Introduction
Multiple myeloma (MM) is a type of plasma
cell-derived malignancy, which leads to the formation of multiple
bone lesions and to disruption in the production of normal blood
cells (1). MM accounts for ~10% of
all hematological malignancies and is the second most common type
of hematological malignancy after non-Hodgkin’s lymphoma (2). Although a number of therapeutic
strategies exist, such as the use of steroids, chemotherapy,
radiotherapy and stem cell transplants, MM remains an incurable
disease (3). Patients with MM
exhibit elevated levels of circulating proteasome. Therefore, it
has been suggested that circulating proteasome levels may serve as
an independent prognostic factor for the survival rates of patients
with MM, and that proteasome therapy may be an effective treatment
approach (4). The first proteasome
inhibitor to be approved for clinical use by the US Food and Drug
Administration (FDA) was bortezomib. Bortezomib, a reversible
proteasome inhibitor, is approved by the FDA for treating
refractory, advanced or rapidly relapsed cases of MM (5). Curcumin is a natural product with
proteasome inhibitory effects that has been studied in a number of
cancers, alone and in combination with other traditional
chemotherapy and radiotherapy agents (6–8).
Curcumin is a primary active ingredient derived from
the spice, turmeric. Curcumin suppresses tumor growth and inhibits
cellular proliferation, invasion, angiogenesis, metastasis and
osteoclastogenesis, which are processes that involve multiple
cellular targets, such as nuclear factor (NF)-κB and
cyclooxygenase-2 (9,10). Curcumin is therefore considered to
be a multi-targeted drug that suppresses NF-κB activation
and reduces MM cell growth and apoptosis. Curcumin inhibits the
proliferation, invasion, metastasis and angiogenesis of a number of
cancers through interaction with a variety of cell signaling
proteins (11), the majority of
which are proteasome target proteins, such as the tumor suppressor
protein, p53, and the pro-apoptotic protein, B-cell lymphoma 2
(Bcl-2) associated X protein (Bax) (12,13).
The tumor suppressor protein, p53 (13), and the pro-apoptotic protein, Bax
(12), are proteasome target
proteins, which are involved in the processes of cancer survival
and carcinogenesis. Despite mutations in the p53 gene
occurring in 50% of all cancers, ~90% of MM cells retain a
functional wild-type p53 (14–17).
The low incidence of mutations and deletions in the p53 gene
make MM an ideal candidate for p53-targeted therapies. Even in
tumors that retain wild-type p53, p53 function is
ultimately inhibited by the action of mouse double minute 2 homolog
(MDM2) (17–19). The life cycle of the p53 protein is
short; during periods of cellular stress, the p53 protein is
regulated by a negative feedback mechanism. Under non-stressful
conditions, p53 is regulated by the negative regulator,
MDM2. Therefore, p53 and MDM2 form a feedback
loop with each other and are maintained at low levels (20).
In the present study, the proliferation rate of the
MM RPMI 8226 cell line was analyzed following treatment with
curcumin. In addition, changes in the expression of the p53,
Bax and MDM2 gene fragments, and in the p53 protein
were examined. Furthermore, the underlying mechanism by which
curcumin promotes RPMI 8226 cell apoptosis was discussed for the
application of curcumin in patients with MM.
Materials and methods
Cell growth curve
In total, RPMI 8226 cells (105/ml; School
of Life Sciences, Zhengzhou University, Zhengzhou, China) were
seeded into six-well plates and cultured in triplicate in RPMI-1640
medium (Invitrogen Life Technologies, Carlsbad, CA, USA) containing
10% fetal bovine serum (Invitrogen Life Technologies), with or
without curcumin. The final concentrations of curcumin were 0, 1,
2.5, 5, 7.5, 10, 15, 20 and 40 μmol/l. The final volume of medium
in each well following the addition of curcumin was 1 ml. The
number of cells in each well was counted every 24 h, and the cells
were cultured as usual with the same medium until 96 h.
MTT
First, 105/ml RPMI-8226 cells were seeded
into 96-well plates and cultured in six repeated wells. The
experimental groups contained the cells and RPMI-1640 medium with
10% fetal bovine serum and curcumin. The positive control groups
contained the cells and RPMI-1640 medium with 10% fetal bovine
serum. The negative control groups contained RPMI-1640 medium alone
with 10% fetal bovine serum. The final concentrations of curcumin
were 1, 2.5, 5, 7.5, 10, 15, 20 and 40 μmol/l. The final volume of
medium in each well following the addition of curcumin was 200 μl.
Next, the surrounding wells were covered with 200 μl
phosphate-buffered saline (PBS). The plates were then incubated at
37°C with 5% CO2 for 24, 48 and 72 h. Following this, 20
μl MTT solution (5 mg/ml in PBS, Sigma-Aldrich, Santa Clara, CA,
USA) was added to each well of the experimental, and positive and
negative control groups. Subsequent to a 4-h incubation at 37°C and
subsequent centrifugation at 1,000 × g for 5 min at 37°C, 200 μl
solution from every well was extracted, and 150 μl
dimethylsulfoxide was added to the wells. After 15 min, the optical
density (OD) at 490 nm was measured using an iMark microplate
absorbance reader (Bio-Rad 550; Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The cell proliferation inhibition ratio was
calculated using the following formula: Cell proliferation
inhibition ratio = 1- (A490 of the experimental groups / A490 of
the control groups) × 100. The A490 of the experimental groups = OD
of the experimental groups - OD of the negative control groups. The
A490 of the positive control groups = OD of the positive control
groups - OD of the negative control groups.
The average half maximal inhibitory concentration of
curcumin from six experiments was obtained by plotting the
percentage of inhibition against the concentration of curcumin.
