Introduction
Esophageal carcinoma is one of the most common
malignant gastrointestinal tumors with a high incidence in China
and is characterized by a unique geographical distribution.
Therefore, it is necessary to elucidate the underlying mechanisms
and to search for chemopreventive tools or drugs to decrease the
risk of carcinogenesis of esophageal epithelial cells.
Studies have indicated that maintenance of telomere
length is important in preventing the consumption of telomere DNA
during cell division. Studies found significant associations
between short telomere length and increased esophageal cancer risk
(1,2).
Telomerase is a ribonucleoprotein enzyme complex
that adds 5′-TTAGGG-3′ repeats onto the ends of human chromosomes,
providing a telomere maintenance mechanism for ~90% of cancers
(3). Telomerase consists of several
subunits, including hTERC that serves as a template during telomere
elongation and a catalytic subunit, human telomerase reverse
transcriptase (hTERT), which has reverse transcriptase activity.
Moreover, telomerase activity is dependent on the expression of 2
main core component genes, hTERT and hTERC. This has prompted a
large number of studies addressing telomerase activity using
telomeric repeat amplification protocol (TRAP) assay (4,5),
amplification of hTERC using in situ hybridization (6–11) or
expression of hTERT using immunohistochemistry (IHC) in cervical
cancer and cervical intraepithelial neoplasia lesions (12–14),
which indicated that the telomerase activation mediated tumor cell
growth.
Studies have shown that inhibition of telomerase
results in gradual erosion of telomeres followed by cessation of
proliferation or apoptosis, and may thus be a promising target for
cancer therapy. Pin2/TRF1 interacting protein X1 (PinX1) was
previously found to be a tumor suppressor and telomerase inhibitor
in vivo. It is expressed in normal human tissues, but is
not, or is less, expressed in tumor tissues. Studies have found
that PinX1 can inhibit telomerase activity in gastric and liver
tumor cells and can induce apoptosis (15–18).
In addition, the expression of PinX1 has been positively correlated
with telomerase activity in leukemia (19,20).
However, some studies on prostate cancer, gastrointestinal cancer
and medulloblastoma indicate that gene polymorphism rather than
PinX1 expression is the key factor in inhibiting telomerase
(21–23) and PinX1 as a microtubule binding
protein plays an important role in stabilizing chromosome (24). In short, the mechanisms by which
PinX1 regulates telomerase/telomere in tumor cells are complex and
may vary in different tumors. The effect of PinX1 on esophageal
carcinoma cell proliferation and the mechanisms by which PinX1
affects telomerase activity have yet to be reported. Therefore, in
the present study, we detected the expression of PinX1 and
telomerase activity in esophageal squamous cell carcinoma (ESCC),
dysplasia of esophageal squamous epithelium and normal esophageal
mucosa in order to explore the relationship with carcinogenesis of
esophageal epithelium; moreover, we created Eca109 stable cell
lines overexpressing PinX1 to investigate the effects of PinX1 on
the telomerase activity and cell proliferation, apoptosis, and
further explored the possible mechanism of PinX1-mediated
carcinogenesis of esophageal epithelial cells.
Materials and methods
Reagents and antibodies
TRIzol reagent and Lipofectamine® 2000
were obtained from Invitrogen (Carlsbad, CA, USA). Horseradish
peroxidase (HRP) AffiniPure goat anti-mouse/rabbit IgG(H+L) was
purchased from Zhongshan Golden Bridge Biotechnology Co., Ltd.
(Beijing, China). FITC AffiniPure goat anti-mouse IgG was from
Jackson ImmunoResearch. Telomere RNA probe kit was from (Dako, USA)
and Telomerase activity kit from Jermaine gene Co., (USA). pCDNA3.1
vector and pCDNA3.1/PinX1 were obtained from Invitrogen.
Geneticin® (G418) was from Gibco (USA). The mouse
anti-hTERT and mouse anti-PinX1 antibodies were from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA).
Clinical materials
One-thousand cases of esophageal tissues from
patients without chemotherapy and radiotherapy were collected from
an esophageal carcinoma high-risk area of China. All specimens were
verified by pathologic diagnosis and we selected 44 cases of ESCC,
50 dysplasia of esophageal squamous epithelium (22 cases of mild
dysplasia and 28 cases of severe dysplasia) and 36 normal
esophageal mucosa to be used to detect the length of telomere and
hTERT protein expression.
Moreover, esophageal carcinoma tissues, paired
adjacent mucosa (2–5 cm from margin of esophageal carcinoma) and
paired normal mucosa (at least 5 cm from margin of esophageal
carcinoma) were obtained from resected surgical specimens of ESCC.
All specimens were verified by pathologic diagnosis, 50 cases of
ESCC, dysplasia of esophageal squamous epithelium and normal
esophageal mucosa were selected from 130 specimens. Fifty ESCC
tissues included 39 cases of well and moderately differentiated
ESCC and 11 cases of poorly differentiated ESCC; fibrous membrane
invasion (n=34), fibrous membrane untouched (n=16); lymph node
metastasis positive (n=17), lymph node metastasis negative (n=33).
Experimental protocols were approved by the Institutional Human
Care and Use Committee of Hebei Medical University.
Cell lines
Human esophageal cancer cells (Eca109) were grown
under humidified air with 5% CO2 in an incubator at 37°C
in RPMI-1640 supplemented with 10% fetal bovine serum, 100 μg/ml
streptomycin and 100 U/ml penicillin.
Fluorescence in situ hybridization (FISH)
for telomere length
The single cell suspension was acquired as
previously described and incubated with FITC-(CCCTAA)3PNA probe for
15 min at 87°C. Then, after overnight hybridization at room
temperature, the cell suspensions were washed, followed by staining
with propidium iodide (PI) solution for 2 h at 4°C. Subsequently,
the cells were analyzed in an Epics-XL II Flow Cytometer (Beckman
Coulter, Miami, FL, USA). Q-FISH was used to represent the telomere
length. Q-FISH = fluorescence intensity in experimental group -
fluorescence intensity in background group.
