Introduction
Cervical cancer is the second most common cancer
among women worldwide and the fourth leading cause of
cancer-associated mortality in women, accounting for 9% of total
new cases of cancer and 8% of all cancer deaths. Over 85% of these
mortalities occur in developing countries with approximately
250,000 women succumbing to the disease annually (1,2).
Although the mortality rate has declined due to improvement in
screening, diagnostic, prognostic and treatment modalities in
recent years, treatments currently available including surgery,
radiotherapy, and chemotherapy are often unsatisfactory,
particularly for patients with advanced stage of cervical cancer
(3,4). Chemotherapy drugs have been widely
used for patients with advanced cervical cancer. However, many
patients acquire resistance to chemotherapeutic agents, leading to
treatment failures (4,5). Additionally, chemotherapy drugs
exhibit high toxicity in normal tissues (4–6).
Therefore, the identification of new drugs with few side-effects
are required to improve the survival and quality of life of
cervical cancer patients.
Natural products are suitable alternatives that can
be used instead of platinum-based drugs since they are much more
effective and have minimal side-effect consequences compared to
synthetic drugs (7,8). Numerous natural products have been
identified as promising sources of drugs for cancer prevention and
treatment, based on their ability to attack multiple molecular
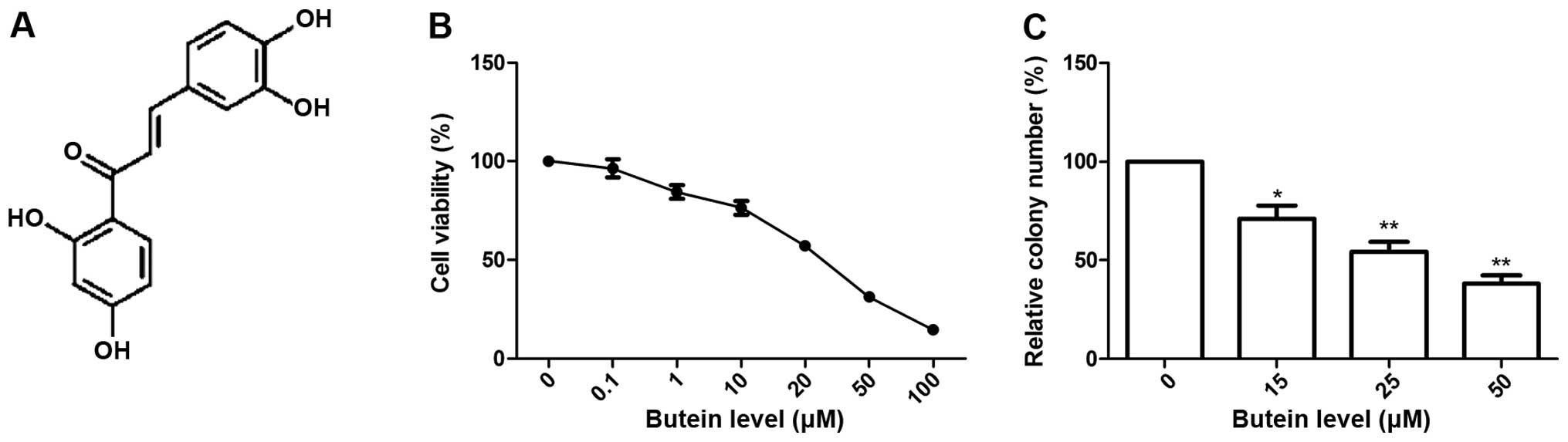
targets (9–13). Butein (2′,3,4,4′-2′,4′,3,4- or
3,4,2′,4′-tetrahy-droxychalcone, chemical structure shown in
Fig. 1A), an important natural
products, can be isolated in various plants including
Toxicodendron vernicifluum (Rhus verniciflua),
Semecarpus anacardium and Dalbergia odorifera, and
has exhibited different pharmacological effects, including
antioxidant, anti-inflammatory, anti-allergic and anti-angiogenic
activities (14–18). Accumulating evidence has
demonstrated that butein suppressed tumor growth by inhibiting cell
proliferation and inducing cell apoptosis in various human cancer
cells, such as colorectal carcinoma (19), lung cancer (20), melanoma (21), prostate cancer (22), leukemia (23), breast carcinoma (24), osteosarcoma cells (25), hepatocellular carcinoma (26) and neuroblastoma cells (27). Butein has recently received wide
attention as a useful chemopreventive and chemotherapeutic agent
due to exhibiting only minimal toxicity in normal cells (28,29). A
preliminary clinical trial on the effects of flavonoids containing
butein in gastric cancer patients showed that this drug was safe
with good tolerability, exhibiting a marked decrease in tumor size
(30). Nonetheless, to the best of
our knowledge, its potential role in apoptosis and tumor
inhibition, as well as the underlying anticancer mechanism in
cervical cancer remains to be elucidated.
The aim of the present study was to evaluate the
potency of butein in inhibiting cervical cancer cell proliferation,
colony formation, migration and invasion, inducing cell apoptosis
and cell-cycle arrest in vitro, as well as to identify the
underlying molecular mechanisms associated with anticancer
activity. In addition, tumor growth ability in nude mice was
detected to define the butein treatment effect in the tumorigenesis
of cervical cancer in vivo.
Materials and methods
Reagents and antibodies
Butein was purchased from Santa Cruz Biotechnology,
Inc. (Santa Cruz, CA, USA). Butein was dissolved in
dimethylsulfoxide (DMSO) and stored at −20°C until use. The
concentration of DMSO used was <1% in the control and the
butein-containing medium.
Dulbecco’s modified Eagle’s medium (DMEM) and fetal
bovine serum (FBS) were purchased from Gibco (Grand Island, NY,
USA). Propidium iodide (PI) and
3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT)
were obtained from Sigma (St. Louis, MO, USA). Stock solutions of
PI and MTT were prepared by dissolving 1 mg of each compound in 1
ml of phosphate-buffered saline (PBS). The solution was stored at
4°C in the dark and used within 1 month. Caspase-3, -8 and -9
Colorimetric Activity Assay kits were purchased from R&D
Systems (Minneapolis, MN, USA). Apoptotic Cells Detection kit
(Annexin V-FITC/ethidium homodimer III staining) was purchased from
PromoKine Systems (Heidelberg, Germany).
For western blot analysis, the antibodies used were:
a mouse monoclonal anti-human Bcl2, mouse monoclonal anti-human
Bax, mouse monoclonal anti-human MMP-2, and mouse monoclonal
anti-human MMP-9, all purchased from Cell Signaling Technology
(Beverly, MA, USA). Mouse monoclonal anti-human AKT, mouse
monoclonal anti-human phosphorylated (p)-AKT, mouse monoclonal
anti-human PI3K, mouse monoclonal anti-human (p)-PI3K, mouse
monoclonal anti-human mTOR, mouse monoclonal anti-human (p)-mTOR,
were obtained from Santa Cruz Biotechnology, Inc. Mouse monoclonal
anti-human β-actin and anti-mouse immuno globulin G-horseradish
peroxidase-conjugated secondary antibodies were purchased from
Sigma-Aldrich (St. Louis, MO, USA).
