Introduction
Non-Hodgkin’s lymphoma (NHL) represents
heterogeneous lymphoproliferative malignancies with differing
patterns of behavior and therapy response (1). Traditionally, ionizing radiation (IR)
therapy plays an important role in the management of NHL. IR alone
is an effective therapy for stages I and II in indolent NHL
patients. While indolent and aggressive NHLs are responsive to IR
and chemotherapy, approximately 50–70% of the patients relapse
(2). It is suggested that a poor
prognosis of NHL may be due to its resistance to cell death
induction by IR or chemotherapy. Therefore, future improvements in
the therapeutic index for radiotherapy are required to target
resistant cells and improve efficacy without toxicity.
Curcumin (diferuloylmethane), a major constituent of
turmeric powder, is extracted from the rhizomes of the plant
Curcuma longa. Numerous studies have demonstrated the
effectiveness of curcumin as an anticancer drug or chemopreventive
agent in laboratory animal models of human carcinogenesis including
NHL (3–6) without cytotoxic effects on healthy
cells (7). Inhibition of cell
growth and induction of apoptosis is the common mechanism by which
curcumin demonstrates its anticancer effects. In addition to the
effectiveness of curcumin alone, it is being currently evaluated in
combination therapy with IR or chemotherapy. Curcumin has exhibited
varying effects on radiation sensitivity in different cancer cell
types and its effect as a radiosensitizer has been supported in a
variety of tumors including prostate, colorectal and ovarian
cancers (8–12). Likewise, we previously demonstrated
that curcumin may enhance IR-induced apoptosis in NHL (13).
Accumulating evidence suggests that multiple
signaling pathways, including inhibition of nuclear factor-κB
(NF-κB), are involved in mediating the effect of curcumin on the
growth suppression of human cancer cells (4,14–18).
The mammalian target of rapamycin (mTOR), an atypical
serine/threonine (S/T) protein kinase, plays a central role in
controlling cell growth, proliferation and metabolism (19,20).
Activation of the mTOR pathway was noted in melanoma (21), squamous cell cancers (22), adenocarcinomas (23), colorectal cancers (24) and lymphomas (25). Increasing lines of evidence suggest
that curcumin may exert its antiproliferative effects by inhibiting
mTOR signaling directly or indirectly and thus may represent a new
class of mTOR inhibitors (26).
The ability of curcumin to alter the redox status of
transformed cells (27) and its
desirable safety profile prompted us to investigate whether it may
also alter radiation sensitivity in NHL. In this study, we
investigated the mechanism of action for curcumin’s effect on IR
cytotoxic activity in NHL cells. Since NHL is often resistant to
IR-induced cell death induction, we specifically examined how
curcumin induces cell death. Therefore, we searched for molecular
mechanisms that were associated with susceptibility to cell death.
We were able to highlight a novel mechanism by which curcumin
caused G2/M cell cycle arrest in NHL cells at concentrations that
were very close to its plasma-achievable concentrations in humans.
In this study, curcumin was identified as a radiosensitizer in NHL
cells. Furthermore, its effects were mediated through the
inhibition of IR-induced mTOR-NF-κB activation.
Materials and methods
Reagents
Ataxia-telangiectasia-mutated (ATM)-kinase
inhibitor, KU55933, was purchased from Calbiochem (an affiliate of
Merck KGaA). Curcumin, RNase A, propidium iodide and rapamycin
(mTOR inhibitor) were purchased from Sigma-Aldrich (St. Louis, MO,
USA). Antibodies against p53 (sc-6243) and β-actin (C4, sc-47778)
were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA). Antibodies against phosphorylated and nonphosphorylated mTOR
(Ser2448), p65, p21Waf1, p53, phospho-ATM (Ser-1981), ATM and DNA
polymerase β-1 were purchased from Cell Signaling Technology, Inc.
(Danvers, MA, USA). Electrophoresis reagents, cDNA synthesis kits,
SYBR-Green Master Mix and iQ5 software were all obtained from
Bio-Rad (Hercules, CA, USA). Unless otherwise indicated, all other
chemicals used in this study were purchased from Sigma-Aldrich.
Cell lines and culture
Three human lymphoma cell lines (Namalwa, Ramos and
Raji) were purchased from the American Type Culture Collection. The
cells were maintained in Roswell Park Memorial Institute
(RPMI)-1640 medium (Sigma-Aldrich) supplemented with 10% fetal calf
serum (Invitrogen Life Technologies, Carlsbad, CA, USA), 100 U/ml
penicillin and 100 mg/ml streptomycin in a humidified incubator at
37°C in the presence of 95% air and 5% CO2.
Cell treatment
Curcumin (98% purity) was dissolved in dimethyl
sulfoxide (DMSO, final concentration 0.5%) to produce a 100 mmol/l
stock solution. IR was delivered in a Siemens Primus accelerator
(Hamburg, Germany) at 6 MV at room temperature (dose rate: 200
cGy/min). Control cells were not irradiated but they were removed
from the incubator and were then placed at the radiation site for
the same period of radiation as the irradiated cells. For combined
treatment with curcumin, rapamycin or KU55933 and IR treatment,
cells were kept at 37°C with the indicated agent for 4 h prior to
IR treatment.
Cell viability assay
Cell viability was determined with the tetrazolium
salt water-soluble tetrazolium salt assay (Roche Diagnostics GmbH,
Penzberg, Germany) as previously described (13). The results were expressed as a
percentage of cell viability for each concentration of curcumin
with respect to the controls.