Polymerase chain reaction (PCR)
In total, 106/ml RPMI-8226 cells in the
logarithmic phase were seeded into 25-cm2 culture
bottles for the positive control and experimental groups, including
the 10- and 15-μmol/l curcumin groups. The final volume of solution
in each culture bottle was 5 ml. The positive control groups
contained the cells and RPMI-1640 medium with 10% fetal bovine
serum. The experimental groups contained the cells, curcumin and
RPMI-1640 medium with 10% fetal bovine serum. The total RNA was
isolated after 48 h from the cells in the culture bottle using
TRIzol reagent (Invitrogen Life Technologies). Next, the RNA was
reverse transcribed into cDNA and quantitative PCR (qPCR) was
performed using the two-step method. Briefly, 25 μl reaction volume
consisting of 12.5 μl of 2X PCR Buffer for KOD FX [ a PCR
amplification enzyme (Qiagen, Venlo, Netherlands)], 5 μl 2 mM
dNTPs, 2 μl of each primer, 0.1 μl KOD, 2.4 μl water and 1 μl DNA.
The standard conditions for PCR were as follows: 95°C for 2 min,
followed by 40 cycles at 95°C for 30 sec, 62°C for 1 min, and a
final extension at 72°C for 5 min. All reactions were performed in
a PerkinElmer 2400 thermocycler (Perkin Elmer Applied Biosystems,
Foster City, CA, USA). The 2−ΔΔCt method was used to
indicate the association between the expression of the target gene
in the experimental group and the expression of the target gene in
the positive control group. The sequences of the primers are shown
in Table I.
 | Table ISequence of primers. |
Table I
Sequence of primers.
| Target gene | Sequence of
primers |
|---|
| p53 |
5′-CCACCATCCACTACAACTACAT-3′
5′-AAACACGCACCTCAAAGC-3′ |
| Bax |
5′-TTTTGCTTCAGGGTTTCATC-3′
5′-GACACTCGCTCAGCTTCTTG-3′ |
| MDM2 |
5′-TACCTACTGATGGTGCTG-3′
5′-TGATTCCTGCTGATTGAC-3′ |
| GAPDH |
5′-GGATTTGGTCGTATTGGG-3′
5′-GGAAGATGGTGATGGGATT-3′ |
Western blot analysis
For the western blot analysis, the cells were
harvested and lysed, and the proteins were separated using a 12.5%
SDS-PAGE gel. The proteins were then transferred to a Hybond-C
membrane (Invitrogen Life Technologies). Next, the membrane was
blocked with Blotto A (5% blocking grade dry milk in Tris-buffered
saline and Tween 20; Invitrogen Life Technologies) and probed using
a monoclonal mouse anti-human p53 primary antibody (dilution,
1:300; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) in Blotto
A. The cells were incubated with the primary antibody at 4°C
overnight, and then with monoclonal goat anti-mouse IgG-horseradish
peroxidase-tagged secondary antibody (dilution, 1:1,000; Santa Cruz
Biotechnology, Inc.) at room temperature for 1 h. Detection of
chemiluminescence was conducted using an enhanced chemiluminescence
western detection reagent (GE Healthcare Bio-Sciences, Pittsburgh,
PA, USA) and developed on a BioMax XAR film (Kodak, Rochester, NY,
USA).
ELISA
In total, 0.5×106/ml RPMI-8226 cells in
the logarithmic phase were seeded into 6-well plates and cultured
in four repeated wells. Control groups, and experimental groups,
including 10 and 15 μmol/l curcumin groups, were used. The final
volume of solution in each well was 1 ml. The control groups
contained the cells and RPMI-1640 medium with 10% fetal bovine
serum. The experimental groups contained the cells, curcumin and
RPMI-1640 medium with 10% fetal bovine serum. The total protein was
isolated after 48 h from the cells in each well of the plates using
100 μl lysis buffer with 1 mM EDTA, according to the manufacturer’s
instructions. The ELISAs were conducted using the p53 kit (RAB0500,
Sigma-Aldrich). The standard and sample groups were set, and the
indicated reagents were added for the indicated time period
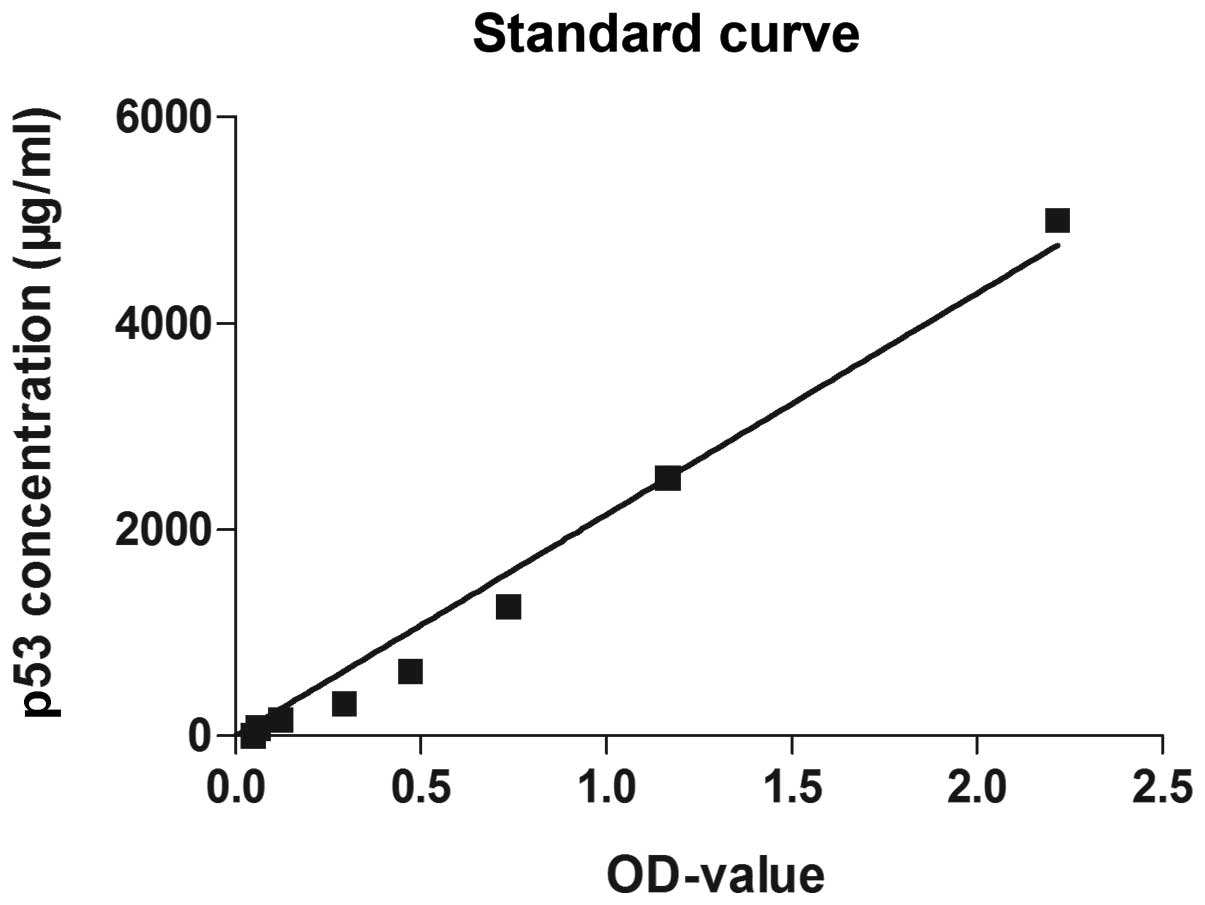
according to the manufacturer’s instructions. The OD of each well
was measured at 490 nm using a microplate reader. A standard curve
was constructed according to the OD values of the standard groups
and a formula was generated based upon this standard curve. The p53
protein concentration of each sample was calculated according to
the formula: p53 concentration = 2318.3ODvalue −
241.19.
