Introduction
Arsenic and arsenic-containing compounds are widely
distributed in the environment and exist in organic and inorganic
forms. Although a well-known poison, arsenic has been used
medicinally for over 2,000 years (1). In particular, administration of
arsenic trioxide (arsenite, AsIII), an arsenic
derivative, has demonstrated a remarkable efficacy in the treatment
of relapsed and refractory acute promyelocytic leukemia (APL)
(1–4). Detailed pharmacokinetic studies of
AsIII in APL patients have been carried out to optimize
treatment (3,5–8). Both
inorganic arsenic and methylated arsenic metabolites accumulate in
red blood cells during repeated administration of AsIII
to APL patients (7). Arsenic
metabolites are also detected in cerebrospinal fluid (6) at concentrations of arsenic necessary
for induction of differentiation (4,9).
Recent data from our laboratory demonstrated that the profiles of
arsenic species in peripheral blood (PB) plasma were very similar
to those of bone marrow (BM) plasma, suggesting that the profiles
of PB plasma could be predictive biomarkers for the treatment
outcome of APL patients (8). These
findings on the pharmacokinetics of AsIII in APL
patients provide new insight into clinical applications of
AsIII, and may contribute to designing better
therapeutic protocols (1).
Multidrug resistance is a major concern for the
clinical use of anticancer drugs. ATP-binding cassette (ABC)
transporters contribute to drug resistance via ATP-dependent drug
efflux (1,10). Multidrug resistance-associated
proteins 1 and 2 (MRP1/2), and multidrug resistance protein 1
(MDR1; also known as P-glycoprotein, P-gp) have been implicated in
the efflux of arsenic, and may contribute to resistance to arsenic
therapy (1,11,12).
Furthermore, we recently demonstrated the MRP2 and aquaporin-9
(AQP9), a member of the aquaporin superfamily, involvement in
arsenic uptake (1,13–15),
contributing to the differential sensitivity of primary
human-derived normal cells to arsenite (14,16–18).
Although these previous findings provide fundamental knowledge for
understanding the cellular handling and elimination pathways of
arsenic in cancer cells as well as normal cells, more studies are
required to provide detailed information on the efficacy and safety
of arsenic for clinical use.
MRP4 transports antiviral agents such as the
nucleoside/nucleotide analogs azidothymidine (AZT), adefovir
[9-(2-phosphonylmethoxyethyl) adenine or PMEA] and tenofovir (TFV)
(19–21), and anticancer drugs such as
camptothecins and methotrexate (22–24).
Of note, recent studies have demonstrated that hematopoietic
progenitor cell differentiation affects expression and function of
MRP4, and that MRP4 has a relevant role in tumor growth and
apoptosis and in the eradication of leukemic stem cells, suggesting
MRP4 as a new potential therapeutic target for acute myeloid
leukemia (25,26). It is noteworthy that MRP4 is
localized to the basolateral membrane of hepatocytes and the apical
membrane of renal proximal tubule cells, distinguishing itself from
other members of the MRP family (21). Considering the localization of MRP4,
biomethylation of arsenic primarily in liver and elimination of
arsenic by the kidney (1), MRP4 is
an ideal candidate for the elimination of arsenic and may
contribute to resistance to AsIII. Only recently have
studies been reported regarding the role of MRP4 in arsenic
resistance (27,28). In the present study, stable human
embryonic kidney epithelial (HEK)293 cells overexpressing MRP4 and
MRP2 were created and used to investigate the cytotoxicity of
AsIII against both MRP overexpressing cells and
reference cells transfected with an empty vector, in order to
clarify whether MRP4 cells have the capacity to confer drug
resistance to AsIII.
Materials and methods
Reagents
Sodium arsenite (AsIII) was purchased
from Tri Chemical Laboratories (Yamanashi, Japan). Cyclosporin A
(CsA), a broad-spectrum inhibitor of ABC transporters, was kindly
provided by Professor Toshihiko Hirano, Department of Clinical
Pharmacology, School of Pharmacy, Tokyo University of Pharmacy and
Life Sciences. 3-[[3-[2-(7-chloroquinolin-2-yl)
vinyl]phenyl]-(2-dimethylcarbamoylethylsulfanyl)methylsulfanyl]
propionic acid (MK571), an inhibitor of MRP4, was purchased from
Merck (Darmstadt, Germany).
Construction of stable HEK293 cell lines
expressing human MRP2 or MRP4 cDNA
Human ABCC2 or ABCC4 cDNA was subcloned into the
pcDNA5/FRT vector (Invitrogen, Carlsbad, CA, USA) as described
previously (19,20). Briefly, HEK293 Flp-In cells were
seeded at 5×105 cells/well in 6-well plates in
Dulbecco's modified Eagle's medium (DMEM) with 10% fetal bovine
serum (FBS) without antibiotics. After 24 h of cultivation, 0.4
µg MRP2 or MRP4 plasmid was transfected into HEK293 Flp-In
cells using Lipofectamine 2000 (Invitrogen) according to the
manufacturer's protocol. Empty pcDNA/FRT vector was transfected as
a negative control (empty vector cells). DMEM fresh media were
added at 6 h post-transfection. The selection of stable cell clones
expressing MRP2 or MRP4 reference plasmid was started in 75
µg/ml hygromycin (Invitrogen) the following day. Media were
changed every 2–3 days, and selection of stable transfectants
generally took 10–14 days. HEK293 cells stably expressing MRP2
reference (MRP2 cells) or MRP4 reference (MRP4 cells) and empty
vector cells were cultured in DMEM supplemented with 10% FBS, 100
U/ml of penicillin and 100 µg/ml of streptomycin at 37°C in
a humidified atmosphere (5% CO2 in air).
MRP2 protein isolation and western blot
analysis
Proteins were isolated using PARIS™ kit (Ambion Life
Technologies, Foster City, CA, USA) according to the manufacturer's
protocol. Proteins (30 µg/lane) were separated on 4–15%
Tris-HCl Criterion gels (Bio-Rad, Hercules, CA, USA) by SDS-PAGE at
80 V and then transferred onto nitrocellulose membranes at 250 mA
for 2 h. Membranes were blocked in 2% skim milk for 1 h at room
temperature and incubated overnight at 4°C with anti-MRP2 antibody
(M2-III-6) at 1:250 dilution (Thermo Fisher Scientific, Rockford,
IL, USA) or anti-GAPDH antibody (ab9483) at 1:100,000 dilution
(Abcam, Cambridge, MA, USA). The following day, membranes were
washed three times with phosphate-buffered saline (PBS) and then
incubated with IRdye 800CW goat anti-mouse fluorescent secondary
antibody (Li-COR, Lincoln, NE, USA) at 1:10,000 dilution. Membranes
were washed with PBS and visualized on an odyssey® Sa
Infrared Imaging system (Li-COR).