Cell cycle analysis
The cell cycle distribution was determined by
staining with propidium iodide (PI) as previously described
(28). After treatment with
curcumin (2 and 10 μmol/l), 1×106 cells were harvested
and fixed in 70% ethanol at 4°C overnight. The cells were washed
twice in ice-cold PBS, were treated with RNase for 1 h and their
DNA was stained with PI solution (50 mg/ml) for 30 min in the dark.
The cell cycle distribution was determined using FACSAria II flow
cyto-meter (Becton Dickinson, San Diego, CA, USA) using Cell Quest
software.
Subcellular fractionation
Namalwa, Ramos and Raji cells were incubated with
various stimuli for the indicated times, and the total and nuclear
extracts were obtained as previously described (13). The protein content was measured by
the Bradford method (Bio-Rad, Hercules, CA, USA). Following
analysis of protein content, 20 μg of both fractions was subjected
to SDS-polyacrylamide gel electrophoresis and examined by western
blotting.
Western blot analysis
Equal amounts of lysate protein (20 μg) were
separated by SDS-polyacrylamide gel electrophoresis, transferred to
polyvinylidene difluoride membranes and blocked in 5% non-fat milk
in PBS for 1 h. The membranes were probed with the primary
antibodies mentioned above in the Reagents section. Membranes were
re-probed with anti-β-actin antibody at the concentration of
1:5,000. The antibodies were detected using a chemiluminescence
detection kit (LumiGLO; Cell Signaling Technology, Inc.) according
to the manufacturer’s instructions.
Statistical analysis
Data are presented as the means ± SEM. Comparison
between groups was performed with the paired Student’s t-test and
the level of statistical significance was determined to be
P<0.05.
Results
Effects of curcumin on cell viability in
human lymphoma cell lines
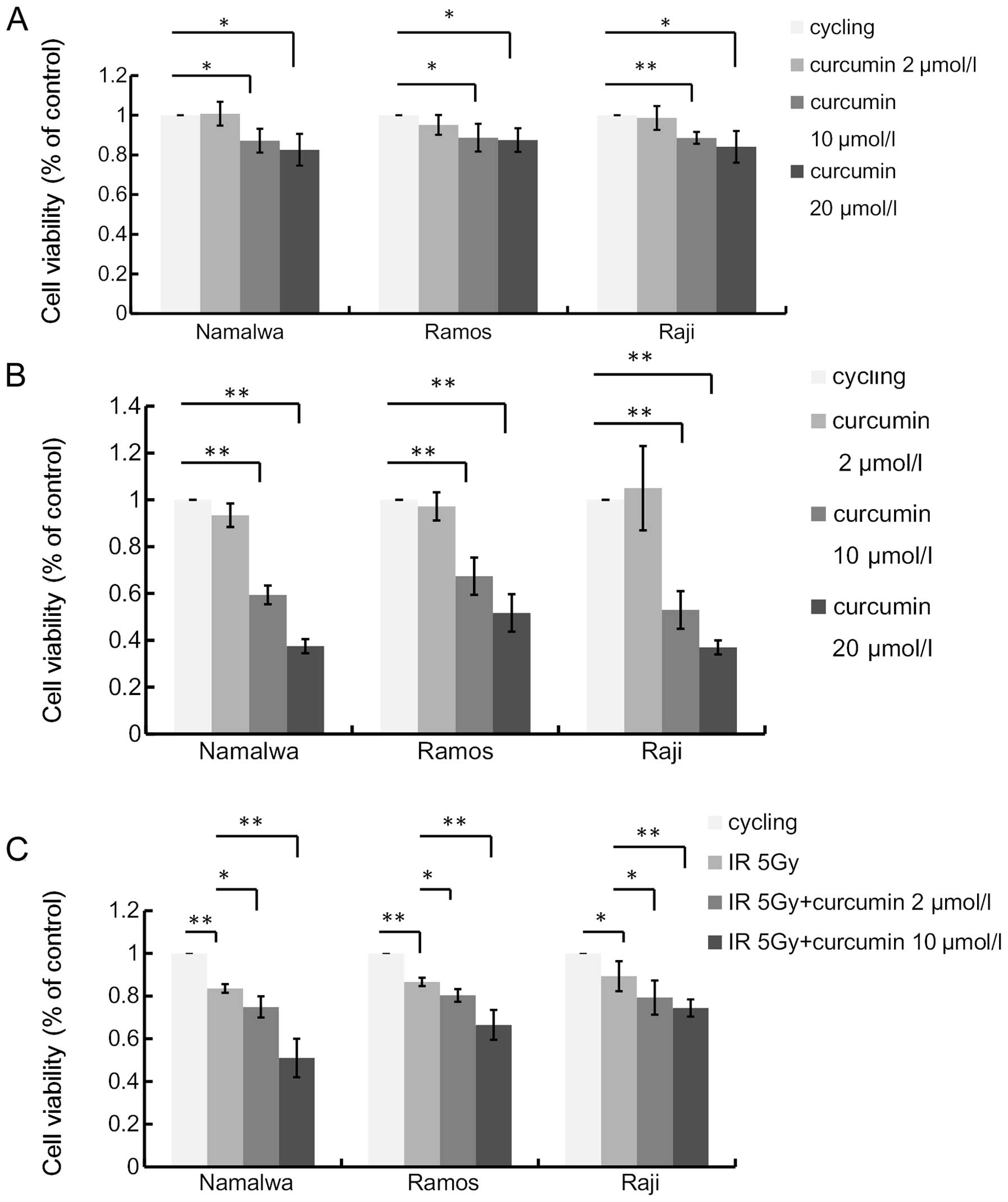
We assessed the antiproliferative effect of curcumin
on NHL cell lines. Namalwa, Ramos and Raji cells were treated with
2, 10 and 20 μmol/l of curcumin for 48 h. The viability of the
three lymphoma cell lines was decreased by curcumin treatment in a
dose-dependent manner (Fig. 1A).
