunate with scorpion toxin
or camptothecin, and other chemotherapeutic drugs have been
published, these treatments have only been applied in vitro
or in animals and, thus, its clinical value remains to be further
explored (36,37). At present, only a small number of
studies have investigated the mechanisms of MET. Jeong et al
(34) demonstrated that artesunate
effectively inhibits the invasion and metastasis of renal cell
carcinoma in vivo and in vitro, and downregulates
focal adhesion kinase, epidermal growth factor receptor and
proto-oncogene c-Met; the sarcoma gene Src is also involved in
these processes. In embryonal rhabdomyosarcoma, artesunate
effectively inhibits the migration and invasion of tumor cells by
upregulating the expression of the cell adhesion molecules neural
cell adhesion molecule 1 and integrin β1 (35). Artesunate significantly inhibits the
migratory and invasive abilities of esophageal squamous cell
carcinoma KYSE-150 cells (38).
Artesunate can increase the adhesion of tumor cells and the
roughness of the cell membrane, as well as reduce the elasticity of
the cell membrane (38). In
addition, artesunate inhibits the migration of ovarian cancer cells
in vitro by inhibiting the TGF-β/WNT signaling pathway
(39). In vivo and in
vitro experiments in HepG2 and SMMC-7721 cells have revealed
that artemisinin significantly inhibits tumor invasion and
metastasis through a mechanism associated with upregulation of
E-cadherin and tissue inhibitor of metallopeptidase 2 expression
and downregulation of metallopeptidase (MMP) 2 expression (40). Dihydroartemisinin inhibited the
metastasis of human ovarian cancer HO8910PM cells by downregulating
the expression of MMP2 and MMP9, and inhibited EMT induced by
platinum-based drugs via the AKT/Snail signaling pathway (41,42).
The present study has established a basis for
further investigation of the molecular mechanism of artemisinin in
relation to EMT. The possibility that the reversal of EMT during
ovarian cancer (i.e. MET) may delay the occurrence of multidrug
resistance or even reverse drug resistance may be of great clinical
significance in the future.
Acknowledgements
Not applicable.
Funding
The present study was supported by grants from the
National Natural Science Foundation of China (grant no. 21707002),
the Anhui Provincial Natural Science Research General Project
(grant no. KJ2015B096by) and the Graduate School of Innovation,
Bengbu Medical College (grant no. Byycx1751).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
WL and LM conceived and planned the study. WL, JL
and XQ performed the experiments. HW and WL participated in the
model establishment. XL, YL and HZ analyzed the data. All authors
read and approved the manuscript and agreed to be accountable for
all aspects of the research in ensuring that the accuracy or
integrity of any part of the work are appropriately investigated
and resolved.
Ethics approval and consent to
participate
All experiments were performed in accordance with a
protocol approved by the Use Committee of Bengbu Medical
College.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Sureechatchaiyan P, Hamacher A, Brockmann
N, Stork B and Kassack MU: Adenosine enhances cisplatin sensitivity
in human ovarian cancer cells. Purinergic Signal. 14:395–408. 2018.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chen W, Zheng R, Zhang S, Zeng H, Zuo T,
Xia C, Yang Z and He J: Cancer incidence and mortality in China in
2013: An analysis based on urbanization level. Chin J Cancer Res.
29:1–10. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2018. CA Cancer J Clin. 68:7–30. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Li J, Shao W and Feng H: MiR-542-3p, a
microRNA targeting CDK14, suppresses cell proliferation,
invasiveness, and tumorigenesis of epithelial ovarian cancer.
Biomed Pharmacother. 110:850–856. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: Acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lu DH, Yang J, Gao LK, Min J, Tang JM, Hu
M, Li Y, Li ST, Chen J and Hong L: Lysine demethylase 2A promotes
the progression of ovarian cancer by regulating the PI3K pathway
and reversing epithelial-mesenchymal transition. Oncol Rep.
41:917–927. 2019.PubMed/NCBI
|
|
8
|
Dong J, Jiang D, Wang Z, Wu G, Miao L and
Huang L: Intra-articular delivery of liposomal
celecoxib-hyaluronate combination for the treatment of
osteoarthritis in rabbit model. Int J Pharm. 441:285–290. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Liu R, Zheng J, Li C, Pang Y, Zheng Q, Xu
X and Liu P: Celecoxib induces epithelial-mesenchymal transition in
epithelial ovarian cancer cells via regulating ZEB1 expression.