IHC for detection of h-TERT protein
The tissues were fixed in 4% formaldehyde. Antigen
recovery was performed using a microwave. The sections were
incubated with primary antibodies against hTERT (1:100) overnight
at 4°C. The following day, the sections were incubated with
polyperoxidase-anti-mouse IgG at 37°C and finally stained with
diaminobenzidine. The sections were imaged with an Olympus
microscope. Positive staining of hTERT protein was located in the
nuclei of epithelial cells. The result criteria: the number of
positive cells was >10%.
Measurement of hTERT and PinX1 protein by
FCM analysis
The tissues in different groups were washed with
phosphate buffered saline (PBS), fixed in 70% ethanol, and the
single cell suspensions were then collected and washed with PBS,
stained with mouse anti-hTERT or PinX1 antibodies at room
temperature for 30 min. Then, the cells were washed 3 times with
PBS and incubated with FITC-anti-mouse IgG at 37°C. The stained
cells were analyzed in an Epics-XL II Flow Cytometer. Fluorescence
index (FI) represents the relative protein expression content. FI =
(fluorescence intensity in experimental group - fluorescence
intensity in control group)/fluorescence intensity in normal
control.
TRAP-silver stain for quantification of
telomerase activity
Biopsy samples were stored in liquid nitrogen. The
extraction of telomerase protein and evaluation of its activity
were measured by the TRAP method using the TRAPeze-XL Telomerase
Detection kit as previously described (12). Briefly, extracts from 100 mg of
frozen esophageal tissues were homogenized in ~200 μl of lysis
buffer. After 30 min of incubation on ice, the suspensions were
centrifuged at 16,000 × g for 30 min at 4°C, after which the
supernatant was frozen rapidly and stored at −70°C. The PCR
conditions were: after 10 min for telomerase extension at 23°C,
95°C for 5 min to activate the Taq polymerase; 27 cycles at 94°C
for 30 sec, 50°C for 30 sec, and 72°C for 90 sec; an extension at
72°C for 10 min; finally, a 4°C incubation. Equal amounts of
amplified products were separated by 12% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE). The gel was
stained with 2% silver nitrate cream for 20 min after fixing with
DNA fixative solution. After washing twice with d.d. water, the gel
was placed in medium of 1.5% NaOH and 0.4% formaldehyde and waved
lightly until clear DNA ladder appeared.
RNA isolation and measurement of PinX1
mRNA by RT-PCR
Total RNA was extracted from esophageal tissues or
Eca109 cells with TRIzol reagent according to the manufacturer’s
instructions. Total RNA (2 μg) was reverse transcribed into cDNA by
AMV reverse transcriptase at 42°C for 1 h and then heated to 94°C
for 5 min in a total reaction volume of 20 μl. PCR conditions used
for PinX1 and internal reference GAPDH were: 94°C for 2 min
followed by 30 cycles of 94°C for 30 sec, 55°C for 30 sec and 72°C
for 30 sec, and 72°C for 5 min. The specific primers were: PinX1
forward, 5′-CCTCAGAACA CTGCCTGGAG-3′ and reverse,
5′-GTTCCACCTGCGTCT CAGAA-3′; GAPDH forward, 5′-CCTGAGGGTTCTTTGT
GCTGA-3′ and reverse, 5′-AAAGGCTCAACCTTCCCCAT-3′. The expected PCR
products were 577 bp and 122 bp for PinX1 and GAPDH, respectively.
The amplicons were analyzed by electrophoresis, imaged using UVI
gel imaging system and quantified using Gel-proAnalyzer3.1
software. Expression levels of PinX1 were normalized to internal
reference GAPDH.
Cell culture and preparation of stable
Eca109 cell lines overexpressing PinX1
Stable transfections of Eca109 cells with pCDNA3.1
vector and pCDNA3.1/PinX1 were performed with
Lipofectamine® 2000, according to the manufacturer’s
instructions. Subsequently, cells were cultured in selection medium
containing 0.5 mg/ml Geneticin® (G418) for 4 weeks
before single clones were isolated. The clones were further
expanded in selection medium containing Geneticin (0.5 mg/ml).
Untransfected cells and cells transfected with pCDNA3.1 were used
as controls. Cells were observed 24–48 h after transfection under a
fluorescence microscope to examine transfection efficiency.
Detection of PinX1 protein by western
blot analysis
Total protein extraction from cultured Eca109 cells
was performed as previously described (25). Protein was quantified with a Bio-Rad
Protein Colorimetric Assay (Bio-Rad, Hercules, CA, USA). Equal
amounts of protein extracts were separated by 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes (Millipore
Corp., Billerica, MA, USA) as previously described (26). The concentration of anti-PinX1
antibody and anti-β-actin antibody was 1:100 (41 kDa) and
1:1,000(42 kDa), respectively. PinX1 protein expression was
quantified by comparison with β-actin.
Measurement of cell proliferation by
MTT
Eca109 cells at the logarithmic phase were
inoculated into 96-well plates with 1×105 cells/well.
Cell viability at 24 h was examined using the MTT method.
OD490 nm values were detected in 6 duplicate wells and
their averages were used to plot growth curve and calculate the
growth inhibition rate of each treatment using the following
formula: Growth inhibition rate Ir = OD490 nm of the
control group - OD490 nm of the treatment
group/OD490 nm of the control group × 100%.
Measurement of cell cycle and apoptosis
by flow cytometry
Forty-eight hours after transfection, Eca109 cells
were collected, washed with PBS, resuspended in PBS at
1×106/ml, and stained with Annexin V and PI for 15 min
in the dark. Apoptotic cells were then analyzed by flow cytometry
and apoptotic index (AI) was calculated using AI = apoptotic
cells/total cells × 100%. Cell cycle was determined after fixing
with precooled 75% ethanol at 4°C and washing with PBS.
