Introduction
Gliomas are intrinsic brain tumours which are rare,
with an incidence of approximately 10 per 100,000 capita (1). The most common type of glioma is
glioblastoma (GB) which is classified as grade IV. It is one of the
most challenging types of cancer to treat and remains incurable. Of
the biological features, such as proliferation, angiogenesis and
invasion which are common to all cancers, perhaps the most
significant one is invasion. Some matrix metalloproteinases or MMPs
are zinc-dependent endopeptidases and have a role in degradation of
the extracellular matrix (ECM). Originally, MMPs were associated
with invasion and metastasis exclusively. Recent studies have
documented the additional involvement of MMPs in several steps of
cancer development which includes cell growth, apoptotic activity,
angiogenesis and immune responses to cancer (2,3). MMPs
also regulate cell growth through cleavage of cell surface-bound
growth factors and receptors, apoptosis through release of death or
survival factors and cell motility by revealing cryptic matrix
signals or cleavage of adhesion molecules, releasing growth factors
sequestered in the ECM or by integrin signalling (4). Also, cell-surface anchored MMPs or
membrane-type MMPs (MT-MMPs) are expressed in gliomas which mediate
extensive local intracerebral invasion and are high in
vascularisation of the tumour (5).
MMP dysregulation and the imbalance between metalloproteases and
their inhibitors (TIMPs) have been implicated as a potential
predictor of ECM production and/or degradation (6,7). TIMPs
are documented widely for their roles on the induction of apoptosis
in different cancer cells in vitro (8).
The prognosis of GB patients remains dismal despite
conventional therapeutic approaches such as radio- and chemotherapy
and various novel surgical approaches that have been introduced for
their management. The current clinical practice guideline for
diagnosis, treatment and follow-up of malignant gliomas has been
reviewed by Stupp et al (9).
Apart from surgery as the initial therapeutic approach,
implantation of carmustine polymers into the resection cavity may
be performed prior to radiotherapy. Chemotherapy, on the other
hand, is the standard care for GB patients with temozolomide (TMZ)
administered daily at a low dose of 75 mg/m2 ~1.5 h
before radiotherapy. Radiotherapy may be administered either as a
standard fractionated focal radiotherapy (e.g. 60 Gy in 30–33
fractions) or in shorter hypofractionated regimens (e.g. 40 Gy in
15 fractions), depending on the patient’s age and condition.
Previous studies (10,11) have suggested that Gliadel wafer
(Carmustine) may offer hope of long-term survival in malignant
glioma patients but adverse systemic effects in some cases were
reported. Nevertheless, there is still a strong need for other
therapeutic interventions in the management of malignant gliomas to
improve the quality of life and possibly the survival time.
Interest in studies on flavonoids has increased due
to their proposed protective role in atherosclerosis and cancer
(12). Moreover, one novel
approach, proposed by Rooprai et al (13) is based on the hypothesis that
micronutrients, such as citrus flavonoids (from tangerine peel) may
have the potential to serve as anticancer agents in the treatment
of malignant gliomas. Their efficacy is based on their ability to
interfere with parameters of invasion. We have proposed the use of
a combination of micronutrients, including selenium and tangeretin,
known to have a variety of therapeutic properties such as anti-
invasive and pro-apoptotic to combat brain tumours (14). We have included curcumin and
polyphenolics from Aronia melanocarpa as part of a very
large study on micronutrients screened in vitro to evaluate
their therapeutic ability, particularly that of the induction of
apoptosis and inhibition of invasion in GBs.
Curcumin (diferuloylmethane; 1,7-bis (4-hydroxy
3-methoxyphenyl)-1,6-heptadiene-3,5-dione) is a polyphenol derived
from Curcuma longa (turmeric), a medicinal plant. Turmeric
contains 3 main components; curcumin (77%), demethoxycurcumin (17%)
and bisdemethoxycurcumin (3%). Together, these components are
referred to as curcuminoids (15,16).
Curcumin has also been reported to cross the rat blood-brain
barrier (BBB) (17). Its
pro-apoptotic potential has been reported in other types of somatic
cancer such as breast, lung and colon (18–20).
It also has a strong safety profile and a number of potent
pleiotropic actions for neuroprotective efficacy. Extensive
research has demonstrated that curcumin affects several targets
including transcription factors, growth regulators, adhesion
molecules, apoptotic genes, angiogenesis regulators and cellular
signalling molecules (21).
Moreover, other researchers have suggested its mode of action in
that there could be participation of the death receptor pathway
through Fas-ligand interaction, caspase-3 activation (22) caspase-8, BID cleavage, cytochrome c
release and anti-apoptotic gene suppression (Bcl-2 and Bcl-xL)
(22,23).
Polypeholics from Aronia melanocarpa
(chokeberry)obtained by solvent extraction (ethanol solution)
include a range of flavonoids, such as chlorogenic acid, (−)
epicatechin, (+) catechin, rutin (flavones) and anthocyanins
including cyanidin-3-galactoside (the primary anthocyanin in
polyphenolics from Aronia melanocarpa),
cyanidin-3-arabinose, cyanidin-3-xyloside and cyanidin-3-glucoside.
Some of these epicatechins are also found in green tea, black tea,
red wine, and cocoa, which include epigallocatechin gallate,
resveratrol, and procyanidin. They have also been extensively
investigated due to their possible role as chemopreventive agents
based on their antioxidant capacities. In addition, catechins were
found to inhibit hepatocyte growth factor receptor (MET kinase)
activation in human colon cancer cells, possibly by competing for
the kinase domain of the MET protein (24).
Apart from our own research on the anticancer
effects of polyphenolics from Aronia melanocarpa on
malignant brain tumours (14) the
only other group who studied its effects in colorectal cancer cell
lines have shown it to inhibit cell growth (25,26).