Statistical analysis
SPSS software version 17.0 (SPSS Inc., Chicago, IL,
USA) was used to perform the statistical analysis. Data from the
control and experiment groups were analyzed by an independent
sample t-test. P<0.05 was considered to indicate a statistically
significant difference.
Results
Growth curve
The results revealed that the cells were in the
logarithmic phase between 24 and 72 h. During this time period, the
number of cells in the logarithmic phase decreased with increasing
concentrations of curcumin. During the same period, the growth of
cells treated with higher curcumin concentrations was slower
(Fig. 1).
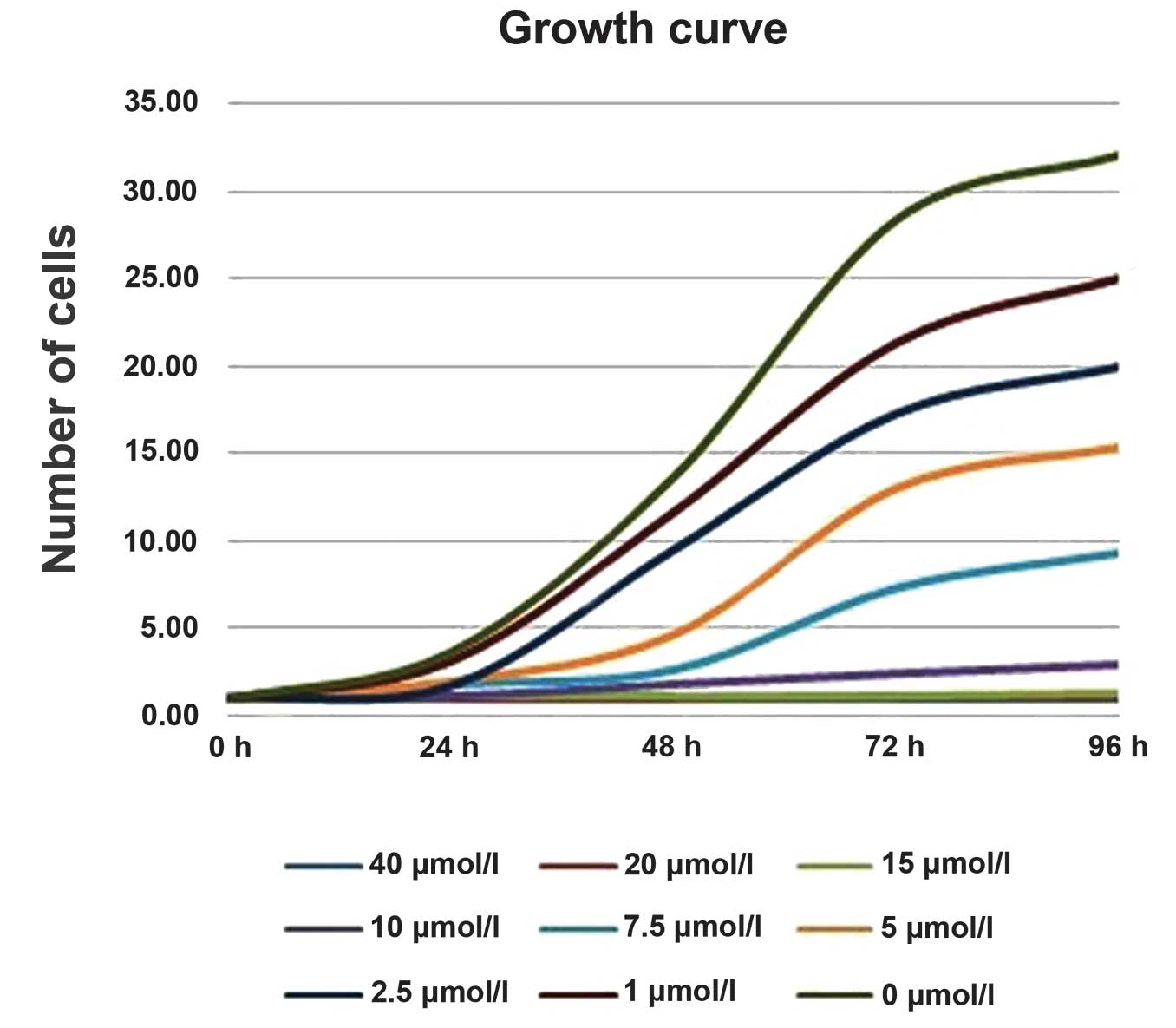 | Figure 1Growth curve representing the growth
of cells treated with different concentrations of curcumin. The
initial number of cells in each well was the same
(105/well). After 96 h, the number of cells in the wells
treated with 0, 1, 2.5, 5, 7.5 and 10 μmol/l curcumin were
>3×106, ~2.5×106, ~2×106,
~1.5×106, <1×106 and <5×105,
respectively. The cells treated with 15, 20 and 40 μmol/l curcumin,
however, barely grew. |
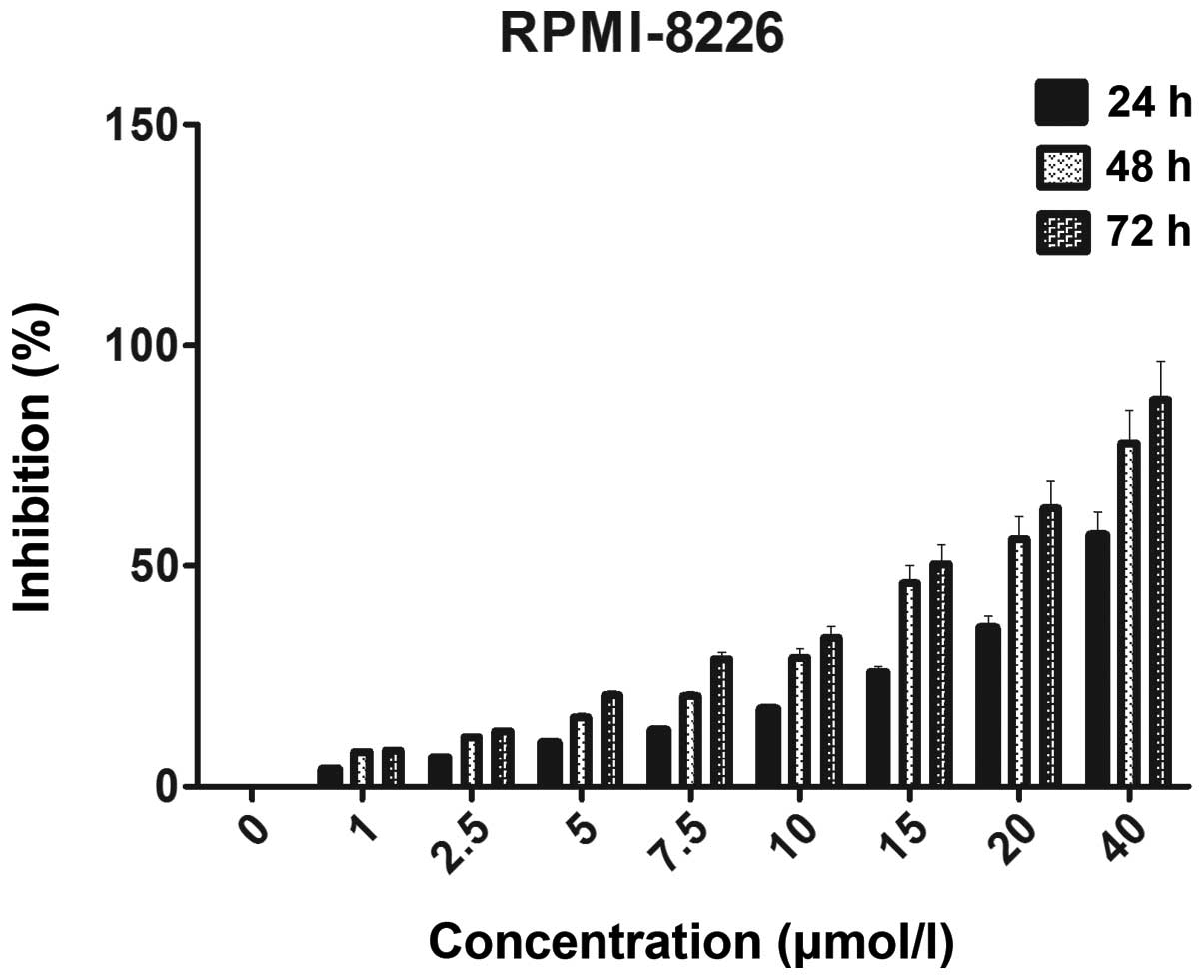
MTT
In order to investigate the effect of curcumin on
the proliferation inhibition of the MM RPMI 8226 cell line, the OD
was measured at 490 nm following 24, 48 and 72 h of treatment with
different concentrations of curcumin. The proliferation inhibition
ratio following 24, 48 and 72 h of treatment with 10 μmol/l
curcumin was 17.6, 29.2 and 33.8%, respectively. The difference in
the OD value was statistically significant between the experimental
and control groups (P<0.05). The proliferation inhibition ratio
following 24, 48 and 72 h of treatment with 15 μmol/l curcumin was
25.8, 46.1 and 50.4%, respectively. The difference in the OD value
was statistically significant between the experimental and control
groups (P<0.05). The results revealed that a higher
concentration of curcumin was more potent than a lower
concentration of curcumin at the same time-point in the growth
suppression of the RPMI 8226 cells, and that a longer duration of
treatment was more potent than a shorter duration of treatment with
the same concentration of curcumin in the growth suppression of the
RPMI 8226 cells (Fig. 2).
