Introduction
Gastric carcinoma is one of the most common causes
of cancer-related mortality worldwide; however, there is limited
effective clinical treatment for this highly malignant tumor
besides surgery and chemotherapy. Furthermore, the treatment
outcome remains unsatisfactory as early diagnosis of gastric cancer
remains difficult and most patients have already developed
metastatic lesions when diagnosed (1). Therefore, it is necessary to search
novel agents to treat stomach cancer patients with adverse
effects.
An increasing number of publications indicate that
vitamin E succinate (VES), a natural derivative of vitamin E,
exhibits powerful anticancer effects in a variety of in vivo
and in vitro cancer models (2–7). Our
previous studies showed that VES induced apoptosis in SGC-7901
human gastric cancer cells via multiple signaling pathways,
including extrinsic Fas, MAP kinase, and endoplasmic reticulum (ER)
stress (8–10).
The relationship between the dysfunction of the
NF-κB signaling pathway and the carcinogenesis of stomach cancer
was previously reported by several research groups (11,12).
NF-κB is a ubiquitous dimeric transcription factor that plays
pivotal roles in regulating the expression of a multitude of
critical genes that regulate cell survival, proliferation,
apoptosis, immune responses, and adaptive responses to change the
intracellular redox balance (13–16).
The constitutive activation of NF-κB, a multi-function nuclear
factor, has been suggested to be a hallmark of highly malignant
tumors (17,18). Therefore, the inhibition of NF-κB
activation has been reported to be a naturally useful strategy for
increasing tissue sensitivity towards cytostatic drug treatment
in vitro and in vivo (19).
Materials and methods
Materials and reagents
Human medium-differentiated gastric cancer SGC-7901
cells were obtained from the Cancer Research Institute of Beijing.
VES, ethylenediaminetetraacetic acid, dimethyl sulfoxide (DMSO),
MTT, Tween-20, sodium dodecyl sulfate (SDS) and PDTC were purchased
from Sigma. RPMI-1640, fetal bovine serum (FBS), TRIzol reagent, an
Annexin V-FITC apoptosis assay kit, and a total-RNA isolation kit
were obtained from Gibco Chemical Co. (Gibco, Rockville, MD, USA).
The GeneAmp RNA PCR kit was provided by Perkin-Elmer Life Sciences
(Boston, MA, USA). The enhanced chemiluminescence (ECL) kit and the
horseradish peroxidase-conjugated antibody were obtained from
Amersham Life Science Inc. Taq polymerase was purchased from Roche
Molecular Biochemicals (Basel, Switzerland). The LightShift
Chemiluminescence EMSA kit was obtained from Pierce (Rockford, IL,
USA). Nuclear and cytoplasmic protein extraction kits and a BCA
protein assay kit were obtained from Viagene Biotech (Ningbo,
China). Rabbit polyclonal antibodies for Bcl-2, Bax, Bak,
caspase-8, caspase-9, caspase-3, PARP, NF-κBp65, TATA binding
protein (TBP) and β-actin were purchased from Santa Cruz
Biotechnology, Inc. Gene primers of c-IAP1, c-IAP-2, NAIP,
survivin, XIAP, and glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) for RT-PCR were constructed by Invitrogen.
Cell culture
SGC-7901 were cultured in RPMI-1640 medium
supplemented with 5% FBS, 2 mM glutamine, 100 μg/ml streptomycin,
100 IU/ml penicillin, and 20 μM sodium bicarbonate at 37°C in a
humidified atmosphere containing 5% CO2. The culture
medium was changed every 2 days.
Cell viability
Cells were plated in 96-well plates, allowed to
attach overnight, and treated with various concentrations of VES
(0–160 μM). Cell viability was estimated using the MTT assay. The
medium was aspirated and DMSO was used to dissolve the crystals.
Absorbance was measured at 570 nm using a microplate reader.
Assessment of apoptosis
Apoptosis was quantified using the Annexin V-FITC
method. Cells were treated with VES (40 μM), and were plated at a
seeding density of 105 per well in 24-well plates and
incubated overnight. Floating and attached cells were harvested and
treated according to the manufacturer’s instructions.
Protein extraction and western blot
analysis
After treatment with or without VES or PDTC, the
cells were collected and proteins were extracted. The protein
concentration was determined by DC Bio-Rad assay according to the
manufacturer’s protocol (Bio-Rad Laboratories, Hercules, CA,
USA).
For western blot analysis, appropriate amounts of
cell lysates (25–50 μg of protein) were separated on 10–15%
SDS-PAGE gel and transferred onto nitrocellulose membranes. The
membranes were blocked using 5% nonfat dry milk and probed using
appropriate primary antibodies in blocking buffer overnight at 4°C.
The membranes were then incubated with appropriate secondary
horseradish peroxidase-conjugated antibodies and detected by
ECL.
Nuclear protein extraction and EMSA
Nuclear protein was extracted from SGC-7901 cells,
treated in the presence or absence of VES or PDTC for 24 h, using
the nuclear and cytoplasmic protein extraction kits. Concentrations
were detected by the BCA assay kit according to the manufacturer’s
instructions. The NF-κBp65 combined assay was performed with a
biotin-labeled oligonucleotide (5′-AGT TGA GGG GAC TTT CCC AGG
C-3′). The complex was separated on 6.5% acrylamide gel in 0.5X TBE
buffer at 4°C. Blots were transferred onto N+ nylon membranes
according to the manufacturer’s instructions.
