Introduction
Mitochondria are recognized as the primary sites of
aerobic oxidation of metabolic fuels in eukaryotic cells (1). It is well established that mitochondria
provide the majority of cellular energy in the form of ATP through
electron transport chains and an oxidative phosphorylation system
(2,3). According to a previous study,
mitochondria contribute to various pivotal cell functions,
including nitrogen metabolism, pyruvate and fatty acid oxidation
and heme biosynthesis (4).
Mitochondria have their own genomic DNA, making them unique among
eukaryotic extranuclear organelles; and the mitochondrial genome is
a circular double-stranded DNA (5).
The mitochondrial transcription termination factor
(MTERF) family members serve important roles in mitochondrial gene
expression (6). The MTERF family
consists of diverse members, including MTERF1, 2, 3 and 4 (7,8). MTERFs,
located in the mitochondria, contain a modular architecture based
on repetitions of a 30 amino acid module (8). A previous study indicated that MTERFs
control mitochondrial DNA (mtDNA) replication and exert important
functions in mitochondria transcription termination and initiation
(9). The differences in
mitochondrial gene organization have been demonstrated to be
associated with a number of functions (8). Among the MTERF family members, MTERF1
is the canonical mitochondrial transcription terminator and is able
to modulate the expression of mitochondrial genes by preventing
L-strand transcription interference within mtDNA (8). However, MTERF1 is dispensable for
ribosomal RNA gene transcription regulation (10). A previous investigation also revealed
that defects in MTERF1 binding may lead to mitochondrial diseases,
including Kearns-Sayre syndrome (11). In post-transcriptional modification
patterns of mitochondrial genes, a mitochondrial encephalomyopathy,
lactic acidosis, and stroke-like episodes (MELAS) mutation in the
mtDNA binding-site for the transcription termination factor was
revealed to cause defects in protein-synthesis in respiration;
however, it does not alter the upstream and downstream mature
transcription levels (12). MTERF2
is highly expressed in liver, heart and skeletal muscle cells
(13). Previous experiments in mice
demonstrated that, when fed on a ketogenic diet (i.e. high fat and
low carbohydrate diet), MTERF2-deficient mice developed myopathy
and memory deficits (14). In the
absence of MTERF2, mtDNA transcription is markedly decreased and a
respiratory defect occurs in neurons (14). MTERF3 is another member of the MTERF
family and is essential for the development of embryos in mice
(15). MTERF4 contributes to the
regulation of mitochondrial translation by targeting NOP2/sun RNA
methyltransferase family member 4 (NSUN4) to large mitochondrial
ribosome, and MTERF4-knockout also leads to mouse embryo death
(16). A previous study also
provided a novel insight into the association between MTERF4 and
MPP+-induced mitochondrial damage (17). In addition, MTERF4 has been
hypothesized to be one of the triggering genes for Parkinson's
disease, which is induced by an environmental toxin (17). Neurological disorders, including
Parkinson's disease and Alzheimer's disease, and also ageing,
diabetes and cancer are associated with mitochondrial dysfunction
resulting from abnormal variations in mitochondrial gene expression
(18,19).
As one of the major causes of mortality worldwide,
lung cancer (LC) is primarily characterized as small cell lung
cancer and non-small cell lung cancer (NSCLC). Over 80% of patients
with LC are characterized as having NSCLC (19). Typically, NSCLC comprises of lung
squamous cell carcinoma (LUSC) and lung adenocarcinoma (LUAD)
(20). A number of studies have
suggested that mitochondrial functions are involved in the
development of LC, particularly mitochondrial dysfunction may lead
to an alternation in bioenergetics and genomic instability in LC
(21). Additionally, a previous
study indicated that mitochondrial fission and fusion are involved
in metabolic function, proliferation and cell survival during the
progression of LC (22). Recently,
various novel approaches and techniques have been used to identify
and investigate novel biomarkers and potential molecular mechanism
in various types of cancer (23–26),
including LC and renal cancer. However, there are >8.2 million
LC-associated mortalities worldwide each year and the mortality
rates of LC remain at unacceptably high levels (20). Inspired by the aforementioned
previous studies, the present study aimed to investigate potential
biological associations between mitochondrial gene expression and
the pathogenesis of NSCLC. MTERFs are understood to regulate
mitochondrial functions through modulating the mitochondrial gene
transcription and protein expression (6,9,10); however, to the best of our knowledge,
there are no reports regarding the prognostic roles of MTERFs in
patients with NSCLC. In addition, the role of MTERFs in NSCLC
remain elusive; therefore, the present study aimed to investigate
the expression and prognostic values of MTERFs in patients with
NSCLC.
Materials and methods
Collection of human lung cancer
tissues and ethics statement
Lung cancer tissues and paired adjacent non-tumor
normal lung tissues from patients with NSCLC were obtained at the
Jilin Province Cancer Hospital (Changchun, China) between January
2012 and December 2012, as described previously (27). The patients included 5 males and 4
females with a mean age of 55.5 years (age range, 45–73 years), A
total of 9 tumor and 9 non-tumor tissue samples were collected from
these patients using video assisted thoracoscopic (VATS) lobectomy.
During surgery, NSCLC and adjacent tissues (5 cm from cancerous
tissues) were obtained and subsequently stored at −80°C.
Pathological data were studied prospectively following surgery.
Cancer tissue specimens were diagnosed by pathology, and adjacent
tissues were diagnosed as no cancer or inflammatory cell
infiltration. All fresh tissues were stored at −80°C for subsequent
experiments. The present study was approved by the Ethics Committee
of Jilin Province Cancer Hospital. Written informed consent was
obtained from all participants prior to enrollment in the present
study. The study protocols were performed based on the Declaration
of Helsinki.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis of gene
expression
Total RNA from human lung tissue was extracted using
TRIzol® reagent (Invitrogen; Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Total RNA (1 µg) was reverse-transcribed
into cDNA using a reverse transcription kit (iScript™ Reverse
Transcription Supermix for RT-qPCR; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Briefly, the temperature protocol for cDNA
synthesis was as follows: 5 min at 25°C, 20 min at 46°C and 1 min
at 96°C. RT-qPCR analysis of gene expression was performed using
the following primers: MTERF1 forward, 5′-TTGGATGACTCGATTTTCAGCA-3′
and reverse, 5′-GCTGTCGTTTCCTTGCCAT-3′; MTERF2 forward,
5′-TTCAGAAAGATGCGATCACCTC-3′ and reverse,
5′-TCATCGGCACCTAGTTCTTGT-3′; MTERF3 forward,
5′-GCTTTGTCAGCCCAACAGATA-3′ and reverse,
5′-CTTGAGCTAGTAGACTGGGAACT-3′; MTERF4 forward,
5′-GCCTGTATGGCTAGGCAGAC-3′ and reverse,
5′-GGAGGCTGTAGTCAGTTTGCG-3′; and GAPDH forward,
5′-ACAACTTTGGTATCGTGGAAGG-3′ and reverse,
5′-GCCATCACGCCACAGTTTC-3′. qPCR analysis was performed using SYBR
Premix Ex Taq (Takara Bio, Inc., Otsu, Japan). PCR products were
monitored with an Mx3000P QPCR system (Agilent Technologies, Inc.,
Santa Clara, CA, USA). The thermocycling conditions were as
follows: 10 sec at 95°C, 40 cycles of 5 sec at 95°C, and 30 sec at
60°C. The mRNA expression levels were analyzed using the
2−ΔΔCq method (28) with
the Mx3000P QPCR system, and the levels of MTERF1-4 were normalized
to the constitutive expression level of GAPDH mRNA.