Statistical analysis
Data are expressed as mean ± standard variation and
analyzed using SPSS 13.0 statistical software package. Differences
between samples were tested using single factor analysis of
variance and LSD method for multiple comparisons. A P-value
<0.05 was considered to indicate a statistically significant
difference. Prior to the comparison, data homogeneity of variance
was first examined using F-test. In the case of heterogeneity of
variance, the approximate variance F-test/Welch method was
used.
Results
Decrease of length of telomere DNA and
increase of hTERT protein in the carcinogenesis of esophageal
epithelial cells
Value of Q-FISH was analyzed by FCM assay in the
normal esophageal epithelium group, the mild dysplasia group, the
severe dysplasia group and the carcinoma group. Q-FISH in the
carcinoma group was lower than in the severe dysplasia group
(P<0.01), but it was lower in the severe dysplasia group than in
the mild dysplasia group (P<0.01, Table I). There was a negative correlation
between telomere length and cytologic grade in esophageal cancer
(r=−0.79, P<0.01). As shown in Table
I, the shorten rate of telomere length in the carcinoma group
was higher compared with that in the mild dysplasia and severe
dysplasia group.
 | Table ILength of telomere DNA in various
lesions of esophageal epithelial cells by FCM (mean ± standard
variation). |
Table I
Length of telomere DNA in various
lesions of esophageal epithelial cells by FCM (mean ± standard
variation).
| | | Telomere length, n
(%) |
|---|
| | |
|
|---|
| Group | n | Q-FISH | Extenda | Shortenb | Normal |
|---|
| Normal | 36 | 50.83±8.86 | | | |
| Mild dysplasia | 22 | 49.51±3.16 | 2 (9.09) | 13 (59.09) | 7 (31.82) |
| Severe
dysplasia | 28 | 36.96±8.02a | 2 (7.14) | 20 (71.43)a | 6 (21.43) |
| Carcinoma | 44 | 27.80±6.59b | 3 (6.82) | 35 (88.64)b | 6 (13.64) |
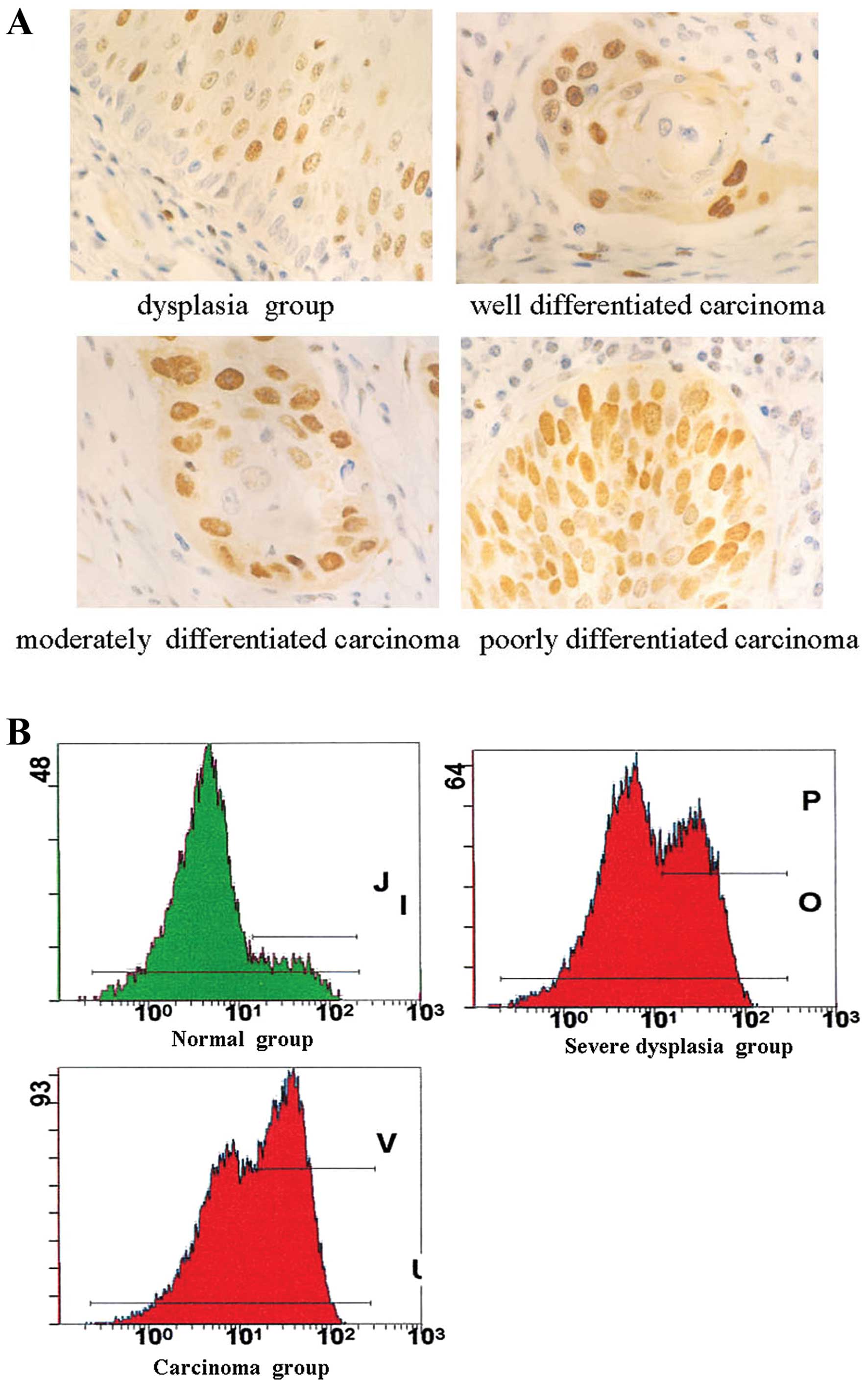
The positive expression of hTERT protein was located
in the nucleus of esophageal epithelial cells and cancerous cells
by immunohistochemical staining (Fig.