Evaluation of mRNA transcription by
RT-PCR
Total RNA was isolated from 106 SGC-7901
cells following treatment according to the manufacturer’s protocol.
First-strand cDNA was synthesized using the GeneAmp RNA PCR kit.
PCR analyses were performed in a final volume of 20 μl of buffer
containing 1 μl of retro-transcription product, deoxyribonucleotide
triphosphates (150 μM each), MgCl2 (2 mM), 1 unit of Taq
polymerase, and each primer at 1 μM. Following inactivation at 95°C
for 1 min, PCR amplification was performed under the following
reaction conditions: 94°C for 1 min, 50°C (c-IAP-2), 58°C (c-IAP-1,
NAIP, XIAP, and GAPDH), 62°C (survivin) for 1 min, 72°C for 1 min,
and a final extension at 72°C for 5 min. We used 15 cycles of
amplification for GAPDH and 30 cycles for the other mRNAs. All PCR
products (10 μl) were analyzed by electrophoresis on 2% (w/v)
agarose gel, photographed, and quantified by densitometric
scanning. The sequence of primers used for RT-PCR is shown in
Table I. The GAPDH gene was used as
a loading control.
 | Table ISequence of primers used in the
present study. |
Table I
Sequence of primers used in the
present study.
| Primers | Sequence |
|---|
| c-IAP-1 | S:
5′-GAAGACATCTCTTCATCGAGG-3′
A: 5′-CCACAGGTGTATTCATCATGAC-3′ |
| c-IAP-2 | S:
5′-TCCTAGCTGCAGATTCGTTC-3′
A: 5′-GGTAACTGGCTTGAACTTGAC-3′ |
| XIAP | S:
5′-GCACGAGCAGGGTTTCTTTATACTGGTG-3′
A: 5′-CTTCTTCACAATACATGGCAGGGTTCCTC-3′ |
| NIAP | S:
5′-CTGGGCCTAGATGCAGTTCAG-3′
A: 5′-ACGGCTCATAAGTCACAAAAGTC-3′ |
| Survivin | S:
5′-TGCCTGGCAGCCCTTTCTCA-3′
A: 5′-TGGCACGGCGCACTTTCTTC-3′ |
| GAPDH | S:
5′-CATCTTCCAGGAGCGAGAT-3′
A: 5′-GCTTCACCACCTTCTTG-3′ |
Statistical analysis
SAS statistical software was used for data analysis.
The significance between the control and treated groups was
assessed by Student’s t-test, and P<0.01 was considered to
