Introduction
Gastric cancer is one of the most common types of
gastrointestinal tumors; its incidence and mortality are among the
highest for all types of malignant tumors. The metastatic
implantation of gastric cancer within the abdominal cavity is the
leading cause of death in patients with gastric cancer, and
effective approaches for its prevention and treatment are
critically needed. The primary process of abdominal implantation of
gastric cancer consists of cancer cell infiltration into the serous
layer of the stomach, detachment into the abdominal cavity, and
formation of an active tumor mass that implants and proliferates to
form metastases (1).
Cell mobility is an important characteristic that
allows tumor cells to invade surrounding tissue and form distant
metastases. Hypoxia can increase tumor cell mobility and migration,
which are closely related to tumor metastasis (2). In a hypoxic microenvironment,
hypoxia-inducible factor-1α (HIF-1α) is highly expressed in tumor
cells (3). HIF-1α is a specific
transcription factor that plays an active role under hypoxic
conditions. High expression of HIF-1α can promote
epithelial-mesenchymal transition (EMT), regulate angiogenesis (via
VEGF), and increase cell adhesion [via integrin β1 (ITGβ1)] to
allow cells to adapt to a hypoxic microenvironment consequently
promoting tumor growth and metastasis (4,5).
As a key regulator of the hypoxia response, HIF-1α
promotes the expression of ITGβ1 to increase the adhesion of
induced cells (5). Adhesion is
important to the process of gastric cancer metastasis. Integrin is
a member of the adhesion factor receptor family. ITGβ1, an
important β subunit of the integrin family, can combine with
different α subunits and form the vast majority of extracellular
matrix receptors. ITGβ1 plays a mediating role in cell-cell and
cell-cell matrix interactions (6).
Enhanced ITGβ1 expression is related to celiac implantation of
cancer (7). Therefore, studies
aimed at examining HIF-1α and ITGβ1 expression are important for
the treatment of tumor recurrence and metastasis.
Animal models have shown that dextran sulphate (DS)
can prevent B-16 melanoma cells from implanting in great omentum
milky spots and the peritoneum and can prolong the survival of mice
with cancerous peritonitis (8). We
hypothesized that DS may inhibit peritoneal metastasis of human
gastric cancer cells through a reduction in HIF-1α and ITGB1
expression. In the present study, gastric cancer cells were exposed
to DS in vitro, and the expression levels of HIF-1α and
ITGβ1 were measured and subjected to correlation analysis.
Furthermore, gastric cancer cells were injected into nude mice to
create a model of gastric carcinoma peritoneal metastasis in
vivo and to observe the influence of DS on the nodules on the
greater omentum. HIF-1α and ITGβ1 expression in the greater omentum
was evaluated for further validation of the in vitro
experiment. Through in vivo and in vitro experiments,
the effects of DS on celiac metastasis of human gastric cancer
cells were evaluated.
Materials and methods
Materials
Cell culture
The human gastric cancer cell line BGC-823 (Beijing
Jin Zijing Biological Pharmaceutical Technology Co., Ltd., Tumor
Cell Library) was cultured in RPMI-1640 medium (Hyclone), fetal
bovine serum (FBS) (Fumeng Gene), trypsin-EDTA (Solarbio), and
streptomycin.
Drugs
DS (Sigma) was dissolved in phosphate-buffered
saline (PBS) for cell culture or in 0.9% normal saline for
injection into nude mice and then sterilized with a 22-µm
filter; the final concentration was 0.3%.
Experimental animals
BALB/c nude mice were purchased from Beijing Tong
Lihua Experimental Animal Technical Co., Ltd. (animal license SCXX,
Beijing, 2006–0009); the animals were 5 to 6 weeks old, male, and
18 to 22 g in weight. All procedures were performed according to
the animal protocol approved by the Ningxia Medical University
Laboratory Animal Centre and internationally recognized guidelines
on animal welfare. Aseptic food and water were available ad
libitum.
Antibodies and reagents
The following antibodies were used: ITGβ1 polyclonal
antibody (Proteintech Group), sheep monoclonal antibody against
ITGβ1, rabbit anti-sheep IgG, goat anti-rabbit IgG and HIF-1α
polyclonal antibody (Proteintech Biological Co., Ltd.) and sheep
anti-rabbit β-actin (Elabscience). The western blot reagents were
as follows: a total protein extraction kit (KeyGen), a protein
quantitative kit (Kang Century), and an enhanced chemiluminescence
kit HRP (Fdbio Science). The PCR reagents were as follows: a total
mRNA extraction kit (Omega), Revert Aid First Strand cDNA Synthesis
kit (Thermo) and PCR kits (Beijing CoWin Biotech Co., Ltd.). The
immunohistochemical reagents used were as follows: goat anti-rabbit
IgG and DAB kit (both from ZSGB-Bio).
Methods for the in vitro experiments
Cell culture
BGC-823 cells were maintained under routine culture
conditions in complete medium in a sterile incubator at a constant
temperature of 37°C with 5% CO2. The medium was replaced
two or three times per week, and the cells were passaged every
three days. The cells were grown to the logarithmic phase in a
60-mm Petri dish. The control and experimental groups received PBS
and DS (final concentration of 0.3%), respectively, and the cells
were placed in a low-oxygen incubator at 37°C with 5%
CO2 and 1% O2. The cells were collected after
2, 8, 12 and 24 h for evaluation.
Immunocytochemical staining
The treated cells were fixed in cold 4%
polyphosphate formaldehyde for 15 min and then washed with PBS. The
fixed cells were permeabilized with 0.5% Triton X-100 for 30 min
and subsequently blocked with goat serum for 30 min at 37°C. After
the cells were washed in PBS, they were incubated with anti-HIF-1α
(1:100) or anti-ITGβ1 (1:100) at 4°C overnight, followed by
incubation with goat anti-rabbit secondary antibodies for 1 h at
room temperature. DAB staining was performed for 3–5 min before
cell evaluation under a microscope. Finally, the cells were
counterstained with haematoxylin. Positive HIF-1α staining was
observed in the nucleus and cytoplasm and was assessed using
Image-Pro Plus 6.0 software (IPP) to obtain the average optical
density (OD) value of the selected field.