The anthocyanins found in this extract are of particular interest
because they are non-toxic and are believed to cross the BBB
(27).
Thus, the aim of this study was to investigate if
curcumin and polyphenolics from Aronia melanocarpa could
induce apoptosis and effect mediators of invasion by looking at the
gene profiles of 8 selected MMPs and 4 TIMPs in an established
glioblastoma cell line (U373) in vitro compared to the
normal astrocytic culture.
Materials and methods
Micronutrients
Curcumin
Curcumin is an orange-coloured powder which was
donated by Indus Biotech, India. It was 97% natural and was
obtained from ground turmeric rhizome by a standard solvent
extraction method. Prior to use for treatment of cell cultures, it
was dissolved in dimethyl sulfoxide (DMSO-Sigma Aldrich USA).
Polyphenolics from Aronia
melanocarpa
The polyphenolics extract from Aronia
melanocarpa is a dark purple-coloured powder which was donated
by Artemis, USA. It is a purified extract containing a number of
flavonoids, including cholorogenic acid (a phenolic acid that is
dominant among the aromatic acids) (−) - epicatechin, (+) -
catechin (catechins), rutin (a flavones) and a range of
anthocyanins: cyanidin-3-O-galactoside, cyanidin-3-O-arabinoside
and cyanidin-3-xyloside (14).
Polyphenolics from Aronia melanocarpa was also diluted in
DMSO prior to use.
Cell cultures
An established cell line, U373, from an adult
patient with GB (grade IV, WHO Classification) was used in this
study. It was kindly donated by Professor Rolf Bjerkvig (Norway)
and has been tested for mycoplasma screening and confirmed to be of
human origin by molecular genetics methods. The control cell
culture for this study was normal astrocytes, (CC2565) used at
passage 11. It was derived from normal human brain cells of an
18-year old male and purchased from Cambrex BioSciences. These two
cell lines were routinely cultured as monolayers in complete
medium, i.e. Dulbecco’s modified Eagle’s medium (DMEM, Sigma
Aldrich USA) supplemented with 10% foetal calf serum and
antibiotics (at the final concentration of 1%
penicillin/streptomycin per ml). The physiological pH was
maintained through equilibration with 5% CO2 atmosphere
in a 37°C humidified incubator.
IC50 determination using
the MTT cytotoxicity assay
(3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide or
MTT is a residual viable biomass assay dependent on the conversion
of MTT to insoluble formazan within living cells, which is
subsequently solubilised in DMSO (28). It was used to determine the
IC50 value for each micronutrient. The formazan in the
solution was quantified spectrophotometrically at 570 nm as an
indicator of cell viability. Assays were carried out in triplicates
in 96-well culture plates. Both CC2565 cell cultures and the U373
cell line were compared when treated separately with each
micronutrient.
Detection of apoptosis with
Annexin-FITC using flow cytometry
The binding of the calcium-dependent
phosphatidylserine binding protein Annexin V to externalised
residues is a hallmark of apoptosis in many cells. A
fluorochrome-labelled Annexin V can be used in combination with a
DNA-binding dye such as propidium iodide to differentiate between
live cells (negative for both dyes), cells in early apoptosis
(Annexin positive but propidium iodide negative) and dead cells
(positive for both dyes). Cells were treated with a range of
concentrations of curcumin and chokeberry and the percentage of
live, dead and apoptotic cells were assessed. Cells were harvested
and washed before being re-suspended in 500 μl of Annexin-V binding
buffer (Pharmingen, USA) with 2.5 μl Annexin-V-FITC (Pharmingen)
and incubated at room temperature for 15 min. Propidium iodide (50
μl) (50 μg/ml, Sigma) was added just before samples were measured.
Samples were analysed on a FACSCalibur (Becton-Dickinson, USA) with
FITC fluorescence being measured using a 530/30 bandpass filter and
propidium iodide fluorescence being measured using a 670 nm
longpass filter. At least 10,000 events were acquired and data were
analysed using the CellQuest software (Becton-Dickinson).
RNA extraction
After 48-h treatment with selected concentrations of
either curcumin or polyphenolics from Aronia melanocarpa,
cells were harvested using Cell Dissociation Solution (Sigma)
before 2 ml of RNAzol reagent (Biogenesis) was added to the pellet.
The next steps were carried out at 4°C. Chloroform (0.2 ml) was
added to the solubilised RNA and vortexed for 15 sec followed by
centrifugation at 4,000 rpm for 30 min. Equal volumes of both the
supernatant (800 μl) taken from the upper phase and isopropanol
were mixed. The sample was incubated for 15 min and centrifuged at
4,000 rpm for 30 min before 2 ml of 70% ethanol was added to the
pellet. This centrifugation was repeated twice for 15 min, each.
The pellet was then resuspended in 100 μl of double autoclaved
water and incubated in a water bath for 15 min at 60°C as to
dissolve the RNA. The RNA (100 μl) was then diluted in 300 μl of
double distilled water for measuring the optical density at 260 nm
in order to determine the RNA concentration. Each sample (3 μl) was
diluted (x100) in double autoclaved water prior to
spectrophotometric analysis, in triplicate. The control used was
double autoclaved distilled water. RNA concentration was calculated
as follows: Mean value × 100 (dilution factor) × 40 μg/ml
(conversion factor for RNA).
Reverse transcription (RT)
reactions
An aliquot (1 μg) of total RNA was reverse
transcribed using 2 μg of random hexamers (Amersham) and 200 units
of Superscript II reverse transcriptase (Invitrogen). The sequences
for primers and probes have been previously described (5) whereas 18S RNA was used as an
endogenous control to account for differences in the extraction and
reverse transcription of the total RNA. PCR was performed using the
manufacturer’s protocol for ABI Prism 7700 Sequence Detection
System (Applied Biosystems). This cDNA synthesis reaction mixture
was pre-incubated for 10 min at 21°C. Reverse transcription
reaction was carried out for 50 min at 42°C and then heated to 95°C
for 5 min to stop the reaction.