Gene alteration frequency and mRNA
expression in NSCLC from the Gene Expression Omnibus (GEO)
database
The present study analyzed the gene alteration
frequency of MTERF1-4 in patients with NSCLC from The Cancer Genome
Atlas (TCGA) database (http://cancergenome.nih.gov) and the PubMed database
(https://www.ncbi.nlm.nih.gov/pubmed)
using the cBioportal for cancer genomics analysis (http://www.cbioportal.org) (29). The present study also analyzed the
mRNA expression levels of MTERF1-4 in NSCLC using the GEO database
(http://www.ncbi.nlm.nih.gov/geo). A
total of two original datasets (GSE10072 and GSE19804) (30) for patients with LC were downloaded
from GEO. The mRNA expression levels of MTERF1-4 in NSCLC samples
and adjacent non-tumor tissues were used for subsequent
analysis.
Prognostic values of MTERFs in
patients with NSCLC
In order to evaluate the prognostic roles of MTERFs
in patients with NSCLC, the present study used public databases to
analyze the expression of these genes and the overall survival rate
(OS) of 1,926 patients with NSCLC, who completed follow-up for 20
years. The data used for analysis were pooled from GEO, TCGA,
European Genome-phenome Archive (https://ega.crg.eu/) and PubMed (http://www.pubmed.com)(31). To obtain Kaplan-Meier
plots for the patients OS rate, the Kaplan-Meier plotter (KM
plotter) database (http://kmplot.com/analysis/index.php?p=service&cancer=lung)
was used. According to the median expression level of all patients
with NSCLC included in the analysis, the patients were divided into
two groups: High expression group and low expression group. Hazard
ratios (HRs), 95% confidence intervals (CIs) and log-rank P-values
were also calculated from the database. P<0.05 was considered to
indicate a statistically significant difference (32). The following Affymetrix IDs were