Measurement of HIF-1α and ITGβ1 mRNA
levels by RT-PCR
Cells that were treated for different periods were
lysed, and total RNA was extracted and reverse transcribed using a
total mRNA extraction kit and a reverse transcription kit according
to the manufacturer's instructions. The primers for HIF-1α and
ITGβ1 were designed and purchased from Shanghai Sangon Biological
Engineering Co., Ltd., and were as follows: ITGβ1 forward,
5′-CGTAGCAAAGGAACAGCAGA-3′ and reverse,
5′-GTAAGACAGGTCCATAAGGTAGTAG-3′, and HIF-1α forward,
5′-GAAAGCGCAAGTCTTCA AAG-3′ and reverse,
5′-TGGGTAGGAGATGGAGATGC-3′. The RT-PCR assay was analyzed by grey
value analysis using Quantity One software; the target/internal
relative grey-scale value indicates the corresponding mRNA
expression level. The ITGβ1 and HIF-1α mRNA expression levels in
cells were compared between the experimental and control groups at
several time-points.
Western blot detection of HIF-1α and
ITGβ1 protein levels
The cells treated with PBS or DS were harvested,
lysed, subjected to SDS-polyacrylamide gel electrophoresis and
transferred to polyvinylidene fluoride (PVDF) membranes. The
membranes were blocked with 10% skim milk for 1.5 h at room
temperature, and the immunoreactivity levels were evaluated by
hybridization with the following antibodies: rabbit polyclonal
anti-HIF-1α (1:1,000), anti-ITGβ1 (1:500) and anti-β-actin
(1:1,000) antibodies and goat anti-rabbit IgG (1:6,000). The signal
was then detected by chemiluminescence using an ECL kit and
exposure in a darkroom or an Amersham Imager 600 instrument. For
grey value analysis with Quantity One, the target/internal relative
grey scale value indicates the corresponding protein expression
level. HIF-1α and ITGβ1 expression levels in cells were compared
between the experimental and control groups at several
time-points.
Methods for the in vivo experiments
Establishment of a nude mouse model of
gastric celiac metastasis
Ninety BALB/c nude mice were randomly divided into a
control group (n=40) and an experimental group (n=50). Experimental
procedures were performed with approval from the Animal Care
Committee of our institute. The nude mice were anaesthetized with
0.5% pentobarbital sodium and the stomach walls were exposed.
BGC-823 cells (2×106/0.2 ml/mouse) and 0.3% DS (1
ml/mouse) suspended in PBS were injected subcutaneously into the
stomach area of the nude mice in the experimental group. Under the
same conditions, the mice in the control group were injected with
BGC-823 cells suspended in PBS and 0.9% saline. The control group
and experimental group were randomly divided into four groups
each.
Mice were euthanized at different
time-points
Each group of nude mice injected with drugs and
tumor cells was sacrificed by cervical dislocation on day 1, 2, 3,
7 or 14; a natural death group was also established. The number,
size and color of tumor nodules in the abdominal cavity,
particularly on the greater omentum, were observed. Two samples of
implanted tumor nodules were collected from each mouse: one sample
was immediately placed in 10% formalin (pH 7.4) for H&E and
immunohistochemical staining, and the other was cryopreserved at
−80°C in a cryogenic vial, which had been disinfected and treated
with DEPC water, for later evaluation.
H&E and immunohistochemical
staining
Formalin-fixed and paraffin-embedded specimens were
sectioned and fixed in 10% formalin for up to 24 h. The slices were
then deparaffinized, the antigen was retrieved, and the samples
were incubated in hydrogen peroxide for 15 min. Primary antibody
was added to each slice and incubated at 4°C overnight; PBS was
used as a negative control. HRP polymer (HRP secondary antibody)
was added to the washed slides, and the slides were incubated at
37°C for 30 min. Then, 100 µl of DAB working solution was
added drop-wise to each slide, and the slides were incubated for
3–10 min. The slides were then washed, counterstained and
dehydrated before the transparent mounting medium and coverslips
were added. The IPP image automatic analysis system was used at
×400 to select a blank area and five representative areas. The OD
value of each area was determined and calculated, and the average
OD value was obtained for the selected field.
RT-PCR detection of HIF-1α and ITGβ1
mRNA expression in the greater omentum
Total RNA was extracted from tissue using a total
mRNA extraction kit. RNA was then reverse transcribed using a
reverse transcription kit according to the manufacturer's
instructions. The primers used for RT-PCR were as follows: ITGβ1
forward, 5′-CGTAGCAAAGGAACAGCAGA-3′ and reverse,
5′-GTAAGACAGGTCCATAAGGTAGTAG-3′; and HIF-1α forward,
5′-GAAAGCGCAAGTCTTCAAAG-3′ and reverse, 5′-TGGGTAGGAGATGGAGATGC-3′.
This process was followed by grey value analysis with Quantity One;
the target/internal relative grey-scale value indicated the
corresponding mRNA expression level. The ITGβ1 and HIF-1α mRNA
expression levels were compared between the experimental and
control groups at several time points.
Western blot detection of HIF-1α and
ITGβ1 protein expression
Tumor tissues were sheared and added to pyrolysis
liquid. The slurry was centrifuged at 14,000 rpm at 4°C for 15 min.
The supernatant was used to extract the total protein, and the BCA
method was used to quantify the protein levels. The proteins were
subjected to SDS-polyacrylamide gel electrophoresis, transferred to
PVDF membranes, and blocked for 1.5 h. Immunoreactivity levels were
evaluated by hybridization using the following antibodies: rabbit
polyclonal anti-HIF-1α and anti-ITGβ1 antibodies and goat
anti-rabbit IgG. The signal was then detected by chemiluminescence
using an ECL kit and exposure in a darkroom or an Amersham Imager
600 instrument. Grey value analysis was performed with Quantity One
to compare HIF-1α and ITGβ1 expression levels between the
experimental and control groups at different time-points.
Statistical analyses
The data were analyzed using SPSS 17.0 statistical
software. The comparison of two sample means was performed using
the two-sample t-test. Multiple samples were compared using
analysis of variance and correlations between the two factors were
analyzed using Pearson's correlation analysis.