However, a lower concentration (2 μmol/l) of curcumin did not
display an impact on the viability of NHL cells (P>0.05).
To examine whether this antiproliferative effect was
permanent or transient, we further assessed the capacity of the
cultures to recover from curcumin treatment. After incubation with
curcumin for 48 h, cells were further incubated for 48 h in the
absence of curcumin. Treatment with 10 and 20 μmol/l curcumin
markedly inhibited the recovery of all cell lines, indicating
substantial injury to DNA repair and replication (Fig. 1B).
Curcumin enhances the antiproliferative
effect of ionizing radiation
To examine the radiosensitization effect of curcumin
on human lymphoma cells, we further investigated the effects of
curcumin pre-treatment on radiation-induced cell death (Fig. 1C). The treatment with 5 Gy IR
resulted in growth inhibition. The percentage of cells that
survived at 48 h were 83.51±2.27, 86.68±2.13 and 89.30±7.21% in
Namalwa, Ramos and Raji cells, respectively. Curcumin significantly
augmented the IR-induced inhibition of survival in the three cell
lines. Although the low dose (2 μmol/l) of curcumin displayed no
cytotoxicity on NHL cells, it enhanced the IR-induced cell death.
Compared with IR alone, the combined treatment led to even less
cell survival down to 74.90±5.52, 80.39±3.28 and 79.30±7.43% in the
three NHL cell lines, respectively (P<0.05). In order to better
demonstrate the potential clinical benefit of curcumin in
combination with radiotherapy, we selected a curcumin dose of 2 and
10 μmol/l for 4 h as the pre-treatment condition used in further
molecular studies. Taken together, these results clearly
demonstrated that curcumin treatment sensitized these tumor cell
lines to IR.
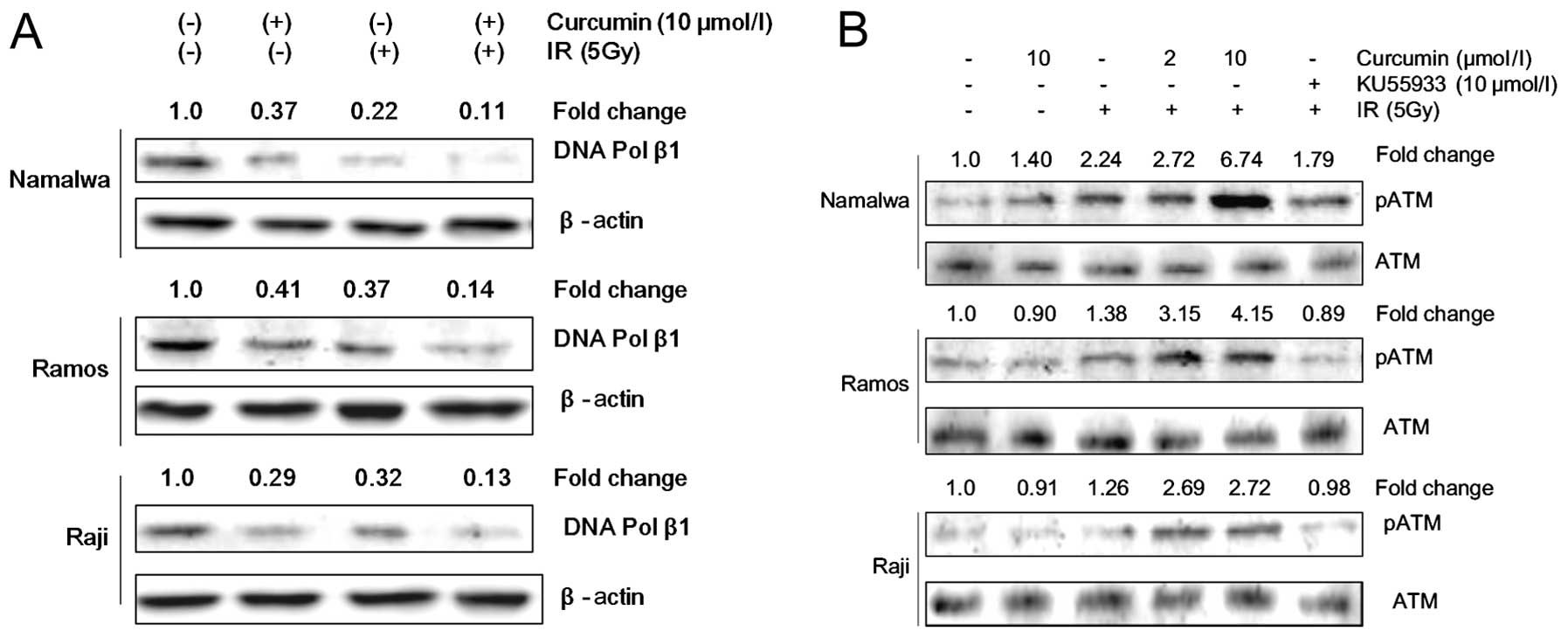
Curcumin induces IR-mediated DNA
damage
DNA polymerase β is crucial in the repair of DNA
strand breaks (29). To investigate
whether the DNA-damaging effect is involved in the sensitization of
the IR-induced antiproliferative effect by curcumin, we analyzed
the expression of molecules involved in the regulation of DNA
damage repair by western blot analysis. Our results demonstrated
that curcumin treatment decreased the expression of DNA polymerase
β and enhanced the IR-induced inhibition of DNA polymerase β
(Fig. 2A).
