Introduction
Chondrosarcoma is a type of bone cancer originating
from cartilaginous tissue and is the second most common bone tumor
(1). Chondrosarcomas respond poorly
to radiotherapy and chemotherapy, which are the currently used
treatment strategies for this malignancy, making the management of
chondrosarcomas a challenge (2).
Chondrosarcoma accounts for 25–30% of all cancers originating from
the skeletal system (3).
Osteoarthritis (OA), the most common joint disease, is a
functionally debilitating condition characterized by degeneration
of articular cartilage. Intrinsic resorption occurs when
chondrocytes, the only cells present in the articular cartilage,
over-exert their capacity for stimulating resorption of the
extracellular matrix (ECM) (4,5).
Articular chondrocytes maintain a dynamic equilibrium between
synthesis and degradation of ECM components, including type II
collagen fibrils surrounding and restraining large hydrated
aggregates of proteoglycans, allowing normal cartilage to function
as a shock absorber and withstand compressive loads (5). ECM degradation and remodeling require
the action of extracellular proteinases, of which matrix
metalloproteinases (MMPs) have been shown to play a significant
role (6). MMPs are a family of
zinc-dependent proteinases that facilitate the breakdown and
turnover of cartilage, and their levels are elevated in joint
tissues of patients with OA and rheumatoid arthritis (RA) (7,8).
Resveratrol (trans-3,4′,5-trihydroxystilbene)
is a natural polyphenolic compound present in various types of
plants, such as grapes and in food products. Its powerful and
diverse biological effects have been well documented in recent
years (5). Resveratrol exhibits
many biological activities, including anticancer,
anti-inflammatory, anti-aging, anti-oxidant, anti-bacterial,
antifungal and antiviral effects (9). Resveratrol has been shown to inhibit
MMPs via modulation of sirtuin-1 (SIRT-1) in human fibrosarcoma
cells. Notably, the activation of SIRT-1 in the presence of
resveratrol specifically inhibits the expression of MMP-9 in HT1080
cells (10). Recent studies have
demonstrated the chondroprotective effects and mechanisms of
resveratrol in chondrocytes stimulated with advanced glycation end
products, including a previous study, in which we delineated the
mechanisms underlying the resveratrol-mediated protection of
cartilage from advanced glycation end product-induced damage, and
addressed the potential therapeutic benefit of resveratrol in the
treatment of OA (11). Another
recent study focused on the effect of resveratrol on metastasis in
4T1 mouse breast cancer cells. Resveratrol inhibited cancer
metastasis both in vitro and in vivo, and this
inhibition was likely due to the reduction in MMP-9 activity caused
by resveratrol (12). Moreover,
resveratrol reduced lung adenocarcinoma cell metastasis by
suppressing heme oxygenase (HO)-1-mediated nuclear factor (NF)-κB
pathway activation and subsequently downregulated the expression of
MMPs. Resveratrol inhibited the transcription-activator function of
HO-1 and subsequently MMP-2 and MMP-9 expression in human lung
cancer cells as well. The inhibitory effects of resveratrol on MMP
expression and invasion of lung cancer cells were, in part, found
to be associated with its regulation of the HO-1-mediated effects
on the NF-κB pathway (13).
The mitogen-activated protein kinase (MAPK) pathway
is an evolutionarily conserved kinase pathway that links
extracellular signals to the machinery that controls fundamental
cellular processes such as proliferation, differentiation, invasion
and apoptosis in various types of cells, including cancer cells
(14). Among the MAPK subtypes, the
extracellular signal-regulated kinases (ERKs) are activated in
response to stimulation with mitogen or growth factor, whereas JNK
and p38 kinase are regulated under conditions of cellular stress
(15). A recent study demonstrated
that induction of MMP-2 and MMP-9 involves multiple signaling
cascades, particularly of the MAPK pathway (16). Other studies showed that epidermal
growth factor receptor (EGFR) regulates MMP function in fibroblasts
through the MAPK and AP-1 pathways (17) and that transforming growth factor
(TGF)-β-induced transcriptional activation of MMP-2 is mediated by
activating transcription factor (ATF)-2 in human breast epithelial
cells (18).
However, the molecular mechanism of MMP-regulated
differentiation of human chondrosarcoma cells by resveratrol
remains largely unclear. Therefore, in the present study, we sought
to investigate the mechanism through which resveratrol regulates
MMP-mediated differentiation, using HTB94 human chondrosarcoma
cells as a model. We also sought to identify the underlying
signaling pathways involved in this process. Our results indicate
that resveratrol suppresses MMP-regulated differentiation through
its effects on the p38 kinase and JNK pathways in HTB94 human
chondrosarcoma cells.
Materials and methods
Materials
Resveratrol (trans-3,4′,5-trihydroxystilbene)
was obtained from Sigma Aldrich (St. Louis, MO, USA). Dulbecco’s
modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were
purchased from Invitrogen (Burlington, ON, Canada). Streptomycin
and penicillin were purchased from Sigma Aldrich. SB203580,
SP600125 and MMP inhibitor III were obtained from Calbiochem (San
Diego, CA, USA). Antibodies specific for MMP-2 (Santa Cruz
Biotechnology, Santa Cruz, CA, USA), MMP-9 (Santa Cruz
Biotechnology), type II collagen (Santa Cruz Biotechnology), SOX-9
(Santa Cruz Biotechnology), phosphorylated (p)p38 (Cell Signaling
Technology, Danvers, MA, USA), p38 (Santa Cruz Biotechnology), pJNK
(Cell Signaling Technology), JNK (Santa Cruz Biotechnology) and
actin (Santa Cruz Biotechnology) were used for the experiments.
Cell lines and culture
The HTB94 (human chondrosarcoma) cell line was
purchased from the American Type Culture Collection (ATCC;
Rockville, MD, USA). HTB94 cells were maintained in DMEM containing
10% FBS, 50 μg/ml streptomycin and 50 U/ml penicillin. Cells were
maintained at 37°C in a humidified atmosphere with 5%
CO2. HTB94 cells were then plated on culture dishes at a
density of 0.9×105 cells/dish, and the cells were
allowed to reach 70% confluency before they were treated with
inhibitors or used in the experiments.