Results
In vitro experimental results
Immunocytochemical staining
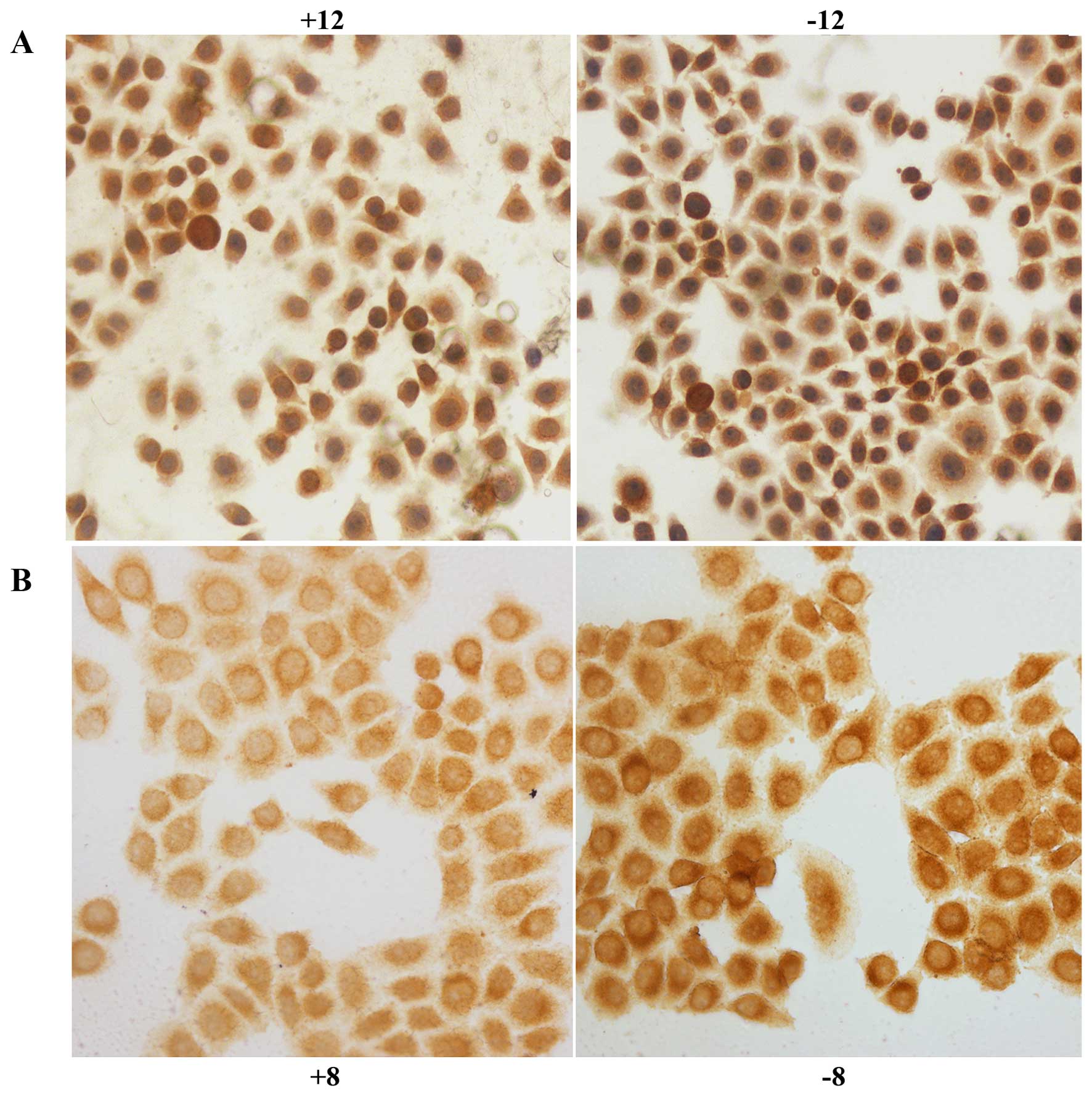
HIF-1α was expressed in the nucleus and cytoplasm of
the control and experimental BGC-823 cell groups; tan color
indicates positive expression. After exposure to hypoxia for 2
(P<0.05), 8 (P<0.01), 12 (P<0.01) or 24 h (P<0.01), the
level of positive expression in the experimental group was
significantly decreased compared with that in the control group
(Fig. 1A). IGTβ1 was expressed in
the cell membrane and the cytoplasm; tan colour indicates positive
expression. After exposure to hypoxia for 2 h, positive expression
in the experimental group was slightly decreased compared with that
in the control group; however, no significant difference was
observed (P>0.05). After exposure to hypoxia for 8 (P<0.01),
12 (P<0.01), or 24 h (P<0.01), the rate of positive
expression was significantly decreased in the experimental group
compared with that noted in the control group (Fig. 1B).
RT-PCR detection of HIF-1α and ITGβ1
mRNA expression in the gastric cancer cells
HIF-1α mRNA expression was significantly lower in
the experimental group than that noted in the control group at each
time point (2, 8, 12, and 24 h). ITGβ1 mRNA expression did not
differ significantly between the two groups at 2 h. However, at 8,
12 and 24 h, the expression levels were significantly lower in the
experimental group than levels in the control group (Fig. 2).