Evaluation of MRP2 membrane expression by
immunocytochemistry
Empty vector and MRP2 reference cells were seeded at
2.5×104/chamber on 4-chamber slides 24 h prior to
staining. The following day cells were washed with cold PBS two
times and permeabilized with cold acetone for 10 min on ice. An
equal volume of 8% paraformaldehyde was added directly to the cells
and incubated for 10 min at room temperature. Cells were washed
with PBS two times and blocked with 2 mg/ml BSA solution for 30 min
at room temperature. Primary anti-MRP2 (M2-III-6) antibody was
added to BSA solution to the final dilution of 1:20. After three
washes with PBS, the slides were incubated with a fluorescently
conjugated secondary antibody (Alexa Fluor® 488; Thermo
Fisher Scientific) for 1 h at room temperature protected from
light. DAPI (Thermo Fisher Scientific) stain at 1:3,000 dilution
was added 20 min before the end of incubation. After three washes
with PBS, images were mounted and then captured using a Retiga
CCD-cooled camera and associated QCapture Pro software (QImaging,
Surrey, BC, Canada).
Functional assays of MRP2 cells
Evaluation of MRP2 transport activity was performed
using a carboxyfluorescein (CF) fluorescent dye retention assay. CF
was applied to cells in its di-acetate (CFDA) form (Sigma-Aldrich,
St. Louis, MO, USA) which is non fluorescent and highly lipophilic;
it freely enters the cells through passive diffusion. Once inside
the cell the acetate groups of CFDA are cleaved by esterases
yielding fluorescent CF, which has low permeability characteristics
and is a substrate for ABC efflux transporters, including MRP2
(29). Briefly, empty vector and
MRP2 overexpressing cells were trypsinized, washed with warm PBS
and re-suspended at a cell density of 1×106 cells/ml in
fresh DMEM without FBS. Cells were then incubated for 30 min at
37°C with 10 µM of CFDA. Following accumulation, cells were
pelleted, washed twice with warm PBS, re-suspended in DMEM media
supplemented with 10% FBS and allowed to efflux for 30 min at 37°C.
Following efflux, cells were pelleted, washed twice with ice-cold
PBS and then re-suspended in ice-cold PBS supplemented with 10% FBS
for analysis. Retention of CF was determined by measuring
fluorescence using flow cytometry performed on a BD FACSCalibur (BD
Biosciences, Mississauga, ON, Canada). Briefly, 10,000 cell events
were collected for each sample. Cells were co-stained with
propidium iodide (PI; Thermo Fisher Scientific) to exclude
non-viable cells from further analysis. CF fluorescence was
measured in FL-1 channel (excitation wavelength 488 nm and emission
wavelength 530 nm), and PI fluorescence was measured in FL-3
channel (excitation wavelength 488 nm and emission wavelength 600
nm). Each experiment was repeated four times. Data were normalized
to empty vector transfected cells and a t-test was used on
normalized data to test for differences in MRP2 overexpressing
cells and reference cells.
Functional assays of MRP4 cells
Transport assays for MRP4 were carried out using two
reported MRP4 substrates [adenine-8-3H-tenofovir
disoproxil (TFV) (3.8 Ci/mmol, 98.1% purity);
3H-9-(2-phosphonylmethoxyethyl)-adenine (PMEA)] (Moravek
Biochemicals, Brea, CA, USA) as described previously (19,20,30).
Briefly, MRP4 and empty vector cells were seeded at
2.5×105 cells/well in triplicate in poly-D-lysine-coated
24-well plates (BD Biosciences, San Jose, CA, USA). After
preincubation for 24 h, cells were incubated with 1 µM TFV
or 100 nM PMEA in glucose-free DMEM supplemented with 10 µM
NaN3 and 10 µM 2-deoxy-D-glucose, respectively,
for 2 h at 37°C. After accumulation, cells were washed with
ice-cold PBS and supplemented with complete DMEM. Supernatant
fractions were collected at 0, 30 and 90 min, and cells were washed
and lysed with 800 µl/well of an aqueous solution of 10%
sodium dodecyl sulfate and 1 N NaOH. Supernatants were added to
Ecolite scintillation fluid (MP Biomedicals, Santa Ana, CA, USA)
and extracellular amounts of TFV and PMEA were analyzed by
scintillation counting. The remaining cell lysate was used to
determine the protein concentration with a BCA™ Protein Assay kit
(Pierce Biotechnology, Rockford, IL, USA); TFV and PMEA levels were
normalized to protein concentrations.
Cell viability assay
The cytotoxicity of AsIII to MRP4, MRP2
and empty vector cells was investigated by XTT dye-reduction assays
according to the method previously described with slight
modifications (14,31). Briefly, the cells were seeded in
96-well plates (Iwaki, Tokyo, Japan) at a density of
5×103 cells/well in 0.1 ml complete DMEM and cultivated
for 24 h. Cultures in triplicate were treated with various
concentrations of AsIII in the presence or absence of
transporter inhibitors at the concentrations indicated. Cells were
pre-incubated with transporter inhibitors at the indicated
concentrations for 30 min. After treatment with AsIII
for an additional 48 h,
2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)-5-[(phenylamino)carbonyl]-2H-tetrazolium
hydroxide (XTT; Sigma, MD, USA) and phenazine methosulfate (Wako
Pure Chemical Industries, Osaka, Japan) were added into each well
at final concentrations of 0.2 mg/ml and 1 mM, respectively. After
incubation at 37°C for 4 h, the plates were mixed, and the
absorbance at 450 nm was measured with a microplate reader (Safire,
Tecan, Switzerland). The relative cell viability was expressed as
the ratio of the absorbance of each treatment group against those
of the corresponding untreated control group. Data are shown as
means ± SD from more than five independent experiments. The
IC50 values of AsIII for all three cell types
were calculated using GraphPad Prism® 5 software.
Statistical analysis
Data were analyzed using Student's t-test and ANOVA
with a Dunnett's post test method. A p-value <0.05 was
considered as statistically significant.