Quantitative PCR (polymerase chain
reaction)
PCRs were performed in 25-μl reaction volumes. Each
reaction volume contained the equivalent of 5 ng of the RT reaction
product, 50% TaqMan 2X PCR Master mix (Applied Biosystem) 100 nM of
each forward and reverse primer and 200 nM probe. Each PCR cycle
consisted of a heat denaturation step for 94°C for 1 min, a
primer-annealing step at 50°C for 30 sec and a polymerization step
for 1 min at 72°C. To determine the relative RNA levels within the
samples, standard curves for the PCR reaction were prepared by
using the cDNA from a standard sample and making 2-fold serial
dilutions covering the range equivalent to 20–0.625 ng of RNA; for
18S analyses, the range was 4–0.125 ng. Messenger RNA levels were
normalized to 18S rRNA levels and are presented with the levels in
the control sample set to 1 (29).
Statistical analysis
The absorbance values obtained from the MTT assay
were analysed using the StatsDirect and Microcal (Origin)
programmes. The average of 6-wells was taken for each concentration
studied and compared with the control cells treated with a medium
(100% viable) in order to obtain the percentage of residual viable
cells. The dose response curve formed (of each micronutrient)
conferred the IC50 values.
Results
Curcumin and polyphenolics from Aronia
melanocarpa reduce glioma cell viability
The MTT viability assay was used to assess the
cytotoxicity of curcumin and polyphenolics from Aronia
melanocarpa on normal astrocytic cultures, CC2565 at passage
number 11 by treating them with the micronutrients compared to
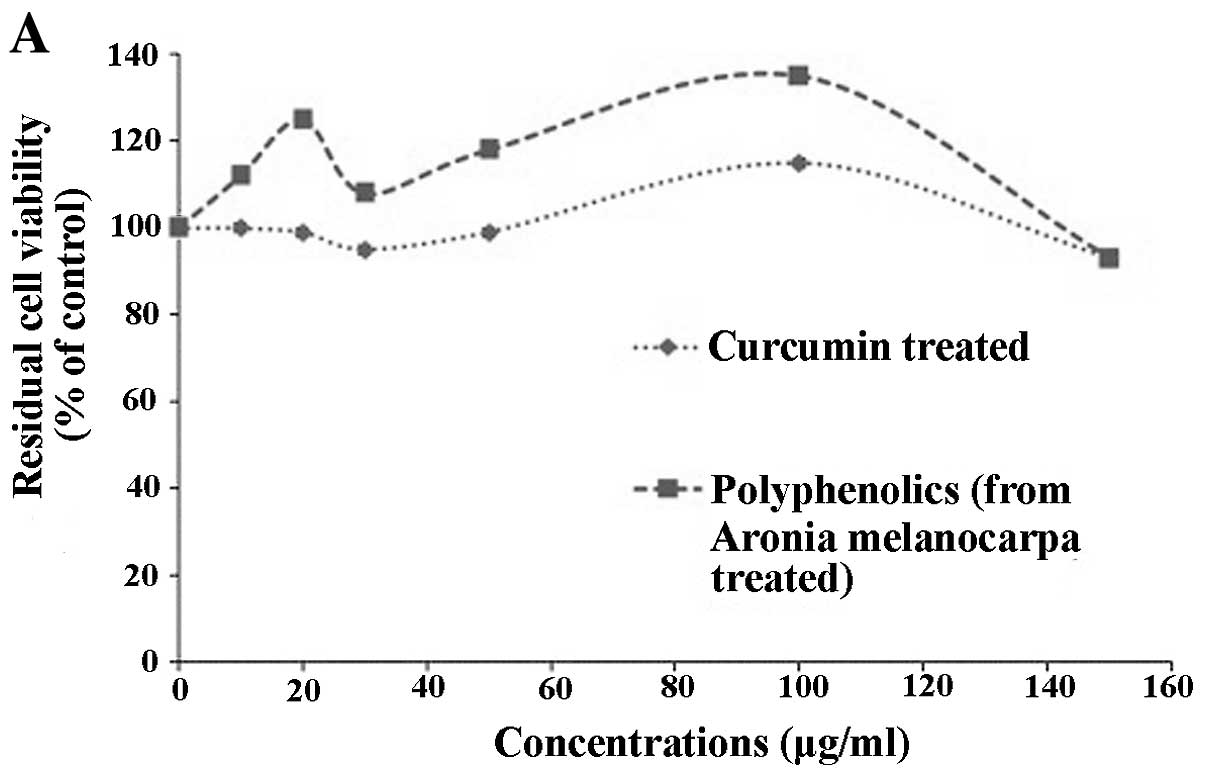
untreated cells (with DMEM only). The data obtained (Fig. 1A) show that generally, curcumin and
polyphenolics from Aronia melanocarpa, at all the
concentrations selected for the study, were not toxic (>90%
viability) to them. In contrast, the U373 established brain tumour
cell line, showed a decrease in cell viability in a dose-responsive
manner (Fig. 1B and C). The
IC50 values for U373, determined for curcumin and
polyphenolics from Aronia melanocarpa were 15 and 200 μg/ml,
respectively. These results indicate that curcumin and
polyphenolics from Aronia melanocarpa, at the concentrations
analysed, are not toxic to normal astrocytic cells but induce cell
death in the U373 cell line.