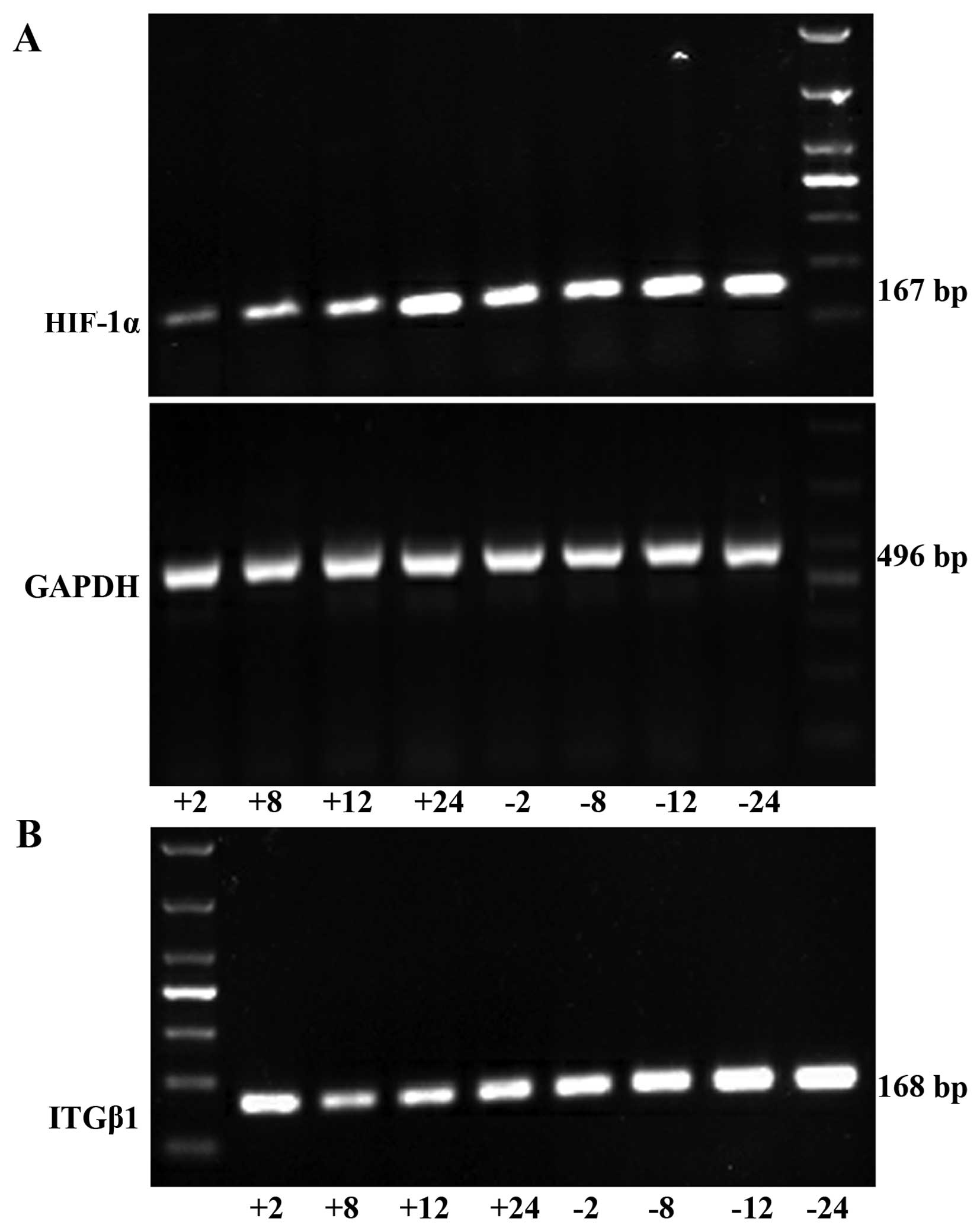 | Figure 2RT-PCR was used to detect HIF-1α (A)
and ITGβ1 (B) mRNA expression levels in the experimental group and
the control group in vitro. (C) Line graph of the gray
values of the relative HIF-1α and ITGβ1 mRNA expression. The labels
+2, +8, +12, +24, indicate the experimental group at 2, 8, 12, and
24 h, respectively, and the labels −2, −8, −12, and −24 indicate
the control group at 2, 8, 12, and 24 h, respectively.
*P<0.05 or **P<0.01, difference between
the experimental group and the control group. |
Western blot detection of HIF-1α and
ITGβ1 protein expression in gastric cancer cells
HIF-1α protein expression was significantly lower in
the experimental group than that noted in the control group at all
time-points (2, 8, 12 and 24 h). ITGβ1 protein expression was
reduced in the experimental group compared with that in the control
group at each time-point; no significant difference between groups
was observed at 2 h, but the differences were significant at 8, 12,
and 24 h (Fig. 3).
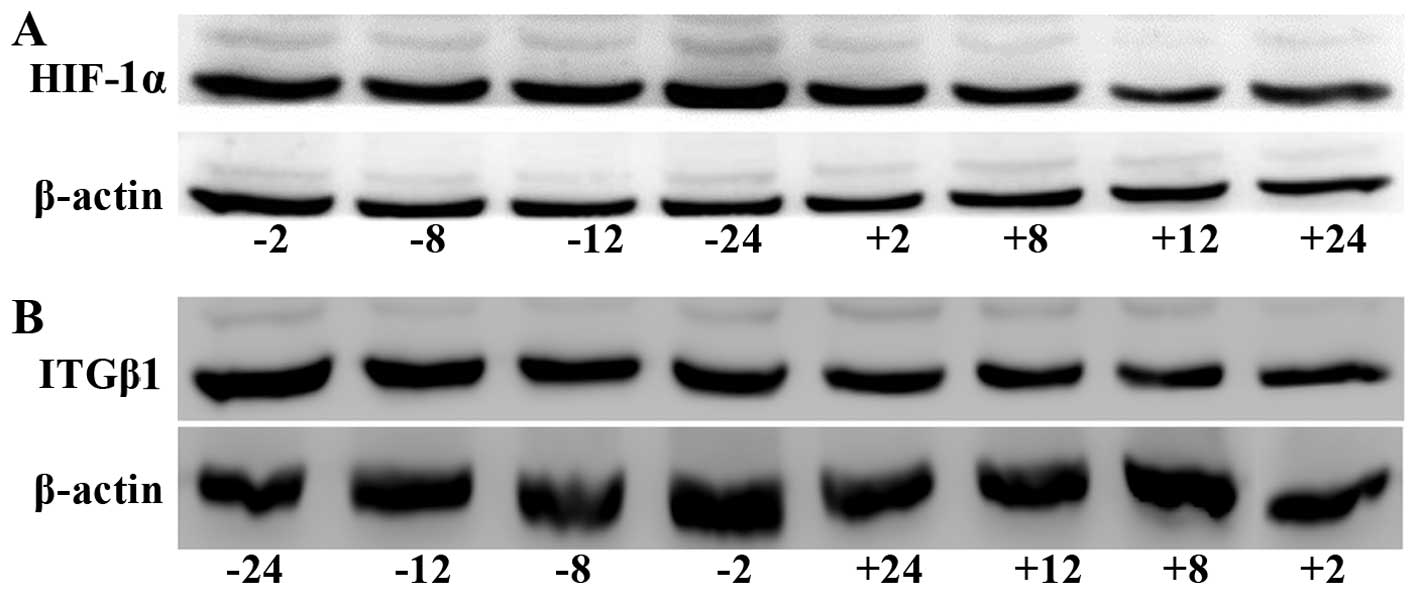 | Figure 3Western blot analysis of HIF-1α (A)
and ITGβ1 (B) protein expression levels in the BGC-823 experimental
and the control cell groups. (C) Line graph of the gray values of
the relative HIF-1α and ITGβ1 protein expression. The labels +2,
+8, +12, +24, indicate the experimental group at 2, 8, 12, and 24
h, respectively, and the labels −2, −8, −12, and −24 indicate the
control group at 2, 8, 12, and 24 h, respectively. *
P<0.05 or **P<0.01, difference between the
experimental group and the control group. |
Correlation analysis of HIF-1α and
ITGβ1 protein expression and time in hypoxia
Pearson's correlation analysis of the HIF-1α and
ITGβ1 protein target/internal grey values and hypoxia time
indicated that ITGβ1 and HIF-1α protein expression levels were
positively correlated with each other and that the level of each
was positively correlated with the time in hypoxia (Table I and Fig. 3).