Results
Confirmation of MRP2 expression and
transporter activity in the MRP2 stable cell line
Protein expression of MRP2 was confirmed using
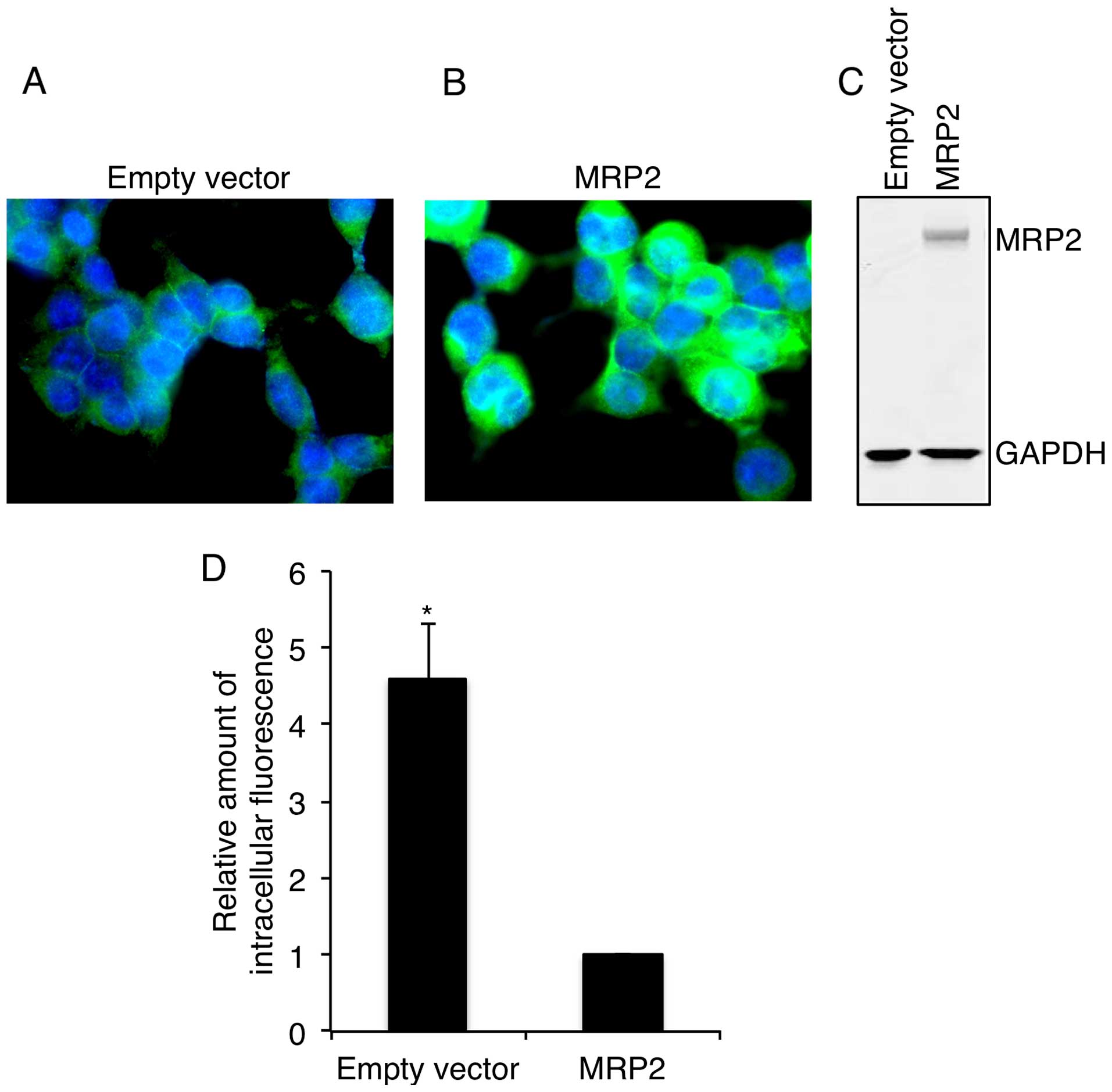
immunocytochemistry and western blotting. As shown in Fig. 1, MRP2 cells had significantly higher
levels of membrane (Fig. 1A and B)
and total (Fig. 1C) MRP2 expression
compared to empty vector transfected cells. Functional activity of
MRP2 was confirmed in MRP2 reference cells using a CF retention
assay. As shown in Fig. 1D,
retention of CF in MRP2 overexpressing cells was 4.5-fold lower
than that in empty vector cells, indirectly confirming high efflux
of fluorescent dye from MRP2 overexpressing cells and, therefore,
providing strong evidence for MRP2 functional activity.
Confirmation of transporter activity in
the MRP4 stable cell line
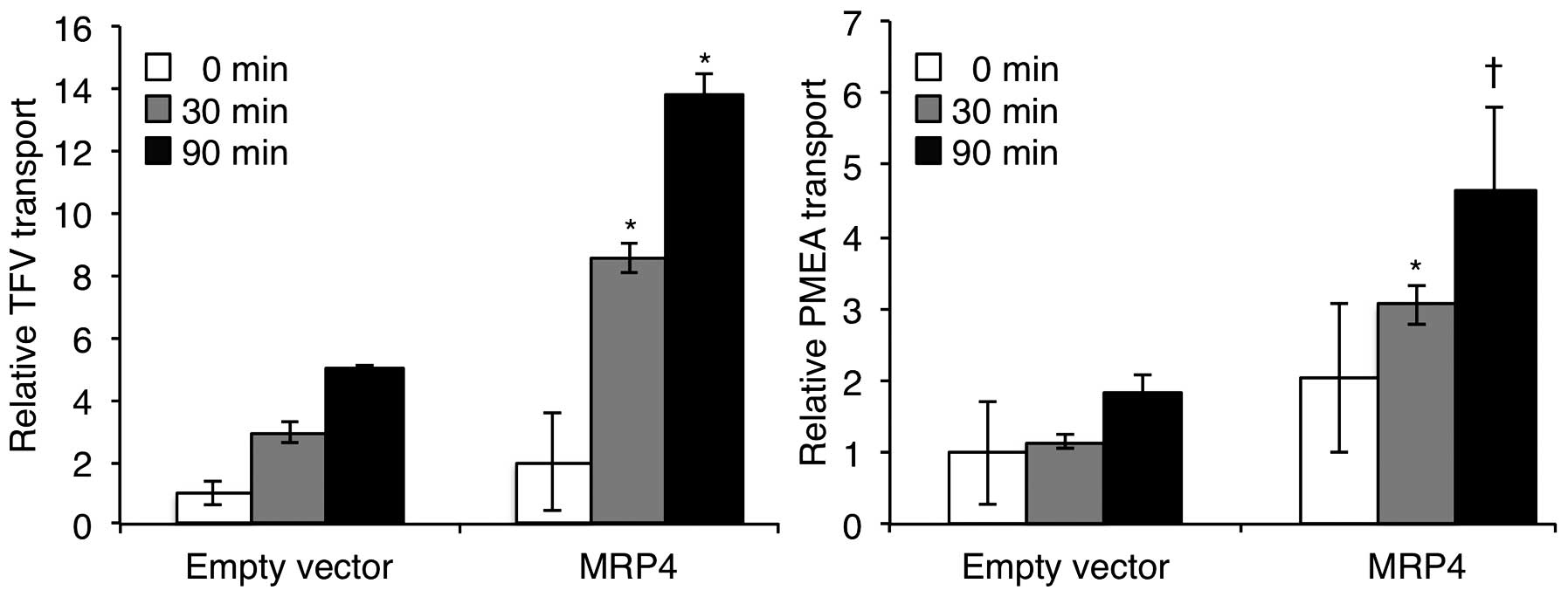
Accumulation of TFV or PMEA in the supernatant
fractions of each cell type was assessed for confirmation of MRP4
function. As shown in Fig. 2, the
accumulation of TFV or PMEA in the supernatant of MRP4 cells
increased with time, and was much higher than that of empty vector
cells. Compared to empty vector cells, an approximately 3-fold and
2.7-fold increase in the efflux of TFV and PMEA, respectively, was
observed in the MRP4 cells at the 30 and 90 min time-points
(Fig. 2), indicating functional
MRP4.