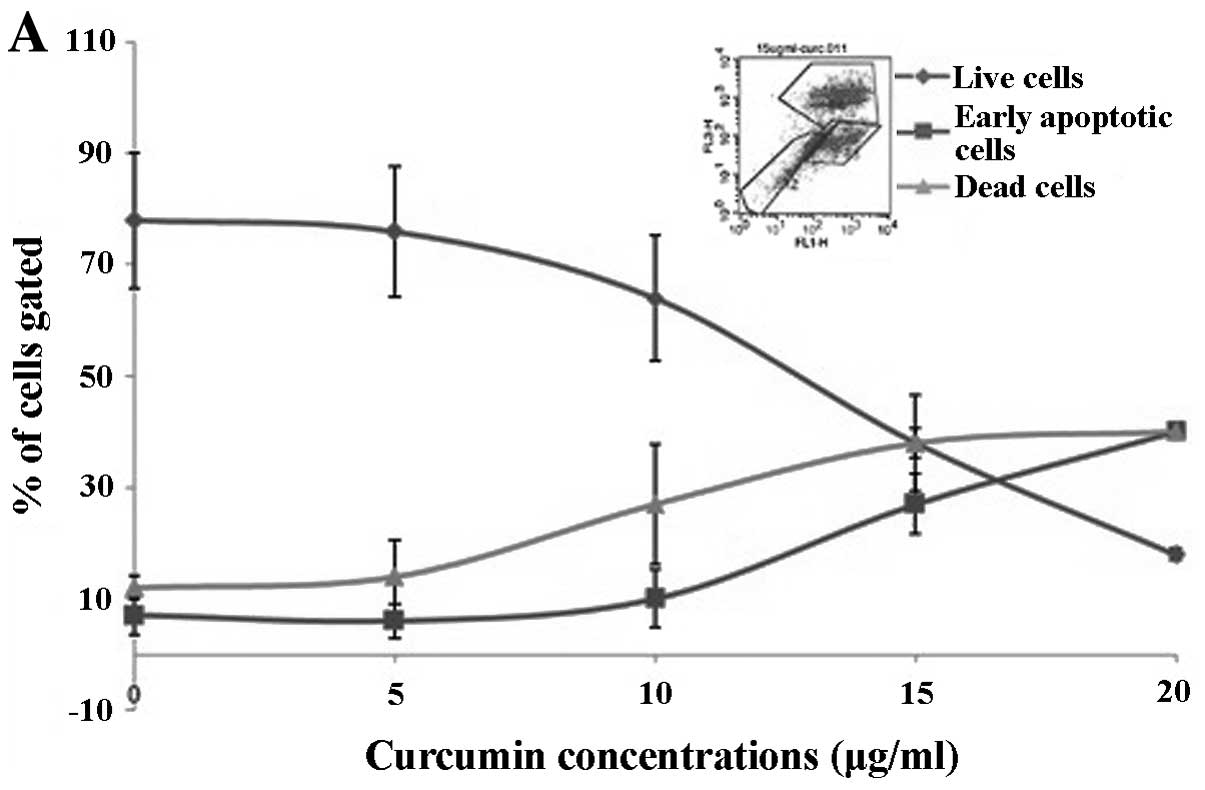
Curcumin and polyphenolics from Aronia
melanocarpa cause cell death by different mechanisms
To evaluate the effects of curcumin and
polyphenolics from Aronia melanocarpa, flow cytometric
assessment of apoptosis and cell death by the Annexin V/propidium
iodide assay was carried out. The advantage of the flow cytometry
is that it is possible to measure not just dead cells but also
early apoptotic cells (30). The
ability to induce cell death by polyphenolics from Aronia
melanocarpa within a range of different concentrations, 30–60
μg/ml (Fig. 2B) and curcumin, 5–20
μg/ml (Fig. 2A), was then assessed,
closer to their IC50 values determined by the MTT
viability assay.
For the polyphenolics from Aronia
melanocarpa, all concentrations studied induced more necrosis
(82%) than apoptosis (11%) in the cells particularly at the highest
concentration of 600 μg/ml (Fig.
2B). This population included the late apoptotic cells (as seen
with both propidium iodide and Annexin V positivity). Percentages
of live, early apoptotic and dead cells were calculated in
comparison to the untreated cells and a representative scatter plot
was included next to each graph plotted to illustrate each cell
population gated. Low percentages of the early apoptotic cells were
gated almost at the same extent at all concentrations studied
(1–15%). The highest two concentrations caused the apoptotic
population of cells to increase but only up to 14 and 11% for cells
treated with 400 and 600 μg/ml of polyphenolics from Aronia
melanocarpa, respectively.
In contrast, higher percentages of the early
apoptotic cell population were seen following treatment with
curcumin, particularly at concentrations of 15 and 20 μg/ml when
the apoptotic cells gated were 26 and 41%, respectively (Fig. 2A). When comparing early apoptotic
and dead cell populations gated, it was seen that as curcumin
concentrations increased, the dead cell population increased more
than the apoptotic one. Interestingly, at the highest concentration
of curcumin (20 μg/ml) used, both early apoptotic and dead cells
appeared to share the same effect due to similar percentage of
cells (~40% for each population) gated. The representative scatter
plots (Fig. 2) show that both
curcumin and polyphenolics from Aronia melanocarpa cause
cell death in the glioma cell line, while polyphenolics from
Aronia melanocarpa mainly induce necrosis. The effect of
curcumin is mediated equally by necrosis and early apoptosis.
