Introduction
Osteosarcoma is a primary malignant tumor of the
skeleton characterized by the direct formation of immature bone or
osteoid tissue by the tumor cells. It is an extremely aggressive
malignancy that arises mostly in the long bones (1). Limited improvements have been made by
using conventional methods including surgery, radiotherapy and
chemotherapy in the past two decades (2). Despite the availability of a myriad of
treatment modalities, including preferred cytotoxic chemotherapy,
dose-limiting toxicity to normal tissues and acquisition of
acquired resistance fails to transcend into optimal clinical
benefit in terms of cure rate in an overwhelming majority of
patients (3,4). Therefore, there is a need for novel
strategies involving less toxic agents for their potential in
improving the prognosis and therapy of patients with
osteosarcoma.
Thymoquinone (TQ) is the bioactive compound derived
from black seed (Nigella sativa) oil. It is an annual herb
that grows in countries bordering the Mediterranean region and in
Western Asian countries including India, Pakistan, and Afghanistan.
In traditional medicine, thymoquinone is known to be the active
principle responsible for many of the seed’s anti-oxidant and
anti-inflammatory effects (5).
Numerous studies have shown that the seeds and oil of this plant
are characterized by a very low degree of toxicity (6). Regarding cancer, recent studies showed
that thymoquinone exerts anti-proliferative and apoptosis-inducing
effects on various tumor cells derived from colorectal carcinoma
(7), lung carcinoma (8), myeloblastic leukemia (9) and prostate carcinoma (10). Thymoquinone has also been shown to
potentiate the antitumor activity of gemcitabine and oxaliplatin in
pancreatic cancer (11).
Mechanistically, thymoquinone has been reported to induce apoptosis
in tumor cells by suppressing NF-κB, Akt activation, and
extracellular signal-regulated kinase signaling pathways and to
also inhibit tumor angiogenesis (11–14).
Although Roepke et al reported that thymoquinone showed
inhibitory effects on human osteosarcoma cells (15), whether thymoquinone inhibits
osteosarcoma in vivo and suppresses osteosarcoma growth
through tumor angiogenesis prevention remains unclear.
In the present study, we first sought to understand
the molecular mechanism of action of thymoquinone in osteosarcoma
cells inducing apoptosis and we tested our hypothesis in
vivo using an orthotropic model of osteosarcoma. Using in
vivo data, we show, for the first time, that thymoquinone
exerts antitumor and anti-angiogenesis activity in osteosarcoma.
These results are correlated with the downregulation of NF-κB and
its downstream proteins such as X-linked inhibitor of apoptosis
(XIAP), survivin and vascular endothelial growth factor (VEGF) in
tumor extracts.
Materials and methods
Reagents
Antibodies were obtained from the following
commercial sources: antibodies against survivin, XIAP, Smac and
β-actin were obtained from Epitomics (Burlingame, CA, USA), and the
caspase-3 antibody was from Abcam (Cambridge, MA, USA).
Anti-retinoblastoma antibody was from Santa Cruz Biotechnology
(Santa Cruz, CA, USA). Thymoquinone was purchased from Sigma (St.
Louis, MO, USA) and was dissolved in DMSO to make 20 mmol/l stock
solution. The 0.1% DMSO alone was set as the control group.
Cell culture
Human osteosarcoma cell line SaOS-2 was purchased
from American Type Culture Collection (ATCC, Rockville, MD, USA).
Mouse osteoblastic cell line MC3T3-El was preserved in our
laboratory. SaOS-2 and MC3T3-El cells were maintained in modified
Eagle’s medium containing 10% fetal bovine serum (FBS), 0.5%
penicillin-streptomycin, and 1% glutamine at 37°C with 5%
CO2. Human umbilical vein endothelial cells (HUVECs;
obtained from ATCC) were cultured in gelatin-coated plates with
M199 medium containing 20% FBS, endothelial cell growth supplement
(50 μg/ml, Sigma) and antibiotics, and incubated at 37°C in 5%
CO2 in air.
Hoechst 33342 staining for apoptotic
nuclei
Morphological changes of SaOS-2 cells were observed
under a fluorescence microscope (Olympus, Tokyo, Japan) by the
Hoechst staining method. SaOS-2 cells were seeded at a density of
2×105 cells per well onto a 12-well plate for 24 h,
followed by incubation with vehicle alone (0.1% DMSO) or 80 μmol/l
thymoquinone for 24 h. Following treatment, cells were fixed with
3.7% formaldehyde for 15 min, permeabilized with 0.1% Triton X-100
and stained with 5 mg/ml of Hoechst 33258 for another 5 min at
37°C. The cells were then washed with PBS and observed under a
fluorescence microscope.
Cell viability inhibition by
thymoquinone
SaOS-2 cells were seeded at a density of
3×103 cells per well in 96-well culture plates. After
overnight incubation, the medium was removed and replaced with
fresh medium containing different concentrations of thymoquinone
(20, 40 and 80 μmol/l). Following a 24-h incubation, cell viability
was determined by CCK-8 assay (Dojin Laboratory, Kumamoto, Japan)
according to the manufacturer’s instructions. Briefly, CCK-8
solution was added to cells in 96-well plates, the cells were then
incubated at 37°C for 60 min, and absorbance was measured at 570 nm
using an MRX Revelation 96-well multiscanner (Dynex Technologies,
Chantilly, VA, USA). This experiment was repeated three times.