AsIII-induced cytotoxicity in
MRP4 and MRP2 cells
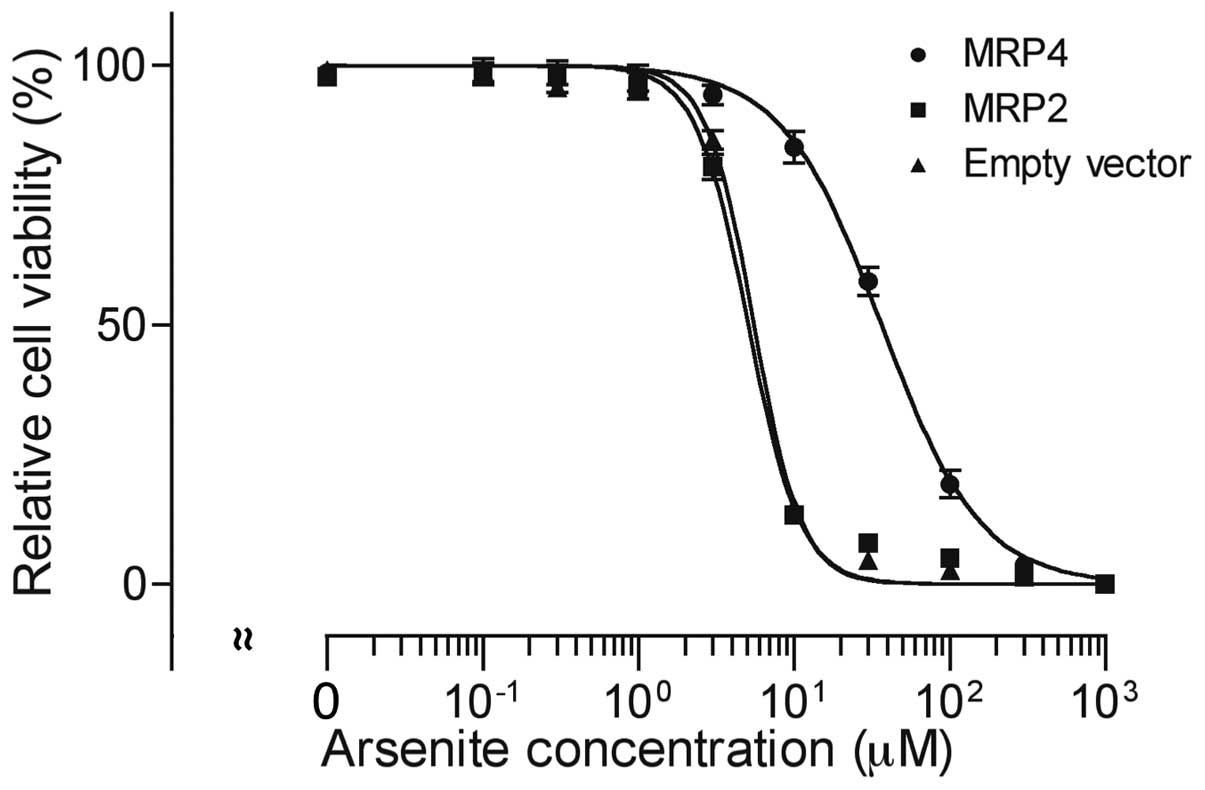
After treatment with 0, 0.1, 0.3, 1, 3, 10, 30, 100,
300 and 1,000 µM of AsIII for 48 h, cell
viability was investigated by XTT assay. A significant
dose-dependent decrease in cell viability was observed in all three
cell types (Fig. 3).
AsIII exhibited much lower cytotoxicity in MRP4 cells in
comparison with MRP2 and empty vector cells. The IC50
values of AsIII were 36.6±8.1, 5.1±0.3 and 5.5±0.3 µM in
MRP4, MRP2 and empty vector cells (MRP4 vs. empty vector,
P<0.01; MRP2 vs. empty vector, P>0.05), respectively.