Considering that phosphorylation of ATM is an
indicator of the presence of DNA double-strand breaks (30), we further analyzed the
phosphorylation of ATM at Ser-1981. We observed an increased
expression of phosphorylated ATM by IR or curcumin in either of the
doses without any changes in the total ATM protein level (Fig. 2B). This effect was more prominent
when NHL cells were treated with a combination of curcumin and
IR.
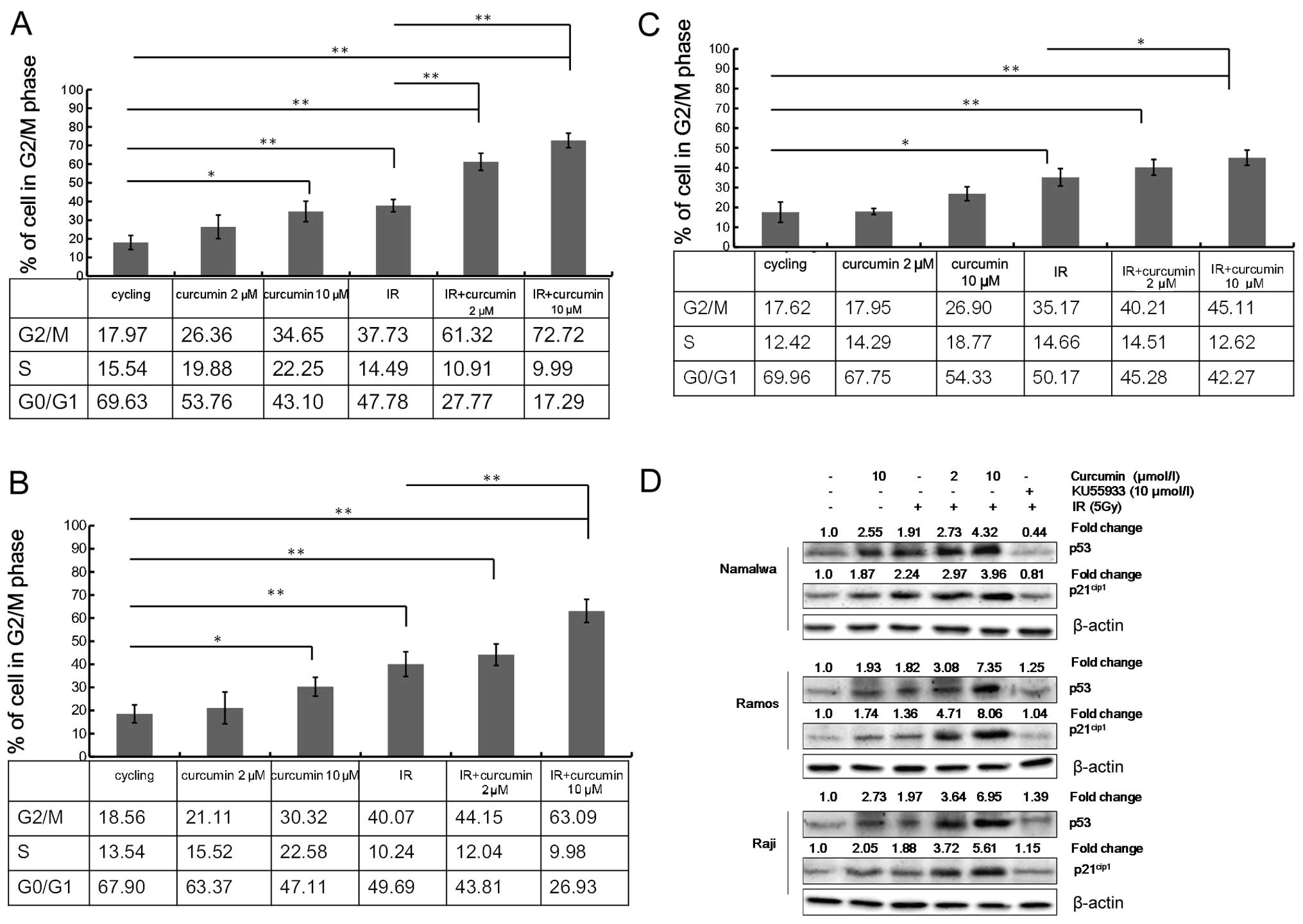
Curcumin enhances IR-induced G2/M arrest
in lymphoma cells
To gain further insight into curcumin’s mechanism of
action for the cell sensitization to IR-induced growth inhibition,
we analyzed the distribution of the cell cycle by flow cytometry.
Both curcumin (10 μmol/l) and IR treatment caused G2/M cell cycle
arrest and this effect was more prominent with the combined
treatment of curcumin and IR. A low concentration of curcumin (2
μmol/l) increased the percentage of lymphoma cells that arrested in
the G2/M phase due to the effect of IR. However, this low dose
alone (without IR) had little effect on G2/M phase arrest. Exposure
of 5 Gy of IR induced G2/M phase arrest in 37.73±3.31, 40.07±3.32
and 35.17±4.38% of Namalwa, Ramos and Raji cells, respectively.