 | Table ICorrelations of HIF-1α and ITGβ1
protein expression levels and time in hypoxia. |
Table I
Correlations of HIF-1α and ITGβ1
protein expression levels and time in hypoxia.
| | Time in hypoxia | HIF-1α | ITGβ1 |
|---|
| Hypoxia time | Pearson's
correlation | 1 | 0.789a | 0.753a |
| Significance
(two-sided) | | 0.020 | 0.031 |
| N | 8 | 8 | 8 |
| HIF-1α | Pearson's
correlation | 0.789a | 1 | 0.906b |
| Significance
(two-sided) | 0.020 | | 0.002 |
| N | 8 | 8 | 8 |
| ITGβ1 | Pearson's
correlation | 0.753a | 0.906b | 1 |
| Significance
(two-sided) | 0.031 | 0.002 | |
| N | 8 | 8 | 8 |
In vivo results
The number of tumor nodules in the
abdominal cavities of mice in the experimental and control
groups
After the injection of drugs and cells, the numbers
of tumor nodules on the greater omentum, abdominal wall and
mesentery gradually increased over time in the experimental and
control groups. The nodules appeared on the greater omentum first
and were the most numerous. The tumor nodules on the greater
omentum were unequal in size and pale with a slightly hard texture
and a smooth surface. Some nodules fused and remained independent
but lacked well-defined borders and well-defined notes for
independent nodules. On the third day, the tumor nodules in the
experimental group and control group showed no obvious differences
(P>0.05). At 7 days (P=0.003) and 14 days (P=0.000), the nodule
number and diameter in the experimental group were significantly
reduced compared with those of the control group (Fig. 4).
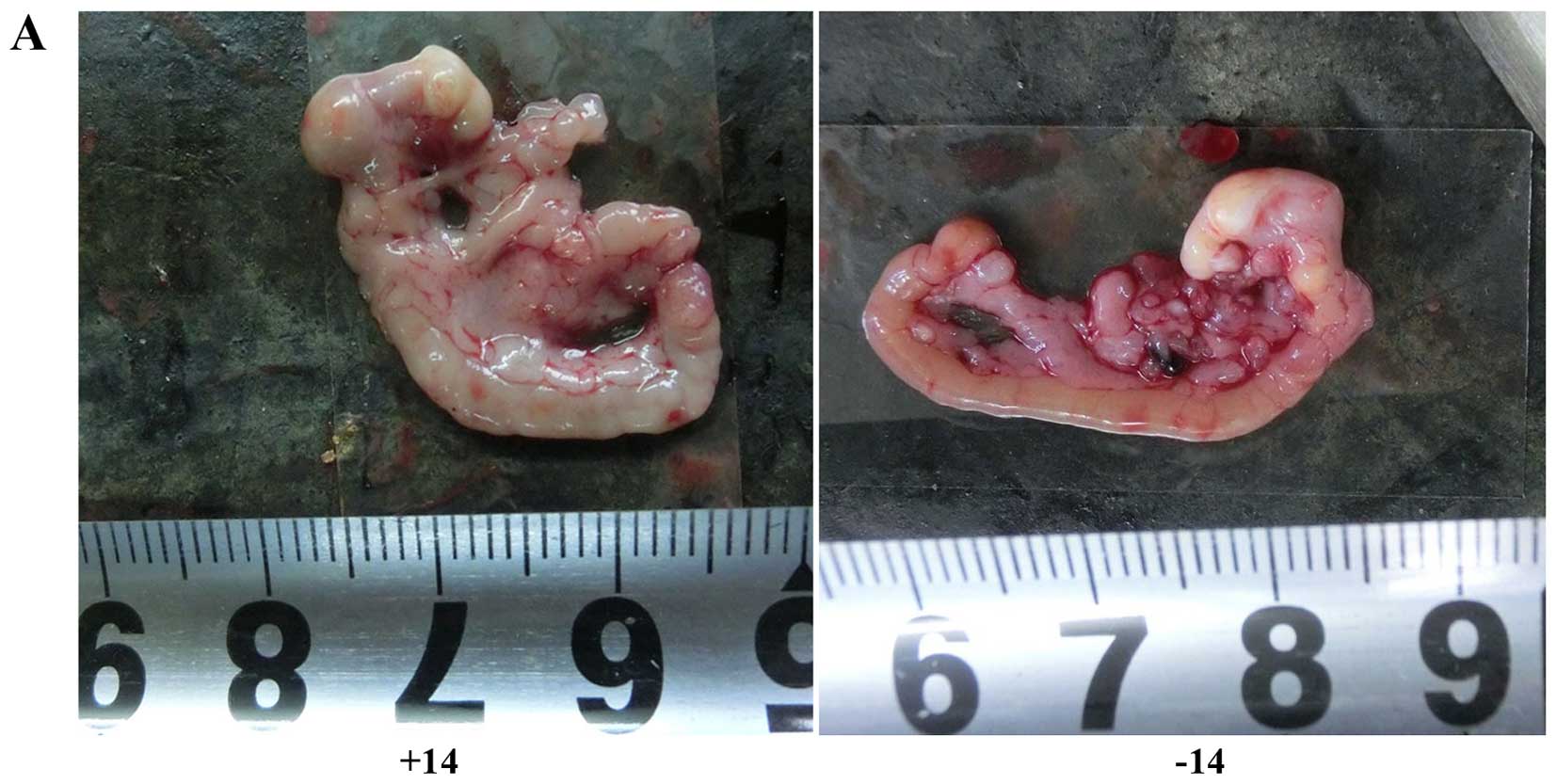 | Figure 4Comparison of abdominal tumor nodules
between the experimental and control groups. (A) Images of the
greater omentum in mice on day 14. (B) Histogram of the numbers of
nodules on the greater omentum. The labels +1, +3, +7, and +14
indicate day 1, 3, 7 and 14 of the experimental group,
respectively, and the labels −1, −3, −7, and −14 indicate day 1, 3,
7 and 14 of the control group, respectively. The numbers of tumor
nodules at day 7 and 14 were significantly reduced in the
experimental group compared with the controls.