Flow cytometric assessment of
apoptosis
The measurement of phosphatidylserine redistribution
in a plasma membrane was conducted according to the protocol
outlined by the manufacturer of the Annexin V-FITC/PI apoptosis
detection kit (Abcam). After exposing the cells to increasing
concentrations of thymoquinone (20, 40 and 80 μmol/l) for 24 h at
6-well plates, harvested 1×105 cells were incubated with
5 μl of Annexin V-FITC and 5 μl of PI for 15 min at room
temperature in the dark and then analyzed by flow cytometry using
the Cell Quest program (Becton-Dickinson, San Jose, CA, USA).
Tube formation assay
The angiogenesis of HUVEC induced by Matrigel was
assessed by the modified methods previously described (12). Briefly, Matrigel (Becton-Dickinson)
was dissolved at 4°C overnight, and each well of pre-chilled
48-well plates was coated with 100 μl Matrigel and incubated at
37°C for 45 min. HUVECs (2×104 cells) were seeded onto
the Matrigel in 250 μl M199 supplemented with 20% FBS and incubated
with various concentrations of thymoquinone (40, 80 and 160 nmol/l)
at 37°C for 24 h in a humidified 5% CO2 atmosphere.
Endothelial cell tube formation was photographed and the light
micrograph images were stored in a computer. Tubular structures
were quantified by manual counting and percent inhibition was
expressed using untreated wells as 100%.
Western blot analysis
At the end of this incubation, cells were washed
with ice-cold PBS and lysed in NP40 lysis buffer (20 mmol/l
Tris-HCl (pH 7.4), 100 mmol/l NaCl, 1% NP40, 0.5% sodium
deoxycholate, 5 mmol/l MgCl2, 0.1 mmol/l
phenylmethylsulfonyl fluoride, and 10 mg/ml of protease inhibitor
mixture). Protein was extracted using Mammalian Protein Extraction
Reagent (Pierce Inc., Rockford, IL, USA) and its concentration was
determined by BCA (Pierce) assay. Proteins (30 μg) were separated
in 10–15% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to a polyphorylated difluoride (PVDF) membrane.
Membranes were incubated with primary antibody overnight at 4°C and
then with the respective secondary antibodies. Immunoreactive bands
were detected by the enhanced chemiluminescence (ECL) kit for
western blotting detection with hyper-ECL film. The same membrane
was reprobed with the anti-β-actin antibody, which was used as an
internal control for protein loading.
Electrophoretic mobility shift assay
Following treatment, the cells were suspended in 400
μl of ice-cold lysis buffer [1 mol/l HEPES (pH 7.9), 1 mol/l KCl,
0.5 mol/l EDTA, 0.1 mol/l EGTA, 0.1 mol/l DTT, 0.1 mol/l PMSF, 2
μg/ml aprotinin, 2 μg/ml leupeptin, and 0.5 mg/ml benzamidine] for
15 min. The cells were allowed to swell on ice for 20 min and then
4.8 μl of 10% Nonidet P-40 was added to every 400 μl cell
suspension, vortexed, and centrifuged for 1 min at 4°C. The nuclear
pellet was resuspended in 30 μl nuclear extraction buffer [2 mol/l
HEPES (pH 7.9), 0.4 mol/l NaCl, 1 mol/l EDTA, 0.1 mol/l EGTA, 0.1
mol/l DTT, 0.1 mol/l PMSF, 2 μg/ml aprotinin, 2 μg/ml leupeptin,
and 0.5 mg/ml benzamidine] and incubated on ice with intermittent
mixing. The tubes were then centrifuged at 10,000 g for 20 min at
4°C, and the supernatant (nuclear extract) was quantified using the
BCA protein assay. Electrophoretic mobility shift assay (EMSA) was
performed by incubating 10 μg of nuclear proteins with IRDye™-700
labeled NF-κB oligonucleotide. The incubation mixture included 2 μg
of poly(deoxyinosinic-deoxycytidylic acid) in a binding buffer. The
DNA-protein complex formed was separated from free oligonucleotide
on 8.0% native polyacrylamide gel using buffer containing 50 mmol/l
Tris, 200 mmol/l glycine (pH 8.5), and 1 mmol/l EDTA and then
visualized by Imager apparatus. Equal protein loading was ensured
by immunoblotting 10 μg of nuclear protein with anti-Rb
antibody.
Mouse osteosarcoma models and
experimental protocol
Male athymic BALB/c nu/nu mice (4–6 weeks old) were
obtained from Wenzhou Medical College. All animals were maintained
in the standard mouse plexiglass cages in a room maintained at
constant temperature and humidity under 12-h light and darkness
cycle. The food, water, and bedding for these immunocompromised
mice were sterilized and changed at least once weekly.
The spontaneously metastatic mouse model was
developed as previously described (16). Briefly, the left tibia was wiped
with 70% ethanol and a 27-gauge needle coupled to a Hamilton
syringe was inserted through the tibial plateau with the knee
flexed, and 1×105 SaOS-2 cells, resuspended in 10 μl
PBS, were injected into the marrow space of the proximal tibia. As
a control, all animals were injected with controled 0.1% DMSO in
the contralateral tibia. After 1 week of implantation, mice were
randomized into 2 groups (n=6): a) vehicle alone (control); b)
thymoquinone (6 mg/kg given daily by intragastric intubation for 15
days) (6). Serial primary tumor
volumes were excised and the final tumor volume was measured using
the formula: π × (d/2)3, where d is the diameter of the
tumor. Mice were sacrificed 5 days after the last treatment. Half
of the tumor tissue was formalin fixed and paraffin embedded for
immunohistochemistry and routine H&E staining. The other half
was snap-frozen in liquid nitrogen and stored at −80°C. H&E
staining confirmed the presence of tumor(s) in each tissue.