valid: 204871_at (MTERF1), 225346_at (MTERF2), 219363_s_at (MTERF3)
and 1557965_at (MTERF4).
Statistical analysis
RT-qPCR assays were analyzed from a minimum of three
independent experiments. Data were analyzed using a paired,
two-tailed Student' t-test or one-way ANOVA using GraphPad Prism
version 6 (GraphPad Software, Inc., La Jolla, CA, USA). Data are
presented as the mean ± standard error of the mean (32). P<0.05 was considered to indicate a
statistically significant difference.
Results
MTERFs gene alteration frequency in
NSCLC
The present study initially analyzed the gene
alteration frequency of MTERFs in NSCLC tissues using the
cBioportal database. As presented in Fig. 1, high amplification rates of MTERF1
and MTERF3 were observed in NSCLC (Fig.
1A and C). MTERF2 was revealed to exhibit a high deep deletion
rate in patients with LUAD and a high mutation rate in patients
with LUSC (Fig. 1B). In addition,
MTERF4 exhibited a high mutation and high deep deletion rate in
patients with LUSC (Fig. 1D). The
mutation of all MTERF genes primarily occurred in the same region,
which contains three leucine zipper sequences (data not shown); and
the leucine zipper sequences are critical for the enzyme functions
(33).
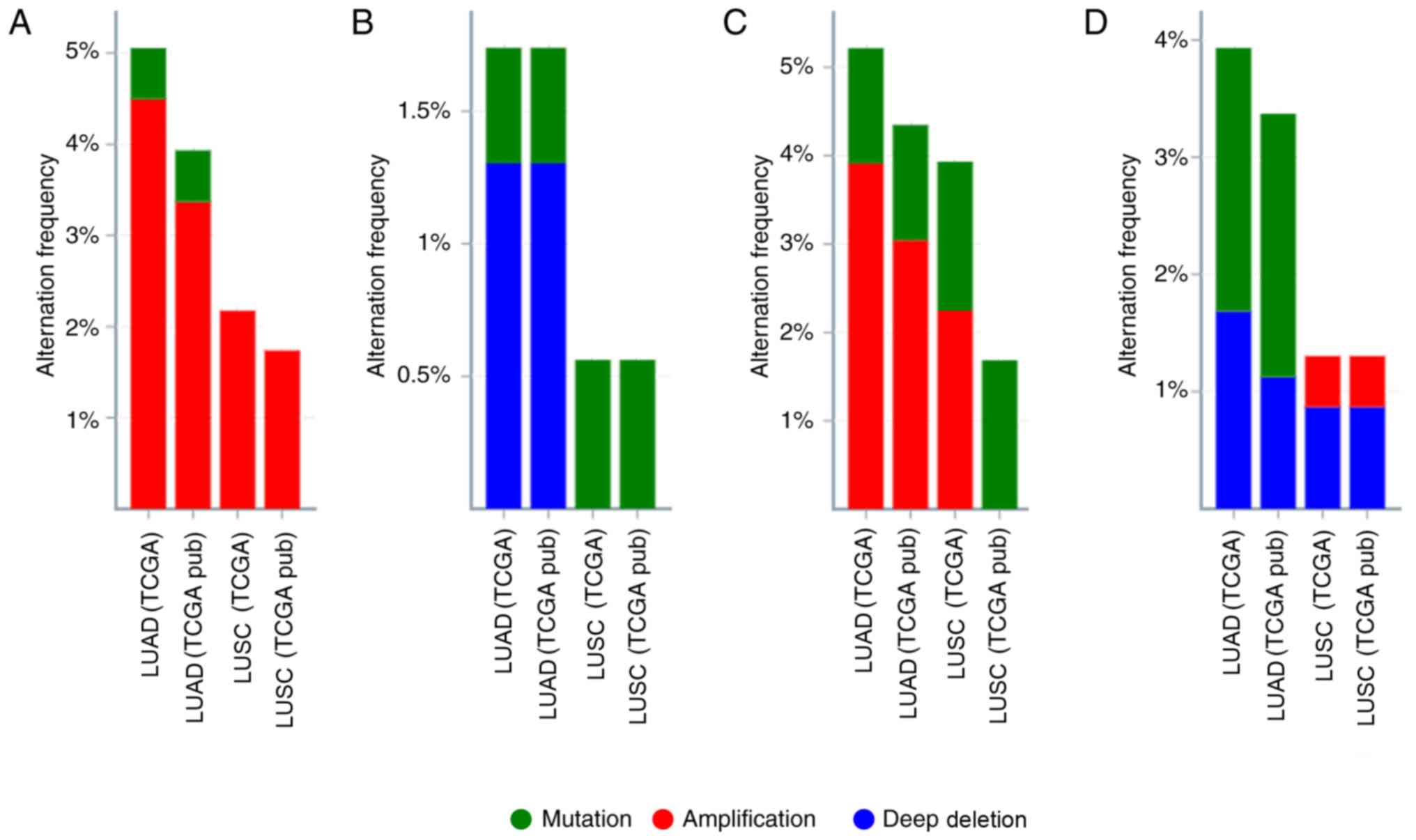 | Figure 1.Alteration frequency of MTERF1-4 gene
expression in NSCLC. The alterations of MTERF1, 2, 3 and 4 gene
expressions were visualized using the cBioPortal for Cancer
Genomics database. Mutations, deletions and amplifications are
presented in different colors. Alteration frequencies of (A)
MTERF1, (B) MTERF2, (C) MTERF3 and (D) MTERF4 gene expression in
NSCLC according to TCGA database. NSCLC, non-small cell lung
cancer; TCGA, The Cancer Genome Atlas; MTERF, mitochondrial
transcription termination factor; pub, PubMed; LUAD, lung