Viability assay (MTT)
HTB94 cells were plated at a density of
0.9×105 cells/well on 96-well plates and incubated
overnight, and the medium was replaced with fresh medium on the
following day. The cells were treated with various concentrations
of resveratrol or left untreated, in the absence or presence of
SB203580, SP600125 or MMP inhibitor III for 24 h. After these
treatments, 10 μl of MTT reagent I (methyl thiazole tetrazolium)
(10 mg/ml) was added to each well. After incubating the cells for
another 24 h, 100 μl of MTT reagent II [solubilization buffer, 10%
SDS with 0.01 N HCl in dimethylsulfoxide (DMSO)] was added to each
well, and the cells were incubated overnight at 37°C. Finally, the
absorbance of the samples was measured at 595 nm by using an
enzyme-linked immunosorbent assay (ELISA) plate reader.
Gelatin zymography
HTB94 cells were seeded on a 35-mm tissue culture
dish and kept overnight for attachment. The next day, the medium
was replaced with DMEM containing 2% FBS, and the cells were
incubated overnight at 37°C. HTB94 cells were treated with the
indicated reagents in DMEM containing 2% FBS. The cells were lysed
to prepare protein extracts, and equal amounts of total protein
were resolved by sodium dodecyl sulfate (SDS)-polyacrylamide gel
electrophoresis (PAGE) on 7.5% polyacrylamide gels containing 0.1%
gelatin. After SDS-PAGE, the gels were washed with 2.5% Triton
X-100 for 90 min and incubated with gelatin incubation buffer (5 mM
CaCl2, 0.2 M NaCl and 50 mM Tris) for 20–24 h. Next, the
gels were stained with Coomassie blue solution for 60–90 min and
destained with a solution containing 30% acetic acid and 10%
methanol. The bands were quantified by densitometric analysis using
the ImageJ software package.
Western blot analysis
HTB94 cells grown in 35-mm tissue culture dishes
were treated with the indicated reagent, harvested and washed with
cold phosphate-buffered saline (PBS). Proteins were extracted using
cold radioimmunoprecipitation assay lysis buffer [50 mM Tris-HCl
(pH 7.4), 150 mM NaCl, 1% Nonidet P-40, 0.1% SDS, supplemented with
protease inhibitors (10 μg/ml aprotinin, 10 μg/ml pepstatin, 10
μg/ml leupeptin, and 1 mM 4-(2-aminoethyl) benzenesulfonyl
fluoride) and phosphatase inhibitors (1 mM NaF and 1 mM sodium
orthovanadate)]. Equal amounts of total cellular proteins were
resolved by SDS-PAGE, and the proteins were transferred to
nitrocellulose membranes (Whatman Schleicher and Schuell, Dachen,
Germany). The membranes were blocked with 5% non-fat dry milk in
Tris-buffered saline (TBS). The membranes were incubated with