Arch Gynecol Obstet. 291:1361–1369. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wang ZL, Fan ZQ, Jiang HD and Qu JM:
Selective Cox-2 inhibitor celecoxib induces epithelial-mesenchymal
transition in human lung cancer cells via activating MEK-ERK
signaling. Carcinogenesis. 34:638–646. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wong YK, Xu C, Kalesh KA, He Y, Lin Q,
Wong WSF, Shen HM and Wang J: Artemisinin as an anticancer drug:
Recent advances in target profiling and mechanisms of action. Med
Res Rev. 37:1492–1517. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhang CJ, Wang J, Zhang J, Lee YM, Feng G,
Lim TK, Shen HM, Lin Q and Liu B: Mechanism-guided design and
synthesis of a mitochondria-targeting artemisinin analogue with
enhanced anticancer activity. Angew Chem Int Ed Engl.
55:13770–13774. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang X, Chen J, Sun L and Xu Y: SIRT1
deacetylates KLF4 to activate Claudin-5 transcription in ovarian
cancer cells. J Cell Biochem. 119:2418–2426. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Kreuzinger C, Gamperl M, Wolf A, Heinze G,
Geroldinger A, Lambrechts D, Boeckx B, Smeets D, Horvat R, Aust S,
et al: Molecular characterization of 7 new established cell lines
from high grade serous ovarian cancer. Cancer Lett. 362:218–228.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Liu MX, Siu MK, Liu SS, Yam JW, Ngan HY
and Chan DW: Epigenetic silencing of microRNA-199b-5p is associated
with acquired chemoresistance via activation of JAG1-Notch1
signaling in ovarian cancer. Oncotarget. 5:944–958. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bonneau C, Rouzier R, Geyl C, Cortez A,
Castela M, Lis R, Daraï E and Touboul C: Predictive markers of
chemoresistance in advanced stages epithelial ovarian carcinoma.
Gynecol Oncol. 136:112–120. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Livney YD and Assaraf YG: Rationally
designed nanovehicles to overcome cancer chemoresistance. Adv Drug
Deliv Rev. 65:1716–1730. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lim BJ, Jung SS, Choi SY and Lee CS:
Expression of metastasis-associated molecules in non-small cell
lung cancer and their prognostic significance. Mol Med Rep.
3:43–49. 2010.PubMed/NCBI
|
|
19
|
Cha BK, Kim YS, Hwang KE, Cho KH, Oh SH,
Kim BR, Jun HY, Yoon KH, Jeong ET and Kim HR: Celecoxib and
sulindac inhibit TGF-β1-induced epithelial-mesenchymal transition
and suppress lung cancer migration and invasion via downregulation
of sirtuin 1. Oncotarget. 7:57213–57227. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bocca C, Bozzo F, Cannito S, Parola M and
Miglietta A: Celecoxib inactivates epithelial-mesenchymal
transition stimulated by hypoxia and/or epidermal growth factor in
colon cancer cells. Mol Carcinog. 51:783–795. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang YP, Wang QY, Li CH and Li XW: COX-2
inhibition by celecoxib in epithelial ovarian cancer attenuates
E-cadherin suppression through reduced Snail nuclear translocation.
Chem Biol Interact. 292:24–29. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Li Z, Hou P, Fan D, Dong M, Ma M, Li H,
Yao R, Li Y, Wang G, Geng P, et al: The degradation of EZH2
mediated by lncRNA ANCR attenuated the invasion and metastasis of
breast cancer. Cell Death Differ. 24:59–71. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Pang MF, Georgoudaki AM, Lambut L,
Johansson J, Tabor V, Hagikura K, Jin Y, Jansson M, Alexander JS,
Nelson CM, et al: TGF-β1-induced EMT promotes targeted migration of
breast cancer cells through the lymphatic system by the activation
of CCR7/CCL21-mediated chemotaxis. Oncogene. 35:748–760. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li X, Zhou Y, Liu Y, Zhang X, Chen T, Chen
K, Ba Q, Li J, Liu H and Wang H: Preclinical efficacy and safety
assessment of artemisinin-chemotherapeutic agent conjugates for
ovarian cancer. EBioMedicine. 14:44–54. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hooft van Huijsduijnen R, Guy RK, Chibale
K, Haynes RK, Peitz I, Kelter G, Phillips MA, Vennerstrom JL,
Yuthavong Y and Wells TN: Anticancer properties of distinct
antimalarial drug classes. PLoS One. 8:e829622013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Fang D, Chen H, Zhu JY, Wang W, Teng Y,
Ding HF, Jing Q, Su SB and Huang S: Epithelial-mesenchymal
transition of ovarian cancer cells is sustained by Rac1 through
simultaneous activation of MEK1/2 and Src signaling pathways.
Oncogene. 36:1546–1558. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee HM, Hwang KA and Choi KC: Diverse
pathways of epithelial mesenchymal transition related with cancer
progression and metastasis and potential effects of endocrine
disrupting chemicals on epithelial mesenchymal transition process.