Immunohistochemical analysis
Xenograft tissues of all mice from both the control
and the treated groups were harvested at the end of the treatment
and fixed with formaldehyde. The fixed tissue was sectioned and
immunostaining was performed using primary antibodies specific for
Ki-67, CD34 and NF-κB with appropriate dilutions and using normal
host serum for negative controls, followed by staining with
appropriate HRP-conjugated secondary antibodies. Results were
expressed as percentage of Ki-67+ cells ± SE per ×40
magnification. A total of ten ×40 fields were examined and counted
from three tumors of each of the treatment groups. Areas of greater
vessel density were then examined under higher magnification (x100)
and counted. Any distinct area of positive staining for CD34 was
counted as a single vessel. Results were expressed as the mean
number of vessels ± SE per high-power field (x100). A total of 20
high-power fields were examined and counted from three tumors of
each of the treatment groups. The slides were developed in
diaminobenzidine and counterstained with a weak solution of
haematoxylin solution stain. H&E was carried out on
paraffin-embedded tissue sections. The stained slides were
dehydrated and mounted in permount and visualized on an Olympus
microscope (Olympus, Japan). Images were captured with an attached
camera linked to a computer.
Statistical analysis
Three independent experiments were performed, and
data are represented as the mean ± SD for the absolute values or
percent of controls. SPSS15.0 software was used for statistical
analysis. Statistically significant differences between values
obtained under different experimental conditions were determined
using the 2-tailed unpaired Student’s t-test or χ2 test.
P<0.05 was considered to indicate statistically significant
differences.
Results
Effect of thymoquinone on cell
viability
In order to investigate whether thymoquinone exerts
anti-viability effects, SaOS-2 cells were treated with increasing
concentrations of thymoquinone (20, 40 and 80 μmol/l) for 24 h, and
then the viability of the cells was examined by CCK-8 assays. As
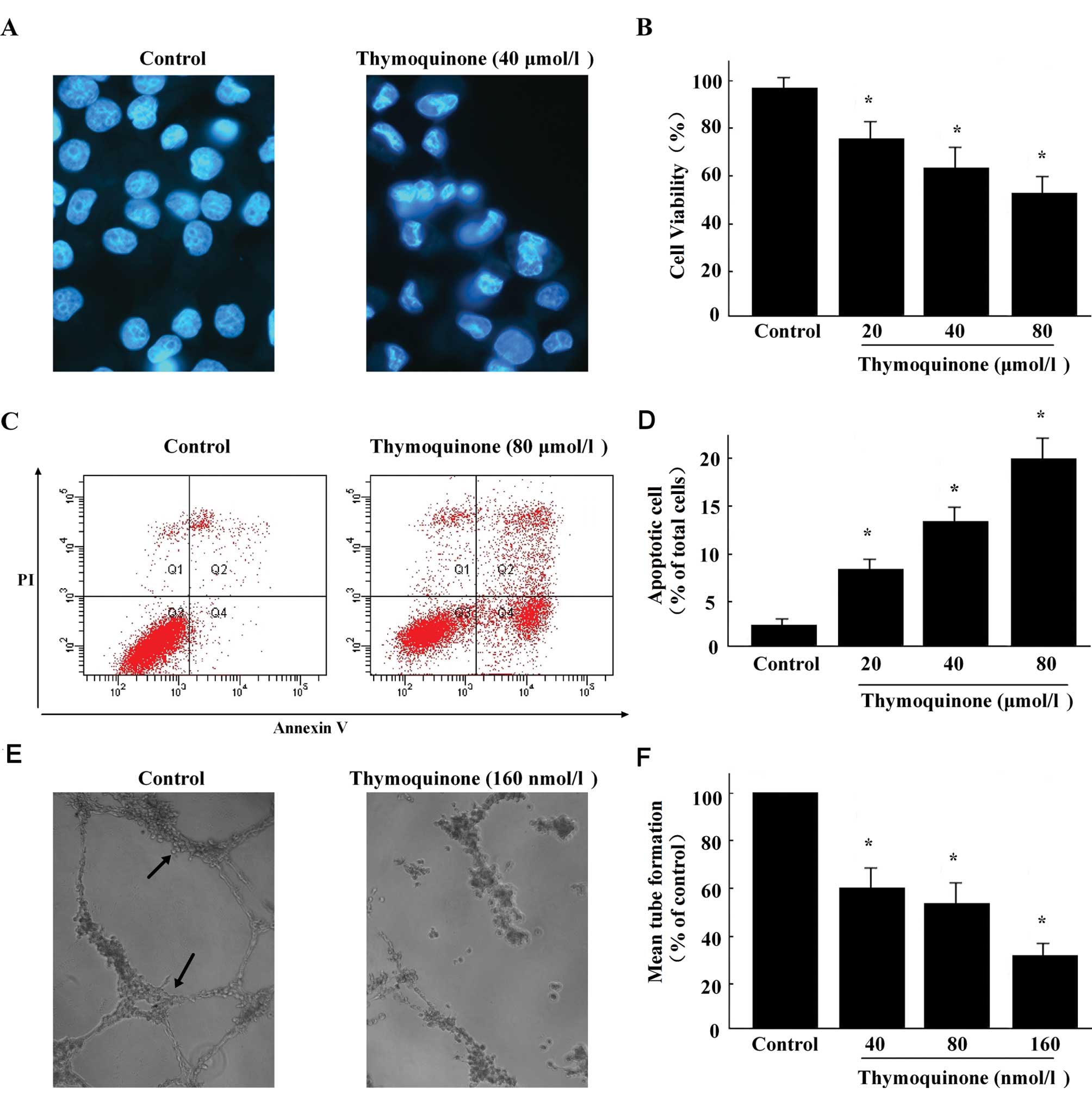
shown in Fig. 1B, the viability of
SaOS-2 cells was decreased dose-dependently in the presence of
thymoquinone.