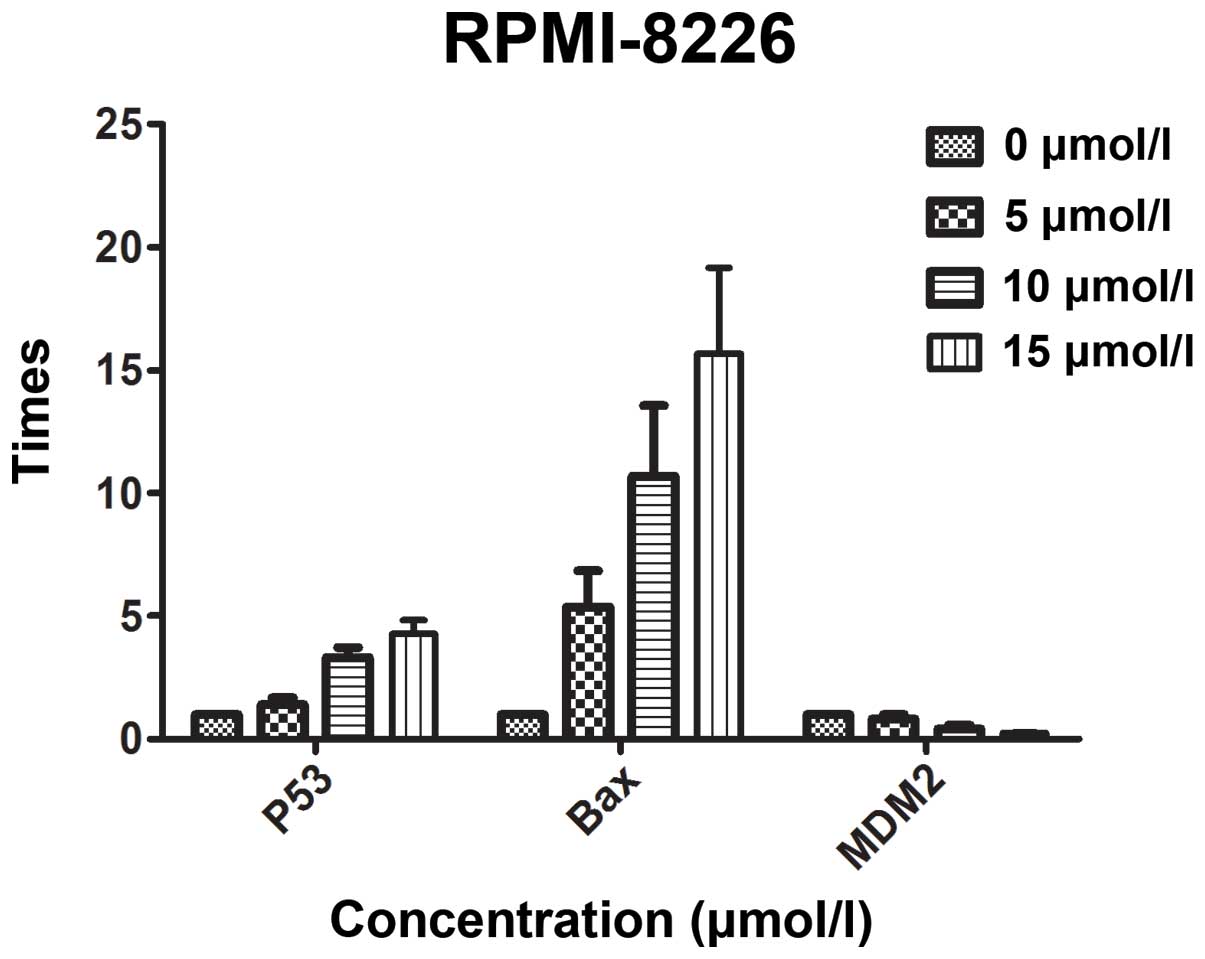
qPCR
The qPCR results revealed that curcumin inhibited
the growth of MM cells in a dose-dependent manner. Following a 48-h
treatment with 10 and 15 μmol/l curcumin, the proliferation
inhibition ratio was 29.2 and 46.1%, respectively. It has been
reported that curcumin regulates the expression of the
apoptosis-related proteins, Bax, Bcl-2 and p53, and that it
regulates the apoptosis of tumor cells via the p53 pathway
(21). The low incidence of
mutations and deletions in the p53 gene make MM an ideal
candidate for p53-targeted therapies (14–17).
In order to investigate whether curcumin inhibits the growth of MM
cells through the p53 pathway, qPCR was used to analyze the
expression of p53, Bax and MDM2 gene fragments
following treatment with different concentrations of curcumin.
Subsequent to a 48-h treatment with 10 μmol/l curcumin, the
expression of the p53, Bax and MDM2 genes was
1.3905, 10.3581 and 0.4046 times higher than the expression of
p53, Bax and MDM2 in the control group,
respectively. Furthermore, following a 48-h treatment with 15
μmol/l, the differences were 2.0871, 12.6826 and 0.2505 times
higher. The differences were statistically significant (P<0.05).
The results are shown in Fig.
3.
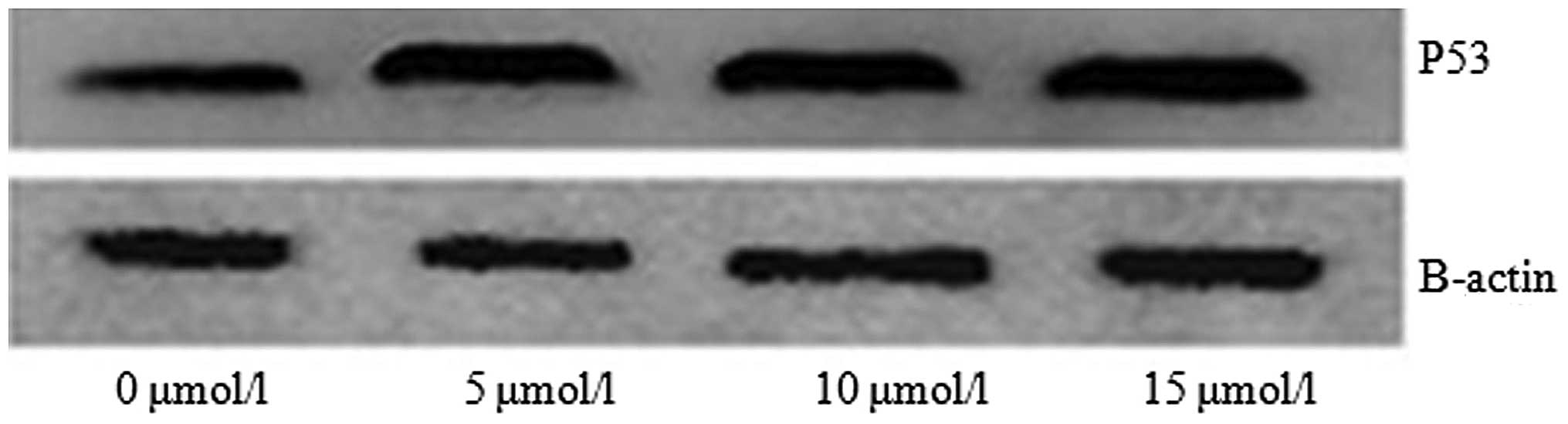
Western blot analysis
The p53 signaling pathway is an important
pathway involved in tumor cell apoptosis. When curcumin was used to
treat the RPMI 8226 cells, the expression of the p53 gene
was increased compared with the control group. In order to detect
the expression of the p53 protein following treatment with
curcumin, a western blot assay was performed. It was revealed that
the expression of the p53 protein in the cells treated with 5, 10
and 15 μmol/l curcumin for 48 h was increased compared with the
control group (Fig. 4).