Cell lines and culture
The HeLa human cervical cancer cell line was
purchased from the Cell Bank of Type Culture Collection of Chinese
Academy of Sciences, Shanghai Institute of Cell Biology (Shanghai,
China). Cultured cells were grown in DMEM supplemented with 10%
fetal calf serum and antibiotics (100 U/ml penicillin and 100 mg/ml
streptomycin) at 37°C in a humidified atmosphere containing 5%
CO2.
Cell viability
Cell viability was determined by MTT assay as
previously described (31).
Briefly, HeLa cells were cultured in 96-well plates and treated
with different concentrations of butein (0–100 μM) for 72 h.
Cell viability was determined via an MTT assay. Absorbance at 570
nm was measured in an ELISA plate reader (Molecular Devices Corp.,
Sunnyvale, CA, USA).
Colony formation assay
HeLa cells were seeded in 6-well plates at a density
of 1×103 cells/well. After being cultured for 24 h, the
cells were treated with the indicated concentrations of butein (0,
15, 25 and 50 μM). The cells were then washed with
drugs-free medium and allowed to grow for 14 days in drugs-free
conditions. After 14 days, the cells were fixed in paraformaldehyde
and stained with Giemsa for 15 min, and images of the colonies were
captured by a digital camera (Olympus, Tokyo, Japan). The
percentage of colony formation was calculated by adjusting control
(untreated cells) to 100%.
Cell cycle analysis
The cells were seeded at a density of
1×106 cells/well in 6-well plates in complete medium for
16 h. At the end of treatment with the indicated concentration of
butein, the cells were collected and fixed with ice-cold 70%
ethanol overnight at −20°C. After centrifugation at 3,000 × g for 5
min, the cell pellets were added into 4 μg/ml PI solution
containing 1% Triton X-100 and 100 μg/ml RNase at 37°C for
30 min. The samples were analyzed by a flow cytometer
(FACSCalibur), and the percentage of cell-cycle phases was
quantified using ModFit LT software 2.0 (BD Biosciences, San Jose,
CA, USA).
Cell apoptosis assays
The cells were plated at a density of
5×105 cells/60-mm Petri dish in complete medium for 16
h, and then treated with the indicated concentrations of butein for
24 h. Apoptotic cells were assessed using an Apoptotic Cells
Detection kit according to the manufacturer’s instructions. The
cells were collected and resuspended in 500 μl of binding
buffer, and 5 μl of Annexin V-FITC and 5 μl of PI
were added. The cells were cultured for 10 min at room temperature.
The samples were analyzed by flow cytometer using CellQuest
software (BD Biosciences).
In addition, the expression of Bax and Bcl-2 were
determined by western blotting 24 h after treatment with the
different concentrations of butein.
Caspase activity assay
Activities of caspase-3, -8 and -9 were detected as
previously described (21).
Briefly, HeLa cells were seeded in 6-well plates at a density of
4×105 cells/well and cultured for 24 h, then treated
with indicated concentration of butein for 2 h. The cells were
washed with cold PBS and collected by centrifugation at 1,500 rpm
for 5 min at 4°C. The cells were lysed using a cell extraction
buffer (BioSource International, Camarillo, CA, USA) with protease
inhibitor cocktail (Sigma) and PMSF (BioSource), incubated on ice
for 30 min and vortexed for 30 sec. The samples were centrifuged at
10,000 rpm for 10 min at 4°C to remove the supernatant. The
caspase-3, -8 and -9 levels in cell lysates were measured using the
caspase-3, -8 and -9 colorimetric kit according to the
manufacturer’s instructions. The optical density was read using a
microplate reader (Thermo Fisher Scientific, Inc., Waltham, MA,
USA) at 405 nm. The relative caspase activity of the untreated
group (0 μM butein treatment) was taken as 100.
Cell invasion and migration assays
The cell invasion and migration assays were carried
out using Transwell inserts (Corning, Corning, NY, USA) according
to the manufacturer’s instructions. For the invasion assay, filters
were precoated with Matrigel (BD Biosciences, Bedford, MA, USA) for
30 min. Approximately 5×104 cells were placed in the
upper chamber in serum-free DMEM, and 500 μl of DMEM
containing 20% FBS and the indicated concentrations of butein (0,
15, 25 and 50 μM) were placed in the lower chamber in the
invasion and migration assays. After 24 h of treatment, the cells
on the upper side of the filters were mechanically removed by
wiping with a cotton swab. Cells migrating to the lower surface of
filter were fixed in 70% ethanol for 15 min and stained with 0.2%
crystal violet for 15 min. The invaded or migrated cells were
photographed and quantified by counting the number of stained cells
in five randomly selected fields under a light microscope
(Olympus).
Reactive oxygen species (ROS)
measurement
Generation of intracellular ROS was examined by flow
cytometry as previously described (32). Briefly, 5×104 HeLa cells
were seeded in 60-mm dishes overnight, and exposed to different
concentrations of butein (0, 15, 25 and 50 μM) for 1 h. The
cells then were stained with 2 μM hematoxylin and eosin
(H&E) or 5 μM H2DCFDA at 37°C for 30 min. The
cells were collected and fluorescence was analyzed using a flow
cytometer. The cells were also pretreated with 10 mM NAC prior to
butein exposure and analysis of ROS generation.
Western blot assay
At the end of treatment, the cells were lysed in the
ice-cold whole cell extract buffer containing protease inhibitors
(Sigma) for 30 min, and centrifuged at 10,000 rpm for 10 min at
4°C. The protein concentration was measured by BCA protein assay
kit (Pierce, Rockford, IL, USA) according to the manufacturer’s
instructions. Equal amounts of total proteins (20 μg) were
subjected to electrophoresis using 8–12% sodium dodecyl
sulfate-polyacrylamide gels (SDS-PAGE). After electrophoresis, the
proteins were transferred to polyvinylidene difluoride membranes.
The membranes were blocked with 5% dry milk in TBS-T overnight at
4°C and then incubated with specific primary antibody at room
temperature for 2 h. After washing with TBS-T twice, the membranes
were incubated with anti-mouse immunoglobulin G-horseradish
peroxidase-conjugated secondary antibodies, followed by washing
three times with TBS-T. The protein bands were then visualized on
the X-ray film using the enhanced chemiluminescence detection
system (PerkinElmer Life and Analytical Sciences, Boston, MA, USA).
Blots were stripped and reprobed with anti-β-actin for the loading
variations. Quantity One software (Bio-Rad) was used to analyze the
intensity of bands on X-ray film by semi-quantification.
Xenograft mouse model
Twenty female BALB/c nude mice (6 weeks old) were
obtained from the Experiments Animal Center of Changchun Biological
Institute (Changchun, China), and maintained under specific
pathogen-free conditions based on the guidelines established by the
Institutional Animal Care and Use Committee of Jilin University.