adenocarcinoma; LUSC, lung squamous cell carcinoma. |
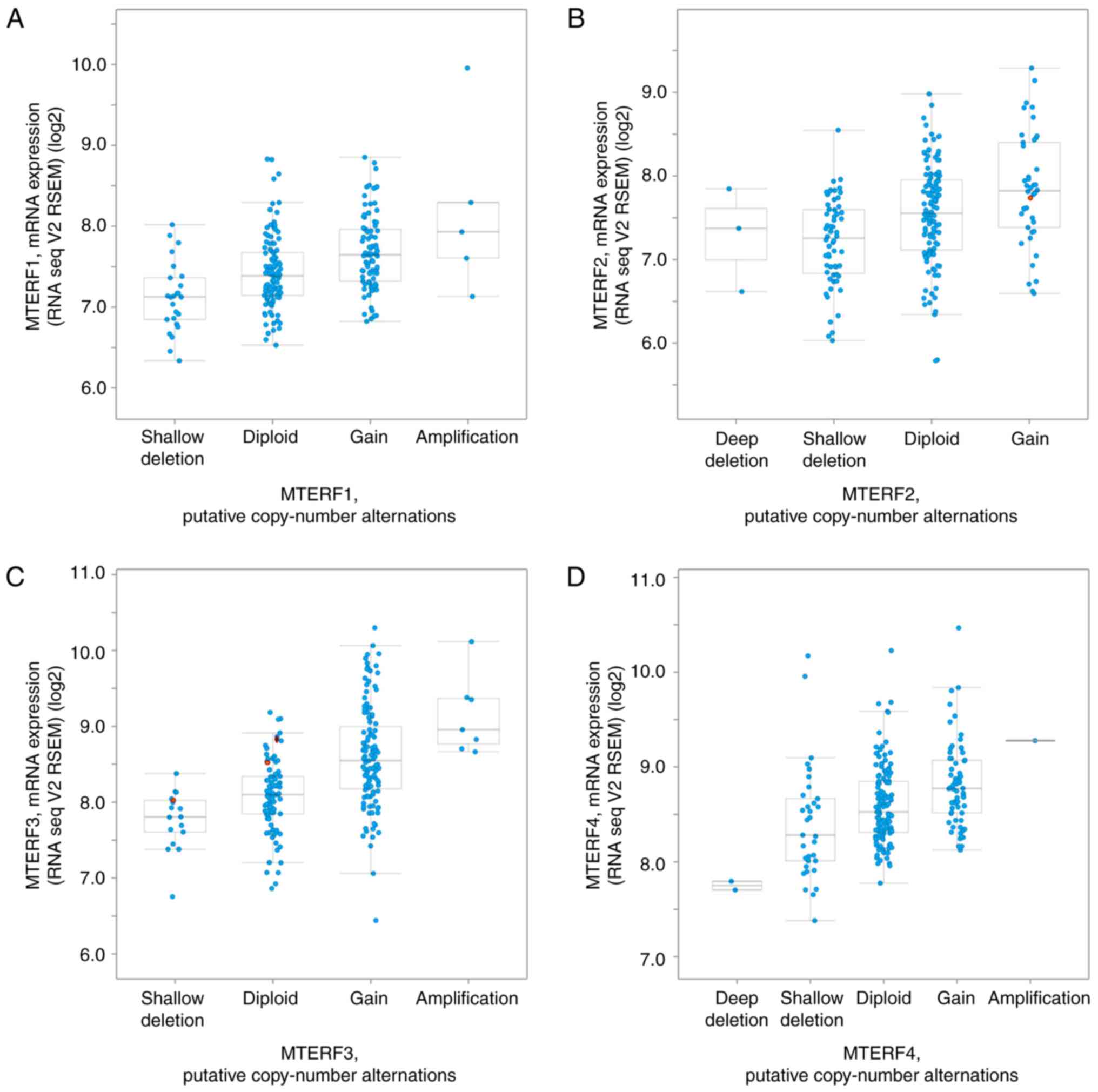
Expression of MTERFs in NSCLC
The present study subsequently analyzed the mRNA
expression of MTERFs and its correlation with the copy numbers
using cBioportal for cancer genomics analysis (http://www.cbioportal.org) as previously described
(28). As illustrated in Fig. 2, the mRNA levels of MTERF1, 2, 3 and
4 were positively associated with the copy number of these genes in
patients with LUAD (Fig. 2). To
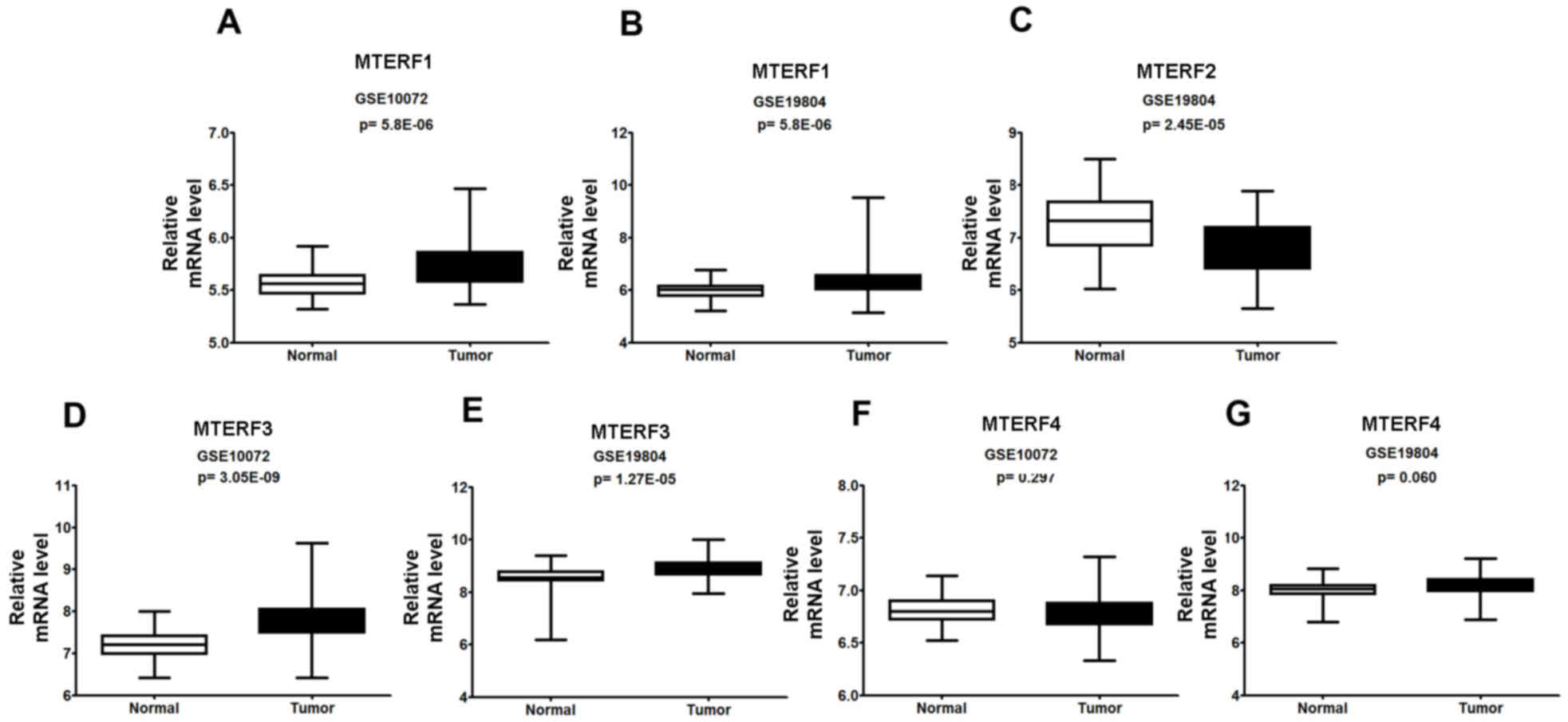
investigate the mRNA expression of MTERFs in NSCLC and normal
tissues, the raw data was retrieved by searching terms ‘GSE#19804′
and ‘GSE#10072’ from the GEO dataset. Additionally, the present
study analyzed the mRNA expression of MTERF1-4 by RT-qPCR in the
fresh isolated NSCLC tissues, which were collected at Jilin
Province Cancer Hospital in 2012, as described previously (27). The subsequent analysis indicated that
the mRNA expression levels of MTERF1 (Fig. 3A and B) and MTERF3 (Fig. 3D and E) were significantly higher
(P<0.001) in NSCLC tissues compared with paired adjacent
non-tumor tissues from micro-array analysis. The mRNA expression of
MTERF2 was significantly decreased in lung tumor tissue (P<0.05;
Fig. 3C). However, the expression of
MTERF4 was not significantly changed in NSCLC tissues when compared
with adjacent non-tumor tissues (Fig. 3F
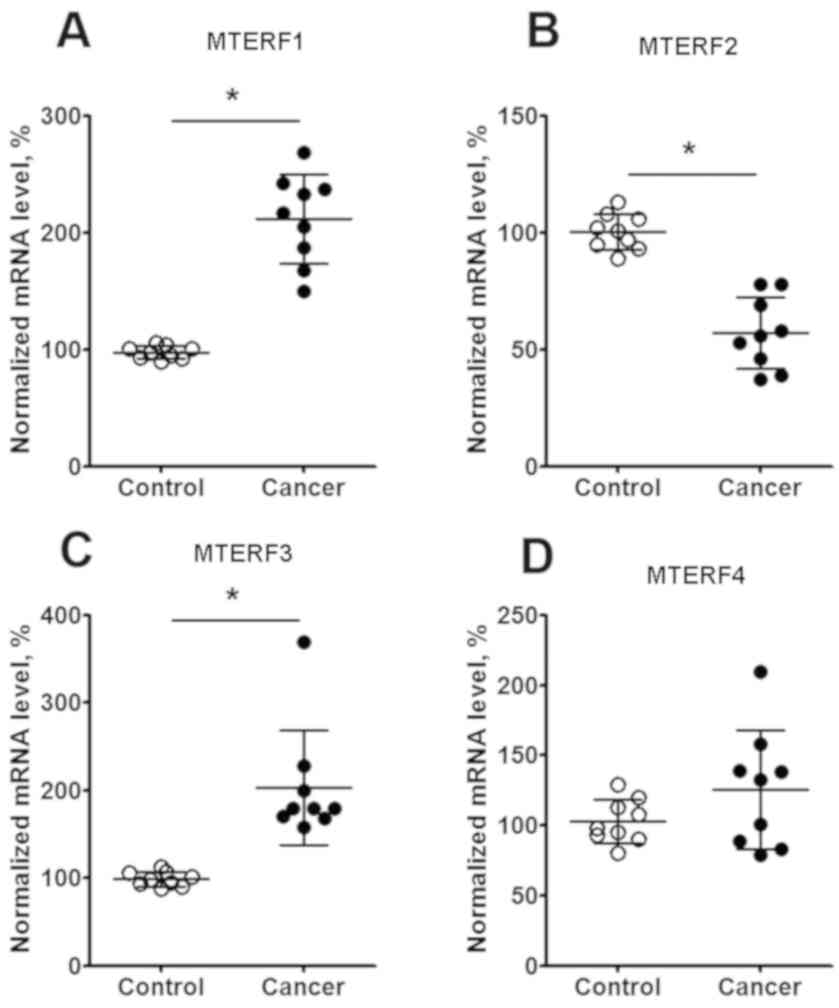
and G). Additionally, qPCR analysis in isolated lung cancer
tissue observed the increase of MTERF1 (Fig. 4A) and MTERF3 (Fig. 4C) mRNA expression (P<0.05), the
decrease of MTERF2 (Fig. 4B) mRNA
expression (P<0.05) by comparing with the non-tumor tissues.
However, the expression MTERF4 in the fresh isolated tumor tissue
has no different by comparing with that from adjacent non-tumor
tissues (Fig. 4D).