indicate a statistically significant difference in the
experiments.
Results
Inhibition of growth
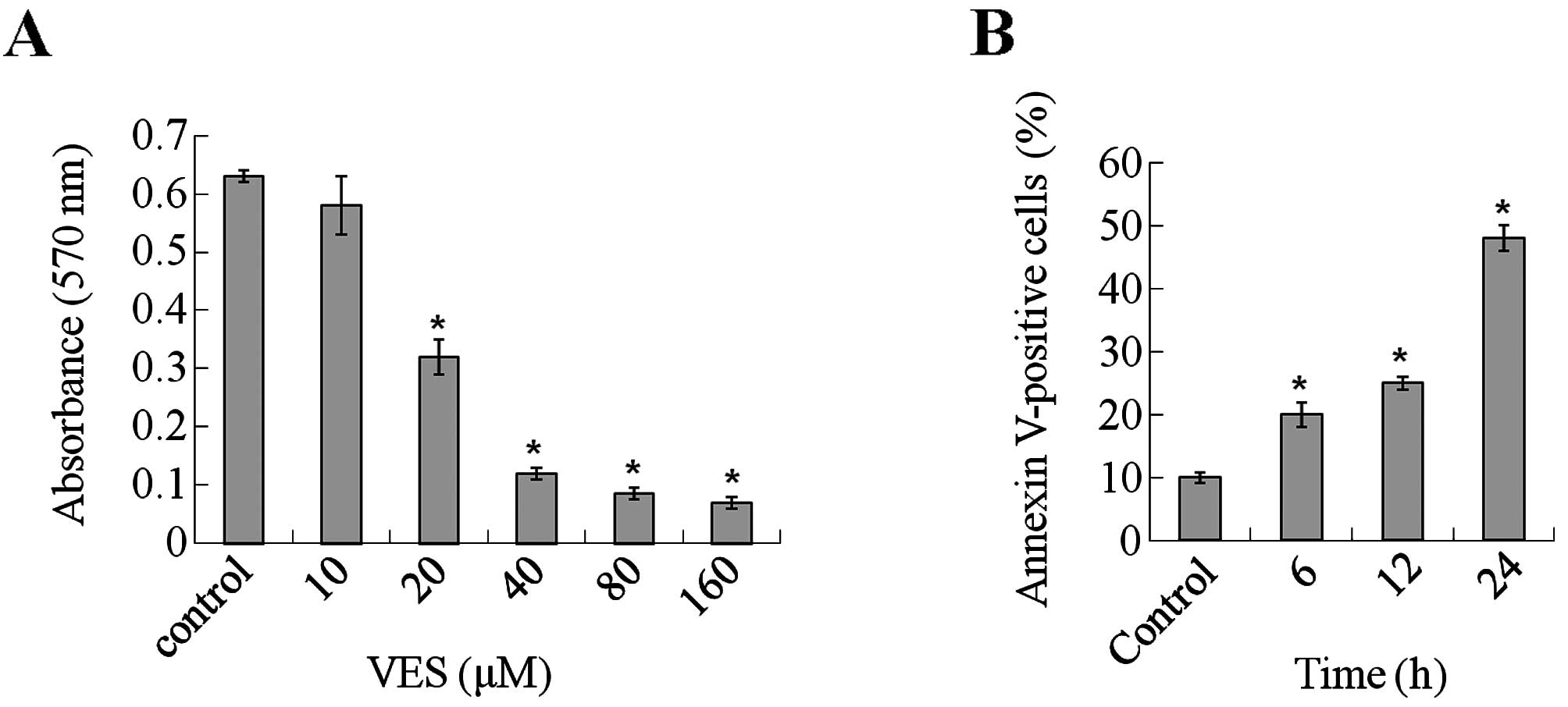
The antitumor effects of VES on human gastric cancer
SGC-7901 cells were evaluated by MTT assay. Various concentrations
of VES (0–160 μM) were used to investigate cell viability after
treatment. As depicted in Fig. 1A,
a significant decrease in cell viability was observed after 24 h of
treatment with 20 to 160 μM VES. Furthermore, when the
concentration of VES was > 40 μM, the cells broke into pieces
quickly after addition of the agent to the culture medium. Hence,
in the present study, moderate concentrations 40 μM were chosen to
study the antitumor effects of VES in SGC-7901 cells.
Induction of apoptosis
For apoptosis assessment, the exposure of
phosphatidylserine on the cell surface was examined by Annexin V
staining. As shown in Fig. 1B, flow
cytometric analysis revealed that the percentage of Annexin
V-positive cells significantly increased following treatment with
VES (40 μM) for 12 h. Following longer treatment, apoptosis became
much more apparent. After VES treatment for 48 h, Annexin
V-positive cells accounted for nearly 50% of all cells counted.
Thus, VES leads to apoptotic cell death in human gastric carcinoma
cells.
Nuclear translocation and DNA-binding
activity of NF-κBp65
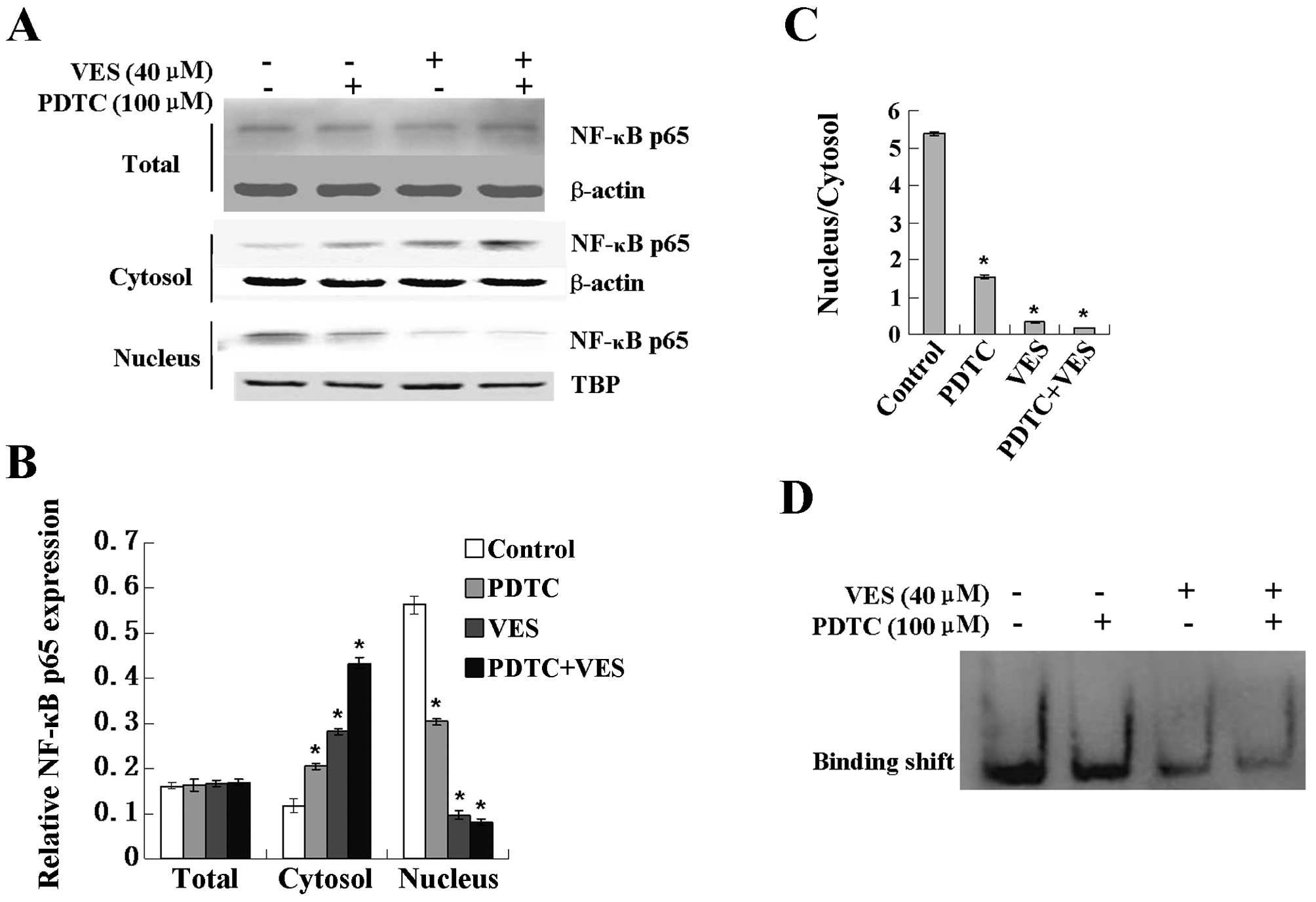
To determine whether the activation of NF-κB was
involved in the apoptosis induced by VES in SGC-7091 cells, PDTC, a
specific inhibitor of the activation of NF-κB, was used in the
following experiments. Western blotting results showed that PDTC
treatment did not influence the expression of the total NF-κBp65
protein level (Fig. 2B). Notably,
there was a distinct increase in cytosolic NF-κBp65 compared with
that in the control group (Fig.