Thymoquinone induces cell morphological
changes
To further examine the cytotoxic effects of
thymoquinone on SaOS-2 cells, the cells were treated with 40 μmol/l
of thymoquinone for 24 h and then observed under fluorescence
microscope by Hoechst 33258 staining. Results showed that there was
an increase in the number of irregular nuclear, fragmented nucleus,
convoluted nucleus and giant nucleus after treatment with
thymoquinone (Fig. 1A), suggesting
that the DNA fragmentation was occurring in these cells.
Detection of apoptosis with Annexin
V
To determine whether the induction of cell death by
thymoquinone could be linked to apoptosis in SaOS-2 cells, we used
a method that allows one to detect concurrently viable, necrotic,
early apoptotic and late apoptotic cells based on distinct
double-staining patterns with a combination of FITC-conjugated
Annexin V and PI.
Results clearly demonstrated that treatment with
thymoquinone (Fig. 1C) for 24 h
increased the percentage of early apoptotic cells in a
dose-dependent manner (Fig. 1D).
These data indicate that thymoquinone exerts cytotoxic effects
which may be mediated by apoptosis on SaOS-2 cells.
Thymoquinone inhibits tube formation of
HUVECs
Since the final event during angiogenesis is the
organization of endothelial cells in a three-dimensional network of
tube, we performed a tube formation assay to investigate the effect
of thymoquinone on the capillary-like structure formation of
HUVECs. Endothelial cells plated on Matrigel align themselves
forming cords, and the tube-like structure formation was maximal
within 15 h (Fig. 1E). Treatment of
cells with thymoquinone resulted in significant inhibition of
tubule formation of HUVECs on Matrigel (Fig. 1F). Our results clearly demonstrate
that thymoquinone is effective in controlling the tube formation of
endothelial cells in vitro.
Thymoquinone inhibits nuclear factor-κB
DNA-binding activity
Next, we analyzed whether thymoquinone could
abrogate constitutively expressed NF-κB in osteosarcoma cells.
SaOS-2 cells were treated with varying doses of thymoquinone (20,
40 and 80 μmol/l) for 24 h and subjected to gel shift assay (EMSA).
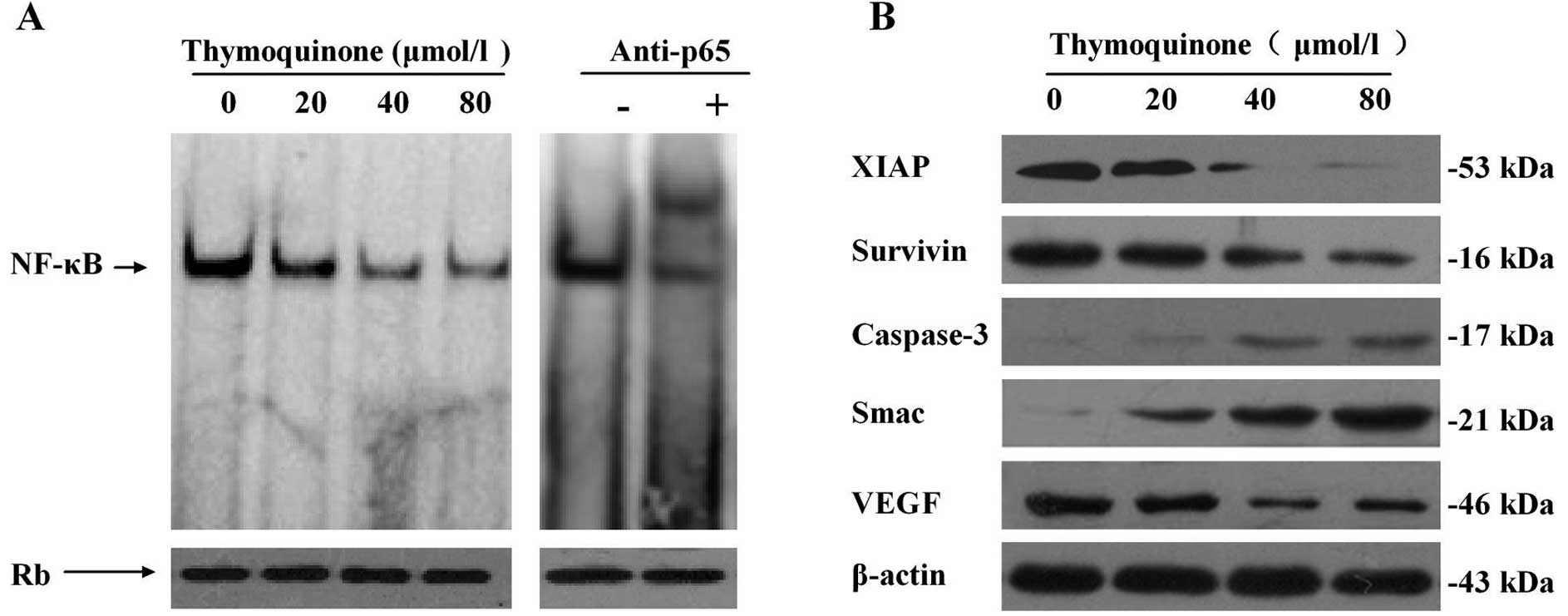
As shown in Fig. 2A, EMSA revealed
that thymoquinone induced a concentration-dependent decrease in
NF-κB DNA binding activity in SaOS-2 cells, which was further
comfirmed by a supershift experiment. These observations provide
strong evidence that thymoquinone is effective in downregulating
NF-κB DNA-binding activity.