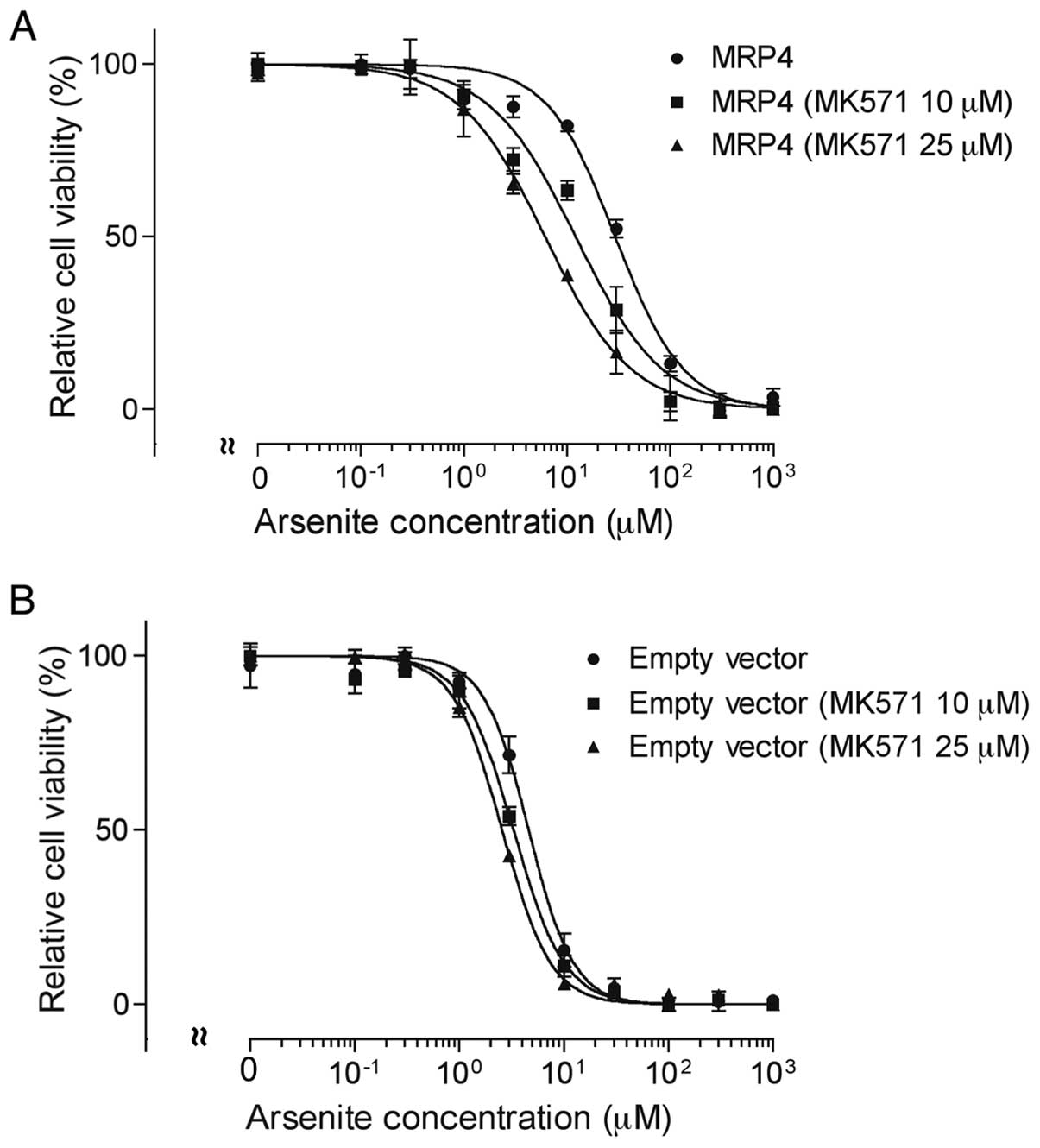
Effect of MK571 on
AsIII-induced cytotoxicity in MRP4 and empty vector
cells
Both MRP4 and empty vector cells were exposed to
various concentrations of AsIII (0–1,000 µM) in
the presence or absence of 10 or 25 µM MK571 for 48 h,
followed by the assessment of cell viability. The IC50
value of AsIII in MRP4 cells decreased significantly
from 28.2±3.3 to 11.2±3.3 and 6.3±1.0 µM by the addition of
10 and 25 µM MK571 (MRP4 without MK571 vs. MRP4 with 10 or
25 µM MK571, P<0.01), respectively (Fig. 4A). Interestingly, the addition of
MK571 also slightly enhanced AsIII-induced cytotoxicity
in empty vector cells, with the IC50 value of
AsIII decreasing from 4.8±0.8 to 3.3±0.4 and 2.5±0.2
µM by the addition of 10 and 25 µM MK571 (empty
vector cells without MK571 vs. empty vector cells with 10 or 25
µM MK571, P<0.05), respectively (Fig. 4B).
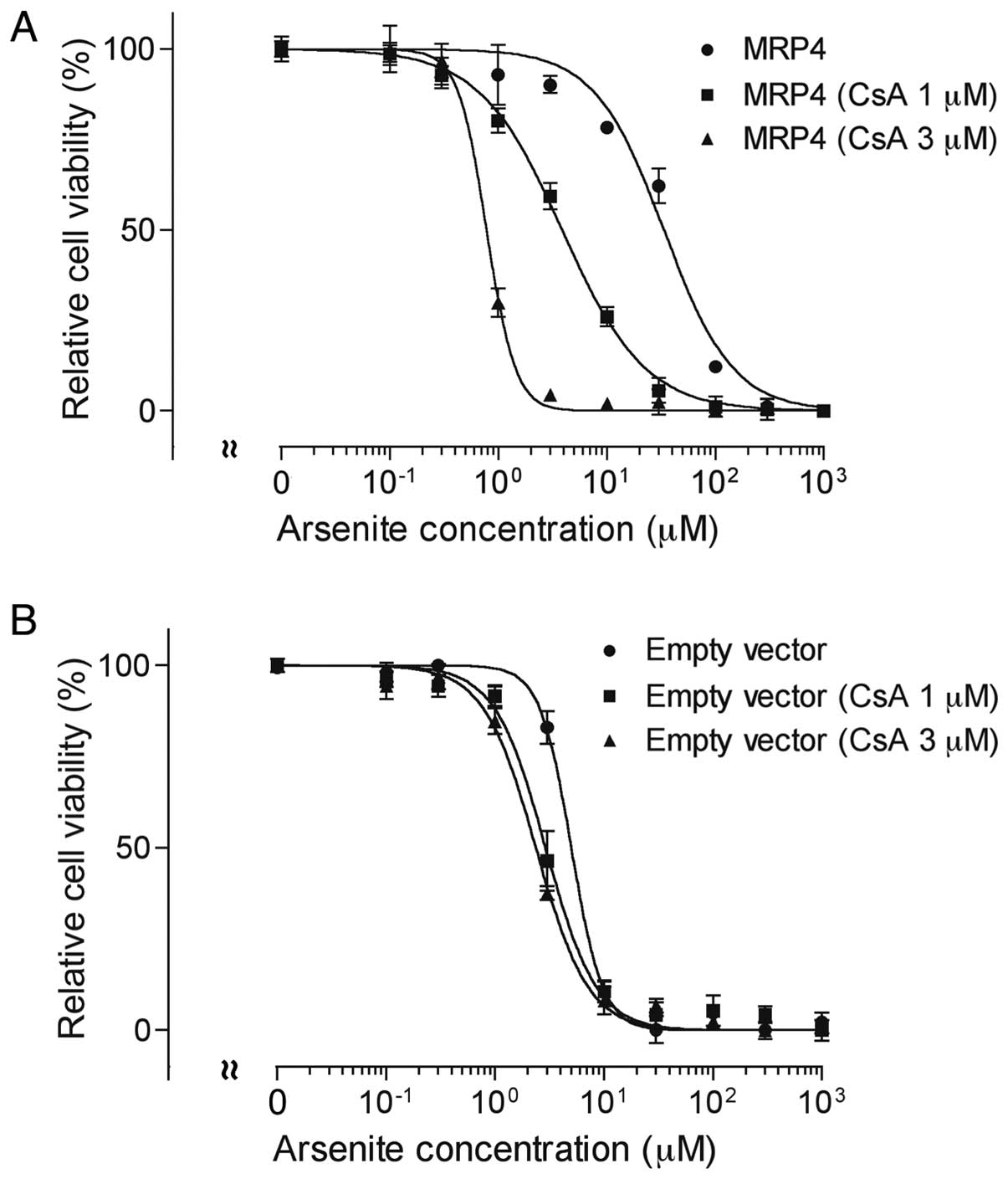
Effect of CsA on AsIII-induced
cytotoxicity in MRP4 and empty vector cells
Both MRP4 and empty vector cells were exposed to
various concentrations of AsIII (0–1,000 µM) in the presence
or absence of 1 or 3 µM CsA for 48 h, followed by the
assessment of cell viability. In comparison to MK571, CsA, a
broad-spectrum inhibitor of ABC transporters, more potently
inhibited MRP4 and increased AsIII-induced cytotoxicity
(Fig. 5A). The IC50
value of AsIII in MRP4 cells decreased ≥90% after the
addition of 1 and 3 µM CsA, respectively (Fig. 5A). There was some increase in
AsIII cytotoxicity as well in empty vector cells
(5.3±0.2 vs 3.1±0.6 and 2.3±0.2 µM) (empty vector cells
without CsA vs. empty vector cells with 1 or 3 µM CsA,
P<0.05) (Fig. 5B).