HeLa cells (2×106) mixed with 10% Matrigel (R&D
Biosystems) were subcutaneously injected into the right flank of
each mouse. Tumor-bearing mice were randomly divided into two
groups with 10 animals each. Animals received either
intraperitoneal (i.p.) injection of 1% polysorbate resuspended in
deionized water (control group) or butein (5 mg/kg in 100
μl) on alternate days for 4 weeks. The mice were weighed
weekly to determine any toxicity associated with butein treatments,
and the tumors were measured using digital vernier calipers. The
volume was calculated as: (π/6 × length × width × height). There
was no mortality during the treatment regimen. At the end of the
study, the animals were euthanized by CO2 asphyxiation,
the tumor tissues were harvested and weighed. Sections of each
tumor tissue were wax-embedded and H&E stained to study cell
apoptosis in vivo by TUNEL using In Situ Cell Death
Detection kit (Roche Diagnostics, Branchburg, NJ, USA) according to
the manufacturer’s instructions.
Statistical analysis
For the in vitro and in vivo studies,
the statistical significance was evaluated by the two-tailed
Student’s t-test and ANOVA, and data are presented as the mean ± SD
(standard deviation). In vitro assessments were performed in
three independent experiments to confirm the results. Statistical
analyses were performed using the GraphPad Prism version 5.01
(GraphPad Software, San Diego, CA, USA). P<0.05 was considered
to indicate a statistically significant result.
Results
Butein decreases cell viability and
colony formation of HeLa cells
To examine the proliferation effects of butein, HeLa
cells were treated with different dose of butein (0–100 μM)
for 72 h, and the percentage of viable cells was determined.
Results shown in Fig. 1B indicated
that butein decreased cell viability of HeLa cells in a
dose-dependent manner. The IC30 values (the effective
dose that inhibits 30% growth) and IC50 values (the
effective dose that inhibits 50% growth) for treatment of HeLa
cells by butein were 14.89 and 24.76 μM, respectively. Based
on the results we selected the concentrations of 15 μM
(IC30), 25 μM (IC50) and 50 μM
(2×IC50) salidroside for subsequent experiments.
The effects of butein on the cell colony formation
of HeLa cells were also investigated. It was found that butein
significantly inhibited the colony formation of HeLa cells in a
dose-dependent manner (P<0.05, Fig.
1C).
Butein-induced G2/M phase arrest and
apoptosis of HeLa cells
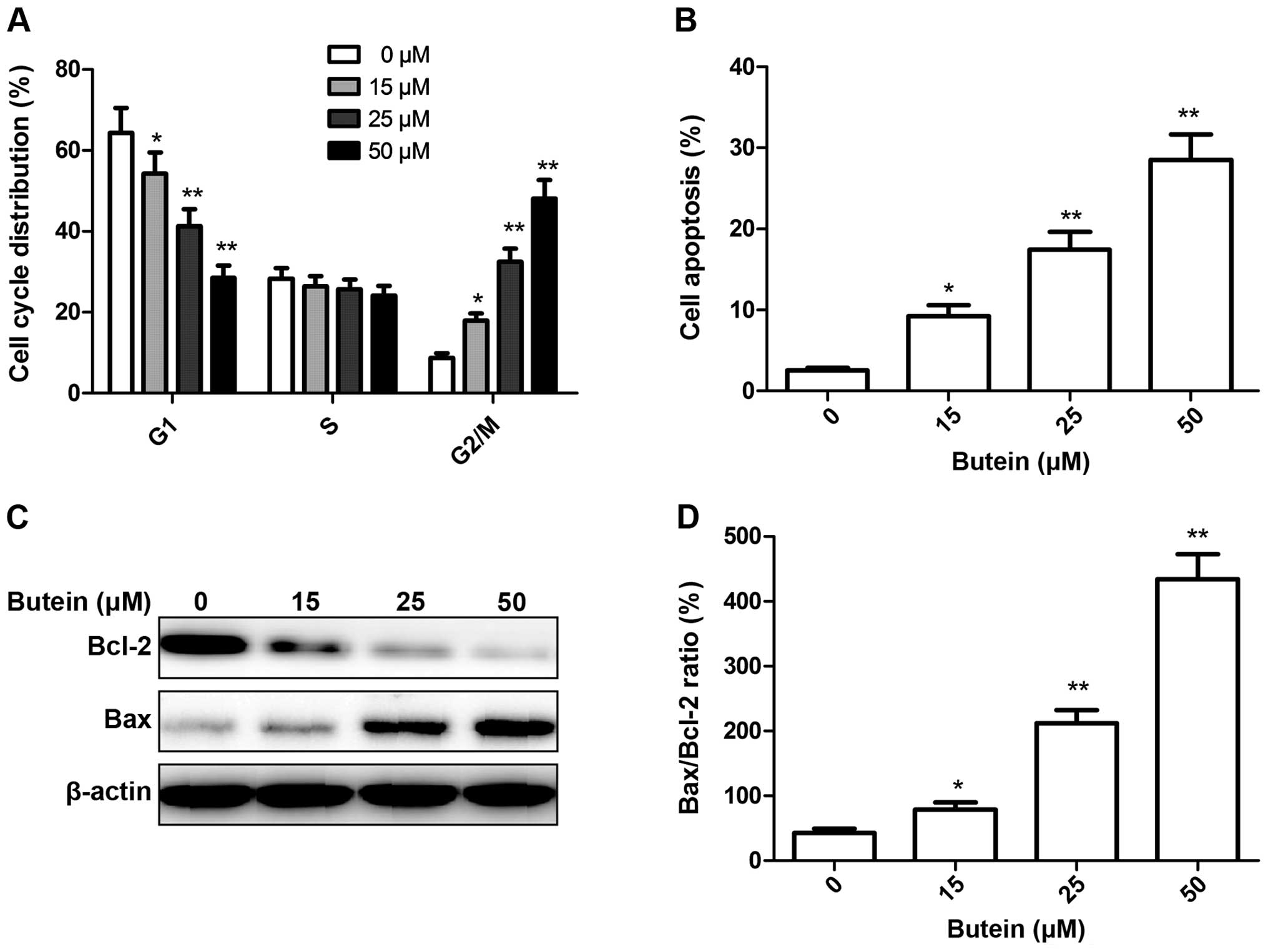
To determine whether the growth inhibitory effect of
butein in HeLa cells was associated with the induction of the cell
cycle, we monitored cell-cycle progression using flow cytometry
following treatment with the indicated concentrations of butein (0,
15, 25 and 50 μM). Exposure to butein resulted in an
increase of G2/M phase cells, accompanied by a decrease in G1 phase
cells (Fig. 2A). The effect
observed at 50 μM butein was the greatest, with ~48% of
cells being in the G2/M phase, compared to ~9% in the control
condition.