antibodies specific for MMP-2, MMP-9, type II collagen, SOX-9,
pp38, p38, pJNK, JNK and actin overnight at 4°C. The membranes were
than washed with TBST (TBS containing Tween-20) and incubated with
horseradish peroxidase-conjugated secondary antibodies (Sigma
Aldrich) for 2 h. The bands were quantified by densitometric
analysis using the ImageJ software package.
Alcian blue staining
HTB94 cells were fixed with 95% methanol at −20°C
for 5 min. Next, the cells were stained with 0.1% Alcian blue in
0.1 M HCl overnight. The cells were washed 3 times with PBS and
incubated with 6 M guanidine HCl for 6 h. Production of sulfated
proteoglycans was evaluated by measuring the absorbance of the
stained cells at 620 nm using an ELISA plate reader.
Immunofluorescence (IF) analysis
HTB94 cells were fixed with cold 3.5%
paraformaldehyde in PBS for 20 min at room temperature. Cells were
permeabilized in PBS containing 0.1% Triton X-100 for 15 min at
room temperature. The fixed cells were subsequently washed with PBS
and incubated for 2 h with antibodies against MMP-9 and type II
collagen. Then, the cells were washed with PBS and incubated with
secondary antibodies for 1 h, after which the cells were washed
again and incubated for 15 min with DAPI (Invitrogen). The cells
were washed 3 times with PBS and observed under a fluorescence
microscope.
Statistical analysis
The results are expressed as the means ± standard
deviation (SD). The significance of differences between
experimental and control groups was assessed using one-way analysis
of variance (ANOVA). Significant differences were defined at the
level of P<0.05.
Results
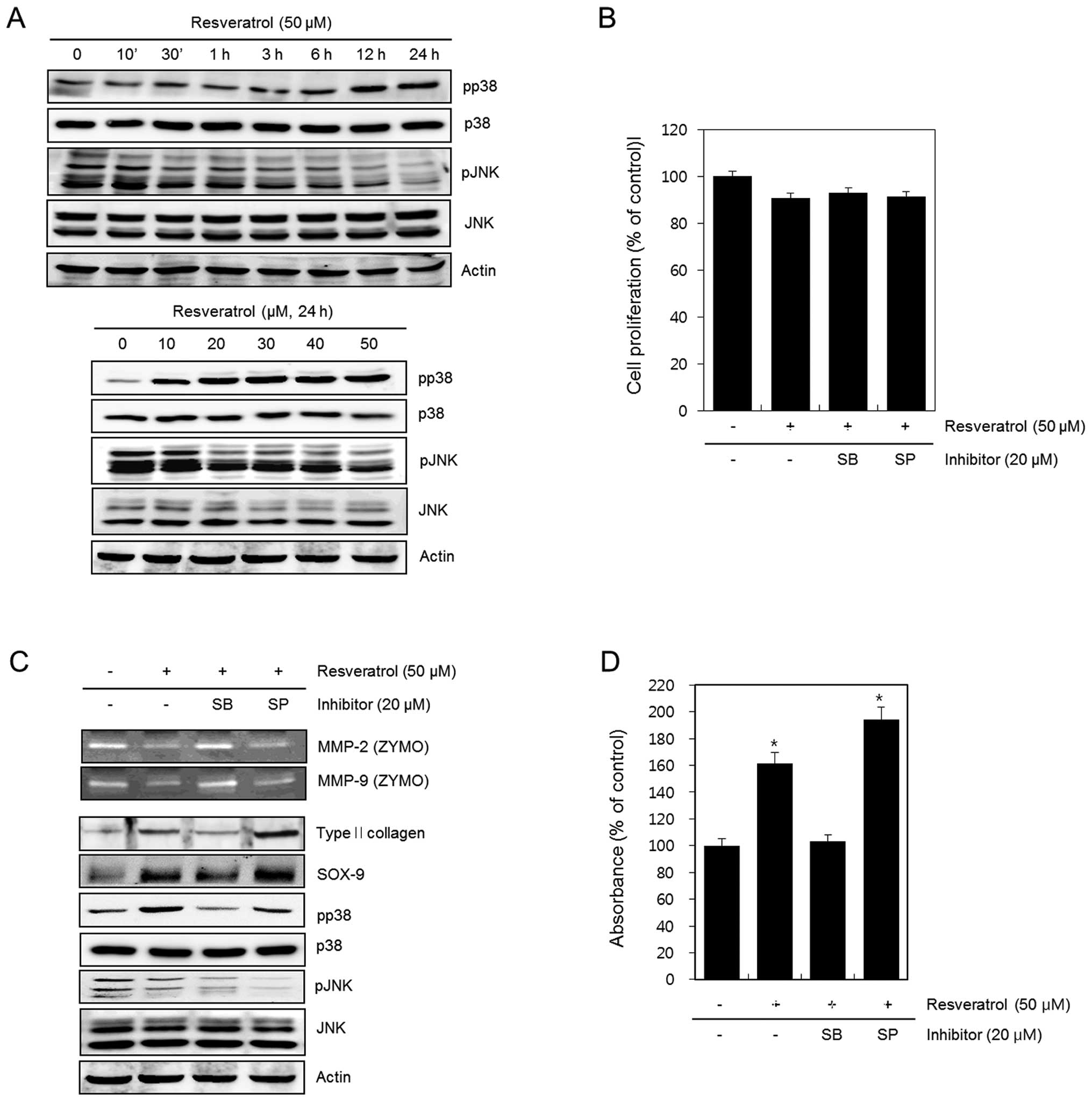
Resveratrol reduces activation of MMPs in
human chondrosarcoma HTB94 cells
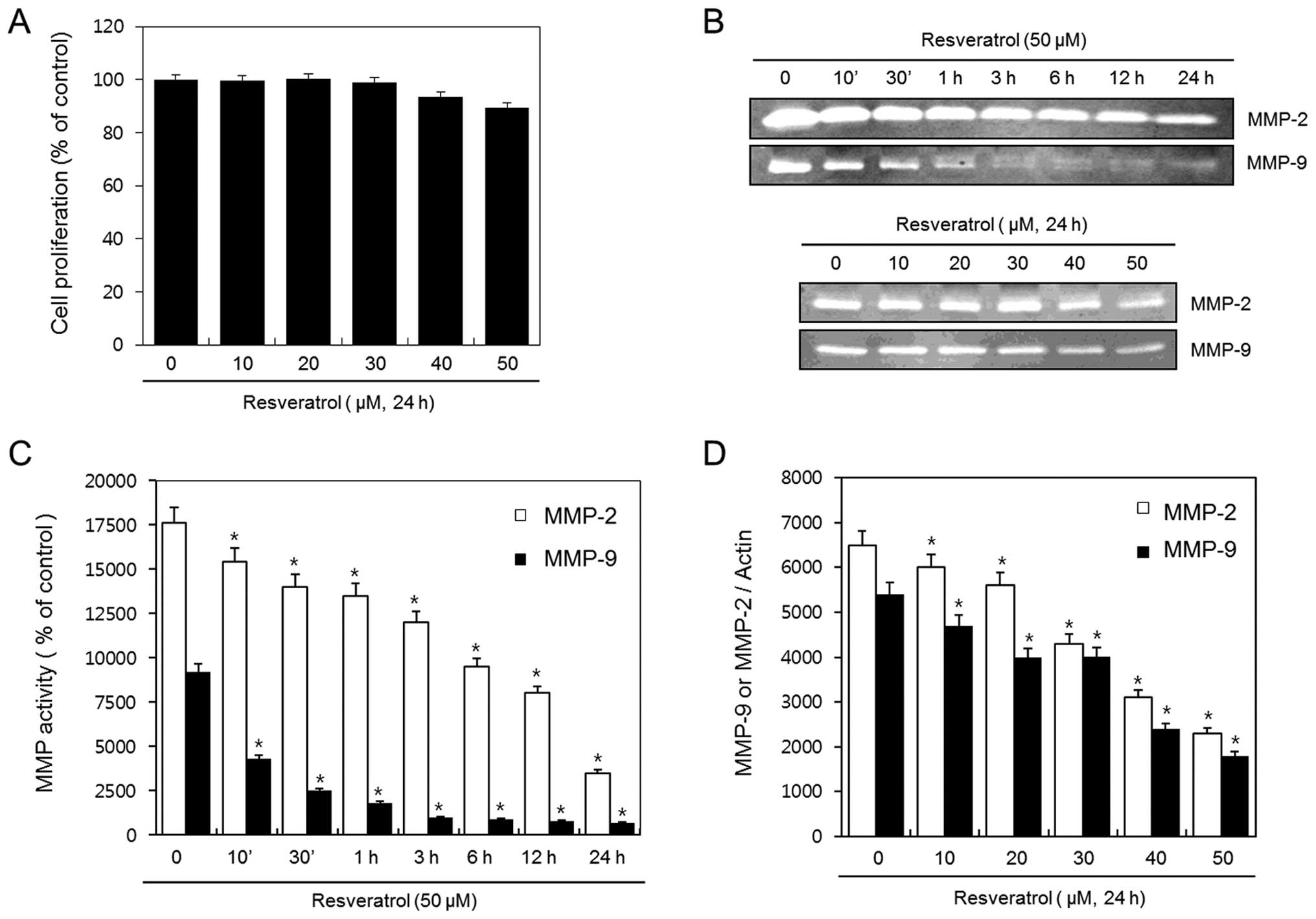
To determine the cytotoxic effects of resveratrol on
HTB94 cells, we treated cells with 10, 20, 30, 40 and 50 μM of
resveratrol for 24 h and evaluated the cell viability using the MTT
assay. The results showed that treatment with resveratrol did not
affect the viability of HTB94 cells (Fig. 1A). Next, HTB94 cells were left
untreated (control) or treated with 50 μM resveratrol for the
indicated time periods or treated with the indicated concentrations
of resveratrol for 24 h. The activation of MMP-2 and MMP-9 was then
analyzed using gelatin zymography. Resveratrol markedly reduced
activation of MMP-2 and MMP-9 in a time- and dose-dependent manner
(Fig. 1B). The zymography data were
quantified by densitometric analyses using ImageJ (Fig. 1C and D). These results suggest that
resveratrol reduces activation of MMP-2 and MMP-9 in HTB94
cells.