However, the pre-treatment of cells with 2 μmol/l of curcumin
followed by IR led to a significant increase in G2/M phase arrest
in Namalwa, Ramos and Raji cells (61.32±4.59, 44.15±4.65 and
40.21±3.97%, respectively). Meanwhile, the proportion of cells in
the G0/G1 phase was concomitantly decreased following the combined
treatment of IR and curcumin (2 μmol/l) (Fig. 3A–C).
Curcumin enhances the IR induction of
cell cycle regulatory proteins
DNA damage generally leads to the activation of the
ATM pathway (31,32). To further delineate the molecular
mechanism of curcumin on enhanced IR-induced G2/M arrest, western
blot analysis was used to determine the effect of curcumin on
selected proteins that are involved in cell cycle regulation.
Treatment of the cells with 10 μmol/l curcumin or IR displayed an
increase in the expression of cyclin-dependent kinase inhibitors
p21cip1 and p53 as compared to the DMSO-treated control
cells (Fig. 3D). The treatment with
combined curcumin and IR further increased the expression of both
the p53 and p21cip1 proteins as compared to the
treatment with either of them alone.
We aimed to explore whether enhanced phosphorylation
of ATM plays a role in the effect of curcumin on IR-induced cell
cycle redistribution. NHL cells were incubated with an ATM-specific
inhibitor, KU55933, (33) and were
further subjected to curcumin treatment for 4 h followed by IR
exposure. Pre-incubation with 10 μmol/l KU55933 prevented
IR-induced ATM phosphorylation but also suppressed
curcumin-enhanced activation of p53 and p21cip1 by IR
(Fig. 3D). Previous studies have
demonstrated that pre-treatment of cells with 20 μmol/l of curcumin
for 4 h significantly suppressed the IR-induced overexpression of
CDC2 and cyclin B1 in all three cell lines (13) These results provide further evidence
for the possible involvement of ATM/p53/p21cip1 in the
G2/M cell cycle arrest caused by curcumin.
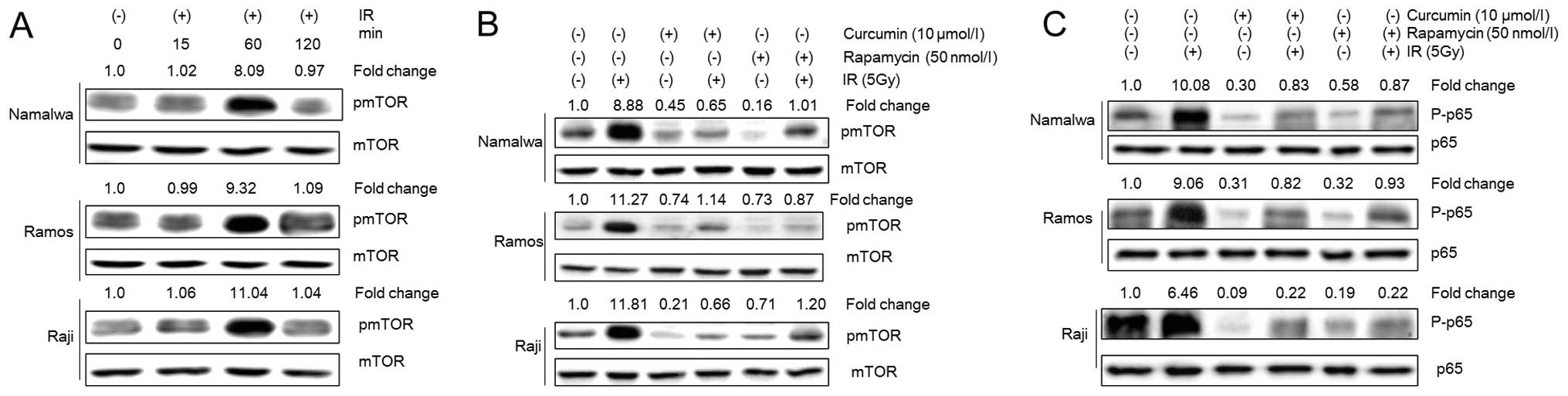
Curcumin inhibits the IR-induced
mTOR-NF-κB pathway activation
To investigate the effect of curcumin on mTOR
activity in lymphoma cells, the regulation of mTOR phosphorylation
by curcumin was examined. The treatment with 10 μmol/l of curcumin
led to decreased phosphorylation of mTOR. Likewise, IR exposure
induced mTOR phosphorylation in NHL cells, which reached the
maximal level approximately 1 h after the irradiation (Fig. 4A). However, pre-treatment with
curcumin (10 μmol/l) inhibited the mTOR phosphorylation that was
induced by IR (Fig. 4B).
A 4-h treatment of NHL cells with rapamycin, a
specific inhibitor of mTOR, markedly inhibited mTOR phosphorylation
in NHL cells. However, when NHL cells were pretreated with
rapamycin followed by IR, the IR-induced mTOR phosphorylation was
suppressed (Fig. 4B).
We previously demonstrated that IR induced p65
phosphorylation in NHL cells 4 h after IR (13) and curcumin pre-treatment inhibited
its nuclear translocation. However, the mechanism through which
curcumin regulates the activation of NF-κB is not clear. Previous
studies have demonstrated that mTOR inhibition led to a reduced
expression of P65 in various cancer cell types (34). In this study, it was demonstrated
that the treatment of NHL cells with rapamycin inhibited
constitutively the expression of p65 and suppressed the IR-induced
p65 phosphorylation. This may be indicative of the possibility that
mTOR regulates the activation of NF-κB in NHL cells (Fig. 4C). Taken together, the results of
this study suggest that the regulation of NF-κB by curcumin in
combination with IR is mediated by mTOR.