Associations between MTERF expression
levels and clinicopathological characteristics of patients with
NSCLC
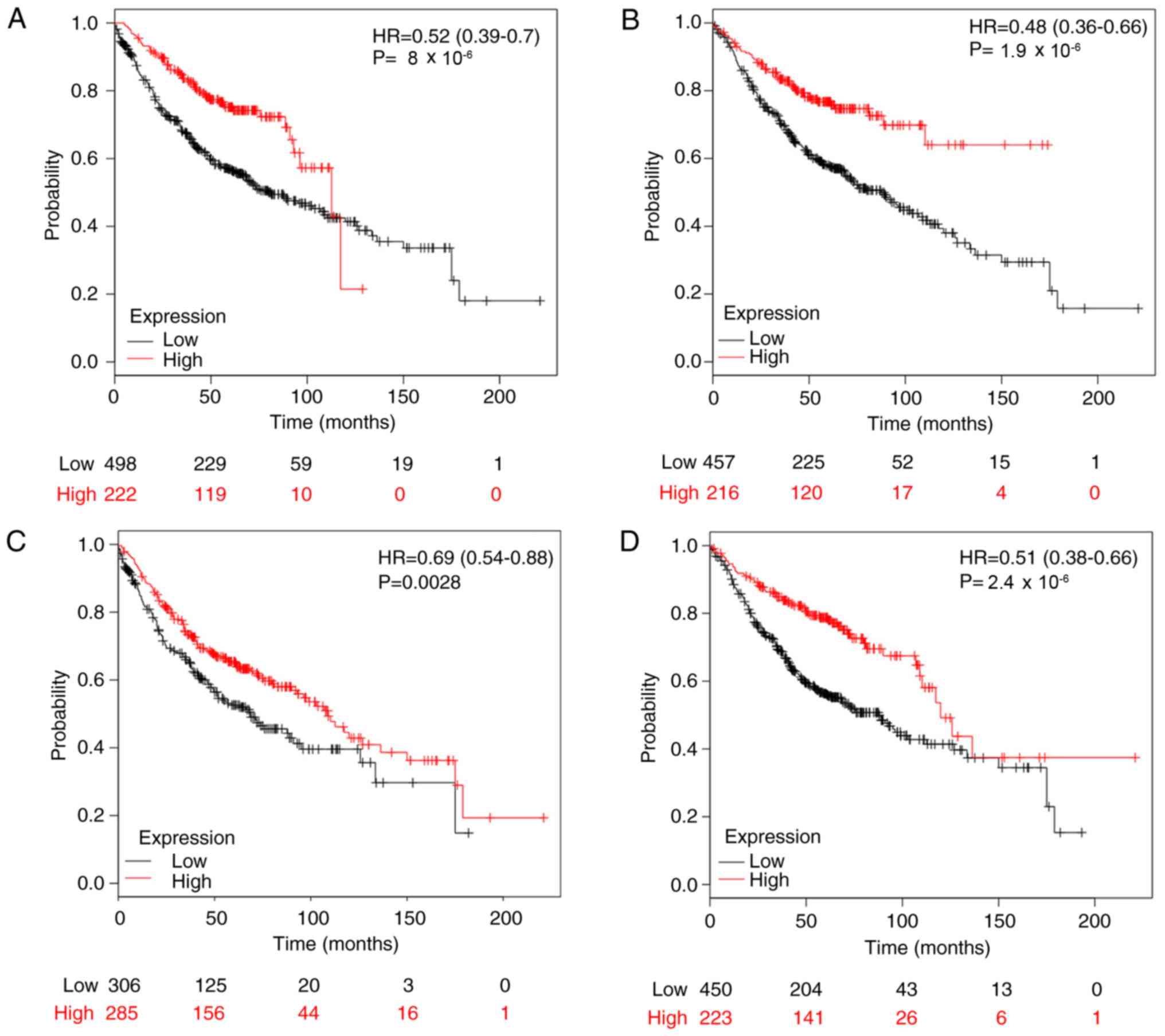
The present study used KM plotter to evaluate the
prognostic value of MTERFs in patients with NSCLC. High MTERF1 mRNA
expression was significantly associated with improved OS rate in
patients with LUAD (n=720; HR, 0.52; 95% CI, 0.39–0.70;
P=8×10−6; Fig. 5A). For
MTERF2, the survival curves were generated for patients with LUAD
(n=673; Fig. 5B). Notably, high mRNA
expression of MTERF2 was significantly associated with improved OS
rate in patients with LUAD (n=673; HR 0.48; 95% CI, 0.36–0.66;
P=1.9×10−6; Fig. 5B).
Furthermore, high MTERF3 mRNA expression was significantly
associated with improved OS rate for patients with LUAD (n=591; HR,
0.69; 95% CI, 0.54–0.88; P=0.0028; Fig.
5C). For MTERF4, a high mRNA expression level was significantly
associated with an improved OS rate for patients with LUAD (n=673;
HR, 0.51; 95% CI, 0.38–0.68; P=2.4×10−6; Fig. 5D). However, for patients with LUSC,
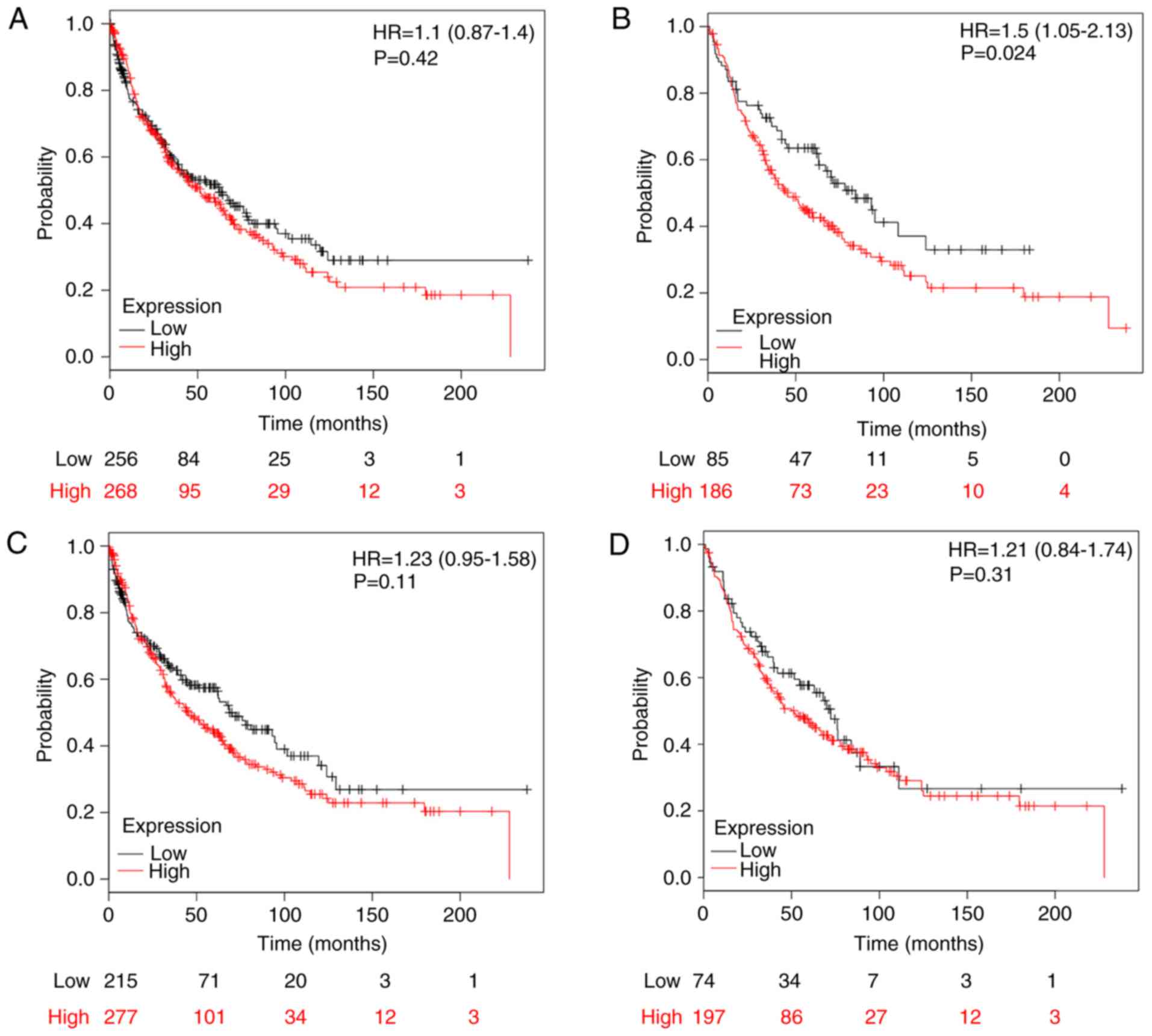
higher mRNA expression levels of MTERF1 (n=524; HR, 1.1; 95% CI,
0.87–1.40; P=0.42; Fig. 6A), MTERF3
(HR, 1.23; 95% CI, 0.95–1.58; P=0.11; Fig. 6C) and MTERF4 (HR, 1.21; 95% CI,
0.84–1.74; P=0.31; Fig. 6D) were not
associated with better OS. Higher mRNA expression of MTERF2 was
also marginally associated with improved OS in patients with LUSC
for MTERF2 (n=271; HR, 1.5; 95% CI, 1.05–2.13; P=0.024; Fig. 6B).