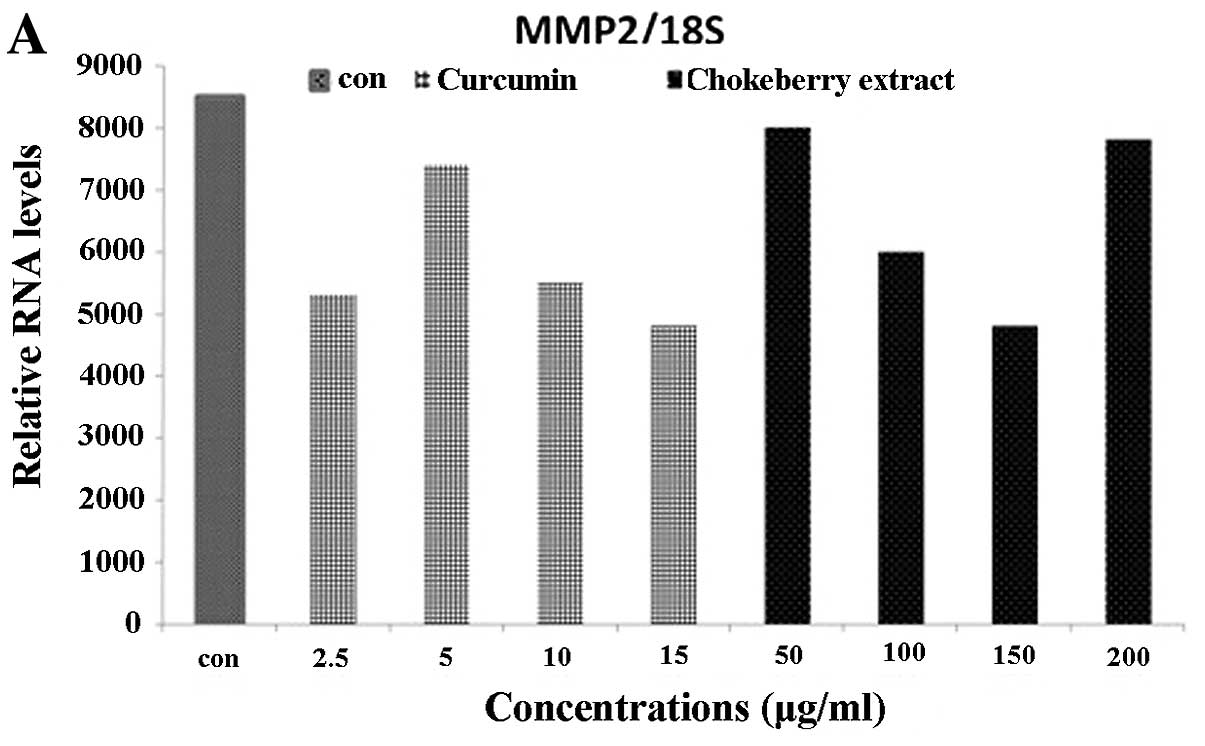
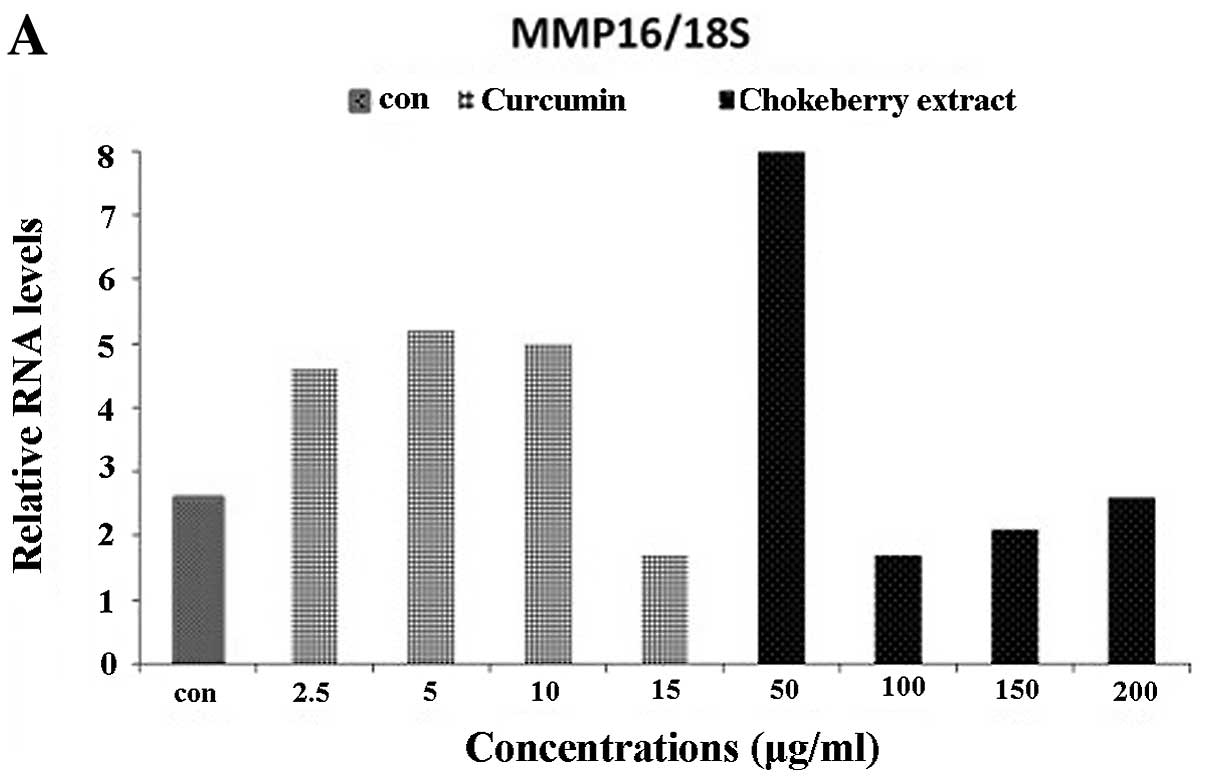
Reduction in MMP gene expression with
polyphenolics from Aronia melanocarpa
Relative RNA expression levels of 8 MMPs (MMP-2, 9,
14, 15, 16, 17, 24 and 25) in the U373 cell line following
treatment with ranges of curcumin 10–50 μg/ml or polyphenolics
50–400 μg/ml from Aronia melanocarpa as shown in Figs. 3 and 4. RNA expression was normalized to 18S
rRNA levels and is expressed relative to the levels in untreated
cells. Generally, the results in Figs.