1A). The positive rate of hTERT in incisal margin normal tissue
was 11.1% (4/36) and those in para-tumorous dysplasia tissues and
carcinoma were 44.0% (22/50) and 86.37% (38/44), the positive
expression rate increased gradually with carcinogenesis of
esophageal epithelial cells (P<0.01).
FI value of hTERT in the normal esophageal
epithelium group, mild dysplasia group, severe dysplasia group and
carcinoma group was 0.87±0.18, 1.13±0.19, 1.39±0.24 and 1.84±0.21
(Table II) (Fig. 1B). FI value of hTERT in the
carcinoma group was higher than in the severe dysplasia group
(P<0.01), and it was higher in the severe dysplasia group than
in the mild dysplasia group (P<0.01). In addition, FI value of
hTERT was positively related with cytologic grade (r=0.84,
P<0.01).
| Table IIContent of hTERT protein in various
lesions of esophageal epithelial cells by FCM (mean ± standard
variation). |
Table II
Content of hTERT protein in various
lesions of esophageal epithelial cells by FCM (mean ± standard
variation).
| Group | n | hTERT FI | F | P-value |
|---|
| Normal | 36 | 0.87±0.18 | 73.52 | <0.01 |
| Mild dysplasia | 22 | 1.13±0.19 | | |
| Severe
dysplasia | 28 | 1.39±0.24a | | |
| Carcinoma | 44 | 1.84±0.21b | | |
There was a negative correlation between Q-FISH
value of telomere and FI value of hTERT (r=−7.49, P<0.01).
Increase of telomerase activity and
decrease of PinX1 in the carcinogenesis of esophageal epithelial
cells
The telomerase activity in different histological
groups is summarized in Table
III. The levels of telomerase activity were 0.073±0.039 in the
normal group, 0.429±0.346 in the dysplasia group, 1.457±0.838 in
the carcinoma group. Telomerase activity was higher in the
carcinoma group compared to the control normal or dysplasia group
(P=0.00). Moreover, the A-value of telomerase activity was related
to the tumor tissue grade and lymph node metastasis (P<0.05),
but not to other clinicopathological features in ESCCs (P>0.05,
Table IV).
| Table IIIDetection of telomerase activity in
various esophageal tissues. |
Table III
Detection of telomerase activity in
various esophageal tissues.
| | Telomerase
activity |
|---|
| |
|
|---|
| Group | n | Positive no. | Rate | A-value |
|---|
| Carcinoma
group | 50 | 42 | 84a | 1.457±0.838a,b |
| Dysplasia
group | 50 | 31 | 62a | 0.429±0.346a |
| Normal group | 50 | 3 | 6 | 0.073±0.039 |
| Table IVRelationship between telomerase
activity and clinicopathological features in human esophageal