Discussion
MRP2 and MRP4 overexpressing cells were used in the
current study to evaluate the contribution of these ABC efflux
transporters to AsIII cytotoxicity. A 6-fold difference
in IC50 values of AsIII between these cells
and a significant reduction of the IC50 value in MRP4
cells provide strong support that MRP4 is a major mediator of
AsIII efflux from cells and plays a major role in
AsIII resistance. MRP2 has been demonstrated to be
involved in the efflux of arsenic, conferring resistance to
AsIII in other experimental systems (12,32,33).
Wild-type and MRP2-deficient Wistar (TR−) rats have been
used to show that MRP2 is responsible for the biliary excretion of
arsenic triglutathione [As(GS)3] and monomethyl arsenic
diglutathione [CH3As(GS)2] (33). These in vivo findings are
supported by cellular transport assays demonstrating that
As(GS)3 is also a substrate for human MRP2 (12). A recent in vitro study using
MRP2-enriched membrane vesicles demonstrated that MRP2 transports
selenobis(S-glutathionyl) arsiniumion
[((GS)2AsSe)−] (32). These previous findings suggest that
glutathione and/or the essential trace element selenium (Se) are
required for the excretion and detoxification of arsenic. In
contrast, using MRP2 overexpressing cells in the present study,
there was no evidence that MRP2 plays a critical role in
AsIII transport and cytotoxicity. These conflicting
results might reflect low levels of glutathione and/or Se
conjugation in MRP2 overexpressing HEK293 cells, although more
detailed analysis of the molecular events such as the amount of
glutathione and its conjugated arsenic compounds is required to
confirm this hypothesis.
Inhibition studies further support the role of MRP4
to confer resistance to AsIII. MK571, an MRP4 inhibitor
(21), was used to demonstrate that
MRP4 confers resistance to a series of camptothecin analogs,
including irinotecan and SN-38 (23,24).
Consistent with these previous studies, the addition of MK571
significantly potentiated AsIII-triggered cytotoxicity
in the MRP4 cells in a dose-dependent manner. CsA, a well-known
broad-spectrum inhibitor of ABC transporters, increased
AsIII-triggered cytotoxicity to a greater degree than
MK571, supporting a role for multiple ABC transporters in
AsIII cytotoxicity. Collectively, our experimental
results implicate MRP4 in resistance to AsIII.
Considering that multidrug resistance-reversing activity of CsA has
been reported in phase II studies with myeloma and acute leukemia,
increased recognition of drug-drug interactions is necessary for
optimal treatment of patients with arsenic-based regimens.
Our results are not completely in agreement with
other studies of arsenic resistance. Increased resistance was not
detected in MRP4-transfected NIH3T3 cells after exposure to sodium
meta-arsenite for 72 h (28).
Furthermore, a recent report demonstrated that after treatment with
increasing concentrations of inorganic and methylated species of
arsenic for 72 h, MRP4-transfected HEK293 cells conferred
resistance to arsenate and methylated species of arsenic except for
arsenite (27). Plausible
explanations for the differences between our findings and previous
reports are differences in cell lines, arsenite reagent, treatment
durations and stable versus transient overexpression systems.
MRP4 has been proposed to contribute to arsenic
elimination due to its characteristic localization in both the
basolateral membrane of hepatocytes and the apical membrane of
renal proximal tubule cells (21,27).
Based on transport studies using MRP4-enriched membrane vesicles,
the diglutathione conjugate of MMAIII, monomethylarsenic
diglutathione [MMA(GS)2], and DMAV are
transported by MRP4 (27). It is
noteworthy that MRP4 has been reported to regulate intracellular
cyclic adenosine monophosphate (cAMP) levels, an endogenous
substrate identified for MRP4 (21), in AML cell lines and contribute to
cell proliferation and differentiation (34). Moreover, Copsel et al
recently reported that MRP4 blockade strongly reduced tumor growth
by inducing cell cycle arrest and apoptosis in U937 xenografted
mice, and further demonstrated that increased cAMP levels and MRP4
inhibition resulted in leukemic stem cell differentiation (25). Of note, a rapid increase in
intracellular cAMP has been linked to all-trans retinoic
acid (ATRA)-induced differentiation in the human APL cell line NB4
and in fresh APL cells (35). More
importantly, cAMP facilitates the degradation of
AsIII-mediated fusion protein promyelocytic leukemia
(PML)-retinoic acid receptor α (RARα), a fusion gene generated by
the t(15;17) translocation in APL and though to play a central role
in the initiation of leukemogenesis (1,4,36).