**P<0.01, difference between the experimental group
and the control group. |
Immunohistochemical detection of the
expression of HIF-1α and ITGβ1 in the experimental and control
groups at different times
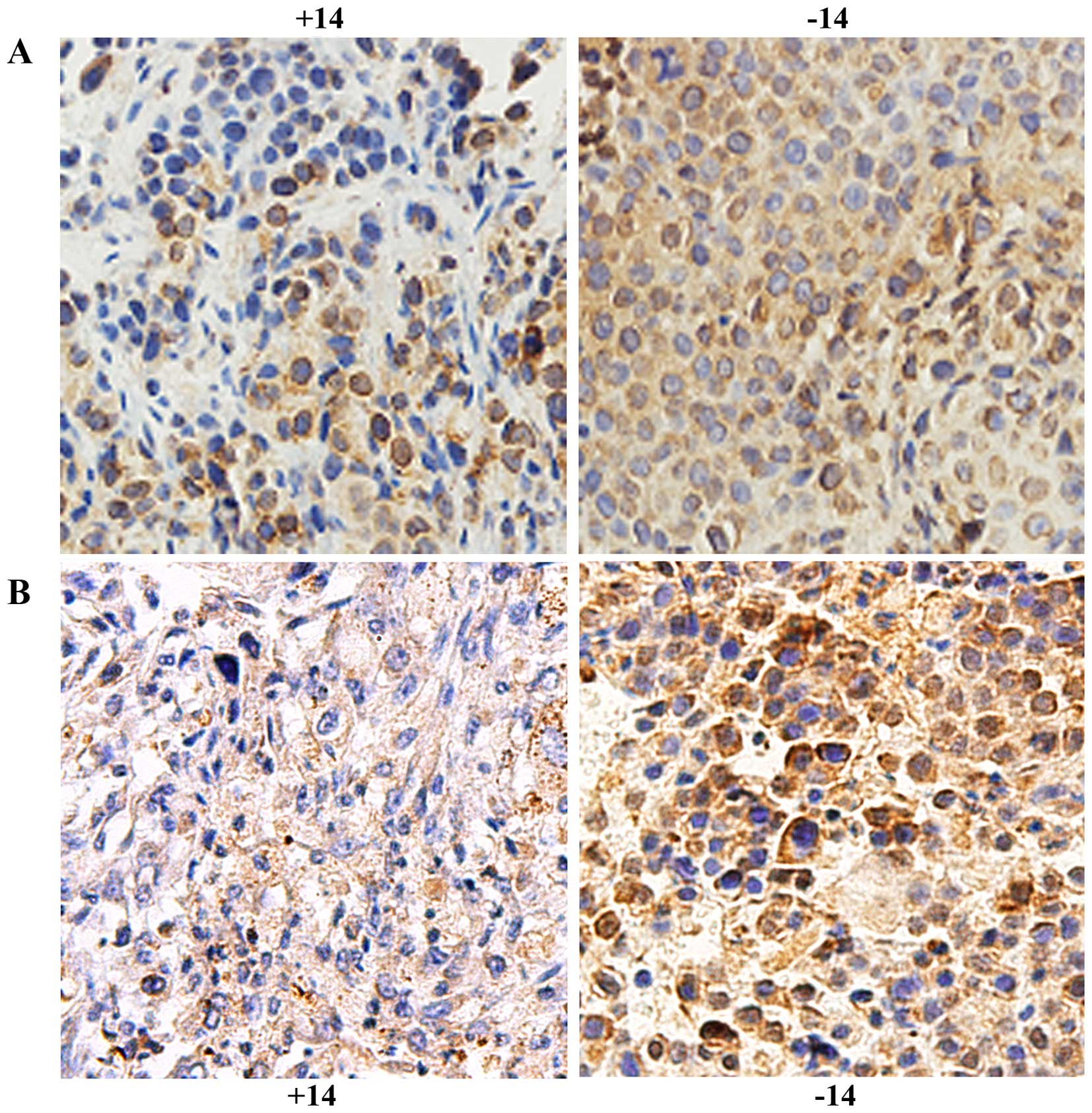
HIF-1α was expressed in the nucleus and cytoplasm.