squamous cell carcinomas. |
Table IV
Relationship between telomerase
activity and clinicopathological features in human esophageal
squamous cell carcinomas.
| | Telomerase
activity | |
|---|
| |
| |
|---|
| Clinicopathological
feature | n | Positive no. | Rate (%) | A-value | P-value |
|---|
| Gender |
| Male | 35 | 30 | 85.71 | 1.253±0.672 | >0.05 |
| Female | 15 | 12 | 80 | 1.702±0.823 | |
| Age (years) |
| ≤60 | 36 | 30 | 83.33 | 1.465±0.672 | >0.05 |
| >60 | 14 | 12 | 85.71 | 1.797±0.952 | |
| Depth of
invasion |
| Fibrous
membrane | 16 | 13 | 81.25 | 1.467±0.923 | >0.05 |
| Fibrous
membrane | 34 | 29 | 85.29 | 1.534±0.782 | |
| Untouched | | | | | |
| Grade |
| Well/moderately
differentiated | 39 | 32 | 82.05 | 1.163±0.438 | <0.05 |
| Poorly
differentiated | 11 | 10 | 90.91 |
2.235±0.814a | |
| Lymph node
metastasis |
| Positive | 17 | 16 | 94.12 |
1.917±0.814a | <0.05 |
| Negative | 33 | 26 | 78.79 | 1.097±0.865 | |
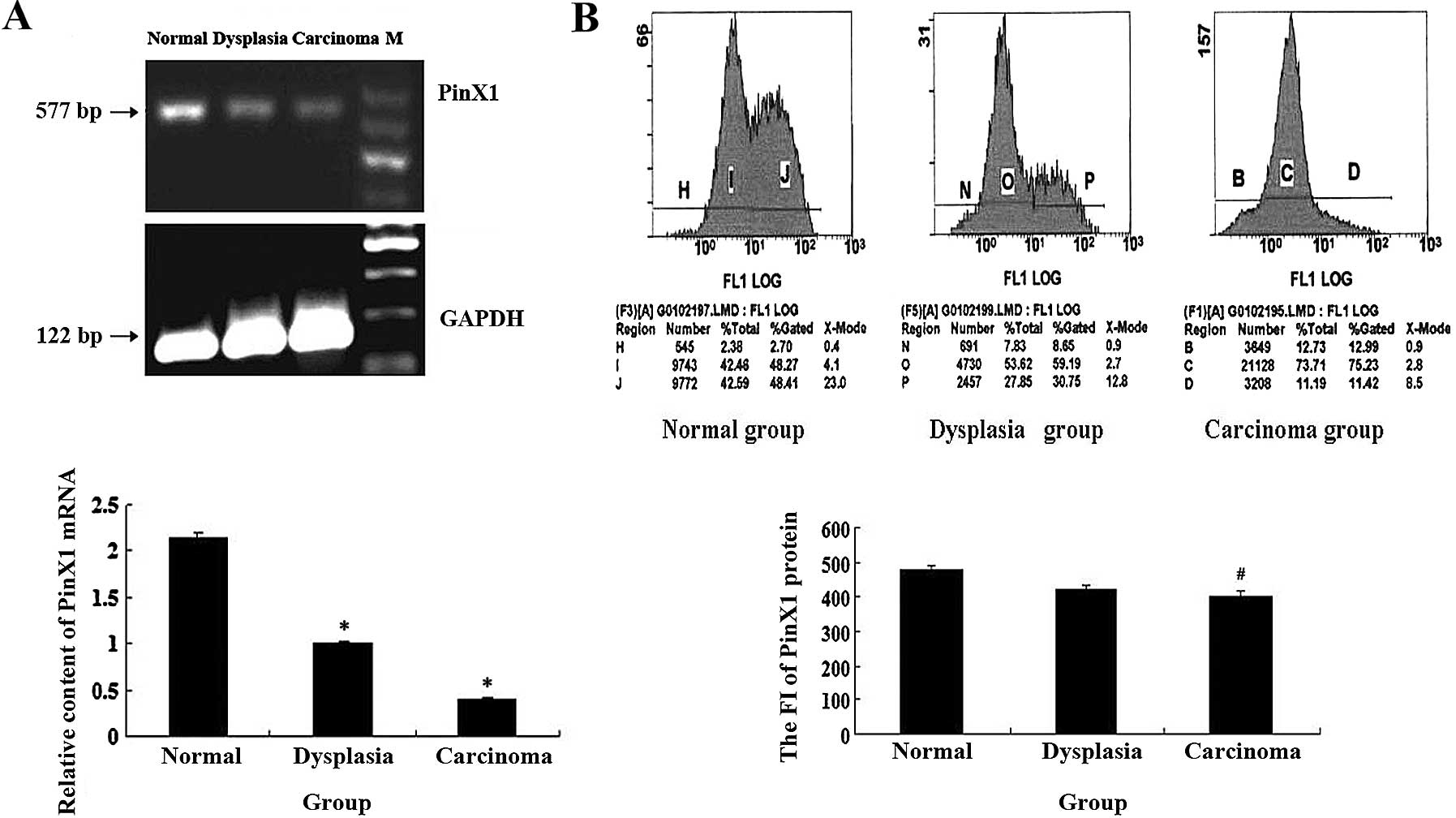
The results of RT-PCR showed that the relative
expression of PinX1 mRNA in esophageal cancer tissues was
significantly lower than that in normal tissues and dysplasia
esophageal tissues (P<0.01) (Fig.
2A). As shown in Fig. 2B, the
PinX1 protein level in esophageal cancer tissues was significantly
lower than in the dysplasia and normal group, and there was a
negative correlation with differentiation, invasive depth and lymph
node metastasis (P<0.05, Table
V) and no correlation with age and gender in ESCCs (P>0.05,
Table V).
| Table VThe relationship between PinX1
protein expression and clinicopathological features in ESCC. |
Table V
The relationship between PinX1
protein expression and clinicopathological features in ESCC.
| Clinicopathological
feature | n | PinX1 protein | P-value |
|---|
| Age (years) |
| ≤60 | 30 | 445.88±22.13 | |
| >60 | 20 | 443.38±23.95 | 0.63 |
| Gender |
| Male | 35 | 442.70±22.57 | |
| Female | 15 | 449.39±23.18 | 0.23 |
| Differentiation
grade |
|
Well/moderately | 39 | 441.04±22.02 | |
| Poorly | 11 | 455.33±22.22 | 0.01 |
| Depth of
invasion |
| Fibrous
membrane | 34 | 448.80±22.76 | |
| Fibrous
membrane | 16 | 436.45±21.02 |
0.02a |
| Untouched | | | |
| Lymph node
metastasis |
| Positive | 17 | 453.91±19.78 | |
| Negative | 33 | 440.14±23.04 | 0.01 |
There was a significant negative correlation between
telomerase activity expression and PinX1 protein expression
(rs=−0.883, P=0.000).