Thymoquinone inhibits the expression of
NF-κB-regulated gene products
We assessed the expression of the NF-κB-regulated
genes XIAP and survivin, the overexpression of which has been
linked to tumor survival, chemoresistance, and radioresistance, and
VEGF, which plays an important role in angiogenesis. Western
blotting revealed that thymoquinone induced a
concentration-dependent decrease in the levels of these molecules
compared with the control treatment in SaOS-2 cells. Western
blotting also showed that thymoquinone increased the expression of
caspase-3 and Smac (Fig. 2B).
Thymoquinone inhibits tumor growth in
nude mice
For the in vivo experiment, 12 mice were
divided into two groups as described in Materials and methods. The
results showed that after 15 days of treatment with thymoquinone,
thymoquinone did not produce significant non-tumor toxicity in
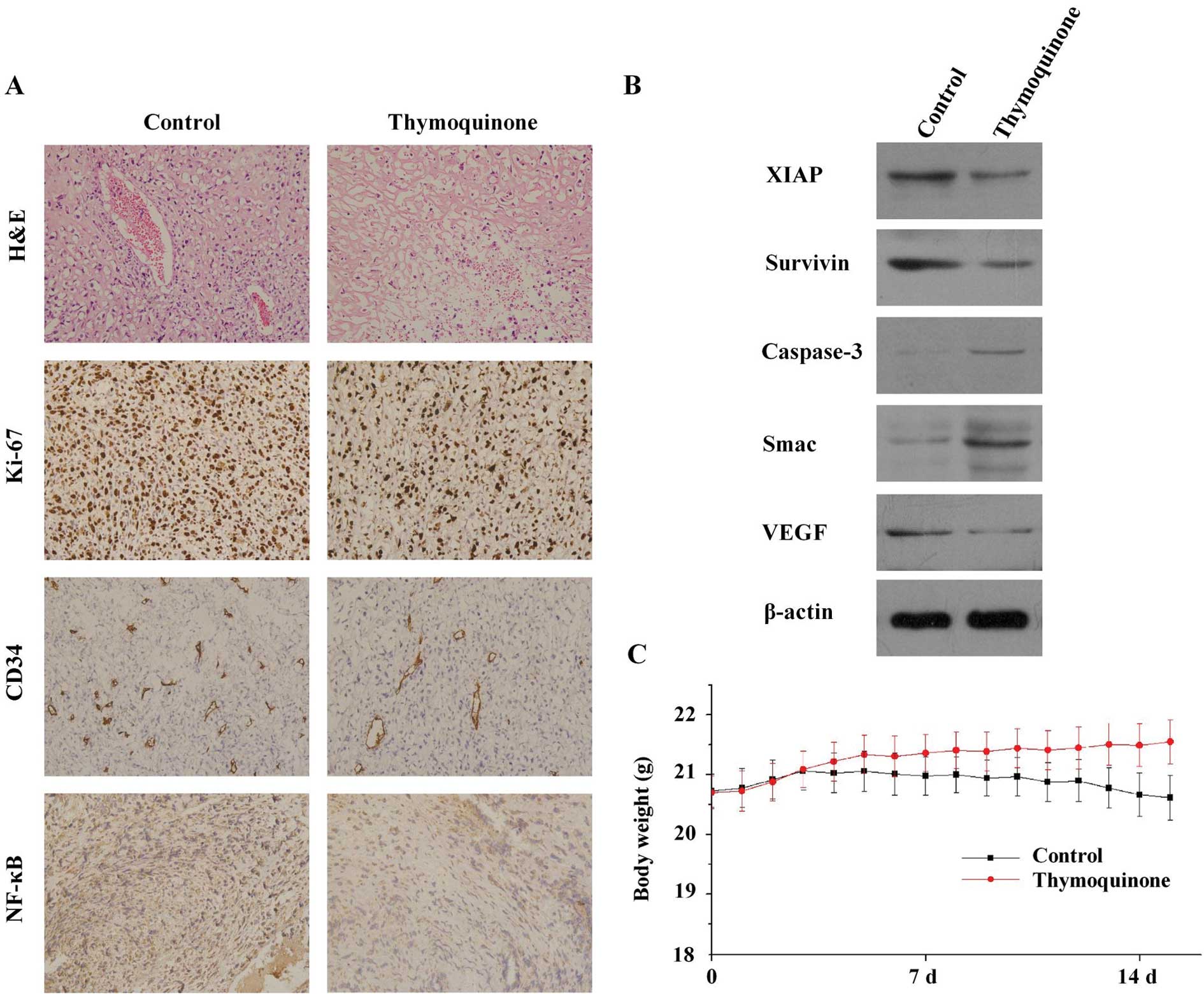
tumor-bearing mice. The average mouse body weight of the control
group decreased from 22.28±1.22 to 21.24±1.32 g, whereas that of
the thymoquinone-treated group increased from 22±1.5 to 24.4±1.2 g
(Fig. 3C). The slight decrease of
the control group is due to the growth of tumors in the xenograft
mice. Next, we determined the mean tumor volume immediately
following euthanization in all mice. The mean tumor volume was
74±25 mm3 in the treated group, vs 126±41 mm3
in the negative control tumors, resulting in a significant
difference of tumor volume in the xenograft model (P<0.05).