Discussion
In the present study, it was demonstrated that
curcumin exerted cytotoxicity on NHL cells at high concentrations.
Furthermore, pre-treatment of these cells with curcumin enhanced
the cell response to ionizing radiation (IR) even at lower
concentrations of curcumin, where curcumin alone failed to display
any direct cytotoxicity in lymphoma cells. We also investigated the
mechanism of action for the cell cycle arrest involved in the
effect of curcumin treatment that is mediated through the DNA
damage pathway. Curcumin was able to increase the G2/M cell cycle
arrest induced by IR. This effect was probably mediated though the
activation of the cell cycle regulators p53 and p21 that was in
turn dependent on the increased ATM phosphorylation indicating the
presence of DNA double-strand breaks. IR induced mTOR
phosphorylation in NHL cells within 1 h after IR exposure and
pre-treatment with curcumin (10 μmol/l) inhibited this effect.
It has been well documented that curcumin possesses
antitumor activity through the inhibition of proliferation,
invasion, angiogenesis and induction of apoptosis and cell cycle
redistribution (35). However,
susceptibility to curcumin varies among different cell lines,
indicating that it has either multiple targets that are expressed
in a cell-specific manner or one target that affects cell-specific
pathways. We reported that the cytotoxic effects of curcumin and IR
in NHL may be mediated through the DNA damage pathway, evidenced by
a reduced expression of DNA polymerase β and an increased ATM
expression. Our results were consistent with previous reports, in
which curcumin caused DNA damage in human pancreatic cancer through
the increased phosphorylation of H2A.X and Chk1 and reduced
expression of DNA polymerase β (36).
We next investigated the mechanism of action for
curcumin and IR in the induction of G2/M phase arrest in NHL cells
that was mediated through the DNA damage pathway. Cells are blocked
in the G2/M phase during DNA damage (32). The critical target of p53 at G2/M is
the Cdk inhibitor p21, which causes the dissociation of the cdc2
and cyclin complex (37).
Consistent with previously published reports (36,38,39),
the present study revealed that pre-treatment with curcumin
enhanced IR-induced cell arrest in the G2/M phase and concomitantly
decreased the percentage of cells in G0/G1 phases in lymphoma. In
the G2/M phase, cells are more susceptible to the cytotoxic effects
of radiotherapy (40,41). At the same time, curcumin enhanced
IR-induced expression of p53 and p21. The IR-induced expression of
p53 and CDK inhibitor p21cip1 as a result of cell
treatment with curcumin may thus provide a potential molecular
mechanism of action for the induction of cell arrest in G2/M
phase.
DNA damage checkpoints are predominantly associated
with the activation of ATM, which plays an important role in DNA
repair (32). In previous studies,
treatment of non-small cell lung cancer A549 cells and pancreatic
cancer cells with curcumin, induced cell cycle arrest through
activation of ATM by its phosphorylation (32). Our results further demonstrated the
ATM phosphorylation at Ser-1981 as a result of curcumin treatment
and IR exposure. To confirm the involvement of ATM phosphorylation
in the IR-induced G2/M phase arrest as a result of curcumin
treatment, NHL cells were pre-incubated with a specific ATM
inhibitor KU55933 followed by IR exposure. Utilization of KU55933
markedly abrogated IR-induced ATM phosphorylation as well as the
expression of p53 and p21cip1. These results suggest
that the phosphorylation of ATM plays a crucial role in G2/M phase
arrest induced by IR and curcumin through the regulation of p53 and
p21cip1 expression.
We previously demonstrated that curcumin sensitizes
NHL cells to IR-induced apoptosis through decreasing the NF-κB
expression and by inhibiting its nuclear localization induced by IR
exposure (13). However, the
details of the mechanism by which curcumin regulates IR-induced
activation of NF-κB remains unclear. As previously suggested, the
activation of mTOR cascade has been hypothesized to increase the
radiation resistance in tumors. Therefore, the mTOR cascade is
triggered as a cellular defense mechanism in response to IR
exposure to prevent radiation-induced cell death. It has been
reported that an mTOR inhibitor was able to sensitize numerous
cancer types to DNA damaging agents including IR (42). In addition, mTOR has been reported
to be a direct target of curcumin in various tumor cell lines
(26). Curcumin inhibited the
phosphorylation of mTOR and its downstream targets in numerous
cancer cell lines. This further led to the inhibition of cell
growth, cell cycle progression and cell proliferation (43,44).
Thus, it is reasonable to hypothesize that mTOR may mediate the
radiosensitization effect of curcumin in NHL through the regulation
of NF-κB activation. The treatment of NHL cells with curcumin in
this study, significantly inhibited the constitutively expressed
mTOR and NF-κB proteins. However, the pre-treatment of NHL cells
with curcumin followed by 5 Gy of IR treatment suppressed
IR-induced mTOR phosphorylation and p65 nuclear translocation. In
addition, rapamycin treatment inhibited the constitutive expression
of p65 and suppressed its IR-induced phosphorylation. Taken
together, these results strongly suggest that the treatment of NHL
with curcumin results in a radiosensitization effect that is
mediated through the inhibition of the mTOR-NF-κB pathway. This may
be the underlying mechanism of action for curcumin’s effect on
IR-induced NHL cell death.
In conclusion, our results indicate that curcumin
may enhance the NHL cell response to IR through the modulation of
the G2/M phase arrest and the inhibition of the mTOR pathway.
Acknowledgements
This study was supported by funding from the
Department of Health of Liaoning Province provided to Q.Q.
(2009A754).