On day 1, no obvious difference was found between the experimental
group and the control group (P>0.05). On days 3, 7 and 14,
expression was significantly reduced in the experimental group
compared with that noted in the control group (P<0.01, Fig. 5A and Table II). ITGβ1 was expressed in the
cytoplasm, and its expression was significantly lower in the
experimental group than that in the control group (P<0.05,
Fig. 5B and Table III).
| Table IIThe average optical density values of
the immunohistochemical staining for HIF-1α expression in the
control and experimental groups at different time-points. |
Table II
The average optical density values of
the immunohistochemical staining for HIF-1α expression in the
control and experimental groups at different time-points.
| Group | Day 1 | Day 3 | Day 7 | Day 14 | F | P-value |
|---|
| Control | 57.87±4.36 | 75.27±3.36 | 95.41±4.81 | 138.0±4.30 | 328.88 | <0.001 |
| Experimental | 54.39±2.18 | 68.04±3.25 | 83.54±3.77 | 88.21±4.51 | 96.195 | <0.001 |
| t | 1.577 | 3.456 | 4.338 | 17.881 | | |
| P-value | 0.154 | 0.009 | 0.002 | <0.001 | | |
| Table IIIImmunohistochemical staining for ITGβ1
protein expression at different time-points in the experimental and
control groups. |
Table III
Immunohistochemical staining for ITGβ1
protein expression at different time-points in the experimental and
control groups.
| Group | Day 1st | Day 3 | Day 7 | Day 14 | Natural death
group | F | P-value |
|---|
| Control | 115.63±14.38 | 109.71±14.95 | 152.86±16.74 | 220.58±16.24 | 185.68±16.63 | 96.89 | <0.001 |
| Experimental | 53.33±4.93 | 53.10±9.26 | 8.12±1.25 | 176.77±13.31 | 129.10±14.10 | | |
| t | −4.10 | −3.22 | −8.62 | −2.09 | −2.60 | | |
| P-value | 0.01 | 0.01 | <0.001 | 0.05 | 0.02 | | |
RT-PCR detection of HIF-1α and ITGβ1
expression on the greater omentum
Analysis of the grey values with image analysis
software showed that the expression levels of HIF-1α and ITGβ1 in
the experimental group were significantly lower than those in the
control group at each time point (Fig.
6 and Tables IV and V).
 | Figure 6RT-PCR was used to detect the HIF-1α
(A) and ITGβ1 (B) mRNA expression in the experimental and control
group in vivo. The labels +1, +2, +3, +7, +14, and +0
indicate the experimental group at days 1, 2, 3, 7, and 14 and the
natural death group, respectively. The labels −1, −2, −3, −7, −14,
and −0 indicate the control group at days 1, 2, 3, 7, and 14 and
the natural death group, respectively. |
| Table IVHIF-1α mRNA expression in the
experimental and control groups at different times. |
Table IV
HIF-1α mRNA expression in the
experimental and control groups at different times.
| Group | Day 1 | Day 3 | Day 7 | Day 14 | F | P-value |
|---|
| Control group | 1.26±0.47 | 1.7±0.34 | 2.71±0.44 | 3.5±0.41 | 100.944 | <0.001 |
| Experimental
group | 0.91±0.18 | 1.4±0.22 | 1.94±0.47 | 2.8±0.46 | 27.963 | <0.001 |
| t | 2.752 | 2.664 | 4.705 | 10.968 | | |
| P-value | 0.016 | 0.015 | 0.001 | <0.001 | | |
| Table VITGβ1 mRNA expression in the
experimental and control groups at different times. |
Table V
ITGβ1 mRNA expression in the
experimental and control groups at different times.
| Group (day) | Day 1 | Day 2 | Day 3 | Day 7 | Day 14 | Natural death | F | P-value |
|---|
| Control group | 1.28±0.08 | 1.3±0.05 | 1.44±0.09 | 2.5±0.02 | 2.24±0.11 | 2.59±0.11 | 190.67 | 0.001 |
| Experimental
group | 0.75±0.03 | 0.7±0.03 | 0.97±0.03 | 1.3±0.04 | 1.50±0.05 | 1.83±0.03 | | |
| t | −6.43 | −7.90 | −5.33 | −26.12 | −4.66 | −18.98 | | |
| P-value | 0.003 | 0.001 | 0.006 | <0.001 | 0.01 | <0.001 | | |
Western blot detection of expression
of HIF-1α and ITGβ1 in tumor nodules
Analysis of the grey values with image analysis
software showed that the expression levels of HIF-1α and ITGβ1 in
the experimental group were significantly lower than those in the
control group at days 3, 7 and 14 (Fig.
7).
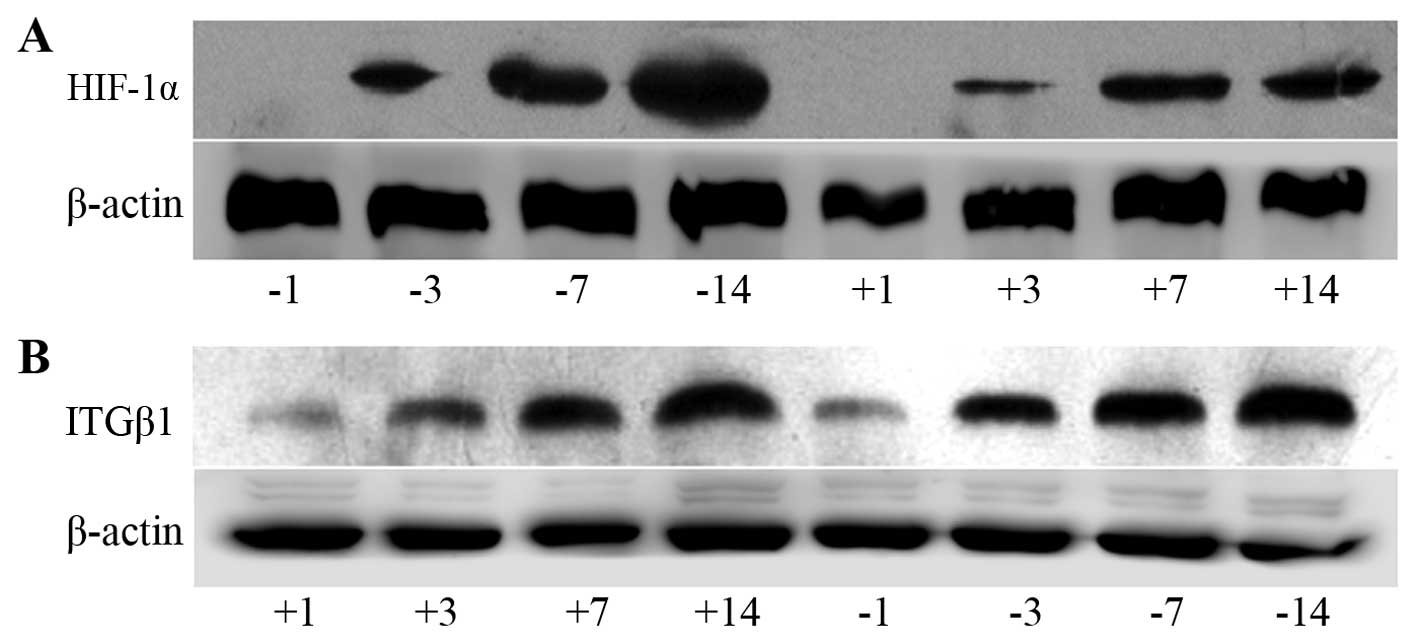 | Figure 7Electrophoresis banding of HIF-1α (A)
and ITGβ1 (B) protein in the experimental and control groups. (C)
Line graph of the gray values of the relative HIF-1α and ITGβ1
protein expression. The labels +1, +3, +7, and +14 indicate the
experimental group at days 1, 3, 7 and 14, respectively, and the
labels −1, −3, −7, and −14 indicate the control group at days 1, 3,
7 and 14, respectively. At days 3, 7 and 14, HIF-1α and ITGβ1
expression levels were significantly lower in the experimental
group than levels noted in the control group. *P<0.05
or **P<0.01, difference between the experimental
group and the control group. |
Correlation analysis of tumor nodule
number, HIF-1α and ITGβ1 protein expression levels and time since
injection
In the experimental and control groups, tumor nodule