2B). However, there was an obvious decrease in the nuclear
NF-κBp65 protein level. Furthermore, as shown in Fig. 2C, the decrease ratio of the nuclear
and cytosolic NF-κBp65 protein level clearly indicates the
possibility that VES induced the nuclear transformation of NF-κBp65
and PDTC enhanced this transformation.
To examine our hypothesis, NF-κB activity was
measured by EMSA. The DNA binding of NF-κB in SGC-7901 cells
presented inhibited a decreasing trend (Fig. 2D). As shown in Fig. 2D, PDTC treatment significantly
enhanced the decrease of NF-κB-DNA binding activity induced by VES
in human gastric SGC-7901.
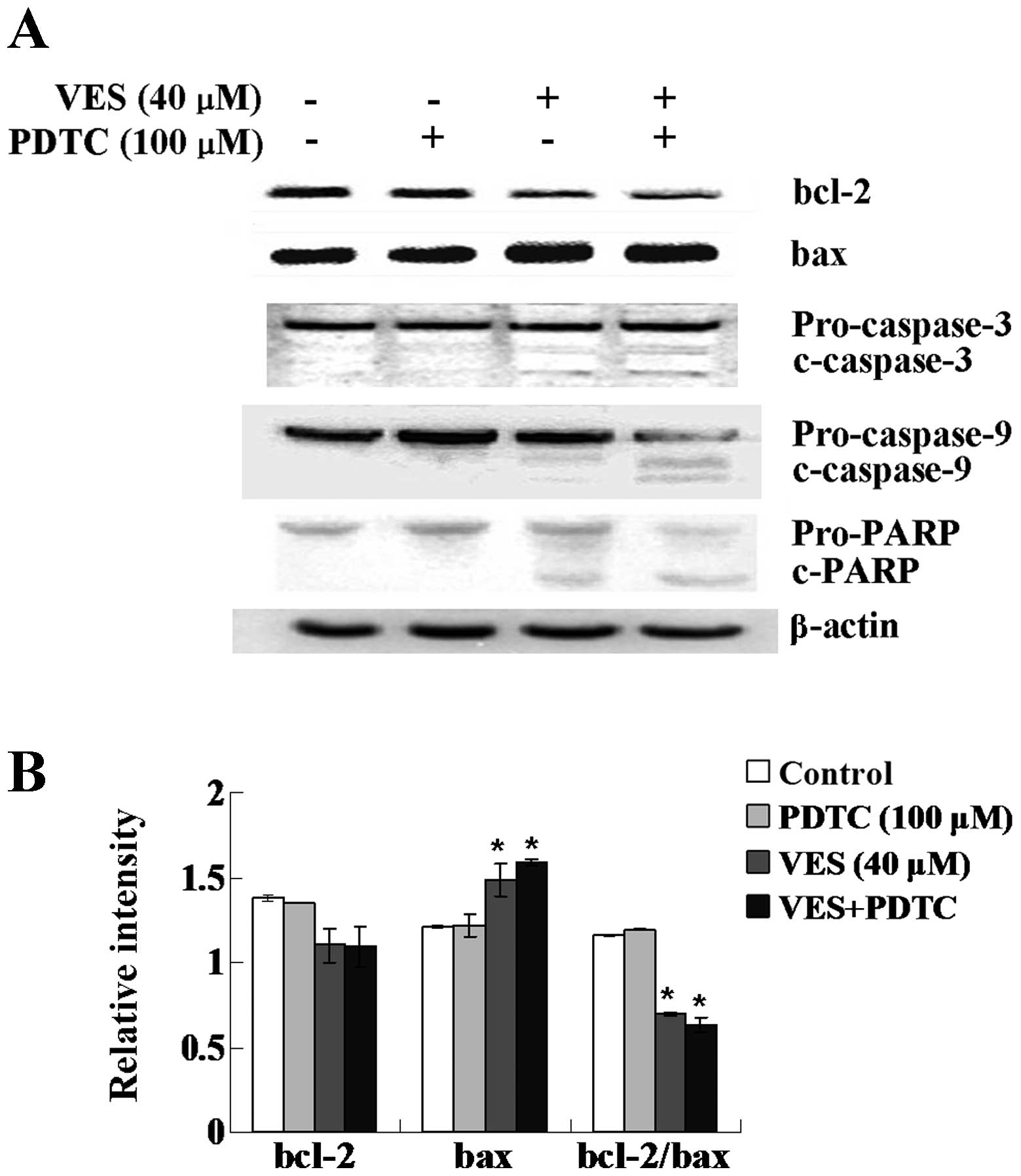
Expression of Bcl-2 family proteins
The protein expression of Bcl-2 family members Bcl-2
and Bax were investigated. VES treatment decreased the protein
expression of Bcl-2 and increased the protein expression of Bax.