To further assess the association of MTERFs with
other clinicopathological factors, the present study determined the
associations of OS rate with the patients' sex (Table I), smoking status (Table II), different clinical stages
(Table III) and surgical margins
(Table IV). As presented in
Table I, high expression levels of
MTERF2 and 4, but not MTERF1 and 3, were significantly associated
with improved OS in both male and female patients with NSCLC. As
presented in Table II, the
expression levels of MTERF1 and 2 were significantly associated
with the OS in patients with NSCLC who smoked and never smoked.
However, a high expression of MTERF3 was significantly associated
with a higher OS in patients who never smoked (HR, 0.25; 95% CI,
0.09–0.69; P=0.0037) but not in patients who smoked. Additionally,
a high expression of MTERF4 was identified to be significantly
associated with OS of patients who smoked (HR, 0.33; 95% CI,
0.19–0.59; P=4.1×10−5) but not patients who never
smoked. As presented in Table III,
low MTERF1-4 mRNA expression was associated with a poorer OS rate
for patients with NSCLC in earlier stages. Particularly, for
patients in stage I, lower expression levels of MTERF1 (P=0.0006),
MTERF2 (P=1.3×10−5), MTERF3 (P=2.4×10−8) and
MTERF4 (P=2.4×10−8) were significantly associated with a
worse OS rates (Table III). For
patients with stage II tumors, the lower expression levels of
MTERF1 (P=0.00053) and MTERF3 (P=0.0063), but not the MTERF2 and
MTERF4 were significantly associated with worse OS rates (Table III). For patients in stage III,
only the lower expression of MTERF2 (P=0.032) but not MTERF1, MTEF3
and MTERF4 was significantly associated with a worse OS rates
(Table III). As presented in
Table IV, a low mRNA expression of
MTERF1-4 was associated with a worse OS in patients with negative
surgical margins.
 | Table I.Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC of different
sexes. |
Table I.
Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC of different
sexes.
| Gene | Sex | Cases, n | HR | 95% CI | P-value |
|---|
| MTERF1 | Male | 1,00 | 0.88 | 0.75–1.04 | 0.14 |
| MTERF1 | Female | 715 | 0.74 | 0.59–0.93 | 0.01 |
| MTERF2 | Male | 659 | 0.77 | 0.61–0.97 | 0.027 |
| MTERF2 | Female | 375 | 0.4 | 0.25–0.64 |
7.4×10−5 |
| MTERF3 | Male | 1,103 | 1.19 | 1.00–1.41 | 0.055 |
| MTERF3 | Female | 619 | 1.25 | 0.93–1.68 | 0.14 |
| MTERF4 | Male | 659 | 0.64 | 0.50–0.82 | 0.0032 |
| MTERF4 | Female | 375 | 0.45 | 0.32–0.63 |
2.0×10−6 |
| Table II.Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC with
different smoking statuses. |
Table II.
Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC with
different smoking statuses.
| Gene | Smoking status | Cases, n | HR | 95% CI | P-value |
|---|
| MTERF1 | Smoked | 820 | 0.66 | 0.51–0.86 | 0.0022 |
| MTERF1 | Never smoked | 205 | 0.24 | 0.13–0.45 |
1.3×10−6 |
| MTERF2 | Smoked | 300 | 0.38 | 0.22–0.63 |
1×10−4 |
| MTERF2 | Never smoked | 141 | 0.25 | 0.11–0.59 | 0.0064 |
| MTERF3 | Smoked | 702 | 1.19 | 0.95–1.50 | 0.12 |
| MTERF3 | Never smoked | 152 | 0.25 | 0.09–0.69 | 0.0037 |
| MTERF4 | Smoked | 300 | 0.33 | 0.19–0.59 |
4.1×10−5 |
| MTERF3 | Never smoked | 141 | 0.36 | 0.16–0.84 | 0.13 |
| Table III.Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC of different
clinical stages. |
Table III.
Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC of different
clinical stages.
| Gene | Clinical stage | Cases, n | HR | 95% CI | P-value |
|---|
| MTERF1 | I | 577 | 0.63 | 0.48–0.82 | 0.0006 |
| MTERF1 | II | 244 | 0.49 | 0.33–0.74 | 0.00053 |
| MTERF1 | III | 70 | 1.4 | 0.81–2.42 | 0.23 |
| MTERF2 | I | 449 | 0.44 | 0.30–0.64 |
1.3×10−5 |
| MTERF2 | II | 161 | 1.21 | 0.77–1.90 | 0.41 |
| MTERF2 | III | 44 | 0.48 | 0.24–0.95 | 0.032 |
| MTERF3 | I | 476 | 0.76 | 0.57–1.01 |
2.4×10−8 |
| MTERF3 | II | 215 | 0.58 | 0.39–0.86 | 0.0063 |
| MTERF3 | III | 53 | 0.71 | 0.38–1.32 | 0.27 |
| MTERF4 | I | 449 | 0.36 | 0.25–0.52 |
2.4×10−8 |
| MTERF4 | II | 161 | 0.62 | 0.36–1.06 | 0.078 |
| MTERF4 | III | 44 | 2.09 | 0.94–4.67 | 0.066 |
| Table IV.Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC with negative
surgical margins. |
Table IV.
Association of MTERF1-4 expression
with the overall survival rate of patients with NSCLC with negative
surgical margins.
| Gene | Cases, n | HR | 95% CI | P-value |
|---|
| MTERF1 | 726 | 0.38 | 0.28–0.51 |
3.7×10−11 |
| MTERF2 | 204 | 0.2 | 0.09–0.42 |
2.6×10−6 |
| MTERF3 | 615 | 0.63 | 0.46–0.86 | 0.036 |
| MTERF4 | 204 | 0.29 | 0.14–0.61 | 0.00052 |
Discussion
The MTERF family consists of four members, termed
MTERF1-4, which have been identified in plants but not in fungi
(8). MTERFs contain leucine
zipper-like heptads, localize to the mitochondria, and are involved
in mitochondrial gene transcription and protein synthesis (8). Among the four MTERFs, MTERF1 and 2 are
uniquely expressed in vertebrates (34). In humans, all MTERFs are expressed in
lung tissue (35). In general, a
number of studies have indicated that MTERFs bind to mtDNA promoter
regions and regulate mitochondrial gene expression (6,7,9,10).