These previous findings raised the possibility that MRP4 would be a
novel promising target in APL therapy. The expression of MRP4
increased in CD34+ cells differentiated toward
megakaryocytes with thrombopoietin, and a similar increase was also
observed in a megakaryoblastic cell line (M-07e) derived from a
patient with megakaryoblastic leukemia, when differentiated toward
megakaryocytes (26). However, the
expression of MRP4 decreased in CD34+ cells
differentiated toward monocytes with G-CSF, suggesting a relevant
role of MRP4 in hematopoietic progenitor cell differentiation
(26). Therefore, considering the
expression status of MRPs, including MRP4, is important for
providing meaningful clinical benefits for patients with different
types of hematological disorders.
It is noteworthy that the MRP4 gene is highly
polymorphic, and numerous nonsynonymous single-nucleotide
polymorphisms (SNPs) have been identified (21). Functional studies have shown that
although there was no evidence for a complete loss of function
allele, two variants (G187W and G487E) show a significantly reduced
function compared to reference MRP4 as evidenced by higher
intracellular accumulation of ATZ and PMEA, two antiviral
substrates for MRP4 (19). Although
no disease has so far been directly linked to altered MRP4
activity, evaluating the functional effects of high frequency
variants on the disposition of arsenic has important implications
for hematologic malignancy patients treated with arsenic-based
regimens.
In conclusion, our results demonstrated the capacity
of MRP4 to confer resistance to AsIII as evidenced by
cell survival assays when treated with AsIII in the
presence or absence of its two differential inhibitors. Given that
MRP4 is widely distributed in the body, and plays a pivotal role in
the drug concentrations achieved clinically, monitoring its
expression levels may have important implications for predicting
not only clinical efficacy but also side effects of arsenite and
its metabolites. Obviously, clinical data are required to support
the role of MRP4 in drug disposition and efficacy. Based on our
findings and previous studies showing that MRP4 could be a major
contributor to arsenic resistance, further investigation into the
correlation between the expression level of MRP4 and treatment
outcome of leukemia patients treated with arsenic-based regimens is
warranted.
References
|
1
|
Yuan B, Yoshino Y, Kaise T and Toyoda H:
Application of arsenic trioxide therapy for patients with leukemia.
Biological Chemistry of Arsenic, Antimony and Bismuth. Sun H: John
Wiley and Sons, Ltd; Chichester: pp. 263–292. 2011
|
|
2
|
Cohen MH, Hirschfeld S, Flamm Honig S,
Ibrahim A, Johnson JR, O'Leary JJ, White RM, Williams GA and Pazdur
R: Drug approval summaries: Arsenic trioxide, tamoxifen citrate,
anastrazole, paclitaxel, bexarotene. Oncologist. 6:4–11. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Shen ZX, Chen GQ, Ni JH, Li XS, Xiong SM,
Qiu QY, Zhu J, Tang W, Sun GL, Yang KQ, et al: Use of arsenic
trioxide (As2O3) in the treatment of acute
promyelocytic leukemia (APL): II. Clinical efficacy and
pharmacokinetics in relapsed patients. Blood. 89:3354–3360.
1997.PubMed/NCBI
|
|
4
|
Soignet SL, Maslak P, Wang ZG, Jhanwar S,
Calleja E, Dardashti LJ, Corso D, DeBlasio A, Gabrilove J,
Scheinberg DA, et al: Complete remission after treatment of acute
promyelocytic leukemia with arsenic trioxide. N Engl J Med.
339:1341–1348. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fujisawa S, Ohno R, Shigeno K, Sahara N,
Nakamura S, Naito K, Kobayashi M, Shinjo K, Takeshita A, Suzuki Y,
et al: Pharmacokinetics of arsenic species in Japanese patients
with relapsed or refractory acute promyelocytic leukemia treated
with arsenic trioxide. Cancer Chemother Pharmacol. 59:485–493.
2007. View Article : Google Scholar
|
|
6
|
Kiguchi T, Yoshino Y, Yuan B, Yoshizawa S,
Kitahara T, Akahane D, Gotoh M, Kaise T, Toyoda H and Ohyashiki K:
Speciation of arsenic trioxide penetrates into cerebrospinal fluid
in patients with acute promyelocytic leukemia. Leuk Res.
34:403–405. 2010. View Article : Google Scholar
|
|
7
|
Yoshino Y, Yuan B, Miyashita SI, Iriyama
N, Horikoshi A, Shikino O, Toyoda H and Kaise T: Speciation of
arsenic trioxide metabolites in blood cells and plasma of a patient
with acute promyelocytic leukemia. Anal Bioanal Chem. 393:689–697.
2009. View Article : Google Scholar
|
|
8
|
Iriyama N, Yoshino Y, Yuan B, Horikoshi A,
Hirabayashi Y, Hatta Y, Toyoda H and Takeuchi J: Speciation of
arsenic trioxide metabolites in peripheral blood and bone marrow
from an acute promyelocytic leukemia patient. J Hematol Oncol.
5:12012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen GQ, Shi XG, Tang W, Xiong SM, Zhu J,
Cai X, Han ZG, Ni JH, Shi GY, Jia PM, et al: Use of arsenic
trioxide (As2O3) in the treatment of acute
promyelocytic leukemia (APL): I. As2O3 exerts
dose-dependent dual effects on APL cells. Blood. 89:3345–3353.
1997.PubMed/NCBI
|
|
10
|
Morjani H and Madoulet C:
Immunosuppressors as multidrug resistance reversal agents. Methods
Mol Biol. 596:433–446. 2010. View Article : Google Scholar
|
|
11
|
Lee TC, Ho IC, Lu WJ and Huang JD:
Enhanced expression of multidrug resistance-associated protein 2
and reduced expression of aquaglyceroporin 3 in an
arsenic-resistant human cell line. J Biol Chem. 281:18401–18407.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Leslie EM, Haimeur A and Waalkes MP:
Arsenic transport by the human multidrug resistance protein 1
(MRP1/ABCC1). Evidence that a tri-glutathione conjugate is
required. J Biol Chem. 279:32700–32708. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bhattacharjee H, Carbrey J, Rosen BP and
Mukhopadhyay R: Drug uptake and pharmacological modulation of drug
sensitivity in leukemia by AQP9. Biochem Biophys Res Commun.
322:836–841. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Yoshino Y, Yuan B, Kaise T, Takeichi M,
Tanaka S, Hirano T, Kroetz DL and Toyoda H: Contribution of
aquaporin 9 and multidrug resistance-associated protein 2 to
differential sensitivity to arsenite between primary cultured
chorion and amnion cells prepared from human fetal membranes.
Toxicol Appl Pharmacol. 257:198–208. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Iriyama N, Yuan B, Yoshino Y, Hatta Y,
Horikoshi A, Aizawa S, Takeuchi J and Toyoda H: Aquaporin 9, a
promising predictor for the cytocidal effects of arsenic trioxide
in acute promyelocytic leukemia cell lines and primary blasts.