The effects of butein on the cell apoptosis of HeLa
cells were also examined. HeLa cells were treated with butein at
different concentrations of 0, 15, 25 and 50 μM for 48 h,
and cell apoptosis was analyzed by flow cytometry (Fig. 2B). It was found that butein
significantly increased total apoptotic rate of HeLa cells in a
dose-dependent manner (P<0.05). Consequently, our results
demonstrated that treatment with butein inhibits HeLa cell growth
by inducing G2/M transition and cell apoptosis.
To determine the potential mechanism of butein, the
effects of cell apoptosis, apoptosis-related protein, Bax and Bcl-2
expression were examined by western blot analysis. The results
revealed a significant upregulation of Bax expression and
downregulation of Bcl-2 expression in HeLa cells treated with
butein in a dose-dependent manner (Fig.
2C). Butein upregu-lated the Bax/Bcl-2 ratio in HeLa cells in a
dose-dependent manner (Fig. 2D),
which triggered the release of cytochrome c from the
mitochondria and stimulated pro-caspase-3, thereby promoting cell
apoptosis.
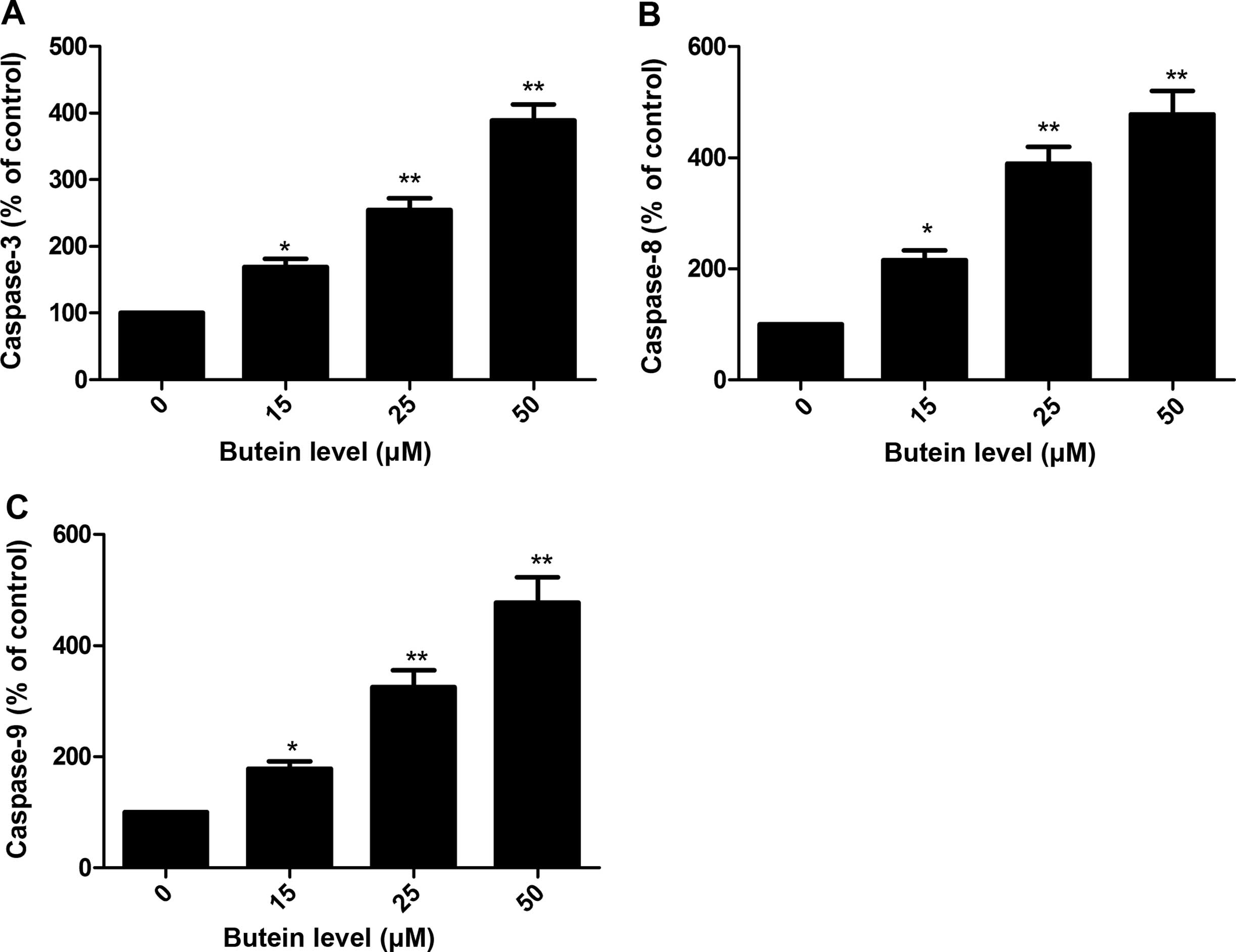
Butein increased caspase-3, -8 and -9
activities of HeLa cells
To examine the contribution of caspases in the
butein-induced apoptosis, the role of caspase-3, -8 and -9 was
investigated after treatment with different concentration butein
(0, 15, 25 and 50 μM). It was found that treatment with
butein resulted in a significant increase in the activities of
caspase-3, -8 and -9 of HeLa cells in a dose-dependent manner
(P<0.05, Fig. 3A–C).