Tumor histology, immunohistochemistry and
protein expression in vivo
H&E evaluation of the tumors from all groups
showed high-grade carcinoma associated with tumor apoptosis and
necrosis (Fig. 3A). However, there
were significant differences in the pattern of necrosis,
inflammatory response and fibrosis among the two groups. In the
control group, the tumor was largely viable with high mitosis and
minimal intratumoral stroma, whereas the peripheral tumor was
largely viable and consisted of large nests of neoplastic cells
with minimal intratumoral stroma. By contrast, in the group
receiving thymoquinone treatment there was marked tumor destruction
throughout the entire tumor.
We next examined the expression of the cell
proliferation marker Ki-67 and the microvessel density marker CD34
in tumor tissues from the two groups. The results in Fig. 3A showed that thymoquinone
significantly downregulated the expression of Ki-67 and CD34 in
tumor tissues compared with the control group. For Ki-67, the
proliferation index of control was 77.2±5.1%, but the group
receiving thymoquinone treatment was 49.3±4.5%. For CD34, the
microvessel density was reduced from 192.3±10.2 to 98.6±14.1 after
the treatment of thymoquinone. Furthermore, immunohistochemical
analysis revealed that the expression of NF-κB was significantly
decreased in tumors derived from mice treated with thymoquinone
compared with untreated mice. Tumors also revealed downregulation
of a few important NF-κB-regulated molecules such as survivin, XIAP
and VEGF proteins, which is consistent with our in vitro
results.
Discussion
Successful treatment with chemotherapeutic agents is
largely dependent on their ability to trigger cell death in tumor
cells. However, despite rapid advances in diagnostic and operative
techniques, osteosarcoma remains one of the most challenging human
malignancies to treat, which is partly due to the advanced stage of
the disease and the de novo chemoresistant behavior to
cytotoxic chemotherapeutic agents. Several previous studies
demonstrated that certain phytochemicals present in medicinal herbs
exert antitumorigenic activity by inducing apoptosis in cancer
cells. Thymoquinone has been reported to exert anti-proliferative
effects on several cancer cells in vitro, which are mediated
through the induction of apoptosis (7–11).
Thymoquinone also enhanced sensitivity to chemotherapeutic agents
in pancreatic and lung cancer (8,11). In
this study, we investigated our hypothesis that thymoquinone exerts
antitumor effects on human osteosarcoma SaOS-2 cell line
proliferation and apoptosis. In agreement with previous studies, we
documented that thymoquinone within the range of tested
concentrations, is able to directly inhibit cell viability
dose-dependently in SaOS-2 cells in vitro, and the
morphological changes of thymoquinone-treated cells were typical of
apoptosis, nuclear condensation, and DNA fragmentation.
Furthermore, inhibition of cell growth was correlated with
apoptotic cell death. Fluorescence-activated cell sorting (FACS)
analysis showed thymoquinone increased the percentages of apoptotic
cells in a dose-dependent manner. In agreement with the results of
FACS, induced apoptosis by thymoquinone in pancreatic cells was
validated by activation of caspase-3 (12). However, the growth of mouse
osteoblastic cells did not significantly change by thymoquinone
until 80 μmol/l for 24 h. Similar observations have been reported
in thymoquinone-treated human pancreatic ductal epithelial cells,
suggesting a moderate concentration of thymoquinone does not show
cytotoxicity to normal cells. In addition to in vitro
results, this is the first report to show that thymoquinone is also
as an effective antitumor agent in a mouse osteosarcoma model, and
significant differences in the percentage of Ki-67-positive cells
were noted in tumors derived from the thymoquinone group relative
to untreated animals. Our results are consistent with a previous
study that no significant variation in body weight was detected in
animals after treatment with thymoquinone (12). Taken together, these data suggest
that thymoquinone is a potential drug candidate for cancer
chemotherapies with low chemotoxic side-effects.
The question remains as to how thymoquinone induces
apoptosis in SaOS-2 cells. Emerging evidence has indicated that
overexpression of the pro-survival molecules survivin and XIAP,
members of the inhibitor of apoptosis protein (IAP) family, are
associated with poor prognosis and increased tumor recurrence
(17). Recent studies have
identified the activation of caspase-3 is blocked by the IAPs
(18). However, second
mitochondria-derived activator of caspase (Smac) (also known as
DIABLO) is released from mitochondria into the cytosol during
apoptosis and functions by eliminating inhibitory effects of IAPs
on caspase-3 (19,20). Once released from mitochondria upon
an apoptotic stimulus, Smac docks to the IAPs via an amino-terminal
Reaper motif. This displaces the IAPs from their caspase-binding
sites, thereby relieving the block on caspase activation (21). In the present study, we noted that
thymoquinone per se was effective as a general inducer of
apoptosis in SaOS-2 cells by downregulating survivin and XIAP, and
upregulating cleaved caspase-3 and Smac. To support our hypothesis,
we present evidence documenting a significant reduction of tumors
in vivo by thymoquinone that was found to be associated with
the inhibition of antiapoptotic survivin and XIAP. Thymoquinone was
effective in downregulating IAP proteins, survivin and XIAP, not
only in osteosarcoma cells in vitro but also in preclinical
in vivo conditions.