Previous studies have suggested that the roles of MTERFs are likely
more complex than just regulating mitochondrial gene expression
(9,10). Human MTERF1 is the most well
characterized family member of the MTERFs, and is a mitochondrial
protein consisting of 342 amino acids (8). MTERF1 modulates mitochondrial gene
replication and regulates mitochondrial gene expression by
arresting mitochondrial RNA polymerase progression (36). The pathogenic MELAS mutation of
MTERF1 has been demonstrated to attenuate the ability of
transcription termination in vitro (11,37).
Additionally, MTERF2 is highly expressed in heart, liver and
skeletal muscle cells (14), and was
demonstrated to bind to the promoter region of mitochondrial genes
and to regulate the initiation of mitochondrial gene transcription
(6,7,9,10). MTERF2-knockout mice exhibit myopathy
and memory deficits, which may be due to decreased mtDNA
transcription (38). In addition,
the loss of MTERF2 in mice results in decreased oxidative
phosphorylation complexes and causes a respiratory defect (14). Similarly, MTERF3 is also involved in
mitochondrial biogenesis in cells of humans and other vertebrates
(39). Knockout of MTERF3 in mice
results in embryo mortality (40),
which indicates that MTERF3 functions as an essential factor in
mouse embryonic development. Furthermore, tissue specific
inactivation of MTERF3 in the heart significantly decreased the
life span of mice, and may affect heart and skeletal muscle
functions by regulating mitochondrial functions (15). MTERF4 has a common fold similar to
MTERF1 and MTERF3, which contains positively charged surfaces and
may be beneficial for nucleic acid interaction (16). In human cells, 3D crystal structure
analysis indicated that MTERF4 is required for mitochondrial
ribosomal biogenesis and translation via forming a stoichiometric
complex with NSUN4 (16). In a human
neuroblast cell line, knockdown of MTERF4 increased the
MPP+-induced mitochondrial dysfunction, which may be due
to increased mtDNA transcription and translation levels (17). Similar to MTERF3, MTERF4-knockdown in
mice also induces embryo mortality (6,40).
Furthermore, the loss of MTERF4 in the mouse heart also disrupts
the transfer RNA pool; however, it increases steady-state levels of
mtDNA transcripts (17). To the best
of our knowledge, until now, the roles of MTERFs in the
pathogenesis of NSCLC have not been investigated.
Mitochondrial functions are essential for eukaryotic
cells (3,4). Defects in mitochondrial gene expression
are associated with numerous diseases, including LC (1). Mitochondrial gene mutations are common
in cancer development and may regulate mitochondrial metabolism
(1). The results of the present
study provide insight into the roles of MTERFs in NSCLC
pathogenesis and suggest an association between mitochondrial gene
transcription and LC. The present study investigated the expression
and prognostic roles of MTERFs in patients with NSCLC, and the
results indicated that the expression levels of MTERF family
members were significantly altered in NSCLC tissue. Notably, high
expression levels of MTERF1, 2, 3 and 4 were significantly
associated with an improved OS for patients with LUAD, and patients
with early stage NSCLC. These results indicate that the expression
levels of MTERFs may exhibit critical prognostic values for
patients with NSCLC; therefore, MTERFs may be used as early
biomarkers for predicting the OS of patients with NSCLC. In
summary, the present study provides novel insights that may promote
further investigations regarding the pathogenesis of NSCLC, and
suggests a novel therapeutic strategy of NSCLC via targeting
mitochondrial transcription. The present study only focuses on the
bioinformatic analysis, and this conclusion may require additional
biological experiments in order to accurately identify the
potential underlying molecular mechanism.
Acknowledgements
Not applicable.
Funding
No funding was received.
Availability of data and materials
The datasets used and/or analyzed during the present
study are available from the corresponding author on reasonable
request.
Authors' contributions
SS, CW, ZH and LM conceived the study and wrote and
revised the manuscript. CY and JC analyzed the gene expression in
lung tissues from patients. SS, CW and LM reviewed, collected and
analyzed the data. XW, YN, ZH and LM designed the study and
acquired the data. All authors contributed to the writing of the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to
participate
All patients gave their full consent to participate
in the present study, and a written consent form was obtained from
each patient. The research protocol was approved by the
Institutional Review Board of Jilin Province Cancer Hospital.
Patient consent for publication
A written consent form was obtained from each
patient.
Competing interests
The authors declare that they have no competing
interests.
Glossary
Abbreviations
Abbreviations:
|
MTERF
|
mitochondrial transcription
termination factor
|
|
NSCLC
|
non-small cell lung cancer
|
|
LC
|
lung cancer
|
|
LUAD
|
lung adenocarcinoma
|
|
LUSC
|
lung squamous cell carcinoma
|
|
KM plotter
|
Kaplan-Meier plotter
|
|
HR
|
hazard ratio
|
|
OS
|
overall survival
|
|
TGCA
|
The Cancer Genome Atlas
|
|
GEO
|
Gene Expression Omnibus
|
|
mtDNA
|
mitochondrial DNA
|
|
NSUN4
|
NOP2/Sun RNA methyltransferase family
member 4
|
References
|
1
|
Baffy G: Mitochondrial uncoupling in
cancer cells: Liabilities and opportunities. Biochim Biophys Acta
Bioenerg. 1858:655–664. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Balaban RS: Regulation of oxidative
phosphorylation in the mammalian cell. Am J Physiol. 258:C377–C389.
1990. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hatefi Y: The mitochondrial electron
transport and oxidative phosphorylation system. Annu Rev Biochem.
54:1015–1069. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
McCommis KS and Finck BN: Mitochondrial
pyruvate transport: A historical perspective and future research
directions. Biochem J. 466:443–454. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Gilkerson R, Bravo L, Garcia I, Gaytan N,
Herrera A, Maldonado A and Quintanilla B: The mitochondrial
nucleoid: Integrating mitochondrial DNA into cellular homeostasis.
Cold Spring Harb Perspect Biol. 5:a0110802013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kleine T and Leister D: Emerging functions
of mammalian and plant mTERFs. Biochim Biophys Acta. 1847:786–797.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kleine T: Arabidopsis thaliana mTERF
proteins: Evolution and functional classification. Front Plant Sci.
3:2332012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Roberti M, Polosa PL, Bruni F, Manzari C,
Deceglie S, Gadaleta MN and Cantatore P: The MTERF family proteins:
Mitochondrial transcription regulators and beyond. Biochim Biophys
Acta. 1787:303–311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Guja KE and Garcia-Diaz M: Hitting the
brakes: Termination of mitochondrial transcription. Biochim Biophys
Acta. 1819:939–947. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Terzioglu M, Ruzzenente B, Harmel J,
Mourier A, Jemt E, López MD, Kukat C, Stewart JB, Wibom R, Meharg
C, et al: MTERF1 binds mtDNA to prevent transcriptional
interference at the light-strand promoter but is dispensable for
rRNA gene transcription regulation. Cell Metab. 17:618–626. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yakubovskaya E, Mejia E, Byrnes J,
Hambardjieva E and Garcia-Diaz M: Helix unwinding and base flipping
enable human MTERF1 to terminate mitochondrial transcription. Cell.