3 and 4 show that RNA levels of
all the MMPs and TIMPs studied were reduced compared to the
untreated cells at the different concentrations of the
micronutrients. However, the extent of reduction in each case was
different as seen in the raw data. In some cases such as MMP-9, a
minimum amount was expressed either in the control or
curcumin/polyphenolics from Aronia melanocarpa-treated
cells. The most prominent reduction was seen in MMP-2 (Fig. 3A) whereas MMP-14, −15 (Fig. 3C-D) and −16 (Fig. 4A) showed slight reduction of the RNA
levels at both IC50 values of each micronutrient. A
similar reduction was seen with MMP-17 (Fig. 4B) but to a greater extent in cells
treated with 200 μg/ml polyphenolics from Aronia
melanocarpa. The level of MMP-24 (Fig. 4C) remained unchanged at the
IC50 value for curcumin but was slightly increased for
the IC50 value of polyphenolics from Aronia
melanocarpa. With MMP-25 (Fig.
4D), negligible changes were observed with either polyphenolics
from Aronia melanocarpa or curcumin at all concentrations
studied.
Discussion
The present study showed that both curcumin and
polyphenolics from Aronia melanocarpa were non-toxic to the
control human brain cell culture, (CC2565) as illustrated in
Fig. 1A. This confirms earlier
reports that curcumin is safe to be consumed even at higher
concentrations because it does not cause harmful effects on normal
cells. Moreover, extensive clinical and metabolic studies including
those of Aggarwal et al (15) have reported that 8–12 g/day is safe
to be consumed (31). Similarly,
the non-toxic effects of anthocyanins and polyphenolics from
Aronia melanocarpa are consistent with other reports in
malignant colon cancer cells studies (32).
Generally, the residual cell viability decreased as
the concentrations increased, suggesting that very high
concentrations could have been toxic to the cells. The
IC50 value for curcumin was 15–20 μg/ml (Fig. 1B). A similar pattern was seen for
cells treated with polyphenolics from Aronia melanocarpa
giving an IC50 of ~200 μg/ml (Fig. 1C). These values probably could not
be achieved physiologically as both curcumin and polyphenolics from
Aronia melanocarpa have rapid and extensive metabolisms
which explains their poor bioavailability. However, as U373 is an
established GB cell line, which has homogenous cell populations of
cells, probably stem cells, unlike the low passage biopsy-derived
primary cultures which are heterogenous. It is therefore not
surprising that it requires a high IC50 value for either
micronutrient compared to primary cultures. The IC50
value was much lower (30 μg/ml) when biopsy-derived cell cultures
were treated with polyphenolics from Aronia melanocarpa
(unpublished data). A brain tumour stem cell study by Inagaki et
al (33) demonstrated a small
population of cancer stem cells (CSCs) with preserved stem cell
properties even after many serial passages under
non-adherent/adherent culture conditions. Other studies have
suggested that cells experiencing in vitro propagation are
likely to be different from the original tumour’s signature,
possibly being responsible for the attenuated resistance observed
(34–36).
Our results also showed that there was a variation
in the proportion of apoptotic and dead cells which could be due to
the cells’ response depending on the state of the cells at the
time. In addition, it also suggested that at high concentration,
the method of cell death may have changed. As seen in Fig. 2B, polyphenolics from Aronia
melanocarpa show a rapid killing from 200 μg/ml upwards whereas
curcumin (Fig. 2A) is more subtle
from 5 μg/ml. Hence, it is implied that unlike curcumin,
polyphenolics from Aronia melanocarpa may not be apoptotic.
In addition, the polyphenolics from Aronia melanocarpa may
appear to kill the cells by a non-apoptotic (or at least a
non-Annexin positive) pathway whereas curcumin appears to target
Annexin V positive populations. The percentage of dead cells as
indicated by propidium iodide staining shows that this did not
change significantly over the range of concentrations used, but the
percentage of apoptotic cells did. This may account for the
discrepancy seen with the MTT assay which generally measures a
final end point rather than early apoptosis. The MTT viability
assay detects metabolically active cells which then do not
necessarily correlate with the flow cytometry data. Recently, other
workers have reported the induction of apoptosis by curcumin in
somatic cancers where the degree of response may not be comparable
to our study on brain tumours. Saha et al showed that at 50
μM (18.4 μg/ml) induced apoptosis in 60% of human lung cancer cells
(37). Moreover, a similar effect
was seen at 10 μmol/l (3.7 μg/ml) in melanoma cells (38) and 20–75 μM (7–28 μg/ml) in breast
and colon cancer (18,20). Perhaps in these different types of
cancer, low concentrations may cause apoptosis while higher ones
may lead to other forms of cell death, such as necrosis and
autophagy.
However, to the best of our knowledge, there are no
other reports in the literature for Aronia melanocarpa’s
potential for induction of apoptosis except for our own recent
findings in another study on its powerful anti-invasive potential
in GB cells (unpublished data). Nevertheless, polyphenolics from
Aronia melanocarpa have been reported to have inhibitory
effects on cell growth and cell cycle at a range of 50–75 μg/ml
(25,26) but non-cytotoxic on healthy human
tissue.
Generally, both curcumin and polyphenolics from
Aronia melanocarpa were found to downregulate the RNA levels
of all the MMP and TIMP RNA studied at their IC50
values. High levels of MMP-2 which has been implicated in invasion
were detected in the untreated cells. Both micronutrients
downregulated MMP2 levels in this study, suggesting their
anti-invasive potential. Whereas, in the U373 cells, hardly any
MMP-9 (Fig. 3B) and MMP-25
(Fig. 4D) expression could be
detected, either in the untreated or curcumin/polyphenolics from
Aronia melanocarpa-treated cells.