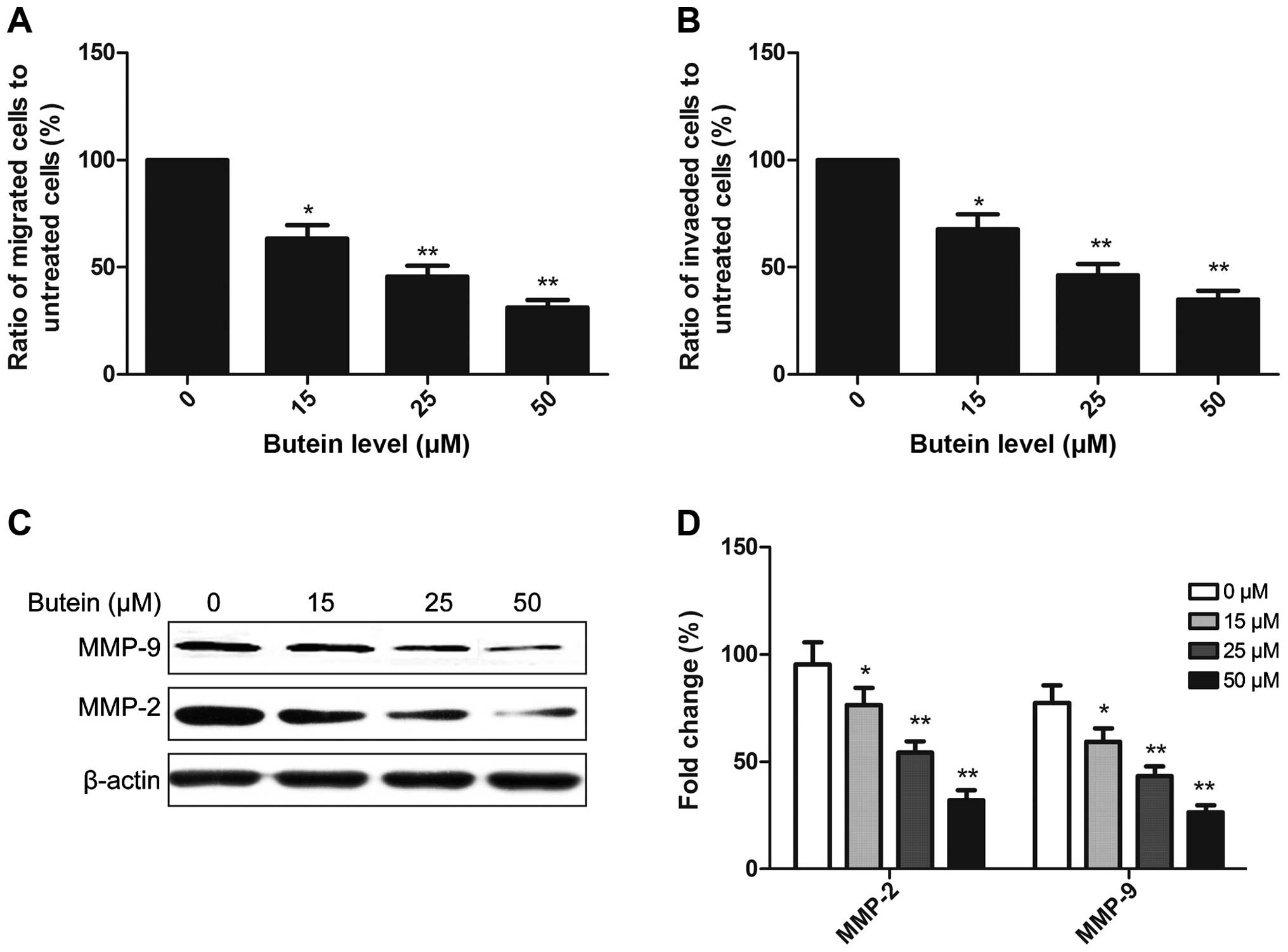
Butein decreases cell migration and
invasion of HeLa cells
To investigate the effect of butein on migration and
invasion of HeLa cells, the cells were seeded on Transwell with
uncoated (for migration) or Matrigel-coated (for invasion) filters.
After 48 h of treatment, we examined the migration activity and
invasive potential of HeLa cells, and found that butein at 15, 25
or 50 μM significantly inhibited the migration and invasion
of HeLa cells (Fig. 4A and B).
These results suggested that butein suppressed the migration and
invasion of HeLa cells in a dose-dependent manner.
To determine the potential mechanism of butein
effect on cell migration and invasion, MMP-2 and MMP-9 expression
was determined by western blotting following treatment with butein.
It was found that butein significantly reduced MMP-2 and MMP-9
expression of HeLa cells in a dose-dependent manner (Fig. 4C and D).
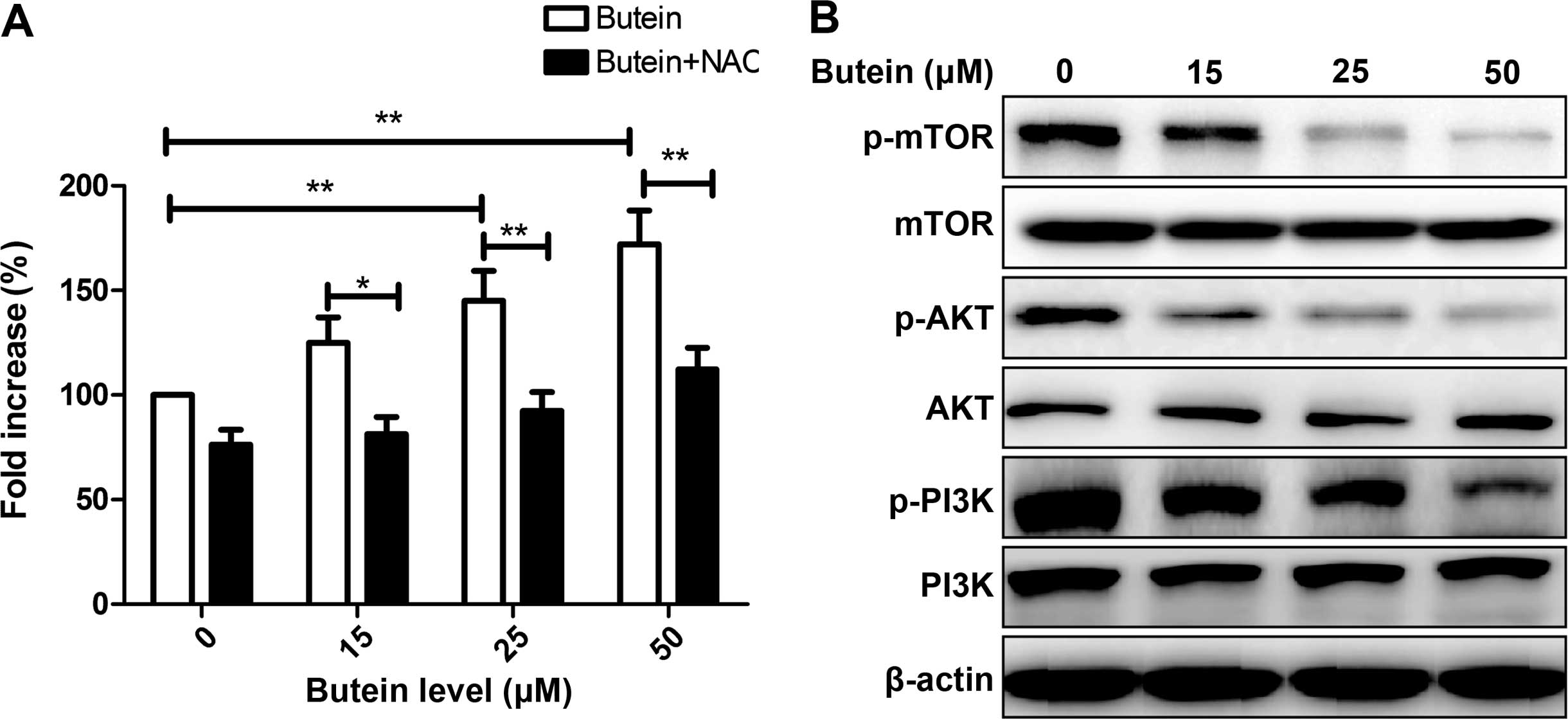
Effect of butein on ROS generation and
the PI3K/AKT/mTOR pathway of HeLa cells
Previous findings have shown that generation of ROS
plays an important role in regulating cell apoptosis and cell cycle
(27,32). To determine whether butein affected
the ROS level, ROS generation was determined using the DCFDA
staining method following treatment with the indicated
concentrations of butein. As shown in Fig. 5A, treatment with butein
significantly induced ROS generation in HeLa cells in a
dose-dependent manner. ROS generation was reduced by pretreatment
with the antioxidant agent NAC (10 mM) (Fig. 5A).