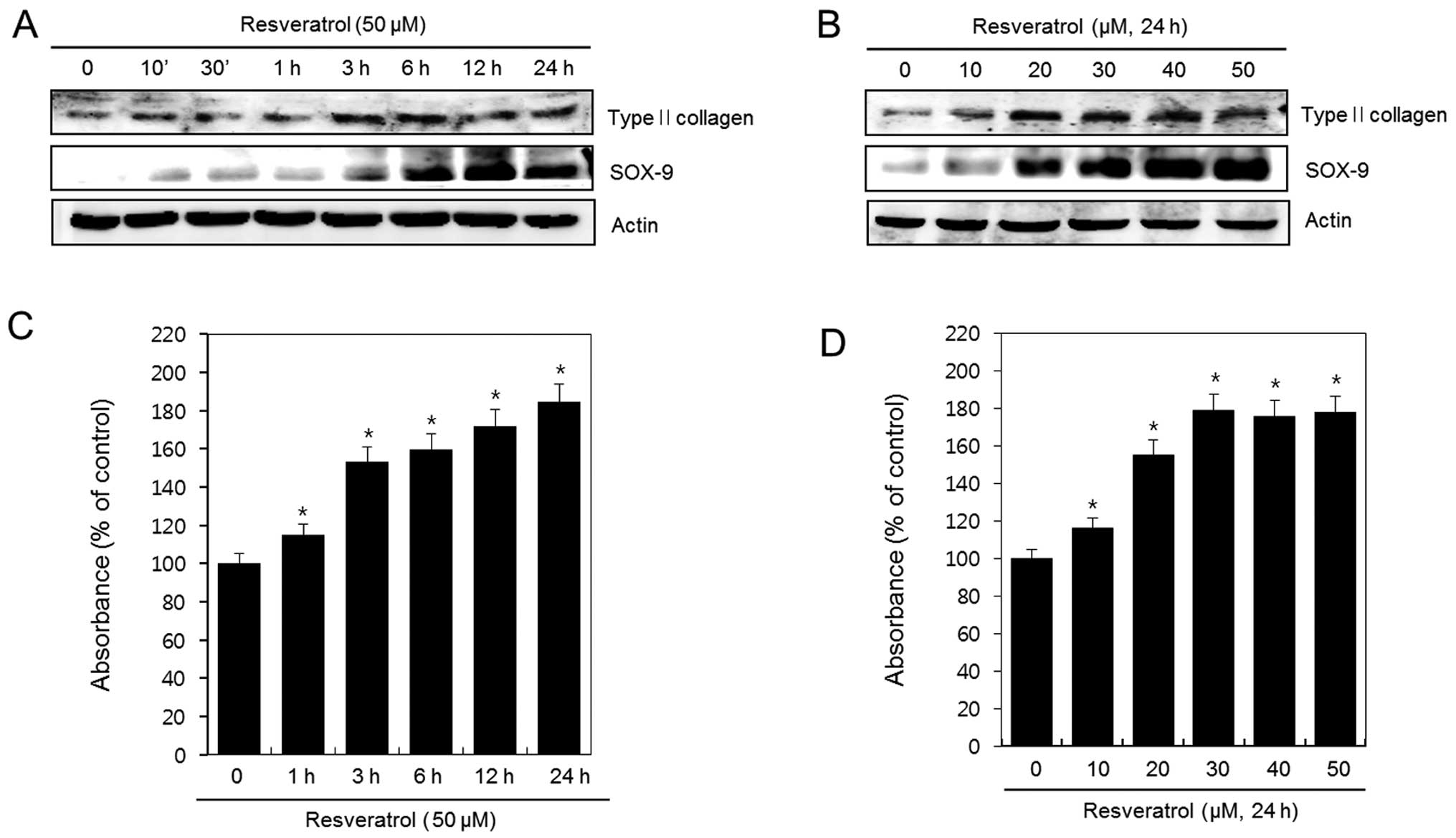
Resveratrol induces differentiation in
human chondrosarcoma HTB94 cells
HTB94 cells were left untreated (control) or treated
with 50 μM of resveratrol for the indicated time periods or treated
with the indicated various concentrations of resveratrol for 24 h.
The expression of type II collagen, SOX-9 and actin was detected
using western blotting. Resveratrol induced a significant increase
in the expression of type II collagen and SOX-9 in a time- and
dose-dependent manner (Fig. 2A and
B). Production of sulfated proteoglycans was analyzed using
Alcian blue staining. Consistent with the expression patterns of
type II collagen, resveratrol induced the production of sulfated
proteoglycans (Fig. 2C and D).
These data suggest that resveratrol increases expression of type II
collagen and SOX-9 and the production of sulfated proteoglycan in
HTB94 chondrosarcoma cells. Taken together, these results indicate
that resveratrol induces differentiation in HTB94 cells.