Angiogenesis is crucial for the growth of solid
tumors not only by supplying oxygen and nutrients for the survival
of tumor cells but also by providing the route for metastatic
spread. Therefore, angiogenesis has been an attractive target for
tumor therapy (22,23). Accumulating evidence has confirmed
the expression of VEGF is closely linked with angiogenesis and has
validated the theory that inhibition of VEGF is a promising
anticancer strategy (24,25). Here, we showed that thymoquinone
effectively inhibited human umbilical vein endothelial cell (HUVEC)
tube formation. Furthermore, we found that thymoquinone inhibited
tumor angiogenesis and prevented osteosarcoma growth in a mouse
osteosarcoma model. In agreement with previous studies (12,26),
our study indicates that thymoquinone can effectively suppress
angiogenesis both in vitro and in vivo at this
experimental condition. Next we showed that thymoquinone
downregulated the expression of VEGF both in vitro and in
vivo, which provided an explanation for its inhibition of
angiogenesis observed in the mouse osteosarcoma model.
Extensive studies have demonstrated that the
rapid-acting primary transcription factor nuclear factor-κB (NF-κB)
is constitutively active in osteosarcoma cell lines (27). Emerging evidence suggests that the
DNA-binding ability of NF-κB has been implicated in survivin and
XIAP expression and the regulation of apoptosis in various cancer
cells (11,25), which underscores the role of NF-κB
activation in mediating chemoresistance and that several
conventional cancer chemotherapeutic agents activate NF-κB leading
to an unfavorable clinical outcome (11,25).
Furthermore, tumor angiogenesis is regulated by numerous
NF-κB-regulated gene products, including VEGF and TNF (25,28).
Finally, evidence indicates the importance of NF-κB in osteosarcoma
and suggests that agents that block NF-κB activation could reduce
chemoresistance and angiogenesis in osteosarcoma and may possibly
be used as a novel therapeutic regimen for osteosarcoma. Previous
studies have demonstrated that curcumin (25), genistein (29) and hyperthermia (30) exert antitumor and anti-angiogenesis
activity through downregulation of NF-κB. In the present study, we
found that thymoquinone abrogates NF-κB activation in osteosarcoma
SaOS-2 cells. In addition, our in vitro results showed that
thymoquinone treatment inhibits NF-κB and exerts anti-proliferative
and apoptosis-inducing effects in SaOS-2 cells, suggesting that
inhibition of NF-κB by thymoquinone is mechanistically associated
with sensitization of osteosarcoma cells to apoptosis. However,
most importantly, these in vitro results, such as the
antitumor activity and inactivation of NF-κB, were recapitulated
in vivo using the mouse osteosarcoma model, which provides a
scientific rationale for the therapeutic exploitation of our
strategy for the treatment of patients with osteosarcoma. These
results provide strong molecular in vitro and in vivo
evidence supporting our hypothesis that inactivation of the NF-κB
signaling pathway by thymoquinone is likely to be an important and
novel strategy for the treatment of osteosarcoma.
In conclusion, our present findings are consistent
with the hypothesis that thymoquinone could downregulate
antiapoptotic and angiogenesis proteins that are regulated by
NF-κB, resulting in loss of osteosarcoma cells to survival. Our
in vitro findings are consistent with the in vivo
results and provide support for the further development of
thymoquinone as an adjunct to conventional chemotherapeutics by
targeted inactivation of NF-κB for the treatment of human
osteosarcoma, and the initiation of clinical trials.
Acknowledgements
The authors thank all the staff in the Laboratory of
Orthopaedic Research Institute and Scientific Research Center of
the Second Affiliated Hospital of Wenzhou Medical College. This
study was supported by grants from the National Natural Science
Foundation of China (31060135/C100302), Project of Science and
Technology Department of Zhejiang Province
(2010C34006/2010C34G2090013), Project of Health Department in
Hainan Province (No.2011-34) and Science-Technology Hall of Hainan
Province (GJXM201102).
References
|
1
|
DuBois S and Demetri G: Markers of
angiogenesis and clinical features in patients with sarcoma.
Cancer. 109:813–819. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lee JA, Kim MS, Kim DH, et al: Relative
tumor burden predicts metastasis-free survival in pediatric
osteosarcoma. Pediatr Blood Cancer. 50:195–200. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Bacci G, Ferrari S, Mercuri M, et al:
Neoadjuvant chemotherapy for osteosarcoma of the extremities in
patients aged 41–60 years - outcome in 34 cases treated with
adriamycin, cisplatinum and ifosfamide between 1984 and 1999. Acta
Orthop. 78:377–384. 2007.
|
|
4
|
Chou AJ, Merola PR, Wexler LH, et al:
Treatment of osteosarcoma at first recurrence after contemporary
therapy: the Memorial Sloan-Kettering Cancer Center experience.
Cancer. 104:2214–2221. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Worthen DR, Ghosheh OA and Crooks PA: The
in vitro anti-tumor activity of some crude and purified components
of blackseed, Nigella sativa L. Anticancer Res. 18:1527–1532.
1998.PubMed/NCBI
|
|
6
|
Ali BH and Blunden G: Pharmacological and
toxicological properties of Nigella sativa. Phytother Res.