141:982–993. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chomyn A, Martinuzzi A, Yoneda M, Daga A,
Hurko O, Johns D, Lai ST, Nonaka I, Angelini C and Attardi G: MELAS
mutation in mtDNA binding site for transcription termination factor
causes defects in protein synthesis and in respiration but no
change in levels of upstream and downstream mature transcripts.
Proc Natl Acad Sci USA. 89:4221–4225. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen Y, Zhou G, Yu M, He Y, Tang W, Lai J,
He J, Liu W and Tan D: Cloning and functional analysis of human
mTERFL encoding a novel mitochondrial transcription termination
factor-like protein. Biochem Biophys Res Commun. 337:1112–1118.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wenz T, Luca C, Torraco A and Moraes CT:
MTERF2 regulates oxidative phosphorylation by modulating mtDNA
transcription. Cell Metab. 9:499–511. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park CB, Asin-Cayuela J, Camara Y, Shi Y,
Pellegrini M, Gaspari M, Wibom R, Hultenby K, Erdjument-Bromage H,
Tempst P, et al: MTERF3 is a negative regulator of mammalian mtDNA
transcription. Cell. 130:273–285. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Camara Y, Asin-Cayuela J, Park CB,
Metodiev MD, Shi Y, Ruzzenente B, Kukat C, Habermann B, Wibom R,
Hultenby K, et al: MTERF4 regulates translation by targeting the
methyltransferase NSUN4 to the mammalian mitochondrial ribosome.
Cell Metab. 13:527–539. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ye X, Han Y, Zhang L, Liu W and Zuo J:
MTERF4 regulates the mitochondrial dysfunction induced by MPP(+) in
SH-SY5Y cells. Biochem Biophys Res Commun. 464:214–220. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wallace DC: A mitochondrial paradigm of
metabolic and degenerative diseases, aging, and cancer: A dawn for
evolutionary medicine. Annu Rev Genet. 39:359–407. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Larsson NG: Somatic mitochondrial DNA
mutations in mammalian aging. Annu Rev Biochem. 79:683–706. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Keith RL and Miller YE: Lung cancer
chemoprevention: Current status and future prospects. Nat Rev Clin
Oncol. 10:334–343. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Roberts ER and Thomas KJ: The role of
mitochondria in the development and progression of lung cancer.
Comput Struct Biotechnol J. 6:e2013030192013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lennon FE and Salgia R: Mitochondrial
dynamics: Biology and therapy in lung cancer. Expert Opin Investig
Drugs. 23:675–692. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hori Y, Otomura N, Nishida A, Nishiura M,
Umeno M, Suetake I and Kikuchi K: Synthetic-molecule/protein hybrid
probe with fluorogenic switch for live-cell imaging of DNA
methylation. J Am Chem Soc. 140:1686–1690. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang H, Feng Z, Del Signore SJ, Rodal AA
and Xu B: Active probes for imaging membrane dynamics of live cells
with high spatial and temporal resolution over extended time scales
and areas. J Am Chem Soc. 140:3505–3509. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Liu LJ, Wang W, Huang SY, Hong Y, Li G,
Lin S, Tian J, Cai Z, Wang HD, Ma DL and Leung CH: Inhibition of
the Ras/Raf interaction and repression of renal cancer xenografts
in vivo by an enantiomeric iridium(iii) metal-based compound. Chem
Sci. 8:4756–4763. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Vellaisamy K, Li G, Ko CN, Zhong HJ,
Fatima S, Kwan HY, Wong CY, Kwong WJ, Tan W, Leung CH and Ma DL:
Cell imaging of dopamine receptor using agonist labeling
iridium(iii) complex. Chem Sci. 9:1119–1125. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huang Z, Yang C, Sun S, Nan Y, Lang Z,
Wang X, Zhao J and Liu Y: Heat shock protein 27, a novel regulator
of transforming growth factor β induced resistance to cisplatin in
a549 cell. Pharmacology. 100:283–291. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cerami E, Gao J, Dogrusoz U, Gross BE,
Sumer SO, Aksoy BA, Jacobsen A, Byrne CJ, Heuer ML, Larsson E, et
al: The cBio cancer genomics portal: An open platform for exploring
multidimensional cancer genomics data. Cancer Discov. 2:401–404.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Landi MT, Dracheva T, Rotunno M, Figueroa
JD, Liu H, Dasgupta A, Mann FE, Fukuoka J, Hames M, Bergen AW, et
al: Gene expression signature of cigarette smoking and its role in
lung adenocarcinoma development and survival. PLoS One.
3:e16512008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Győrffy B, Surowiak P, Budczies J and
Lánczky A: Online survival analysis software to assess the
prognostic value of biomarkers using transcriptomic data in
non-small-cell lung cancer. PLoS One. 8:e822412013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Huang LS, Mathew B, Li H, Zhao Y, Ma SF,
Noth I, Reddy SP, Harijith A, Usatyuk PV, Berdyshev EV, et al: The
mitochondrial cardiolipin remodeling enzyme lysocardiolipin
acyltransferase is a novel target in pulmonary fibrosis. Am J
Respir Crit Care Med. 189:1402–1415. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fernandez-Silva P, Martinez-Azorin F,
Micol V and Attardi G: The human mitochondrial transcription
termination factor (mTERF) is a multizipper protein but binds to
DNA as a monomer, with evidence pointing to intramolecular leucine
zipper interactions. EMBO J. 16:1066–1079. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wobbe L and Nixon PJ: The mTERF protein
MOC1 terminates mitochondrial DNA transcription in the unicellular
green alga Chlamydomonas reinhardtii. Nucleic Acids Res.
41:6553–6567. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Xu Q, Zhang F, He H, Xu S, Li K, Liu S, Li
Y and Wu Q: Expression profile of mouse Mterfd2, a novel component
of the mitochondrial transcription termination factor (MTERF)
family. Genes Genet Syst. 86:269–275. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Arnold JJ, Smidansky ED, Moustafa IM and
Cameron CE: Human mitochondrial RNA polymerase: Structure-function,
mechanism and inhibition. Biochim Biophys Acta. 1819:948–960. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Hess JF, Parisi MA, Bennett JL and Clayton
DA: Impairment of mitochondrial transcription termination by a
point mutation associated with the MELAS subgroup of mitochondrial
encephalomyopathies. Nature. 351:236–239. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wredenberg A, Lagouge M, Bratic A,
Metodiev MD, Spåhr H, Mourier A, Freyer C, Ruzzenente B, Tain L,
Grönke S, et al: MTERF3 regulates mitochondrial ribosome biogenesis
in invertebrates and mammals. PLoS Genet. 9:e10031782013.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Roberti M, Bruni F, Loguercio Polosa P,
Manzari C, Gadaleta MN and Cantatore P: MTERF3, the most conserved
member of the mTERF-family, is a modular factor involved in
mitochondrial protein synthesis. Biochim Biophys Acta.
1757:1199–1206. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Peralta S, Wang X and Moraes CT:
Mitochondrial transcription: Lessons from mouse models. Biochim
Biophys Acta. 1819:961–969. 2012. View Article : Google Scholar : PubMed/NCBI
|