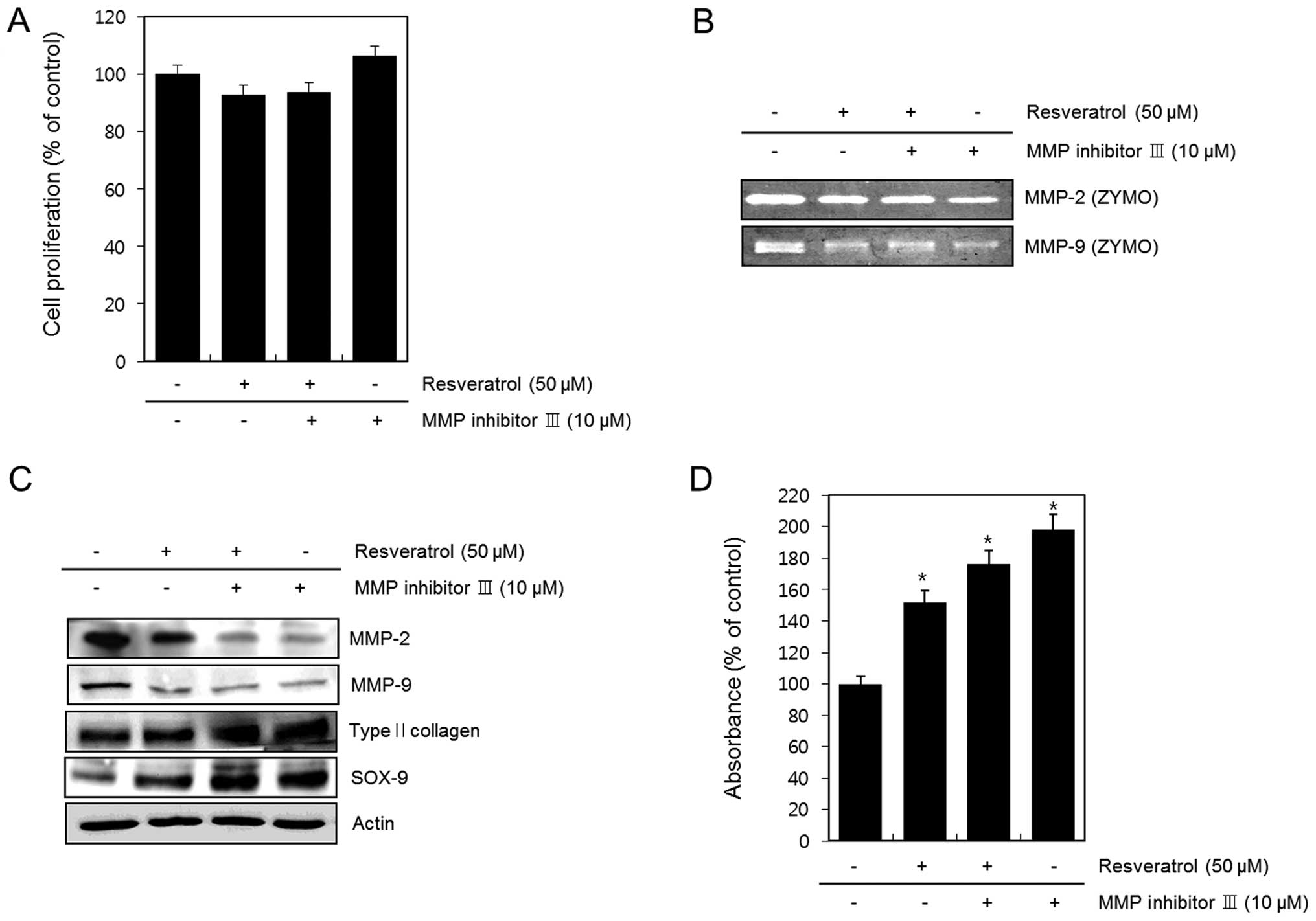
Resveratrol-induced differentiation
occurs via its effects on MMPs
We left HTB94 cells untreated (control) or treated
them with resveratrol in the absence or presence of 10 μM MMP
inhibitor III for 24 h. To evaluate the cytotoxic effects on cell
viability, we performed an MTT assay. The results indicated that
co-treatment with resveratrol and MMP inhibitor III did not affect
the viability of HTB94 cells (Fig.
3A). Activation of MMP-2 and MMP-9 was assessed using gelatin
zymography. We found that MMP inhibitor III suppressed the activity
of MMP-2 and MMP-9, and that the extent of this suppression was
greater in the presence of resveratrol (Fig. 3B). Expression of MMP-2, MMP-9, type
II collagen, SOX-9 and actin was assessed by western blot analysis.
Inhibition of MMP-2 and MMP-9 with MMP inhibitor III further
increased the resveratrol-induced expression of type II collagen
and SOX-9 (Fig. 3C). Consistent
with the expression patterns of type II collagen, inhibition of
MMP-2 and MMP-9 with MMP inhibitor III further enhanced the
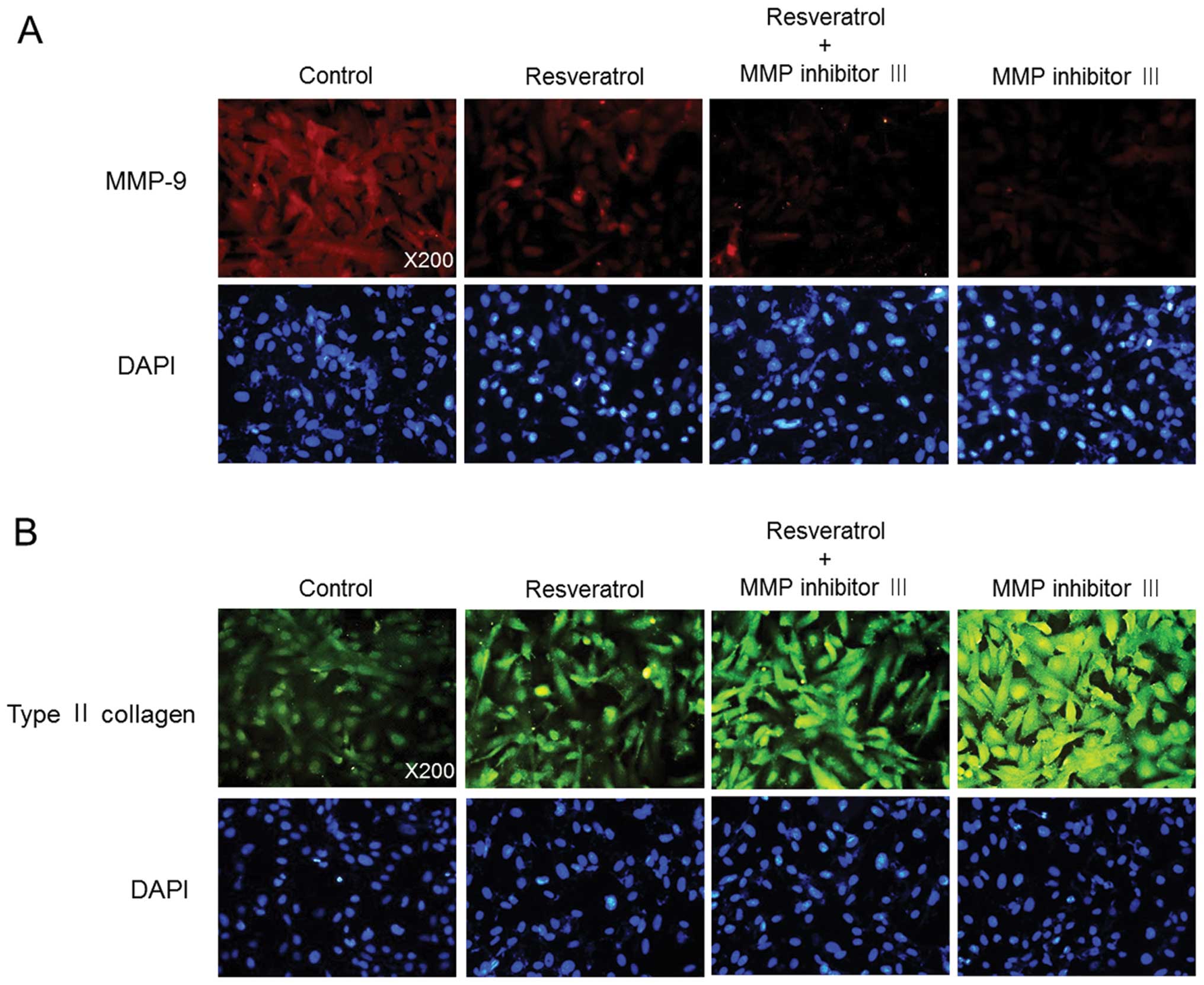