References
|
1
|
Bulut E, Bekcioglu B, Gunhan O and Sener
I: Diffuse large B-cell lymphoma with oral manifestations. J
Craniofac Surg. 22:1144–1147. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Hainsworth JD: Monoclonal antibody therapy
in lymphoid malignancies. Oncologist. 5:376–384. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li L, Aggarwal BB, Shishodia S, Abbruzzese
J and Kurzrock R: Nuclear factor-kappaB and IkappaB kinase are
constitutively active in human pancreatic cells, and their
down-regulation by curcumin (diferuloylmethane) is associated with
the suppression of proliferation and the induction of apoptosis.
Cancer. 101:2351–2362. 2004. View Article : Google Scholar
|
|
4
|
Aggarwal BB, Banerjee S, Bharadwaj U, Sung
B, Shishodia S and Sethi G: Curcumin induces the degradation of
cyclin E expression through ubiquitin-dependent pathway and
up-regulates cyclin-dependent kinase inhibitors p21 and p27 in
multiple human tumor cell lines. Biochem Pharmacol. 73:1024–1032.
2007. View Article : Google Scholar
|
|
5
|
Liu HL, Chen Y, Cui GH and Zhou JF:
Curcumin, a potent anti-tumor reagent, is a novel histone
deacetylase inhibitor regulating B-NHL cell line Raji
proliferation. Acta Pharmacol Sin. 26:603–609. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Alaikov T, Konstantinov SM, Tzanova T,
Dinev K, Topashka-Ancheva M and Berger MR: Antineoplastic and
anticlastogenic properties of curcumin. Ann NY Acad Sci.
1095:355–370. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Syng-Ai C, Kumari AL and Khar A: Effect of
curcumin on normal and tumor cells: role of glutathione and bcl-2.
Mol Cancer Ther. 3:1101–1108. 2004.PubMed/NCBI
|
|
8
|
Khafif A, Lev-Ari S, Vexler A, et al:
Curcumin: a potential radio-enhancer in head and neck cancer.
Laryngoscope. 119:2019–2026. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chendil D, Ranga RS, Meigooni D,
Sathishkumar S and Ahmed MM: Curcumin confers radiosensitizing
effect in prostate cancer cell line PC-3. Oncogene. 23:1599–1607.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sandur SK, Deorukhkar A, Pandey MK, et al:
Curcumin modulates the radiosensitivity of colorectal cancer cells
by suppressing constitutive and inducible NF-kappaB activity. Int J
Radiat Oncol Biol Phys. 75:534–542. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yallapu MM, Maher DM, Sundram V, Bell MC,
Jaggi M and Chauhan SC: Curcumin induces chemo/radio-sensitization
in ovarian cancer cells and curcumin nanoparticles inhibit ovarian
cancer cell growth. J Ovarian Res. 3:112010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Javvadi P, Segan AT, Tuttle SW and
Koumenis C: The chemopreventive agent curcumin is a potent
radiosensitizer of human cervical tumor cells via increased
reactive oxygen species production and overactivation of the
mitogen-activated protein kinase pathway. Mol Pharmacol.
73:1491–1501. 2008. View Article : Google Scholar
|
|
13
|
Qiao Q, Jiang Y and Li G: Curcumin
improves the antitumor effect of X-ray irradiation by blocking the
NF-kappaB pathway: an in vitro study of lymphoma. Anticancer Drugs.
23:597–605. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Anand P, Sung B, Kunnumakkara AB,
Rajasekharan KN and Aggarwal BB: Suppression of pro-inflammatory
and proliferative pathways by diferuloylmethane (curcumin) and its
analogues dibenzoylmethane, dibenzoylpropane, and
dibenzylideneacetone: role of Michael acceptors and Michael donors.
Biochem Pharmacol. 82:1901–1909. 2011. View Article : Google Scholar
|
|
15
|
Wang Y, Rishi AK, Wu W, et al: Curcumin
suppresses growth of mesothelioma cells in vitro and in vivo, in
part, by stimulating apoptosis. Mol Cell Biochem. 357:83–94. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Lev-Ari S, Starr A, Vexler A, et al:
Inhibition of pancreatic and lung adenocarcinoma cell survival by
curcumin is associated with increased apoptosis, down-regulation of
COX-2 and EGFR and inhibition of Erk1/2 activity. Anticancer Res.
26:4423–4430. 2006.PubMed/NCBI
|
|
17
|
Liao S, Xia J, Chen Z, et al: Inhibitory
effect of curcumin on oral carcinoma CAL-27 cells via suppression
of Notch-1 and NF-kappaB signaling pathways. J Cell Biochem.
112:1055–1065. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Binion DG, Otterson MF and Rafiee P:
Curcumin inhibits VEGF-mediated angiogenesis in human intestinal
microvascular endothelial cells through COX-2 and MAPK inhibition.
Gut. 57:1509–1517. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Huang S and Houghton PJ: Targeting mTOR
signaling for cancer therapy. Curr Opin Pharmacol. 3:371–377. 2003.
View Article : Google Scholar
|
|
20
|
Li R, Wang R, Zhai R and Dong Z: Targeted
inhibition of mammalian target of rapamycin (mTOR) signaling
pathway inhibits proliferation and induces apoptosis of laryngeal
carcinoma cells in vitro. Tumori. 97:781–786. 2011.PubMed/NCBI
|
|
21
|
Jazirehi AR, Wenn PB and Damavand M:
Therapeutic implications of targeting the PI3Kinase/AKT/mTOR
signaling module in melanoma therapy. Am J Cancer Res. 2:178–191.