17:299–305. 2003. View
Article : Google Scholar
|
|
7
|
Gali-Muhtasib H, Diab-Assaf M, Boltze C,
et al: Thymoquinone extracted from black seed triggers apoptotic
cell death in human colorectal cancer cells via a p53-dependent
mechanism. Int J Oncol. 25:857–866. 2004.PubMed/NCBI
|
|
8
|
Jafri SH, Glass J, Shi RH, Zhang SL,
Prince M and Kleiner-Hancock H: Thymoquinone and cisplatin as a
therapeutic combination in lung cancer: in vitro and in vivo. J Exp
Clin Cancer Res. 29:872010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
El-Mahdy MA, Zhu QZ, Wang QE, Wani G and
Wani AA: Thymoquinone induces apoptosis through activation of
caspase-8 and mitochondrial events in p53-null myeloblastic
leukemia HL-60 cells. Int J Cancer. 117:409–417. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kaseb AO, Chinnakannu K, Chen D, et al:
Androgen receptor- and E2F-1-targeted thymoquinone therapy for
hormone-refractory prostate cancer. Cancer Res. 67:7782–7788. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Banerjee S, Kaseb AO, Wang ZW, et al:
Antitumor activity of gemcitabine and oxaliplatin is augmented by
thymoquinone in pancreatic cancer. Cancer Res. 69:5575–5583. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yi TF, Cho SG, Yi ZF, et al: Thymoquinone
inhibits tumor angiogenesis and tumor growth through suppressing
AKT and extracellular signal-regulated kinase signaling pathways.
Mol Cancer Ther. 7:1789–1796. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sethi G, Ahn KS and Aggarwal BB: Targeting
nuclear factor-kappa B activation pathway by thymoquinone: Role in
suppression of antiapoptotic gene products and enhancement of
apoptosis. Mol Cancer Res. 6:1059–1070. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Chehl N, Chipitsyna G, Gong QK, Yeo CJ and
Arafat HA: Anti-inflammatory effects of the Nigella sativa
seed extract, thymoquinone, in pancreatic cancer cells. HPB.
11:373–381. 2009.
|
|
15
|
Roepke M, Diestel A, Bajbouj K, et al:
Lack of p53 augments thymoquinone-induced apoptosis and caspase
activation in human osteosarcoma cells. Cancer Biol Ther.
6:160–169. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Al-Romaih K, Somers GR, Bayani J, et al:
Modulation by decitabine of gene expression and growth of
osteosarcoma U2OS cells in vitro and in xenografts: identification
of apoptotic genes as targets for demethylation. Cancer Cell Int.
7:142007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Srinivasula SM and Ashwell JD: IAPs:
what’s in a name? Mol Cell. 30:123–135. 2008.
|
|
18
|
Yang YL and Li XM: The IAP family:
endogenous caspase inhibitors with multiple biological activities.
Cell Res. 10:169–177. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Verhagen AM, Ekert PG, Pakusch M, et al:
Identification of DIABLO, a mammalian protein that promotes
apoptosis by binding to and antagonizing IAP proteins. Cell.
102:43–53. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Du CY, Fang M, Li YC, Li L and Wang XD:
Smac, a mitochondrial protein that promotes cytochrome c-dependent
caspase activation by eliminating IAP inhibition. Cell. 102:33–42.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Srinivasula SM, Hegde R, Saleh A, et al: A
conserved XIAP-interaction motif in caspase-9 and Smac/DIABLO
regulates caspase activity and apoptosis. Nature. 410:112–116.
2001. View
Article : Google Scholar : PubMed/NCBI
|
|
22
|
Bergers G and Benjamin LE: Tumorigenesis
and the angiogenic switch. Nat Rev Cancer. 3:401–410. 2003.
View Article : Google Scholar
|
|
23
|
Cooney MM, van Heeckeren W, Bhakta S,
Ortiz J and Remick SC: Drug insight: vascular disrupting agents and
angiogenesis - novel approaches for drug delivery. Nat Clin Pract
Oncol. 3:682–692. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yu HG, Yu LL, Yang YN, et al: Increased
expression of RelA/nuclear factor-kappa B protein correlates with
colorectal tumorigenesis. Oncology. 65:37–45. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kunnumakkara AB, Guha S, Krishnan S,
Diagaradjane P, Gelovani J and Aggarwal BB: Curcumin potentiates
antitumor activity of gemcitabine in an orthotopic model of
pancreatic cancer through suppression of proliferation,
angiogenesis, and inhibition of nuclear factor-kappa B-regulated
gene products. Cancer Res. 67:3853–3861. 2007. View Article : Google Scholar
|
|
26
|
Erdurmus M, Yagci R, Yilmaz B, et al:
Inhibitory effects of topical thymoquinone on corneal
neovascularization. Cornea. 26:715–719. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lamoureux F, Picarda G, Rousseau J, et al:
Therapeutic efficacy of soluble receptor activator of nuclear
factor-kappa B-Fc delivered by nonviral gene transfer in a mouse
model of osteolytic osteosarcoma. Mol Cancer Ther. 7:3389–3398.
2008. View Article : Google Scholar
|
|
28
|
Aggarwal BB: Nuclear factor-kappa-B: the
enemy within. Cancer Cell. 6:203–208. 2004.
|
|
29
|
Banerjee S, Zhang YX, Wang ZW, et al: In
vitro and in vivo molecular evidence of genistein action in
augmenting the efficacy of cisplatin in pancreatic cancer. Int J
Cancer. 120:906–917. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Adachi S, Kokura S, Okayama T, et al:
Effect of hyperthermia combined with gemcitabine on apoptotic cell
death in cultured human pancreatic cancer cell lines. Int J
Hyperthermia. 25:210–219. 2009. View Article : Google Scholar : PubMed/NCBI
|