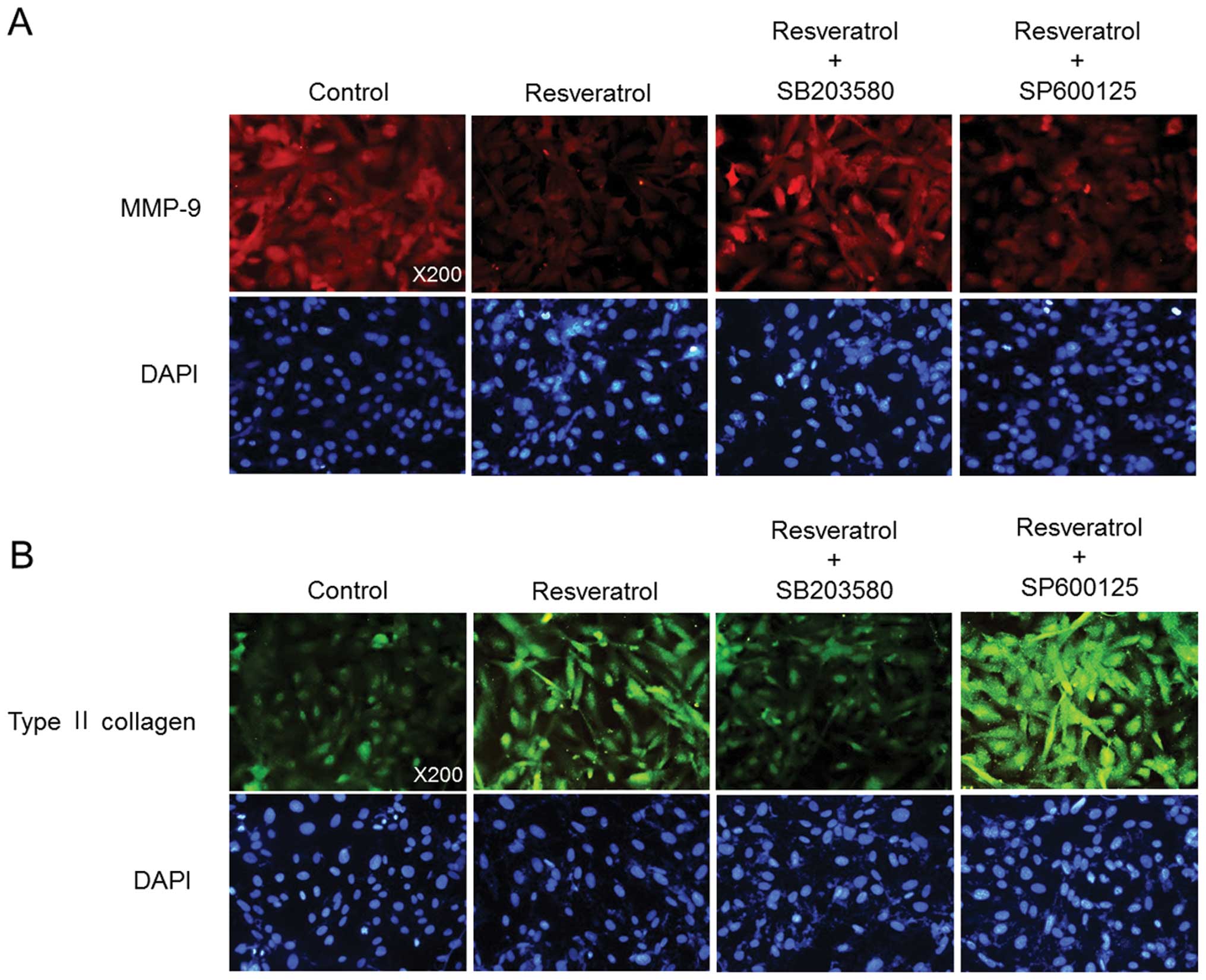
resveratrol-induced production of sulfated proteoglycans (Fig. 3D). Moreover, consistent with the
western blotting data, immunofluorescence analysis showed that
treatment with MMP inhibitor III enhanced the resveratrol-mediated
suppression of MMP-9 expression and increase in type II collagen
expression (Fig. 4).
These data suggest that resveratrol decreases MMP-2-
and MMP-9-regulated expression of type II collagen and SOX-9, and
the production of sulfated proteoglycan in HTB94 chondrosarcoma
cells. Taken together, our results indicate that resveratrol
reduces MMP-regulated differentiation in HTB94 cells.
Resveratrol reduces MMP-regulated differentiation
via p38 kinase and JNK pathways in HTB94 cells. We sought to
identify the upstream signaling pathways involved in the
resveratrol-mediated suppression of MMP-regulated differentiation
in chondrosarcoma cells. HTB94 cells were left untreated (control),
treated with 50 μM of resveratrol for the indicated time periods or
treated with the various concentrations of resveratrol for 24 h.