Overexpression of PinX1 in telomerase
activity, cell proliferation and apoptosis in Eca109 cells
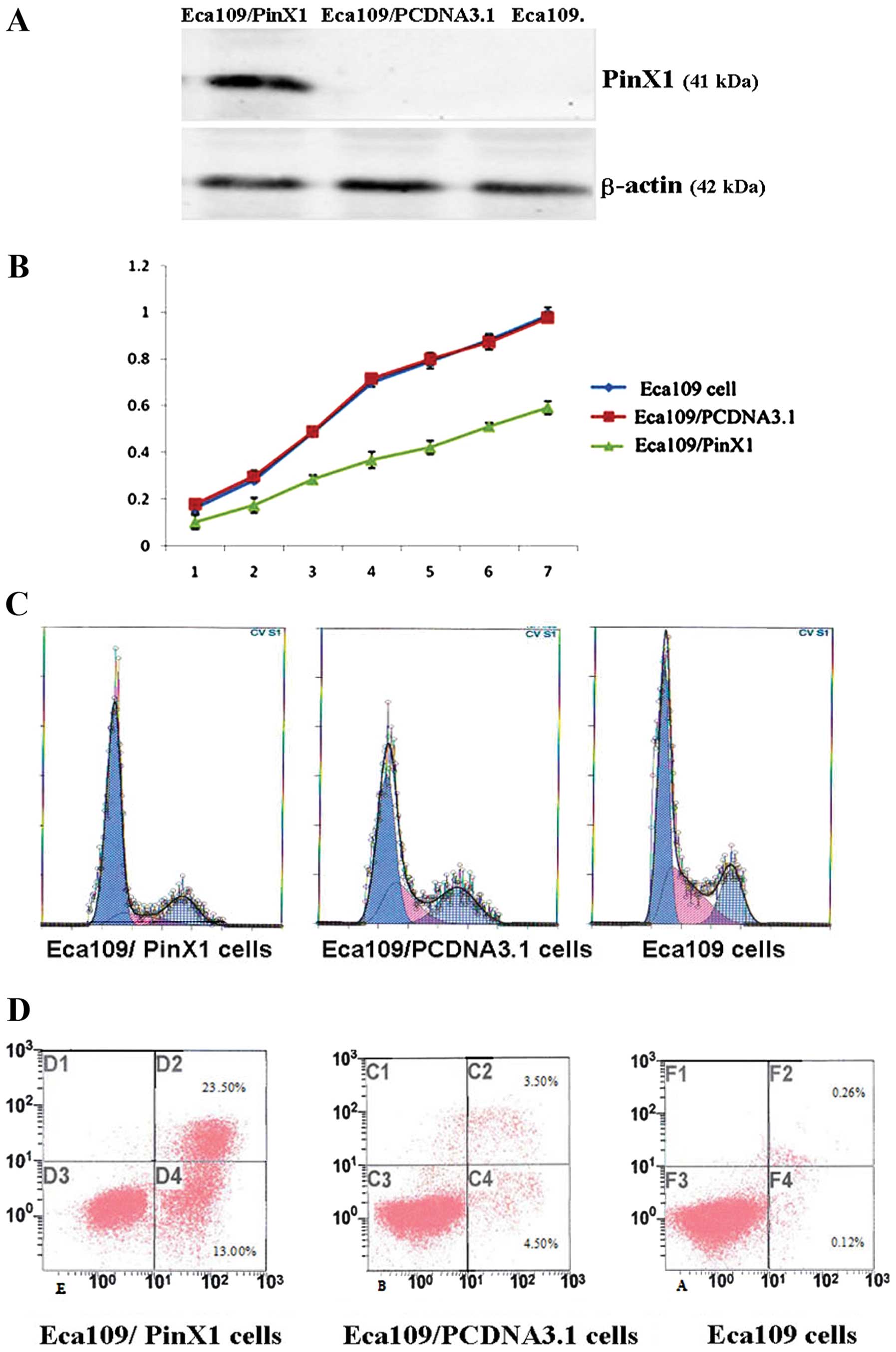
To investigate the effect of PinX1 on telomerase
activity and cell growth, Eca109 cells were transfected with
expression vector for PinX1 (pCR3.1/PinX1) or control vector
(pCR3.1). The expression of PinX1 protein was 4.5-fold greater than
that of the control group (Fig.
3A). We further examined the effect of PinX1 on the telomerase
activity by TRAP-PCR, cell proliferation and apoptosis by FCM. In
Eca109 cells, PinX1 overexpression decreased Eca109 cell growth
(Fig. 3B) and blocked cells into
the G0/G1 stage (Fig. 3C) (Table VI). The apoptosis rate was higher
in PinX1-transfected Eca109 than in PinX1-untransfected cells or
cells transfected with void vectors only by FCM (Fig. 3D) (Table VI).
| Table VIChanges of cell cycle distribution,
PI and apoptotic rate in different groups. |
Table VI
Changes of cell cycle distribution,
PI and apoptotic rate in different groups.
| Cell | G0/G1 | PI | Apoptotic rate |
|---|
| Eca109 | 53.14±4.83 | 33.17±0.47 | 0.27±0.18 |
|
Eca109/PCDNA3.1 | 47.27±3.73 | 35.64±0.65 | 3.40±1.09 |
| Eca109/PinX1 | 70.58±5.26a | 13.58±0.59a | 23.28±5.73a |
The expression of telomerase activity was lower in
PinX1-transfected Eca109 than in PinX1-untransfected cells or cells
transfected with vectors only (P<0.05, Table VII).
| Table VIIDetection of telomerase activity in
various cells. |
Table VII
Detection of telomerase activity in
various cells.
| Cell | A-value | F | P-value |
|---|
| Eca109 | 2.446±0.652 | | |
|
Eca109/PCDNA3.1 | 2.137±0.475 | | |
| Eca109/PinX1 | 0.874±0.439a | 163.25 | 0.000 |
There was a significant positive correlation between
telomerase activity expression and PI expression
(rs=0.451, P=0.000).
Discussion
Telomere shortening results in chromosomal
instability which, in the absence of normal cellular senescence
processes, can lead to cancer development (27). In the present study, we collected
esophageal tissue in a high incidence area of China in order to
detect the length of telomere. The results showed that telomere
shortening occurred in the carcinogenesis of esophageal epithelial
cells, which indicated that telomere shortening is an important
event in the carcinogenesis of esophageal squamous epithelial
cells.
Telomerase is a special reverse transcriptase that
is composed of RNA and protein and regulates the length of
telomere. hTERT is the key component in telomerase and plays an
important role in genetic stability and maintenance of chromosomes.
Several studies have indicated that telomerase is only slightly
expressed in normal cells, but its expression and activity are
enhanced in most immortalized tumor cells (28,29).
Previous studies have verified that telomerase is significantly
related to the incidence of human malignant tumors (30). Enhancement of its activity is the
power source of constantly increased proliferation, invasion and
metastasis of tumor cells. Therefore, downregulation of telomerase
activity in tumor cells may be one of the important therapeutic
measures to inhibit tumor growth. Enhanced hTERT/CMV promoter can
reduce telomerase activity, eventually leading to the death of
tumor cells (31). The PinX1 gene
is located on chromosome 8p22–23 region and can attenuate
telomerase activity, inhibit growth of tumor cells and induce
apoptosis. Lack of endogenous PinX1 leads to increased telomerase
activity and tumorigenicity in nude mice. Therefore, PinX1 is
considered a telomerase inhibitor and tumor suppressor. Our study
indicated that increase of telomerase activity and hTERT protein
followed by decrease of PinX1 mRNA and protein occurred in the
carcinogenesis of esophageal epithelial cells and there was a
significantly negative correlation between PinX1 and tumor grade,
depth of infiltration and lymph node metastasis. From these
results, we deduced that the PinX1 gene may be used as one of the
molecular markers to determine the malignant degree, metastasis
potency and predicting progression of ESCC.