It is well known that the PI3K/AKT/mTOR signaling
pathways play keys role in cell proliferation, survival, motility
and angiogenesis. We evaluated the effect of butein on this
signaling pathway. Measurements of the phosphorylation/activation
pattern of PI3K, AKT and mTOR were carried out by western blotting
24 h after treatment with the indicated concentration of butein.
Treatment of HeLa cells with butein caused a dose-dependent
inhibition in the expression of phosphorylated PI3K, AKT and mTOR,
while the total protein levels of PI3K, AKT and mTOR in different
concentrations of butein group were not altered (Fig. 5B).
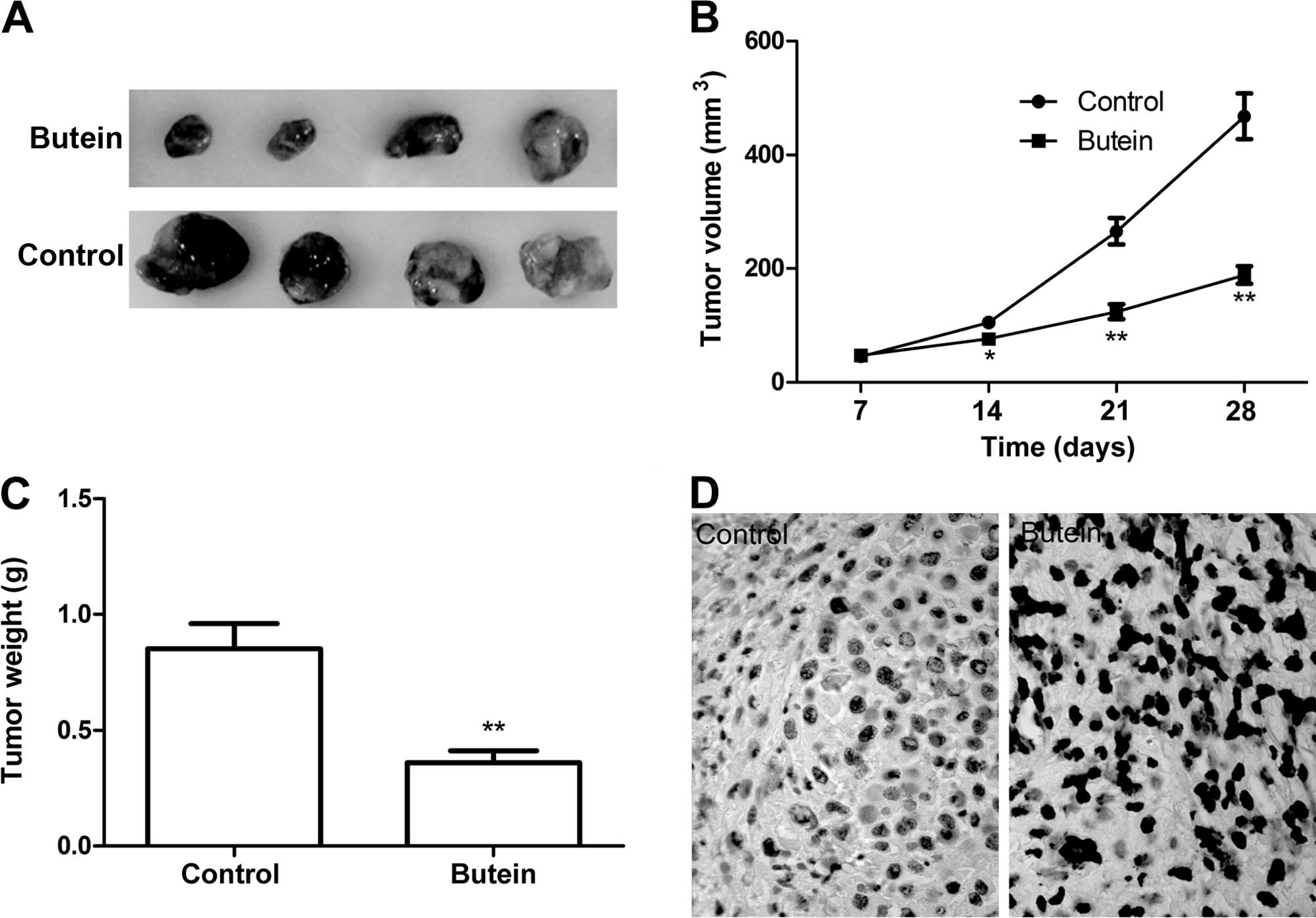
Butein inhibits cervical tumor growth in
a xenograft mouse model
We performed a HeLa xenograft mouse study to test
the effectiveness of butein in preventing tumor growth. Results
showed that treatment with butein (5 mg/kg) on alternate days for 4
weeks caused a significant inhibition of HeLa tumor xenograft
growth in the butein treatment group compared to the control group
(untreated group) (Fig. 6A–C). We
did not observe any gross signs of toxicity and/or possible
side-effects/mortality during the butein treatment relative to the
control group, suggesting butein is a safe anticancer agent.
In addition, we determined cell apoptosis of tumor
tissue by TUNEL. The results of TUNEL showed that butein obviously
induced the cell apoptosis of tumor tissue compared to the control
group (Fig. 6D). These data
suggested that butein treatment suppressed the tumor growth of
cervical cancer in vivo.
Discussion
Cervical cancer is the fourth leading cause of
cancer-associated mortality in women worldwide (1,2). The
development of novel strategies and anticancer drugs with fewer
side effects for cervical cancer therapy is crucial. Butein is a
polyphenol and belongs to one of the chalcone flavonoid subgroups.
Chalcones including butein have been approved for clinical trials
for the treatment of cancer and viral and cardiovascular diseases,
and findings suggest that these compounds reached reasonable plasma
concentrations without causing toxicity (28,33).
Butein is one of the compound of chalcones that possess anticancer
abilities against varying human cancer cells (19–27).
In the present study, the anticancer activities of butein were
investigated in a cervical cancer model in vitro and in
vivo. The in vitro studies demonstrate that butein
provides an effective therapeutic approach for cervical cancer
since butein significantly inhibits proliferation, migration and
invasion, and induced tumor apoptosis in HeLa cells in
dose-dependent manners. The in vivo mouse models also
confirmed that butein suppresses tumor growth with minor side
effects in the nude mouse model. These findings suggest that butein
serves as a potential therapeutic agent for the treatment of
cervical cancer.
The balance of expression of pro- and anti-apoptotic
members of the Bcl-2 family of proteins has been shown to play an
important role in apoptosis (34).