The expression of pp38, p38, pJNK, JNK and actin was analyzed using
western blotting. The results showed that resveratrol markedly
induced the phosphorylation of p38 and reduced the phosphorylation
of JNK in a time- and dose-dependent manner (Fig. 5A). We left HTB94 cells untreated
(control) or treated them with resveratrol in the absence or
presence of 20 μM SB203580 or 20 μM SP600125 for 24 h. To determine
the effect of resveratrol on the viability of HTB94 cells, we
performed an MTT assay. We found that co-treatment with resveratrol
and SB203580 or SP600125 did not affect the viability of HTB94
cells (Fig. 5B). Next, activation
of MMP-2 and MMP-9 was detected using gelatin zymography, and
expression of type II collagen, SOX-9, pp38, pJNK and actin was
assessed using western blotting. Inhibition of p38 kinase with
SB203580 enhanced the effects of resveratrol on MMP-2 and MMP-9
expression, as well as its effects on the expression of type II
collagen and SOX-9. Similar effects were observed upon co-treatment
of cells with resveratrol and the JNK inhibitor SP600125 (Fig. 5C). Moreover, resveratrol-mediated
production of sulfated proteoglycans, as assessed using Alcian blue
staining, was suppressed in cells co-treated with the p38 kinase
inhibitor SB203580 and resveratrol; inhibition of JNK with SP600125
in the presence of resveratrol further increased the
resveratrol-induced production of sulfated proteoglycans (Fig. 5D). Moreover, consistent with the
results of western blotting, the results of immunofluorescence
analysis showed that inhibition of p38 kinase with SB203580
enhanced the suppression of MMP-9 by resveratrol and inhibited
resveratrol-induced type II collagen expression (Fig. 6A). Inhibition of JNK kinase with
SP600125 further increased the resveratrol-induced type II collagen
expression (Fig. 6B).
Taken together, these results suggest that
resveratrol attenuates MMP-regulated differentiation via the p38
kinase and JNK pathways in HTB94 chondrosarcoma cells.
Discussion
Resveratrol is a grape polyphenol, and is known to
have anti-cancer, anti-oxidant, anti-inflammatory and
neuroprotective activities. Resveratrol has been shown to prevent
angiogenesis and cell migration in endothelial cells, and these
properties of resveratrol make it a good candidate for use in the
prevention of tumor progression (19). Resveratrol was recently shown to
inhibit tumor cell adhesion to endothelial cells by blocking the
expression of intracellular adhesion molecule-1 (20), and the resveratrol analog
4-[2-(3,5-dimethoxyphenyl)vinyl] pyridine was found to reduce the
extent of differentiation of 3T3-L1 adipocytes (21).
Chondrosarcoma is a highly malignant cancer with
high potential to invade locally and cause distant metastasis, with
a predilection for metastasis to the lungs (22). Clinically, surgical resection
remains the primary mode of treatment for chondrosarcoma. In the
absence of effective adjuvant treatment, this mesenchymal
malignancy has a poor prognosis, and therefore, it is important to
explore novel and more effective therapeutic strategies (23). MMPs constitute a family of enzymes
that enable degradation of cartilage. These proteolytic enzymes
attack and degrade components of the ECM. Importantly, they
contribute to the breakdown of type II collagen and other matrix
proteins (4). Type II collagen is a
major component of cartilage, where it is present at a high
concentration that is essential for the functional maintenance of
tissue (24). Moreover, in
carcinomas, stromal cells participate in cancer progression by
producing proteases such as MMPs. The expression of MMPs is a
prognostic factor in human chondrosarcomas (25). Recent studies have demonstrated that
endothelin-1 promotes MMP-13 production and migration in human
chondrosarcoma cells through the focal adhesion kinase
(FAK)/phosphatidylinositol 3-kinase (PI3K)/Akt/mammalian target of
rapamycin (mTOR) pathways. These data suggest that endothelin-1
activates FAK/PI3K/AKT/mTOR signaling, which in turn activates IκB
kinase α/β and NF-κB, resulting in increased MMP-13 expression and
the migration of human chondrosarcoma cells (26). Interleukin (IL)-1β induces MMP-13
expression by activation of the p38 MAPK/c-Fos/AP-1 and Janus
kinase (JAK)/signal transducers and activators of transcription
(STAT) pathways in chondrocytes. The p38 MAPK/c-Fos/AP-1 and
JAK2/STAT1/2 pathways are involved in MMP-13 induction in
IL-1β-treated HTB94 human chondrocytes. Blocking these signaling
pathways may have chondroprotective effects in cartilage
degeneration (27). Another study
demonstrated downregulation of MMP-13 by the root extract of
Cyathula officinalis Kuan and its constituents in
IL-1β-treated chondrocytes. The results also showed that C.
officinalis Kuan constituents may have the potential for
conferring protection against cartilage-degrading disorders
(28). Moreover, CCN3, a
cysteine-rich protein family, enhanced the migration of
chondrosarcoma cells by increasing MMP-13 expression via the
αvβ3/αvβ5 integrin receptor, FAK, PI3K, Akt, p65 and NF-κB
signal-transduction pathways (29).
In the present study, we showed that resveratrol
suppressed the activation of MMP-2 and MMP-9 in HTB94 human
chondrosarcoma cells. Moreover, resveratrol induced the expression
of type II collagen and SOX-9 and sulfated proteoglycan production.
Inhibition of MMP-2 and MMP-9 with MMP inhibitor III further
enhanced these effects of resveratrol. In addition, resveratrol
markedly induced phosphorylation of p38 and reduced phosphorylation
of JNK in HTB94 cells. Inhibition of p38 kinase with SB203580
enhanced resveratrol-mediated suppression of MMP-2 and MMP-9 and
inhibited resveratrol-induced type II collagen and SOX-9
expression. Inhibition of JNK with SP600125 decreased
resveratrol-mediated suppression of MMP-2 and MMP-9, and further
increased resveratrol-induced effects on type II collagen and
SOX-9, and on the production of sulfated proteoglycans. In
conclusion, the results of the present study suggest that
resveratrol reduces the MMP-regulated differentiation of human
chondrosarcoma cells via the p38 kinase and JNK pathways.
Acknowledgements
The present study was supported by a grant from the
Korean Health Technology Research and the Development Project,
Ministry of Health and Welfare, Republic of Korea
(A120960-1201-0000300).
References
|
1
|
Zyada MM and Shamaa AA: Is collagenase-3
(MMP-13) expression in chondrosarcoma of the jaws a true marker for
tumor aggressiveness? Diagn Pathol. 3:262008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tang CH and Tsai CC: CCL2 increases MMP-9
expression and cell motility in human chondrosarcoma cells via the
Ras/Raf/MEK/ERK/NF-κB signaling pathway. Biochem Pharmacol.