However, the mechanism of PinX1 functioning in tumor
cells has yet to be fully elucidated. Some studies indicate that
the PinX1 gene can inhibit telomerase activity and induce cell
apoptosis and expression of PinX1 is negatively correlated with
hTERT expression and telomerase activity in tumor cells. For
example, Lai et al(32)
reported that overexpression of PinX1 decreased hTERT mRNA by 21%,
reduced telomerase activity, inhibited cell growth, migration and
wound healing ability, arrested cells in the G0/G1 phase, and
increased AI. Zhang et al(33) reported that silencing the PinX1 gene
using short hairpin RNA can lead to significant shortening of
telomere and growth inhibition of telomerase-positive tumor cells,
but not telomerase-negative tumor cells, indicating PinX1 affects
telomere length and tumorigenicity through regulating telomerase
activity. Wang et al(34)
constructed and transfected PinX1 and PinX1-siRNA eukaryotic
expression vectors into gastric cancer cells and found that
downregulation of PinX1 significantly enhanced telomerase activity
compared with cells transfected with PinX1 vector, indicating that
PinX1 is a telomerase inhibitor and inhibits tumorigenesis and
development possibly through the telomerase/telomere pathway. To
better understand the role of PinX1 in esophageal cancer cells, we
also successfully established overexpression of PinX1, the
transfection of which significantly increased endogenous PinX1
protein in Eca109 cells. Moreover, PinX1 overexpression inhibited
telomerase activity and Eca109 cell growth, and blocked cells into
the G0/G1 stage, induced apoptosis, suggesting that PinX1 is a
telomerase inhibitor and inhibits tumorigenesis and development
possibly through the telomerase/telomere pathway.
Zhou and Lu (35)
considered that PinX1 inhibits telomerase activity by binding to
hTERT through its TID domain, which consequently results in
telomere shortening, cell senescence and increase of tumorigenicity
in nude mice. Banik and Counter (36) showed that inhibition of telomerase
activity by PinX1 requires its binding to both hTERT and hTR. In
the present study, we found that there were significantly negative
correlations between the expression of PinX1 and hTERT, telomerase
activity in carcinogenesis of esophageal epithelial cells,
respectively, thus we considered that the inhibition of PinX1 in
telomerase activity may contribute to downregulation of hTERT.
However, the precise mechanism requires further examination.
In summary, our data demonstrate that the length
shortening of telomere is an important characteristic in the
carcinogenesis of esophageal epithelial cells, followed by increase
of telomerase activity and downregulation of PinX1. Overexpression
of PinX1 blocked Eca109 cell proliferation and induced cell
apoptosis by downregulating the telomerase activity. However,
further studies are required to examine the precise role of PinX1
in telomerase activity in the pathogenesis of esophageal
cancer.
Acknowledgements
This study was supported by the Hebei Natural
Science Foundation (H2012206107, 062611136D-5).
References
|
1
|
Risques RA, Vaughan TL, Li X, et al:
Leukocyte telomere length predicts cancer risk in Barrett’s
esophagus. Cancer Epidemiol Biomarkers Prev. 16:2649–2655.
2007.
|
|
2
|
Xing J, Ajani JA, Chen M, et al:
Constitutive short telomere length of chromosome 17p and 12q but
not 11q and 2p is associated with an increased risk for esophageal
cancer. Cancer Prev Res (Phila). 2:459–465. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Cohen SB, Graham ME, Lovrecz GO, et al:
Protein composition of catalytically active human telomerase from
immortal cells. Science. 315:1850–1853. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pao CC, Tseng CJ, Lin CY, et al:
Differential expression of telomerase activity in human cervical
cancer and cervical intraepithelial neoplasia lesions. J Clin
Oncol. 15:1932–1937. 1997.PubMed/NCBI
|
|
5
|
Anderson S, Shera K, Ihle J, et al:
Telomerase activation in cervical cancer. Am J Pathol. 151:25–31.
1997.
|
|
6
|
Heselmeyer-Haddad K, Janz V, Castle PE, et
al: Detection of genomic amplification of the human telomerase gene
(TERC) in cytologic specimens as a genetic test for the diagnosis
of cervical dysplasia. Am J Pathol. 163:1405–1416. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Heselmeyer-Haddad K, Sommerfeld K, White
NM, et al: Genomic amplification of the human telomerase gene
(TERC) in pap smears predicts the development of cervical
cancer. Am J Pathol. 166:1229–1238. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Andersson S, Wallin KL, Hellström AC, et
al: Frequent gain of the human telomerase gene TERC at 3q26
in cervical adenocarcinomas. Br J Cancer. 95:331–338. 2006.
View Article : Google Scholar
|
|
9
|
Hopman A, Theelen W, Hommelberg P, et al:
Genomic integration of oncogenic HPV and gain of the human
telomerase gene TERC at 3q26 are strongly associated events in the
progression of uterine cervical dysplasia to invasive cancer. J
Pathol. 210:412–419. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sokolova I, Algeciras-Schimnich A, Song M,
et al: Chromosomal biomarkers for detection of human papillomavirus
associated genomic instability in epithelial cells of cervical
cytology specimens. J Mol Diagn. 9:604–611. 2007. View Article : Google Scholar
|
|
11
|
Caraway NP, Khanna A, Dawlett M, et al:
Gain of the 3q26 region in cervicovaginal liquid-based pap
preparations is associated with squamous intraepithelial lesions
and squamous cell carcinoma. Gynecol Oncol. 110:37–42. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hashimoto Y, Murakami Y, Uemura K, et al:
Detection of human telomerase reverse transcriptase (hTERT)
expression in tissue and pancreatic juice from pancreatic cancer.