2012.PubMed/NCBI
|
|
22
|
Chakraborty S, Mohiyuddin SM, Gopinath KS
and Kumar A: Involvement of TSC genes and differential expression
of other members of the mTOR signaling pathway in oral squamous
cell carcinoma. BMC Cancer. 8:1632008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Darb-Esfahani S, Faggad A, Noske A, et al:
Phospho-mTOR and phospho-4EBP1 in endometrial adenocarcinoma:
association with stage and grade in vivo and link with response to
rapamycin treatment in vitro. J Cancer Res Clin Oncol. 135:933–941.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ekstrand AI, Jonsson M, Lindblom A, Borg A
and Nilbert M: Frequent alterations of the PI3K/AKT/mTOR pathways
in hereditary nonpolyposis colorectal cancer. Fam Cancer.
9:125–129. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kuo SH, Hsu CH, Chen LT, et al: Lack of
compensatory pAKT activation and eIF4E phosphorylation of lymphoma
cells towards mTOR inhibitor, RAD001. Eur J Cancer. 47:1244–1257.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Beevers CS, Chen L, Liu L, Luo Y, Webster
NJ and Huang S: Curcumin disrupts the mammalian target of
rapamycin-raptor complex. Cancer Res. 69:1000–1008. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Scapagnini G, Foresti R, Calabrese V,
Giuffrida Stella AM, Green CJ and Motterlini R: Caffeic acid
phenethyl ester and curcumin: a novel class of heme oxygenase-1
inducers. Mol Pharmacol. 61:554–561. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kumar R and Atlas I: Interferon alpha
induces the expression of retinoblastoma gene product in human
Burkitt lymphoma Daudi cells: role in growth regulation. Proc Natl
Acad Sci USA. 89:6599–6603. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Neijenhuis S, Verwijs-Janssen M, van den
Broek LJ, Begg AC and Vens C: Targeted radiosensitization of cells
expressing truncated DNA polymerase β. Cancer Res. 70:8706–8714.
2010.PubMed/NCBI
|
|
30
|
Shrivastav M, Miller CA, De Haro LP, et
al: DNA-PKcs and ATM co-regulate DNA double-strand break repair.
DNA Repair (Amst). 8:920–929. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lukas C, Bartkova J, Latella L, et al: DNA
damage-activated kinase Chk2 is independent of proliferation or
differentiation yet correlates with tissue biology. Cancer Res.
61:4990–4993. 2001.PubMed/NCBI
|
|
32
|
Huang M, Miao ZH, Zhu H, Cai YJ, Lu W and
Ding J: Chk1 and Chk2 are differentially involved in homologous
recombination repair and cell cycle arrest in response to DNA
double-strand breaks induced by camptothecins. Mol Cancer Ther.
7:1440–1449. 2008. View Article : Google Scholar
|
|
33
|
Hickson I, Zhao Y, Richardson CJ, et al:
Identification and characterization of a novel and specific
inhibitor of the ataxia-telangiectasia mutated kinase ATM. Cancer
Res. 64:9152–9159. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Rashid A, Liu C, Sanli T, et al:
Resveratrol enhances prostate cancer cell response to ionizing
radiation. Modulation of the AMPK, Akt and mTOR pathways. Radiat
Oncol. 6:1442011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Kunnumakkara AB, Anand P and Aggarwal BB:
Curcumin inhibits proliferation, invasion, angiogenesis and
metastasis of different cancers through interaction with multiple
cell signaling proteins. Cancer Lett. 269:199–225. 2008. View Article : Google Scholar
|
|
36
|
Sahu RP, Batra S and Srivastava SK:
Activation of ATM/Chk1 by curcumin causes cell cycle arrest and
apoptosis in human pancreatic cancer cells. Br J Cancer.
100:1425–1433. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Taylor WR and Stark GR: Regulation of the
G2/M transition by p53. Oncogene. 20:1803–1815. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Calaf GM, Echiburu-Chau C, Wen G, Balajee
AS and Roy D: Effect of curcumin on irradiated and
estrogen-transformed human breast cell lines. Int J Oncol.
40:436–442. 2012.PubMed/NCBI
|
|
39
|
Lin YC, Chen HW, Kuo YC, Chang YF, Lee YJ
and Hwang JJ: Therapeutic efficacy evaluation of curcumin on human
oral squamous cell carcinoma xenograft using multimodalities of
molecular imaging. Am J Chin Med. 38:343–358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Moragoda L, Jaszewski R and Majumdar AP:
Curcumin induced modulation of cell cycle and apoptosis in gastric
and colon cancer cells. Anticancer Res. 21:873–878. 2001.PubMed/NCBI
|
|
41
|
Liu E, Wu J, Cao W, et al: Curcumin
induces G2/M cell cycle arrest in a p53-dependent manner and
upregulates ING4 expression in human glioma. J Neurooncol.
85:263–270. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Schiewer MJ, Den R, Hoang DT, et al: mTOR
is a selective effector of the radiation therapy response in
androgen receptor-positive prostate cancer. Endocr Relat Cancer.
19:1–12. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Johnson SM, Gulhati P, Arrieta I, et al:
Curcumin inhibits proliferation of colorectal carcinoma by
modulating Akt/mTOR signaling. Anticancer Res. 29:3185–3190.
2009.PubMed/NCBI
|
|
44
|
Beevers CS, Li F, Liu L and Huang S:
Curcumin inhibits the mammalian target of rapamycin-mediated
signaling pathways in cancer cells. Int J Cancer. 119:757–764.
2006. View Article : Google Scholar : PubMed/NCBI
|