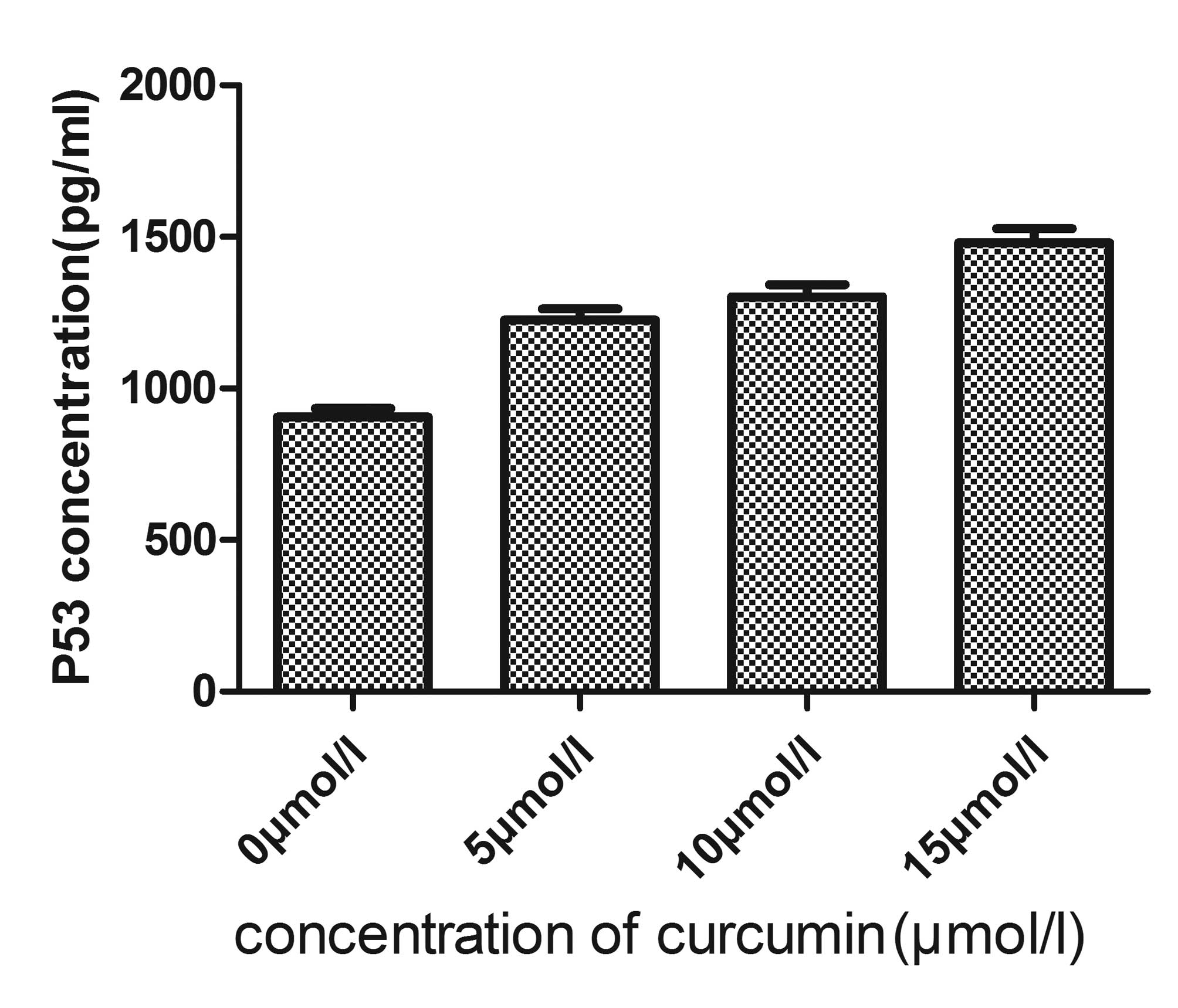
ELISA
As the expression of the p53 gene fragment
was increased in the RPMI 8226 cells treated with curcumin, the
present study next sought to determine the effect of curcumin on
p53 protein expression in the MM RPMI 8226 cell line. The RPMI 8226
cells were treated with 5, 10 and 15 μmol/l curcumin for 48 h. The
total proteins were then isolated and the expression level of the
p53 protein was determined. The results revealed that the
expression of the p53 protein was upregulated following treatment
with curcumin in a dose-dependent manner (Table II; Figs. 5 and 6). These results indicated that curcumin
promotes p53 protein expression, and may induce apoptosis through
the p53 pathway.
| Table IIp53 protein content of each sample
(mean ± standard deviation; n=6). |
Table II
p53 protein content of each sample
(mean ± standard deviation; n=6).
| Curcumin
concentration, μmol/l | p53 protein,
pg/ml |
|---|
| 0 | 906.035±28.324 |
| 5 |
1226.024±36.536 |
| 10 |
1302.629±40.007 |
| 15 |
1481.220±45.510 |
Discussion
MM is a B-cell malignancy whereby plasma cells grow
abnormally in the bone marrow and secret monoclonal immunoglobulin
or an M protein fragment, which ultimately leads to relative organ
or tissue injury. Although MM is sensitive to a variety of
cytotoxic drugs in the initial and relapsed treatment periods, the
relief is only temporary. Therefore, MM remains an incurable
disease. A number of drugs, including bortezomib, a proteasome
inhibitor, and thalidomide, an inhibitor of tumor necrosis factor
production (22,23), have been tested during the search
for an effective treatment for MM. Bortezomib, the first reversible
proteasome inhibitor approved by the US FDA for treating
refractory, advanced or rapidly relapsed MM (5), has been researched extensively.