The decreased ratio of Bcl-2 and Bax is shown as in Fig. 3B. The trend became more apparent
with PDTC treatment added.
Cleavage of caspase-9, caspase-3 and
PARP
Cleavage of caspase-9, caspase-3 and subsequent
proteolytic cleavage of PARP were assessed by western blot
analysis. As illustrated in Fig.
3A, VES treatment led to cleavage of caspase-9, caspase-3 and
PARP, all of which indicate induction of apoptosis. Notably,
pretreatment with PDTC reinforced the VES-induced cleavage.
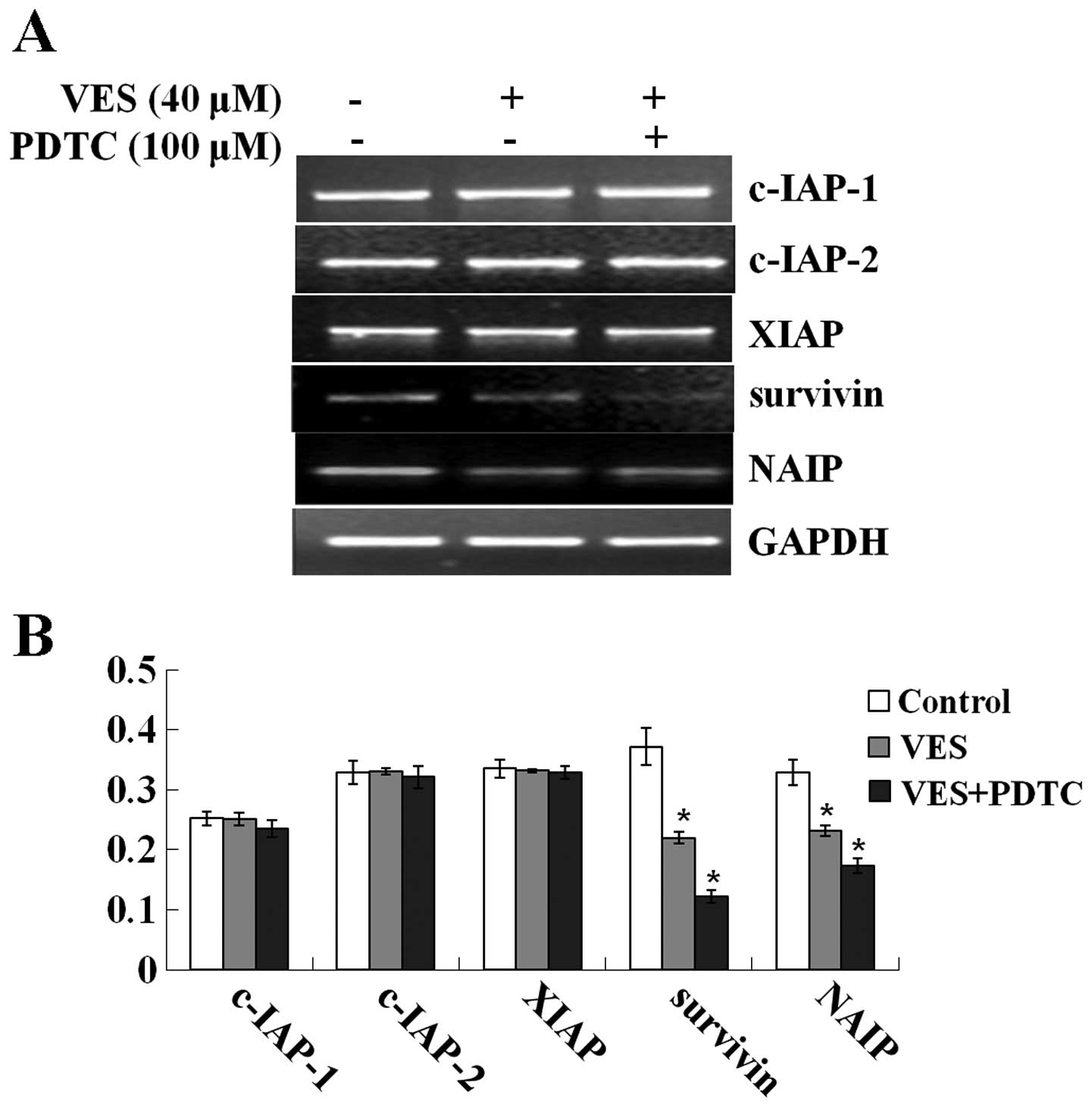
Transcription of IAP genes
VES or PDTC treatment had no effect on the gene
transcription of c-IAP-1, c-IAP-2, and XIAP compared with the
control group, whereas gene mRNA levels of survivin and NAIP
markedly decreased after treatment with VES (Fig. 4). Furthermore, PDTC intensified the
decrease of gene mRNA levels.