Oncol Rep. 29:2362–2368. 2013.PubMed/NCBI
|
|
16
|
Yuan B, Ohyama K, Bessho T and Toyoda H:
Contribution of inducible nitric oxide synthase and
cyclooxygenase-2 to apoptosis induction in smooth chorion
trophoblast cells of human fetal membrane tissues. Biochem Biophys
Res Commun. 341:822–827. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Yuan B, Ohyama K, Bessho T, Uchide N and
Toyoda H: Imbalance between ROS production and elimination results
in apoptosis induction in primary smooth chorion trophoblast cells
prepared from human fetal membrane tissues. Life Sci. 82:623–630.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yuan B, Ohyama K, Takeichi M and Toyoda H:
Direct contribution of inducible nitric oxide synthase expression
to apoptosis induction in primary smooth chorion trophoblast cells
of human fetal membrane tissues. Int J Biochem Cell Biol.
41:1062–1069. 2009. View Article : Google Scholar
|
|
19
|
Abla N, Chinn LW, Nakamura T, Liu L, Huang
CC, Johns SJ, Kawamoto M, Stryke D, Taylor TR, Ferrin TE, et al:
The human multidrug resistance protein 4 (MRP4, ABCC4): Functional
analysis of a highly polymorphic gene. J Pharmacol Exp Ther.
325:859–868. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kelly L, Fukushima H, Karchin R, Gow JM,
Chinn LW, Pieper U, Segal MR, Kroetz DL and Sali A: Functional hot
spots in human ATP-binding cassette transporter nucleotide binding
domains. Protein Sci. 19:2110–2121. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Russel FG, Koenderink JB and Masereeuw R:
Multidrug resistance protein 4 (MRP4/ABCC4): A versatile efflux
transporter for drugs and signalling molecules. Trends Pharmacol
Sci. 29:200–207. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Chen ZS, Lee K, Walther S, Raftogianis RB,
Kuwano M, Zeng H and Kruh GD: Analysis of methotrexate and folate
transport by multidrug resistance protein 4 (ABCC4): MRP4 is a
component of the methotrexate efflux system. Cancer Res.
62:3144–3150. 2002.PubMed/NCBI
|
|
23
|
Tian Q, Zhang J, Chan SY, Tan TM, Duan W,
Huang M, Zhu YZ, Chan E, Yu Q, Nie YQ, et al: Topotecan is a
substrate for multidrug resistance associated protein 4. Curr Drug
Metab. 7:105–118. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tian Q, Zhang J, Tan TM, Chan E, Duan W,
Chan SY, Boelsterli UA, Ho PC, Yang H, Bian JS, et al: Human
multidrug resistance associated protein 4 confers resistance to
camptothecins. Pharm Res. 22:1837–1853. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Copsel S, Bruzzone A, May M, Beyrath J,
Wargon V, Cany J, Russel FG, Shayo C and Davio C: Multidrug
resistance protein 4/ATP binding cassette transporter 4: A new
potential therapeutic target for acute myeloid leukemia.
Oncotarget. 5:9308–9321. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Oevermann L, Scheitz J, Starke K, Köck K,
Kiefer T, Dölken G, Niessen J, Greinacher A, Siegmund W, Zygmunt M,
et al: Hematopoietic stem cell differentiation affects expression
and function of MRP4 (ABCC4), a transport protein for signaling
molecules and drugs. Int J Cancer. 124:2303–2311. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Banerjee M, Carew MW, Roggenbeck BA,
Whitlock BD, Naranmandura H, Le XC and Leslie EM: A novel pathway
for arsenic elimination: Human multidrug resistance protein 4
(MRP4/ABCC4) mediates cellular export of dimethylarsinic acid
(DMAV) and the diglutathione conjugate of monomethylarsonous acid
(MMAIII). Mol Pharmacol. 86:168–179. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lee K, Klein-Szanto AJ and Kruh GD:
Analysis of the MRP4 drug resistance profile in transfected NIH3T3
cells. J Natl Cancer Inst. 92:1934–1940. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Lee G and Piquette-Miller M: Influence of
IL-6 on MDR and MRP-mediated multidrug resistance in human hepatoma
cells. Can J Physiol Pharmacol. 79:876–884. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schuetz JD, Connelly MC, Sun D, Paibir SG,
Flynn PM, Srinivas RV, Kumar A and Fridland A: MRP4: A previously
unidentified factor in resistance to nucleoside-based antiviral
drugs. Nat Med. 5:1048–1051. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kikuchi H, Yuan B, Yuhara E, Imai M,
Furutani R, Fukushima S, Hazama S, Hirobe C, Ohyama K, Takagi N, et
al: Involvement of histone H3 phosphorylation via the activation of
p38 MAPK pathway and intracellular redox status in cytotoxicity of
HL-60 cells induced by Vitex agnus-castus fruit extract. Int J
Oncol. 45:843–852. 2014.PubMed/NCBI
|
|
32
|
Carew MW and Leslie EM: Selenium-dependent
and -independent transport of arsenic by the human multidrug
resistance protein 2 (MRP2/ABCC2): Implications for the mutual
detoxification of arsenic and selenium. Carcinogenesis.
31:1450–1455. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kala SV, Neely MW, Kala G, Prater CI,
Atwood DW, Rice JS and Lieberman MW: The MRP2/cMOAT transporter and
arsenic-glutathione complex formation are required for biliary
excretion of arsenic. J Biol Chem. 275:33404–33408. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Copsel S, Garcia C, Diez F, Vermeulem M,
Baldi A, Bianciotti LG, Russel FG, Shayo C and Davio C: Multidrug
resistance protein 4 (MRP4/ABCC4) regulates cAMP cellular levels
and controls human leukemia cell proliferation and differentiation.
J Biol Chem. 286:6979–6988. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Zhao Q, Tao J, Zhu Q, Jia PM, Dou AX, Li
X, Cheng F, Waxman S, Chen GQ, Chen SJ, et al: Rapid induction of
cAMP/PKA pathway during retinoic acid-induced acute promyelocytic
leukemia cell differentiation. Leukemia. 18:285–292. 2004.
View Article : Google Scholar
|
|
36
|
Zhu Q, Zhang JW, Zhu HQ, Shen YL, Flexor
M, Jia PM, Yu Y, Cai X, Waxman S, Lanotte M, et al: Synergic
effects of arsenic trioxide and cAMP during acute promyelocytic
leukemia cell maturation subtends a novel signaling cross-talk.
Blood. 99:1014–1022. 2002.PubMed/NCBI
|