An increased the ratio of pro-apoptotic (Bax) to anti-apoptotic
(Bcl-2) proteins altered the membrane potential of the
mitochondria, released apoptogenic factors to the cytoplasma,
activated the caspase cascade and eventually lead to cell apoptosis
(34,35). It has been reported that butein
induced cell apoptosis via the upregulation of Bax expression and
downregulation of Bcl-2 expression in neuroblastoma cells, and
activation of caspase-3 and cleavage of PARP (27). Caspases are known for being
activated during apoptosis in a self-amplifying cascade, playing a
crucial role in the execution of apoptosis (23). In the present study, we found that
butein significantly induced cell apoptosis of HeLa cells, and
increased caspase-3, -8 and -9 activity, and upregulated the
Bax/Bcl-2 ratio suggesting butein induces cell apoptosis by
regulating the Bax/Bcl-2 ratio and caspase activity.
Alteration of intracellular reactive oxygen species
(ROS) production modulates several physiological functions
including stimulating mitotic cell division, regulating cell
apoptosis and inducing cell senescence (36). It has been demonstrated that
elevated ROS binds to lipids, proteins or DNA to produce oxidative
stress and even tually cause cell death (37). Several studies have shown that
butein induced apoptosis in cancer cells via the elevation of
intracellular ROS levels. For example, Moon et al (32) showed that butein induces ROS
generation, modulates JNK, ATM and Chk activity, and causes G2/M
arrest in hepatoma cells. Yang et al (38) reported that butein induced apoptosis
through a generation of ROS and deregulation of ERK1/2 and p38MAPK
in triple-negative [triple-negative breast cancer (TNBC)]
MDA-MB-231 cells. Chen et al (27) demonstrated that butein-triggered
neuroblastoma cells undergo apoptosis via the generation of ROS,
alteration of the Bcl-2/Bax ratio, and cleavage of pro-caspase-3
and PARP. Consistent with these studies, the present study showed
that butein inhibited cervical cancer cell growth and induced cell
apoptosis through the generation of ROS.
PI3K/Akt is major signal transduction pathway
regulating cell apoptosis, proliferation, differentiation and
metastasis (39). The constitutive
activation of the PI3K/Akt signaling pathway has been shown to
function as a major determinant of tumor cell growth and survival
in a number of solid tumors (40).
Downstream in the PI3K/Akt pathway there is a distal component
known as mTOR (39). Treatment of
human PCa cells with butein resulted in a decrease in the
expression of phosphorylation PI3K (p-PI3K) and phosphorylation of
Akt (p-AKT) (22). Liu et al
found that butein inhibited the Akt/mTOR/p70S6K translational
machinery, leading to the downregulation of MMP-9 and uPA, and
subsequently exerting an antimetastatic effect (41). In the present study, our results
showed that butein inhibited p-PI3K, p-AKT and p-mTOR, suggesting
that butein suppressed cervical cancer growth, at least partially
by regulating the PI3K/AKT/mTOR signaling pathway.
In conclusion, to the best of our knowledge, this is
the first study to show that butein inhibits cell viability, colony
formation, migration and invasion, and induced G2/M phase arrest in
HeLa cells. Our results also show that butein significantly
induced-HeLa cell apoptosis by regulating the Bcl-2/Bax ratio, and
increasing caspase-3, -8 and -9 activity. The in vivo
results showed that butein suppressed tumor growth in a xenograft
mouse model. In addition, butein increased the generation of ROS,
and inhibited PI3K/AKT/mTOR pathway activation, which may
contribute to a decrease in oxidative stress and suppression of
tumor growth. Collectively, our findings suggest that butein serves
as a chemotherapeutic agent for the future treatment of cervical
cancer.
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Martin-Hirsch PL and Wood NJ: Cervical
cancer. BMJ Clin Evid. Jul 27–2011.0818PubMed/NCBI
|
|
3
|
Duenas-Gonzalez A, Serrano-Olvera A,
Cetina L and Coronel J: New molecular targets against cervical
cancer. Int J Womens Health. 6:1023–1031. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
de Freitas AC, Gomes Leitão Mda C and
Coimbra EC: Prospects of molecularly-targeted therapies for
cervical cancer treatment. Curr Drug Targets. 16:77–91. 2015.
View Article : Google Scholar
|
|
5
|
Jakubowicz J, Blecharz P, Skotnicki P,
Reinfuss M, Walasek T and Luczynska E: Toxicity of concurrent
chemoradiotherapy for locally advanced cervical cancer. Eur J
Gynaecol Oncol. 35:393–399. 2014.PubMed/NCBI
|
|
6
|
Bazaeva IIa, Gorbunova VA, Kravets OA,
Khokhlova SV, Limareva SV, Panov VO, Strel’tsova ON and Tarachkova
EV: Chemoradiotherapy for locally advanced cervical cancer. Vopr
Onkol. 60:280–287. 2014.In Russian.
|
|
7
|
Butler MS: The role of natural product
chemistry in drug discovery. J Nat Prod. 67:2141–2153. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Paterson I and Anderson EA: Chemistry. The
renaissance of natural products as drug candidates. Science.
310:451–453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tsuda H, Ohshima Y, Nomoto H, Fujita K,
Matsuda E, Iigo M, Takasuka N and Moore MA: Cancer prevention by
natural compounds. Drug Metab Pharmacokinet. 19:245–263. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nobili S, Lippi D, Witort E, Donnini M,
Bausi L, Mini E and Capaccioli S: Natural compounds for cancer
treatment and prevention. Pharmacol Res. 59:365–378. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huang WY, Cai YZ and Zhang Y: Natural
phenolic compounds from medicinal herbs and dietary plants:
Potential use for cancer prevention. Nutr Cancer. 62:1–20. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
McCarty MF: Current prospects for
controlling cancer growth with non-cytotoxic agents - nutrients,
phytochemicals, herbal extracts, and available drugs. Med
Hypotheses. 56:137–154. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Lazzeroni M, Gandini S, Puntoni M, Bonanni
B, Gennari A and DeCensi A: The science behind vitamins and natural
compounds for breast cancer prevention. Getting the most prevention
out of it. Breast. 20(Suppl 3): S36–S41. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moon DO, Choi YH, Moon SK, Kim WJ and Kim
GY: Butein suppresses the expression of nuclear factor-kappa
B-mediated matrix metalloproteinase-9 and vascular endothelial
growth factor in prostate cancer cells. Toxicol In Vitro.
24:1927–1934. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kojima R, Kawachi M and Ito M: Butein
suppresses ICAM-1 expression through the inhibition of IκBα and
c-Jun phosphorylation in TNF-α- and PMA-treated HUVECs. Int
Immunopharmacol. 24:267–275. 2015. View Article : Google Scholar
|
|
16
|
Rasheed Z, Akhtar N, Khan A, Khan KA and
Haqqi TM: Butrin, isobutrin, and butein from medicinal plant Butea
monosperma selectively inhibit nuclear factor-kappaB in activated
human mast cells: Suppression of tumor necrosis factor-alpha,
interleukin (IL)-6, and IL-8. J Pharmacol Exp Ther. 333:354–363.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee SH, Seo GS and Sohn DH: Inhibition of
lipopolysaccha-ride-induced expression of inducible nitric oxide
synthase by butein in RAW 264.7 cells. Biochem Biophys Res Commun.