Discussion
The potential anticancer effect of VES in human
gastric carcinoma cells has been indicated by our previous in
vivo experiment. We found that VES induced apoptosis in
SGC-7901 human gastric cancer cells via multiple signaling
pathways, including extrinsic Fas, MAP kinase, endoplasmic
reticulum (ER) stress, the couple of ER stress and unfolded protein
response (UPR) (8–10, 20–22).
In addition, VES enhanced DOXO anticancer efficiency via promotion
of DOXO influx and suppression of MDR-1 mediated DOXO efflux
(21). Recent studies showed that
NF-κB was not only related to immune response and inflammatory
reaction, but was also involved in regulating cell proliferation,
apoptosis and migration (23–25).
Involvement of the NF-κB signaling pathway in the genesis and
progression of malignant gastric cancer led us to undertake this
study to explore the mechanism of induction of apoptosis of VES as
well as the relationship between this mechanism and the NF-κB
signaling pathway in human gastric carcinoma cells.
In this study, moderate concentrations of VES (10–40
μM) exhibited apparent growth inhibition effects and induced
apoptosis in human gastric carcinoma SGC-7901 cells. The
concentration that significantly decreased SGC-7901 cell viability
and induced apoptosis was similar to that in other cancer cell
cultures, such as leukemia and breast (26–28).
VES treatment caused reductions in nuclear and an
increase in cytosolic NF-κBp65 protein levels accompanied by
fixedness in total NF-κBp65 protein levels. These results are
consistent with those of previous reports (29,30).
Several mechanisms for NF-κB inhibition are available, such as
inhibition of the inhibitor of NF-κB (IκB) degradation,
phosphorylation of the antagonist of IκB by inhibition of IκB
kinase α, and NF-κB nuclear translocation (31–34).
According to our results, VES may regulate the NF-κB pathway by
inhibiting the nuclear translocation of NF-κBp65 in SGC-7901 cells.
With the involvement of PDTC, this hypothesis is confirmed.
Overexpression of NF-κB-responsive proteins, such as
Bcl-2 family members and IAPs, resulting from the constitutive
activation of NF-κB may critically contribute to the genesis and
progression of cancer and represent an important cause of tumor
drug resistance (35,36). As previously reported, high NF-κB
expression is associated with high Bcl-2 expression in breast
cancer and leukemic cells (37).
Several Bcl-2 family members are known to be involved in the
apoptosis-inducing effect of VES in tumor cells in vitro
(38,39). We further verified that the
intrinsic pathway of cell death is mediated by Bcl-2 family
proteins and the balance between pro- and anti-apoptotic Bcl-2
family proteins. This balance may terminally determine
mitochondrial disruption, cytochrome c release, and caspase
activation until apoptosis. Only this ratio dictates whether or not
a cell responds to a proximal apoptotic stimulus (40). Caspases are known to be important
mediators of apoptosis and contribute to the overall apoptotic
morphology by cleaving various cellular substrates (41,42).
Activation of caspases is the terminal phase of programmed cell
death, and this step is contracted by anti-apoptotic molecules of
the Bcl-2 family (43). Our results
confirm that VES-mediated changes in Bcl-2 family members trigger
the release of mitochondrial cytochrome c, which, in turn,
accounts for the cleavage of caspase-9, caspase-3 and PARP and
ultimately contributes to cell death. Furthermore, this effect is
enhanced by PDTC, an inhibitor of NF-κB.
IAP family proteins play a role in oncogenesis via
their effective suppression of apoptosis (44). NF-κB is known to regulate the
expression of anti-apoptotic genes such as IAP-1, IAP-2, XIAP, NAIP
and survivin (45). Thus, in this
study, whether or not VES treatment regulates the gene
transcription of IAP family proteins was determined. Inhibition of
constitutive NF-κBp65 activation could trigger decreases in
survivin and NAIP mRNA levels without changing the levels of other
inhibitors. Hence, only survivin and NAIP are involved in the
apoptosis-inducing effects of VES, which are caused by the
inhibition of constitutive NF-κBp65 activation in VES-treated human
gastric carcinoma cells. In contrast to our previous studies
carried out using other types of human cancer cells (46). This result indicated that survivin
and NAIP may be involved in the VES-induced apoptosis in SGC-7901.
However, the inhibition of nuclear translocation of NF-κBp65 did
not downregulate the expression of all anti-apoptotic genes. There
may be other nuclear translocation factors which can regulate the
translocation of IAP family genes.
In conclusion, our results demonstrate that
VES-induced apoptosis in human gastric carcinoma SGC-7901 cells is
accompanied by changes in Bcl-2 family members, cleavage of
caspases, and inhibition of the NF-κB signaling pathway. Nuclear
translocation of NF-κBp65 is markedly inhibited after treatment of
SGC-7901 cells with VES. Additional studies are required to
understand the biochemical mechanisms involved in the induction of
apoptosis caused by treatment with VES in human gastric cancer
cells. We believe that NF-κB may be a favorable candidate protein
for developing new molecular-targeted therapies against human
gastric cancer.