However, bortezomib is expensive, and the majority of patients with
MM cannot afford to be treated with it. Curcumin is an inexpensive,
natural plant ingredient with protease inhibitor effects, which has
been studied, alone or in combination with traditional chemotherapy
and radiotherapy agents against a number of cancers (6–8)
Therefore, the present study analyzed the inhibitory effects of
curcumin on the MM RPMI 8226 cell line, and examined the underlying
mechanism that promotes the apoptosis of RPMI 8226 cells.
The results of the present study revealed that
following treatment with curcumin, the growth of the MM RPMI 8226
cell line was inhibited in a concentration- and time-dependent
manner, which was in agreement with the results of a study by
Bharti et al (24). In the
RPMI 8226 cells treated with curcumin, the expression of p53
protein was upregulated, which suggested that curcumin may promote
the apoptosis of MM cells by upregulating p53 protein expression. A
number of previous in vitro and in vivo studies have
indicated that curcumin exhibits a variety of pharmacological
effects, including antitumor, anti-inflammatory and antioxidant
activities, and that the side-effects of treatment are minor
(25). Other studies have revealed
that curcumin can inhibit the proliferation of MM cells by
downregulating the expression of interleukin-6 and NF-κB.
Curcumin also prevents osteoclast-inducing osteogenesis and
improves the resistance of MM cells to conventional chemotherapy
drugs (26). Bharti et al
(24) confirmed that curcumin can
promote and induce the apoptosis of MM cells (24), and suppress osteoclastogenesis by
inhibiting the receptor activator of NF-κB ligand signal
(27). Therefore, curcumin exerts
its antitumor effect through the mechanisms of inhibiting the
proliferation of tumor cells, regulating the expression of
oncogenes and anticancer genes, and inducing cell cycle arrest and
apoptosis.
Curcumin also inhibits the proliferation, invasion,
metastasis and angiogenesis of a number of cancers by interacting
with a variety of cell signaling proteins (11), the majority of which are proteasome
target proteins, including the tumor suppressor protein, p53, and
the pro-apoptotic protein, Bax (12,13).
The pro-apoptotic protein, Bax (12), and the tumor suppressor protein, p53
(13), are two types of proteasome
target proteins involved in the processes of cancer survival and
carcinogenesis. It has been reported that curcumin effects the
expression of the apoptosis-related proteins, Bax, Bcl-2 and p53,
and that it has the ability to regulate the apoptosis of tumor
cells via the p53 pathway (21). Furthermore, previous studies have
demonstrated that curcumin can induce apoptosis through
p53-dependent and -independent pathways during the treatment
of endometriosis (28), and that it
can upregulate the expression of p53 protein and Bax mRNA in
thioacetamide-induced liver fibrosis (29). Therefore, it was hypothesized that
curcumin may inhibit the proliferation and induce the apoptosis of
MM cells through a p53-mediated pathway, which may be a
novel therapeutic target for clinical use.
Despite mutations in the p53 gene occurring
in 50% of all cancers, ~90% of MM cells retain functional wild-type
p53 (14–16). Even in tumors that retain wild-type
p53, p53 function is ultimately inhibited by the
action of MDM2 (17–19). The life cycle of the p53 protein is
short; during periods of cellular stress, the p53 protein is
regulated by a negative feedback mechanism. Under non-stressful
conditions, p53 is regulated by the negative regulator, MDM2.
Therefore, p53 and MDM2 form a feedback loop with each other and
are maintained at a low level (20).
The present study demonstrated that with the
intervention of curcumin, the growth of the MM RPMI 8226 cells was
inhibited in a concentration- and time-dependent manner. Using qPCR
to detect the mRNA expression of p53, Bax and
MDM2, it was revealed that in the RPMI 8226 cells treated
with curcumin, the expression of the p53 and Bax
genes was upregulated, while the expression of the MDM2 gene
was downregulated. Curcumin has the ability to upregulate
p53 and Bax, and downregulate MDM2. Since the
action of p53 is inhibited by MDM2, the
downregulation in MDM2 may reduce the inhibition of p53. The
results of the western blot analysis and the ELISA indicated that
when 5, 10 and 15 μmol/l curcumin was administered to the MM RPMI
8226 cells for 48 h, the expression of the p53 protein was
upregulated. The level of p53 protein expressed in the 15, 10 and 5
μmol/l curcumin experimental groups was higher than the amount of
p53 protein expressed in the control group. Although only one MM
cell line was investigated in the present study, curcumin
significantly upregulated p53 and Bax and
downregulated the negative inhibitor for p53, MDM2.
Therefore, it can be concluded that curcumin may inhibit the
proliferation and induce the apoptosis of MM cells through a
p53-mediated pathway different from the one identified in
the study by Bharti et al (24). Further studies and the use of other
MM cell lines are required in order to investigate this hypothesis
and provide evidence for the clinical application of curcumin,
which may provide a novel therapeutic target and an effective
treatment strategy.
Acknowledgements
The authors are grateful for the collaboration of
the participating college and its staff. The authors would like to
thank Dr Weiquan Lu from the Department of Cancer Prevention, Henan
Cancer Hospital, Zhengzhou, China, for help with the statistical
analysis.
References
|
1
|
Raab MS, Podar K, Breitkreutz I, et al:
Multiple myeloma. Lancet. 374:324–339. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Garcia-Sanz R, Mateos MV and San Miguel J:
Multiple myeloma. Med Clin (Barc). 129:104–115. 2007.(In Spanish).