Mol Cell Endocrinol. 457:103–113. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu Y, Wang C, Su J, Xie Q, Ma L, Zeng L,
Yu Y, Liu S, Li S, Li Z and Sun L: Tolerance to endoplasmic
reticulum stress mediates cisplatin resistance in human ovarian
cancer cells by maintaining endoplasmic reticulum and mitochondrial
homeostasis. Oncol Rep. 34:3051–3060. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Yang SH, Sharrocks AD and Whitmarsh AJ:
MAP kinase signalling cascades and transcriptional regulation.
Gene. 513:1–13. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mak MP, Tong P, Diao L, Cardnell RJ,
Gibbons DL, William WN, Skoulidis F, Parra ER, Rodriguez-Canales J,
Wistuba II, et al: A Patient-derived, pan-cancer EMT signature
identifies global molecular alterations and immune target
enrichment following epithelial-to-mesenchymal transition. Clin
Cancer Res. 22:609–620. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xia L, Zhang B, Yan Q and Ruan S: Effects
of saponins of patrinia villosa against invasion and metastasis in
colorectal cancer cell through NF-κB signaling pathway and EMT.
Biochem Biophys Res Commun. 503:2152–2159. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, Huang P, Liu X, Xiang Y, Zhang T,
Wu Y, Xu J, Sun Z, Zhen W, Zhang L, et al: Polyphyllin I inhibits
growth and invasion of cisplatin-resistant gastric cancer cells by
partially inhibiting CIP2A/PP2A/Akt signaling axis. J Pharmacol
Sci. 137:305–312. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Po JW, Roohullah A, Lynch D, DeFazio A,
Harrison M, Harnett PR, Kennedy C, de Souza P and Becker TM:
Improved ovarian cancer EMT-CTC isolation by immunomagnetic
targeting of epithelial EpCAM and mesenchymal N-cadherin. J Circ
Biomark. 7:18494544187826172018. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jeong DE, Song HJ, Lim S, Lee SJ, Lim JE,
Nam DH, Joo KM, Jeong BC, Jeon SS, Choi HY and Lee HW: Repurposing
the anti-malarial drug artesunate as a novel therapeutic agent for
metastatic renal cell carcinoma due to its attenuation of tumor
growth, metastasis, and angiogenesis. Oncotarget. 6:33046–33064.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Beccafico S, Morozzi G, Marchetti MC,
Riccardi C, Sidoni A, Donato R and Sorci G: Artesunate induces ROS-
and p38 MAPK-mediated apoptosis and counteracts tumor growth in
vivo in embryonal rhabdomyosarcoma cells. Carcinogenesis.
36:1071–1083. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang L, Chen F, Zhang Z, Chen Y and Wang
J: Synthesis and biological evaluation of a novel
artesunate-podophyllotoxin conjugate as anticancer agent. Bioorg
Med Chem Lett. 26:38–42. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Q, Wang W, Liu Y, Lian B, Zhu Q, Yao L
and Liu T: The biological characteristics of a novel
camptothecin-artesunate conjugate. Bioorg Med Chem Lett.
25:148–152. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shi R, Cui H, Bi Y, Huang X, Song B, Cheng
C, Zhang L, Liu J, He C, Wang F, et al: Artesunate altered cellular
mechanical properties leading to deregulation of cell proliferation
and migration in esophageal squamous cell carcinoma. Oncol Lett.
9:2249–2255. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Marchion DC, Xiong Y, Chon HS, Al Sawah E,
Bou Zgheib N, Ramirez IJ, Abbasi F, Stickles XB, Judson PL, Hakam
A, et al: Gene expression data reveal common pathways that
characterize the unifocal nature of ovarian cancer. Am J Obstet
Gynecol. 209:576.e1–576.e16. 2013. View Article : Google Scholar
|
|
40
|
Weifeng T, Feng S, Xiangji L, Changqing S,
Zhiquan Q, Huazhong Z, Peining Y, Yong Y, Mengchao W, Xiaoqing J
and Wan-Yee L: Artemisinin inhibits in vitro and in vivo invasion
and metastasis of human hepatocellular carcinoma cells.
Phytomedicine. 18:158–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wu B, Hu K, Li S, Zhu J, Gu L, Shen H,
Hambly BD, Bao S and Di W: Dihydroartiminisin inhibits the growth
and metastasis of epithelial ovarian cancer. Oncol Rep. 27:101–108.
2012.PubMed/NCBI
|
|
42
|
Qin Y, Yang G, Li M, Liu HJ, Zhong WL, Yan
XQ, Qiao KL, Yang JH, Zhai DH, Yang W, et al: Dihydroartemisinin
inhibits EMT induced by platinum-based drugs via Akt-Snail pathway.
Oncotarget. 8:103815–103827. 2017. View Article : Google Scholar : PubMed/NCBI
|