Surgery. 143:113–125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Saha B, Chaiwun B, Tsao-Wei DD, et al:
Telomerase and markers of cellular proliferation are associated
with the progression of cervical intraepithelial neoplasia lesions.
Int J Gynecol Pathol. 26:214–222. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Branca M, Giorgi C, Ciotti M, et al:
Upregulation of telomerase (hTERT) is related to the grade of
cervical intraepithelial neoplasia, but is not an independent
predictor of high-risk human papillomavirus, virus persistence, or
disease outcome in cervical cancer. Diagn Cytopathol. 34:739–748.
2006. View
Article : Google Scholar
|
|
15
|
Kondo T, Oue N, Mitani Y, et al: Loss of
heterozygosity and histone hypoacetylation of the PINX1 gene are
associated with reduced expression in gastric carcinoma. Oncogene.
24:157–164. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ma Y, Wu L, Liu C, et al: The correlation
of genetic instability of PINX1 gene to clinico-pathological
features of gastric cancer in the Chinese population. J Cancer Res
Clin Oncol. 135:431–437. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liao C, Zhao MJ, Zhao J, et al:
Over-expression of LPTS-L in hepatocellular carcinoma cell line
SMMC-7721 induces crisis. World J Gastroenterol. 8:1050–1052.
2002.PubMed/NCBI
|
|
18
|
Liao C, Zhao M, Song H, et al:
Identification of the gene for a novel liver-related putative tumor
suppressor at a high-frequency loss of heterozygosity region of
chromosome 8p23 in human hepatocellular carcinoma. Hepatology.
32:721–727. 2000. View Article : Google Scholar
|
|
19
|
Sun J, Huang H, Zhu Y, et al: The
expression of telomeric proteins and their probable regulation of
telomerase during the differentiation of all-trans-retinoic
acid-responsive and -resistant acute promyelocytic leukemia cells.
Int J Hematol. 82:215–223. 2005. View Article : Google Scholar
|
|
20
|
Sun J, Tan YM, Huang H, et al: Expression
of telomerase inhibitor Pinx1 in leukemia cells and its correlation
with telomerase activity. Chinese J Pathophysiology. 22:1725–1728.
2006.
|
|
21
|
Hawkins GA, Chang BL, Zheng SL, et al:
Mutational analysis of PINX1 in hereditary prostate cancer.
Prostate. 60:298–302. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Akiyama Y, Maesawa C, Wada K, et al: Human
PinX1, a potent telomerase inhibitor, is not involved in human
gastrointestinal tract carcinoma. Oncol Rep. 11:871–874.
2004.PubMed/NCBI
|
|
23
|
Chang Q, Pang JC, Li J, et al: Molecular
analysis of PinX1 in medulloblastomas. Int J Cancer. 109:309–314.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yuan K, Li N, Jiang K, et al: PinX1 is a
novel microtubule-binding protein essential for accurate chromosome
segregation. J Biol Chem. 284:23072–23082. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hao J, Zhu L, Zhao S, et al: PTEN
ameliorates high glucose-induced lipid deposits through regulating
SREBP-1/FASN/ACC pathway in renal proximal tubular cells. Exp Cell
Res. 317:1629–1639. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Feng X, Hao J, Liu Q, et al: HMGB1
mediates IFN-γ-induced cell proliferation in MMC cells through
regulation of cyclin D1/CDK4/p16 pathway. J Cell Biochem.
113:2009–2019. 2012.
|
|
27
|
Wentzensen IM, Mirabello L, Pfeiffer RM
and Savage SA: The association of telomere length and cancer: a
meta-analysis. Cancer Epidemiol Biomarkers Prev. 20:1238–1250.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Klingelhutz AJ: The roles of telomeres and
telomerase in cellular immortalization and the development of
cancer. Anticancer Res. 19:4823–4830. 1999.PubMed/NCBI
|
|
29
|
Wang XW, Xiao J, Zhao S, et al: Expression
of telomerase subunits and its relationship with telomerase
activity in nasopharyngeal carcinoma. Zhonghua Yi Xue Za Zhi.
81:553–556. 2001.(In Chinese).
|
|
30
|
Liu H, Liu S, Wang H, et al: Genomic
amplification of the human telomerase gene (hTERC) associated with
human papillomavirus is related to the progression of uterine
cervical dysplasia to invasive cancer. Diagnostic Pathol.
7:1472012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Shen CX, Wen Z, Qian YH, et al: Targeted
gene therapy of nasopharyngeal cancer in vitro and in vivo by
enhanced thymidine kinase expression driven by human TERT promoter
and CMV enhancer. J Exp Clin Cancer Res. 29:942010. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lai XF, Shen CX, Wen Z, et al: PinX1
regulation of telomerase activity and apoptosis in nasopharyngeal
carcinoma cells. J Exp Clin Cancer Res. 31:122012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Zhang B, Bai YX, Ma HH, et al: Silencing
PinX1 compromises telomere length maintenance as well as
tumorigenicity in telomerase-positive human cancer cells. Cancer
Res. 69:75–83. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Wang HB, Wang XW, Zhou G, et al: PinX1
inhibits telomerase activity in gastric cancer cells through
Mad1/c-Myc pathway. J Gastrointest Surg. 14:1227–1234. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhou XZ and Lu KP: The
Pin2/TRF1-interacting protein PinX1 is a potent telomerase
inhibitor. Cell. 107:347–359. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Banik SS and Counter CM: Characterization
of interactions between PinX1 and human telomerase subunits hTERT
and hTR. J Biol Chem. 279:51745–51748. 2004. View Article : Google Scholar : PubMed/NCBI
|