323:125–132. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Chan SC, Chang YS, Wang JP, Chen SC and
Kuo SC: Three new flavonoids and antiallergic, anti-inflammatory
constituents from the heartwood of Dalbergia odorifera. Planta Med.
64:153–158. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Yit CC and Das NP: Cytotoxic effect of
butein on human colon adenocarcinoma cell proliferation. Cancer
Lett. 82:65–72. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li Y, Ma C, Qian M, Wen Z, Jing H and Qian
D: Butein induces cell apoptosis and inhibition of cyclooxygenase-2
expression in A549 lung cancer cells. Mol Med Rep. 9:763–767.
2014.
|
|
21
|
Cui Z, Song E, Hu DN, Chen M, Rosen R and
McCormick SA: Butein induces apoptosis in human uveal melanoma
cells through mitochondrial apoptosis pathway. Curr Eye Res.
37:730–739. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Khan N, Adhami VM, Afaq F and Mukhtar H:
Butein induces apoptosis and inhibits prostate tumor growth in
vitro and in vivo. Antioxid Redox Signal. 16:1195–1204. 2012.
View Article : Google Scholar :
|
|
23
|
Kim NY, Pae HO, Oh GS, Kang TH, Kim YC,
Rhew HY and Chung HT: Butein, a plant polyphenol, induces apoptosis
concomitant with increased caspase-3 activity, decreased Bcl-2
expression and increased Bax expression in HL-60 cells. Pharmacol
Toxicol. 88:261–266. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Wang Y, Chan FL, Chen S and Leung LK: The
plant polyphenol butein inhibits testosterone-induced proliferation
in breast cancer cells expressing aromatase. Life Sci. 77:39–51.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jang HS, Kook SH, Son YO, Kim JG, Jeon YM,
Jang YS, Choi KC, Kim J, Han SK, Lee KY, et al: Flavonoids purified
from Rhus verniciflua Stokes actively inhibit cell growth and
induce apoptosis in human osteosarcoma cells. Biochim Biophys Acta.
1726:309–316. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rajendran P, Ong TH, Chen L, Li F,
Shanmugam MK, Vali S, Abbasi T, Kapoor S, Sharma A, Kumar AP, et
al: Suppression of signal transducer and activator of transcription
3 activation by butein inhibits growth of human hepatocellular
carcinoma in vivo. Clin Cancer Res. 17:1425–1439. 2011. View Article : Google Scholar
|
|
27
|
Chen YH, Yeh CW, Lo HC, Su SL, Hseu YC and
Hsu LS: Generation of reactive oxygen species mediates
butein-induced apoptosis in neuroblastoma cells. Oncol Rep.
27:1233–1237. 2012.PubMed/NCBI
|
|
28
|
Yadav VR, Prasad S, Sung B and Aggarwal
BB: The role of chalcones in suppression of NF-κB-mediated
inflammation and cancer. Int Immunopharmacol. 11:295–309. 2011.
View Article : Google Scholar :
|
|
29
|
Kang DG, Lee AS, Mun YJ, Woo WH, Kim YC,
Sohn EJ, Moon MK and Lee HS: Butein ameliorates renal concentrating
ability in cisplatin-induced acute renal failure in rats. Biol
Pharm Bull. 27:366–370. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee SH, Choi WC, Kim KS, Park JW, Lee SH
and Yoon SW: Shrinkage of gastric cancer in an elderly patient who
received Rhus verniciflua Stokes extract. J Altern Complement Med.
16:497–500. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Hamzeloo-Moghadam M, Aghaei M, Fallahian
F, Jafari SM, Dolati M, Abdolmohammadi MH, Hajiahmadi S and
Esmaeili S: Britannin, a sesquiterpene lactone, inhibits
proliferation and induces apoptosis through the mitochondrial
signaling pathway in human breast cancer cells. Tumour Biol.
36:1191–1198. 2015. View Article : Google Scholar
|
|
32
|
Moon DO, Kim MO, Choi YH, Hyun JW, Chang
WY and Kim GY: Butein induces G2/M phase arrest and
apoptosis in human hepatoma cancer cells through ROS generation.
Cancer Lett. 288:204–213. 2010. View Article : Google Scholar
|
|
33
|
Opletalova V, Jahodar L, Jun D and Opletal
L: Chalcones (1,3-diarylpropen-1-ones) and their analogs as
potential therapeutic agents in cardiovascular system diseases.
Ceska Slov Farm. 52:12–19. 2003.
|
|
34
|
Weerasinghe P, Hallock S, Tang SC and
Liepins A: Role of Bcl-2 family proteins and caspase-3 in
sanguinarine-induced bimodal cell death. Cell Biol Toxicol.
17:371–381. 2001. View Article : Google Scholar
|
|
35
|
Cory S and Adams JM: The Bcl2 family:
Regulators of the cellular life-or-death switch. Nat Rev Cancer.
2:647–656. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
36
|
Valko M, Leibfritz D, Moncol J, Cronin MT,
Mazur M and Telser J: Free radicals and antioxidants in normal
physiological functions and human disease. Int J Biochem Cell Biol.
39:44–84. 2007. View Article : Google Scholar
|
|
37
|
Ali-Seyed M, Bhuvaneswari R, Soo KC and
Olivo M: Photolon™ - Photosensitization induces apoptosis via
ROS-mediated crosstalk between mitochondria and lysosomes. Int J
Oncol. 39:821–831. 2011.PubMed/NCBI
|
|
38
|
Yang LH, Ho YJ, Lin JF, Yeh CW, Kao SH and
Hsu LS: Butein inhibits the proliferation of breast cancer cells
through generation of reactive oxygen species and modulation of ERK
and p38 activities. Mol Med Rep. 6:1126–1132. 2012.PubMed/NCBI
|
|
39
|
Fresno Vara JA, Casado E, de Castro J,
Cejas P, Belda-Iniesta C and González-Barón M: PI3K/Akt signalling
pathway and cancer. Cancer Treat Rev. 30:193–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chen YL, Law PY and Loh HH: Inhibition of
PI3K/Akt signaling: An emerging paradigm for targeted cancer
therapy. Curr Med Chem Anticancer Agents. 5:575–589. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu SC, Chen C, Chung CH, Wang PC, Wu NL,
Cheng JK, Lai YW, Sun HL, Peng CY, Tang CH, et al: Inhibitory
effects of butein on cancer metastasis and bioenergetic modulation.
J Agric Food Chem. 62:9109–9117. 2014. View Article : Google Scholar : PubMed/NCBI
|