It is well documented that high levels of MT-MMPs,
such as MMP-14 are expressed by gliomas and it facilitates MMP-2
activation thus playing a role in the regulation of invasiveness
(39). Although our results show
that there were some increases at the concentrations below the IC
value of either curcumin or polyphenolics from Aronia
melanocarpa in MMP-14 (Fig. 3C)
and −15 (Fig. 3D) the expression of
both MMPs was reduced to a similar extent by 15 μg/ml curcumin when
compared with the control. MMP-16 expression was also reduced at
all concentrations studied (Fig.
4A). Thus, the reduction of the expression of MMP-14, −15 and
−16 may suggest inhibition of invasion. MMP-24 is a cell
membrane-anchored MMP which is brain specific and mainly expressed
in the cerebellum (40).
Surprisingly, its levels elevated generally with both
micronutrients (Fig. 4C), similar
to the effect seen on MMP-25 with the treatment of selenium
(41).
In conclusion, it is possible that curcumin and
polyphenolics from Aronia melanocarpa with its complex and
ranged phenolic compounds could selectively induce apoptosis or
inhibit invasion in the established malignant brain tumour cells,
U373. Further studies include comparison of the therapeutic
potential of these and other micronutrients in primary cultures,
particularly with the use of spheroids in 3D invasion assays and
in vitro angiogenesis assays.
Acknowledgements
The authors are grateful to the Have a Chance Inc.,
RoseTrees Trust, CALM and Ministry of Higher Education, Malaysia
for their financial support.
References
|
1
|
McKinney PA: Brain tumours: incidence,
survival and aetiology. J Neurol Neurosurg Psychiatry. 75:1–12.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Egelblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nature Rev.
2:161–174. 2002.PubMed/NCBI
|
|
3
|
Bourboulia D and Stetler-Stevenson WG:
Matrix metalloproteinases (MMPs) and tissue inhibitors of
metalloproteinases (TIMPs): positive and negative regulators in
tumor cell adhesion. Semin Cancer Biol. 20:161–168. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cauwe B, Van den Steen PE and Opdenakker
G: The biochemical, biological, and pathological kaleidoscope of
cell surface substrates processed by matrix metalloproteinases.
Crit Rev Biochem Mol Biol. 42:113–185. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Nuttal RK, Pennington CJ, Taplin J, Wheal
A, Yong VW, Forsyth PA and Edwards DR: Elevated membrane-type
matrix metalloproteinases in gliomas revealed by profiling
proteases and inhibitors in human cancer cells. Mol Cancer Res.
1:333–345. 2003.PubMed/NCBI
|
|
6
|
Gomez DE, Alonso DF, Yoshiji H and
Thorgeirsson UP: Tissue inhibitors of metalloproteinases:
structure, regulation and biological functions. Eur J Cell Biol.
74:111–122. 1997.PubMed/NCBI
|
|
7
|
Skrzydlewska E, Sulkowska M, Koda M and
Sulkowski S: Proteolytic-antiproteolytic balance and its regulation
in carcinogenesis. World J Gastroenterol. 11:1251–1266. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baker AH, Edwards DR and Murphy G:
Metalloproteinase inhibitors: biological actions and therapeutic
opportunities. J Cell Sci. 115:3719–3727. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Stupp R, Tonn JC, Brada M and
Pentheroudakis G: High-grade malignant glioma: ESMO clinical
practice guidelines for diagnosis, treatment and follow-up. Ann
Oncol. 21:V190–V193. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Westphal M, Hilt DC, Bortey E, Delavault
P, Olivares R, Warnke PC, Whittle IR, Jaaskelainen J and Ram Z: A
phase 3 trial of local chemotherapy with biodegradable carmustine
(BCNU) wafers (Gliadel wafers) in patients with primary malignant
glioma. Neuro Oncol. 5:79–88. 2003.PubMed/NCBI
|
|
11
|
Weber EL and Goebel EA: Cerebral edema
associated with gliadel wafers: two case studies. Neuro Oncol.
7:84–89. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tapiero H, Tew KD, Nguyen Ba G and Mathe
G: Polyphenols: do they play a role in the prevention of human
pathologies? Biomed Pharmacother. 56:200–207. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Rooprai HK, Kandanearatchi A, Maidment SL,
Christidou M, Trillo-Pazos G, Dexter DT, Rucklidge GJ, Widmer W and
Pilkington GJ: Evaluation of the effects of swainsonine, captopril,
tangeretin and nobiletin on the biological behaviour of brain
tumour cells in vitro. Neuropathol Appl Neurobiol. 27:29–39. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Rooprai HK, Christidou M and Pilkington
GJ: The potential for strategies using micronutrients and
heterocyclic drugs to treat invasive gliomas. Acta Neurochir
(Wien). 145:683–690. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aggarwal BB, Kumar A and Bharti AC:
Anticancer potential of curcumin: preclinical and clinical studies.
Anticancer Res. 23:363–398. 2003.PubMed/NCBI
|
|
16
|
Sharma RA, Gescher AJ and Steward WP:
Curcumin: the story so far. Eur J Cancer. 41:1955–1968. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Datla KP, Zbarsky V and Dexter D: Effects
of anaesthetics on the loss of nigrostriatal dopaminergic neurons
by 6-hydroxydopamine in rats. J Neural Transm. 113:583–591. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pillai GR, Srivastava AS, Hassanein TI,
Chauhan DP and Carrier E: Induction of apoptosis in human lung
cancer cells by curcumin. Cancer Lett. 208:163–170. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Simon A, Allais DP, Durox JL, Basly JP,
Durand-Fontainer S and Delage C: Inhibitory effect of curcuminoids
on MCF-7 cell proliferation and structure-activity relationships.