View Article : Google Scholar
|
|
3
|
Krstevska SB, Sotirova T, Balkanov T and
Genadieva-Stavric S: Treatment approach of nontransplant patients
with multiple myeloma. Mater Sociomed. 26:348–351. 2014. View Article : Google Scholar
|
|
4
|
Jakob C, Egerer K, Liebisch P, et al:
Circulating proteasome levels are an independent prognostic factor
for survival in multiple myeloma. Blood. 109:2100–2105. 2007.
View Article : Google Scholar
|
|
5
|
Adams J and Kauffman M: Development of the
proteasome inhibitor Velcade (Bortezomib). Cancer Invest.
22:304–311. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Milacic V, Banerjee S, Landis-Piwowar KR,
Sarkar FH, Majumdar AP and Dou QP: Curcumin inhibits the proteasome
activity in human colon cancer cells in vitro and in vivo. Cancer
Res. 68:7283–7292. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dhillon N, Aggarwal BB, Newman RA, et al:
Phase II trial of curcumin in patients with advanced pancreatic
cancer. Clin Cancer Res. 14:4491–4499. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Garcea G, Jones DJ, Singh R, et al:
Detection of curcumin and its metabolites in hepatic tissue and
portal blood of patients following oral administration. Br J
Cancer. 90:1011–1015. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
von Metzler I, Krebbel H, Kuckelkorn U, et
al: Curcumin diminishes human osteoclastogenesis by inhibition of
the signalosome-associated I kappaB kinase. J Cancer Res Clin
Oncol. 135:173–179. 2009. View Article : Google Scholar
|
|
10
|
Kunnumakkara AB, Anand P and Aggarwal BB:
Curcumin inhibits proliferation, invasion, angiogenesis and
metastasis of different cancers through interaction with multiple
cell signaling proteins. Cancer Lett. 269:199–225. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kudo C, Yamakoshi H, Sato A, et al: Novel
curcumin analogs, GO-Y030 and GO-Y078, are multi-targeted agents
with enhanced abilities for multiple myeloma. Anticancer Res.
31:3719–3726. 2011.PubMed/NCBI
|
|
12
|
Li B and Dou QP: Bax degradation by the
ubiquitin/proteasome-dependent pathway: involvement in tumor
survival and progression. Proc Natl Acad Sci USA. 97:3850–3855.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scheffner M, Werness BA, Huibregtse JM,
Levine AJ and Howley PM: The E6 oncoprotein encoded by human
papillomavirus types 16 and 18 promotes the degradation of p53.
Cell. 63:1129–1136. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chng WJ, Price-Troska T, Gonzalez-Paz N,
et al: Clinical significance of TP53 mutation in myeloma. Leukemia.
21:582–584. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chang H, Qi C, Yi QL, Reece D and Stewart
AK: p53 gene deletion detected by fluorescence in situ
hybridization is an adverse prognostic factor for patients with
multiple myeloma following autologous stem cell transplantation.
Blood. 105:358–360. 2005. View Article : Google Scholar
|
|
16
|
Avet-Loiseau H, Li JY, Godon C, et al: P53
deletion is not a frequent event in multiple myeloma. Brit J
Haematol. 106:717–719. 1999. View Article : Google Scholar
|
|
17
|
Saha MN, Micallef J, Qiu L and Chang H:
Pharmacological activation of the p53 pathway in haematological
malignancies. J Clin Pathol. 63:204–209. 2010. View Article : Google Scholar
|
|
18
|
Saha MN, Jiang H, Jayakar J, Reece D,
Branch DR and Chang H: MDM2 antagonist nutlin plus proteasome
inhibitor velcade combination displays a synergistic anti-myeloma
activity. Cancer Biol Ther. 9:936–944. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vassilev LT: MDM2 inhibitors for cancer
therapy. Trends Mol Med. 13:23–31. 2007. View Article : Google Scholar
|
|
20
|
Shangary S and Wang S: Targeting the
MDM2-p53 interaction for cancer therapy. Clin Cancer Res.
14:5318–5324. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Song G, Mao YB, Cai QF, Yao LM, Ouyang GL
and Bao SD: Curcumin induces human HT-29 colon adenocarcinoma cell
apoptosis by activating p53 and regulating apoptosis-related
protein expression. Braz J Med Biol Res. 38:1791–1798. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Barlogie B, Zangari M, Spencer T, et al:
Thalidomide in the management of multiple myeloma. Semin Hematol.
38:250–259. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Adams J: Proteasome inhibition in cancer:
development of PS-341. Semin Oncol. 28:613–619. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Bharti AC, Donato N, Singh S and Aggarwal
BB: Curcumin (diferuloylmethane) down-regulates the constitutive
activation of nuclear factor-kappa B and IkappaBalpha kinase in
human multiple myeloma cells, leading to suppression of
proliferation and induction of apoptosis. Blood. 101:1053–1062.
2003. View Article : Google Scholar
|
|
25
|
Child JA, Morgan GJ, Davies FE, et al;
Medical Research Council Adult Leukaemia Working Party. High-dose
chemotherapy with hematopoietic stem-cell rescue for multiple
myeloma. N Engl J Med. 348:1875–1883. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Sung B, Kunnumakkara AB, Sethi G, Anand P,
Guha S and Aggarwal BB: Curcumin circumvents chemoresistance in
vitro and potentiates the effect of thalidomide and bortezomib
against human multiple myeloma in nude mice model. Mol Cancer Ther.
8:959–970. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Bharti AC, Takada Y and Aggarwal BB:
Curcumin (diferuloylmethane) inhibits receptor activator of
NF-kappa B ligand-induced NF-kappa B activation in osteoclast
precursors and suppresses osteoclastogenesis. J Immunol.
172:5940–5947. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jana S, Paul S and Swarnakar S: Curcumin
as anti-endometriotic agent: implication of MMP-3 and intrinsic
apoptotic pathway. Biochem Pharmacol. 83:797–804. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang ME, Chen YC, Chen IS, Hsieh SC, Chen
SS and Chiu CH: Curcumin protects against thioacetamide-induced
hepatic fibrosis by attenuating the inflammatory response and
inducing apoptosis of damaged hepatocytes. J Nutr Biochem.
23:1352–1366. 2012. View Article : Google Scholar : PubMed/NCBI
|