Acknowledgements
This study was supported in part by grants from the
National Natural Science Foundation of China (No. 81172651) to
K.W.
References
|
1
|
Foukakis T, Lundell L, Gubanski M and Lind
PA: Advances in the treatment of patients with gastric
adenocarcinoma. Acta Oncol. 46:277–285. 2007. View Article : Google Scholar
|
|
2
|
Tomasetti M, Gellert N, Procopio A and
Neuzil J: A vitamin E analogue suppresses malignant mesothelioma in
a preclinical model: a future drug against a fatal neoplastic
disease? Int J Cancer. 109:641–642. 2004. View Article : Google Scholar
|
|
3
|
Neuzil J, Weber T, Gellert N and Weber C:
Selective cancer cell killing by alpha-tocopheryl succinate. Br J
Cancer. 84:87–89. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Malafa MP, Fokum FD, Mowlavi A, Abusief M
and King M: Vitamin E inhibits melanoma growth in mice. Surgery.
131:85–91. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kline K, Yu W and Sanders BG: Vitamin E
and breast cancer. J Nutr. 134:S3458–S3462. 2004.
|
|
6
|
Wu K, Zhao Y, Liu BH, et al:
RRR-alpha-tocopheryl succinate inhibits human gastric cancer
SGC-7901 cell growth by inducing apoptosis and DNA synthesis
arrest. World J Gastroenterol. 8:26–30. 2002.PubMed/NCBI
|
|
7
|
Dong LF, Jameson VJ, Tilly D, et al:
Mitochondrial targeting of vitamin E succinate enhances its
pro-apoptotic and anti-cancer activity via mitochondrial complex
II. J Biol Chem. 286:3717–3728. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wu K, Li Y, Zhao Y, et al: Roles of Fas
signaling pathway in vitamin E succinate-induced apoptosis in human
gastric cancer SGC-7901 cells. World J Gastroenterol. 8:982–986.
2002.PubMed/NCBI
|
|
9
|
Wu K, Zhao Y, Li GC and Yu WP: c-Jun
N-terminal kinase is required for vitamin E succinate-induced
apoptosis in human gastric cancer cells. World J Gastroenterol.
10:1110–1114. 2004.PubMed/NCBI
|
|
10
|
Huang X, Zhang Z, Jia L, Zhao Y, Zhang X
and Wu K: Endoplasmic reticulum stress contributes to vitamin E
succinate-induced apoptosis in human gastric cancer SGC-7901 cells.
Cancer Lett. 296:123–131. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Guo JL, Zheng SJ, Li YN, et al:
Toxicarioside A inhibits SGC-7901 proliferation, migration and
invasion via NF-κB/bFGF signaling. World J Gastroenterol.
18:1602–1609. 2012.PubMed/NCBI
|
|
12
|
Long YM, Ye S, Rong J and Xie WR: Nuclear
factor kappa B: a marker of chemotherapy for human stage IV gastric
carcinoma. World J Gastroenterol. 14:4739–4744. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhu BS, Xing CG, Lin F, Fan XQ, Zhao K and
Qin ZH: Blocking NF-kappaB nuclear translocation leads to
p53-related autophagy activation and cell apoptosis. World J
Gastroenterol. 17:478–487. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Li X and Stark GR: NFkappaB-dependent
signaling pathways. Exp Hematol. 30:285–296. 2002. View Article : Google Scholar
|
|
15
|
Karin M and Lin A: NF-kappaB at the
crossroads of life and death. Nat Immunol. 3:221–227. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Gilmore TD: The Re1/NF-kappa B/I kappa B
signal transduction pathway and cancer. Cancer Treat Res.
115:241–265. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bharti AC and Aggarwal BB: Nuclear
factor-kappa B and cancer: its role in prevention and therapy.
Biochem Pharmacol. 64:883–888. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chen W, Li Z, Bai L and Lin Y: NF-kappaB
in lung cancer, a carcinogenesis mediator and a prevention and
therapy target. Front Biosci. 16:1172–1185. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sebens S, Arlt A and Schafer H: NF-kappaB
as a molecular target in the therapy of pancreatic carcinoma.
Recent Results Cancer Res. 177:151–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wu K, Shan YJ, Zhao Y, Yu JW and Liu BH:
Inhibitory effects of RRR-alpha-tocopheryl succinate on
benzo(a)pyrene (B(a)P)-induced forestomach carcinogenesis in female
mice. World J Gastroenterol. 7:60–65. 2001.PubMed/NCBI
|
|
21
|
Zhang X, Peng X, Yu W, et al:
Alpha-tocopheryl succinate enhances doxorubicin-induced apoptosis
in human gastric cancer cells via promotion of doxorubicin influx
and suppression of doxorubicin efflux. Cancer Lett. 307:174–181.