Cancer Lett. 129:111–116. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Shi M, Cai Q, Yao L, Mao Y, Ming Y and
Ouyang G: Antiproliferation and apoptosis induced by curcumin in
human ovarian cancer cells. Cell Biol Int. 30:221–226. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Anand P, Sundaram C, Jhurani S,
Kunnumakkara AB and Aggarwal BB: Curcumin and cancer: an ‘old-age’
disease with an ‘age-old’ solution. Cancer Lett. 267:133–164.
2008.
|
|
22
|
Duvoix A, Roman B, Delhalle S,
Schnekenburger M, Morceau F, Henry E, Dicato M and Diederich M:
Chemopreventive and therapeutic effects of curcumin. Cancer Lett.
223:181–190. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Anto RJ, Mukhopadhyay A, Dening K and
Aggarwal BB: Curcumin (diferuloylmethane) induces apoptosis through
activation of caspase-8, BID cleavage and cytochrome c release: its
suppression by ectopic expression of Bcl-2 and Bcl-xl.
Carcinogenesis. 23:143–150. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Larsen CA, Bisson WH and Dashwood RH: Tea
catechins inhibit hepatocyte growth factor receptor (MET kinase)
activity in human colon cancer cells: kinetic and molecular docking
studies. J Med Chem. 52:6543–6545. 2009. View Article : Google Scholar
|
|
25
|
Malik M, Zhao C, Schoene N, Guisti MM,
Moyer MP and Magnuson BA: Anthocyanin-rich extract from Aronia
meloncarpa E induces a cell cycle block in colon cancer but not
normal colonic cells. Nutr Cancer. 46:186–196. 2003.
|
|
26
|
Zhao C, Giusti MM, Malik M, Moyer MP and
Magnuson BA: Effects of commercial anthocyanin-rich extracts on
colonic cancer and nontumorigenic colonic cell growth. J Agric Food
Chem. 52:6122–6128. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Andress-Lacueva C, Shukitt-Hale B, Galli
RL, Jaurengui O, Lamuela-Raventos RM and Joseph JA: Anthocyanins in
aged blueberry-fed rats are found centrally and may enhance memory.
Nutr Neurosci. 8:111–120. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Nikkah G, Tonn JC, Hoffmann O, Kraemer HP,
Darling JL, Schahenmayr W and Schonmayr R: The MTT assay for
chemosensitivity testing of human tumours of the central nervous
system. J Neurooncol. 13:13–24. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pennington CJ and Edwards DR: Real time
PCR expression profiling of MMPs and TIMPs. Matrix
Metalloproteinase Protocols. Methods Mol Biol. 622:159–173. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Vermes I, Haanen C, Steffens-Nakken H and
Reutelingsperger C: A novel assay for apoptosis. Flow cytometric
detection of phosphatidylserine expression on early apoptotic cells
using fluorescein labelled Annexin V. J Immunol Methods. 184:29–51.
1995. View Article : Google Scholar
|
|
31
|
Anand P, Kunnumakkara AB, Newman RA and
Aggarwal BB: Bioavailability of curcumin: problems and promises.
Mol Pharm. 4:807–818. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhang Y, Vareed SK and Nair MG: Human
tumor cell growth inhibition by nontoxic anthocyanidins, the
pigments in fruits and vegetables. Life Sci. 76:1465–1472. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Inagaki A, Soeda A, Oka N, Kitajima H,
Nagakawa J, Motohashi T, Kunisada T and Iwama T: Long-term
maintenance of brain tumor stem cell properties under at
non-adherent and adherent culture conditions. Biochem Biophys Res
Commun. 361:586–592. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Szakacs G and Gottesman MM: Comparing
solid tumours with cell lines: implications for identifying drug
resistance genes in cancer. Mol Interv. 4:323–325. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Singh SK, Clarke ID, Terasaki M, et al:
Identification of a cancer stem cell in human brain tumors. Cancer
Res. 63:5821–5828. 2003.PubMed/NCBI
|
|
36
|
Galli R, Binda E, Orfanelli U, et al:
Isolation and characterization of tumorigenic, stem-like neural
precursors from human glioblastoma. Cancer Res. 64:7011–7021. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Saha A, Kuzuhara T, Echigo N, et al:
Apoptosis of human lung cancer cells by curcumin mediated through
up-regulation of ‘growth arrest and DNA damage inducible genes 45
and 153’. Biol Pharm Bull. 33:1291–1299. 2010.
|
|
38
|
Bill MA, Fuchs JR, Li C, et al: Curcumin
induces proapoptotic effects against human melanoma cells and
modulates the cellular response to immunotherapeutic cytokines. Mol
Cancer Ther. 8:2726–2735. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Nakada M, Kita D, Futami K, Yamashita J,
Fujimoto N, Sato H and Okada Y: Roles of membrane type 1 matrix
metalloproteinases and tissue inhibitor of metalloproteinases 2 in
invasion and dissemination of human malignant glioma. J Neurosurg.
94:464–473. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sekine-Aizawa Y, Hama E, Watanabe K,
Tsubuki S, Kanai-Azuma M, Kanai Y, Arai H, Aizawa H, Iwata N and
Saido TC: Matrix metalloproteinase (MMP) system in brain:
identification and characterization of brain-specific MMP highly
expressed in cerebellum. Eur J Neurosci. 13:935–948. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Rooprai HK, Kyriazis I, Nuttall RK,
Edwards DR, Zicha D, Aubyn D, Davies D, Gullan R and Pilkington GJ:
Inhibition of invasion and induction of apoptosis by selenium in
human malignant brain tumour cells in vitro. Int J Oncol.
30:1263–1271. 2007.PubMed/NCBI
|