83:335–344. 2012.PubMed/NCBI
|
|
3
|
Power PF, Mak IW, Singh S, Popovic S,
Gladdy R and Ghert M: ETV5 as a regulator of matrix
metalloproteinase 2 in human chondrosarcoma. J Orthop Res.
31:493–501. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lu YC, Jayakumar T, Duann YF, et al:
Chondroprotective role of sesamol by inhibiting MMPs expression via
retaining NF-κB signaling in activated SW1353 cells. J Agric Food
Chem. 59:4969–4978. 2011.PubMed/NCBI
|
|
5
|
Im HJ, Li X, Chen D, et al: Biological
effects of the plant-derived polyphenol resveratrol in human
articular cartilage and chondrosarcoma cells. J Cell Physiol.
227:3488–3497. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hidalgo M and Eckhardt SG: Development of
matrix metalloproteinase inhibitors in cancer therapy. J Natl
Cancer Inst. 93:178–193. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Abeles AM and Pillinger MH: The role of
the synovial fibroblast in rheumatoid arthritis: cartilage
destruction and the regulation of matrix metalloproteinases. Bull
NYU Hosp Jt Dis. 64:20–24. 2006.PubMed/NCBI
|
|
8
|
Tchetverikov I, Lohmander LS, Verzijl N,
et al: MMP protein and activity levels in synovial fluid from
patients with joint injury, inflammatory arthritis, and
osteoarthritis. Ann Rheum Dis. 64:694–698. 2005. View Article : Google Scholar
|
|
9
|
Brisdelli F, D’Andrea G and Bozzi A:
Resveratrol: a natural polyphenol with multiple chemopreventive
properties. Curr Drug Metab. 10:530–546. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee SJ and Kim MM: Resveratrol with
antioxidant activity inhibits matrix metalloproteinase via
modulation of SIRT1 in human fibrosarcoma cells. Life Sci.
88:465–472. 2009.
|
|
11
|
Liu FC, Hung LF, Wu WL, et al:
Chondroprotective effects and mechanisms of resveratrol in advanced
glycation end products-stimulated chondrocytes. Arthritis Res Ther.
12:R1672010. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee HS, Ha AW and Kim WK: Effect of
resveratrol on the metastasis of 4T1 mouse breast cancer cells
in vitro and in vivo. Nutr Res Pract. 6:294–300.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu PL, Tsai JR, Charles AL, et al:
Resveratrol inhibits human lung adenocarcinoma cell metastasis by
suppressing heme oxygenase 1-mediated nuclear factor-κB pathway and
subsequently downregulating expression of matrix
metalloproteinases. Mol Nutr Food Res. 54(Suppl 2): S196–S204.
2010.PubMed/NCBI
|
|
14
|
Dhillon AS, Hagan S, Rath O and Kolch W:
MAP kinase signalling pathways in cancer. Oncogene. 26:3279–3290.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kim SJ, Ju JW, Oh CD, et al: ERK-1/2 and
p38 kinase oppositely regulate nitric oxide-induced apoptosis of
chondrocytes in association with p53, caspase-3, and
differentiation status. J Biol Chem. 277:1332–1339. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cohen M, Meisser A, Haenggeli L and
Bischof P: Involvement of MAPK pathway in TNF-α-induced MMP-9
expression in human trophoblastic cells. Mol Hum Reprod.
12:225–232. 2006.
|
|
17
|
Kajanne R, Miettinen P, Mehlem A, et al:
EGF-R regulates MMP function in fibroblasts through MAPK and AP-1
pathways. J Cell Physiol. 212:489–497. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kim ES, Sohn YW and Moon A: TGF-β-induced
transcriptional activation of MMP-2 is mediated by activating
transcription factor (ATF) 2 in human breast epithelial cells.
Cancer Lett. 252:147–156. 2007.
|
|
19
|
Azios NG and Dharmawardhane SF:
Resveratrol and estradiol exert disparate effects on cell
migration, cell surface actin structures, and focal adhesion
assembly in MDA-MB-231 human breast cancer cells. Neoplasia.
7:128–140. 2005. View Article : Google Scholar
|
|
20
|
Park JS, Kim KM, Kim MH, et al:
Resveratrol inhibits tumor cell adhesion to endothelial cells by
blocking ICAM-1 expression. Anticancer Res. 29:355–362.
2009.PubMed/NCBI
|
|
21
|
Hwang JT, Kim S, Choi I and Choi SY:
Resveratrol analog 4-[2-(3,5-dimethoxyphenyl)vinyl]pyridine reduces
differentiation of the 3T3-L1 adipocyte. Pharm Biol. 51:96–99.
2013.
|
|
22
|
Hou CH, Chiang YC, Fong YC and Tang CH:
WISP-1 increases MMP-2 expression and cell motility in human
chondrosarcoma cells. Biochem Pharmacol. 81:1286–1295. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tang CH, Chen CF, Chen WM and Fong YC:
IL-6 increases MMP-13 expression and motility in human
chondrosarcoma cells. J Biol Chem. 286:11056–11066. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Shinomura T, Ito K, Hook M and Kimura JH:
A newly identified enhancer element responsible for type II
collagen gene expression. J Biochem. 152:565–575. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jawad MU, Garamszegi N, Garamszegi SP, et
al: Matrix metalloproteinase 1: role in sarcoma biology. PLoS One.
5:e142502010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wu MH, Lo JF, Kuo CH, et al: Endothelin-1
promotes MMP-13 production and migration in human chondrosarcoma
cells through FAK/PI3K/Akt/mTOR pathways. J Cell Physiol.
227:3016–3026. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lim H and Kim HP: Matrix
metalloproteinase-13 expression in IL-1β-treated chondrocytes by
activation of the p38 MAPK/c-Fos/AP-1 and JAK/STAT pathways. Arch
Pharm Res. 34:109–117. 2011.
|
|
28
|
Park HY, Lim H, Kim HP and Kwon YS:
Downregulation of matrix metalloproteinase-13 by the root extract
of Cyathula officinalis Kuan and its constituents in
IL-1β-treated chondrocytes. Planta Med. 77:1528–1530. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Tzeng HE, Chen JC, Tsai CH, et al: CCN3
increases cell motility and MMP-13 expression in human
chondrosarcoma through integrin-dependent pathway. J Cell Physiol.
226:3181–3189. 2011. View Article : Google Scholar : PubMed/NCBI
|