2011. View Article : Google Scholar
|
|
22
|
Huang X, Li L, Zhang L, et al: Crosstalk
between endoplasmic reticulum stress and oxidative stress in
apoptosis induced by alpha-tocopheryl succinate in human gastric
carcinoma cells. Br J Nutr. 109:1–9. 2013. View Article : Google Scholar
|
|
23
|
Chen X, Kandasamy K and Srivastava RK:
Differential roles of RelA (p65) and c-Rel subunits of nuclear
factor kappa B in tumor necrosis factor-related apoptosis-inducing
ligand signaling. Cancer Res. 63:1059–1066. 2003.PubMed/NCBI
|
|
24
|
Sun SC and Xiao G: Deregulation of
NF-kappaB and its upstream kinases in cancer. Cancer Metastasis
Rev. 22:405–422. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lin A and Karin M: NF-kappaB in cancer: a
marked target. Semin Cancer Biol. 13:107–114. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Neuzil J, Svensson I, Weber T, Weber C and
Brunk UT: alpha-tocopheryl succinate-induced apoptosis in Jurkat T
cells involves caspase-3 activation, and both lysosomal and
mitochondrial destabilisation. FEBS Lett. 445:295–300. 1999.
View Article : Google Scholar
|
|
27
|
Wang XF, Witting PK, Salvatore BA and
Neuzil J: Vitamin E analogs trigger apoptosis in
HER2/erbB2-overexpressing breast cancer cells by signaling via the
mitochondrial pathway. Biochem Biophys Res Commun. 326:282–289.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yu W, Sanders BG and Kline K:
RRR-alpha-tocopheryl succinate-induced apoptosis of human breast
cancer cells involves Bax translocation to mitochondria. Cancer
Res. 63:2483–2491. 2003.PubMed/NCBI
|
|
29
|
Crispen PL, Uzzo RG, Golovine K, et al:
Vitamin E succinate inhibits NF-kappaB and prevents the development
of a metastatic phenotype in prostate cancer cells: implications
for chemoprevention. Prostate. 67:582–590. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Dalen H and Neuzil J: Alpha-tocopheryl
succinate sensitises a T Lymphoma cell line to TRAIL-induced
apoptosis by suppressing NF-kappaB activation. Br J Cancer.
88:153–158. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mitsiades N, Mitsiades CS, Richardson PG,
et al: The proteasome inhibitor PS-341 potentiates sensitivity of
multiple myeloma cells to conventional chemotherapeutic agents:
therapeutic applications. Blood. 101:2377–2380. 2003. View Article : Google Scholar
|
|
32
|
Zheng B, Georgakis GV, Li Y, et al:
Induction of cell cycle arrest and apoptosis by the proteasome
inhibitor PS-341 in Hodgkin disease cell lines is independent of
inhibitor of nuclear factor-kappaB mutations or activation of the
CD30, CD40, and RANK receptors. Clin Cancer Res. 10:3207–3215.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cilloni D, Messa F, Arruga F, et al: The
NF-kappaB pathway blockade by the IKK inhibitor PS1145 can overcome
imatinib resistance. Leukemia. 20:61–67. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dai Y, Pei XY, Rahmani M, Conrad DH, Dent
P and Grant S: Interruption of the NF-kappaB pathway by Bay 11-7082
promotes UCN-01-mediated mitochondrial dysfunction and apoptosis in
human multiple myeloma cells. Blood. 103:2761–2770. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Li H and Lin X: Positive and negative
signaling components involved in TNFalpha-induced NF-kappaB
activation. Cytokine. 41:1–8. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Müerköster S, Arlt A, Sipos B, et al:
Increased expression of the E3-ubiquitin ligase receptor subunit
betaTRCP1 relates to constitutive nuclear factor-kappaB activation
and chemoresistance in pancreatic carcinoma cells. Cancer Res.
65:1316–1324. 2005.
|
|
37
|
Viatour P, Bentires-Alj M, Chariot A, et
al: NF-kappa B2/p100 induces Bcl-2 expression. Leukemia.
17:1349–1356. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Al-Harbi S, Hill BT, Mazumder S, et al: An
antiapoptotic BCL-2 family expression index predicts the response
of chronic lymphocytic leukemia to ABT-737. Blood. 118:3579–3590.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Kelly PN and Strasser A: The role of Bcl-2
and its pro-survival relatives in tumourigenesis and cancer
therapy. Cell Death Differ. 18:1414–1424. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Oltvai ZN, Milliman CL and Korsmeyer SJ:
Bcl-2 heterodimerizes in vivo with a conserved homolog, Bax, that
accelerates programmed cell death. Cell. 74:609–619. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Pop C and Salvesen GS: Human caspases:
activation, specificity, and regulation. J Biol Chem.
284:21777–21781. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Olsson M and Zhivotovsky B: Caspases and
cancer. Cell Death Differ. 18:1441–1449. 2011. View Article : Google Scholar
|
|
43
|
Gross A, McDonnell JM and Korsmeyer SJ:
BCL-2 family members and the mitochondria in apoptosis. Genes Dev.
13:1899–1911. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Gyrd-Hansen M, Darding M, Miasari M, et
al: IAPs contain an evolutionarily conserved ubiquitin-binding
domain that regulates NF-kappaB as well as cell survival and
oncogenesis. Nat Cell Biol. 10:1309–1317. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Trocoli A and Djavaheri-Mergny M: The
complex interplay between autophagy and NF-kappaB signaling
pathways in cancer cells. Am J Cancer Res. 1:629–649.
2011.PubMed/NCBI
|
|
46
|
Neuzil J, Tomasetti M, Zhao Y, et al:
Vitamin E analogs, a novel group of “mitocans,” as anticancer
agents: the importance of being redox-silent. Mol Pharmacol.
71:1185–1199. 2007.
|