number on the greater omentum, HIF-1α and ITGβ1 target/internal
grey values, and drug and cell injection time were subjected to
Pearson's correlation analysis. Nodule number and HIF-1α and ITGβ1
protein expression levels were all positively correlated with the
time since injection. HIF-1α and ITGβ1 protein expression levels
were positively correlated with tumor nodule number, and HIF-1α and
ITGβ1 protein expression levels were positively correlated with
each other (Table VI and Fig. 7).
| Table VICorrelation of HIF-1α and ITGβ1
protein levels and drug and cell injection times in
vivo. |
Table VI
Correlation of HIF-1α and ITGβ1
protein levels and drug and cell injection times in
vivo.
| | Injection time | Tumor nodule
number | HIF-1α protein
level | ITGβ1 protein
level |
|---|
| Injection time | Pearson's
correlation | 1 | 0.908a | 0.847a | 0.932a |
| Significance
(two-sided) | | 0.002 | 0.008 | 0.001 |
| N | 8 | 8 | 8 | 8 |
| Tumor nodule
number | Pearson's
correlation | 0.908a | 1 | 0.991a | 0.941a |
| Significance
(two-sided) | 0.002 | | 0.000 | 0.000 |
| N | 8 | 8 | 8 | 8 |
| HIF-1α protein | Pearson's
correlation | 0.847a | 0.991a | 1 | 0.909a |
| Significance
(two-sided) | 0.008 | 0.000 | | 0.002 |
| N | 8 | 8 | 8 | 8 |
| ITGβ1 protein | Pearson's
correlation | 0.932a | 0.941a | 0.909a | 1 |
| Significance
(two-sided) | 0.001 | 0.000 | 0.002 | |
| N | 8 | 8 | 8 | 8 |
Discussion
Gastric cancer is one of the most common
gastrointestinal malignant tumors in China. The morbidity and
mortality of gastric cancer are among the highest for all types of
malignant tumors, and recurrence and metastasis are the primary
causes of the high mortality rate of gastric cancer. Currently,
more than half of patients with gastric cancer eventually die of
peritoneal implantation. Therefore, methods for preventing gastric
cancer and treating it after its spread to the abdominal cavity are
urgently needed. The primary process of abdominal implantation of
gastric cancer consists of cancer cell infiltration into the serous
layer of the stomach, detachment into the abdominal cavity, and
formation of an active tumor mass that implants and proliferates to
form metastases (1).
After gastric cancer cells disperse into the
abdominal cavity, they are likely to grow in the intra-abdominal
viscera and greater omentum. Surgical treatment often fails to
eradicate these cells; therefore, patient survival rates remain
low. Postoperative chemotherapy could destroy micrometastases and
residual tumors, and it is considered an effective treatment for
postoperative gastric cancer metastasis. However, due to the
existence of the peritoneal serous membrane barrier, most of the
intravenous chemotherapeutic drugs fail to reach the abdominal
cavity, and drugs directly injected into the abdominal cavity more
easily achieve effective concentrations. Common chemotherapeutic
drugs have a small molecular weight and are absorbed easily in the
abdominal cavity; therefore, their efficacy is reduced. The present
study used DS, a type of macromolecular dextran derivative with a
molecular weight of 500,000; this substance is absorbed slowly in
the abdominal cavity and therefore has a high level of safety and
few side effects. Guided by results that indicate that DS
suppresses cell adhesion and cell cycle progression, both of which
are essential for metastasis, DS could be used as an
anti-metastatic drug (9). The
present study used DS to prevent the growth of gastric cancer cells
in a nude mouse celiac implantation model and administered the same
amount of PBS or normal saline to a control group to study the
possible mechanism by which DS inhibits the peritoneal metastasis
of gastric cancer cells.
During tumor invasion of the surrounding tissue,
which occurs in the process of metastasis, active cell mobility is
an important factor. A lack of oxygen can improve tumor cell
mobility and migration and is closely related to tumor metastasis
(2). The oxygen level is a basic
characteristic of the solid tumor microenvironment and can affect
multiple stages of tumor development. When tumor growth reaches a
particular stage, such that oxygen demand exceeds the supply or
immature blood vessels within the tumor stroma rise and collapse,
the local microenvironment enters a hypoxic condition. HIF-1 is
primarily composed of HIF-1α and HIF-1β subunits. HIF-1α is the
only oxygen-sensitive subunit and is primarily expressed in hypoxic
cells (10). In a hypoxic
microenvironment, cancer cells highly express HIF-1α (3), which is the primary factor in the
hypoxic state, and plays an active specific role in the hypoxic
response. HIF-1α is widely found in mammals and humans and can
connect upstream and downstream signals in the process of oxygen
activation. HIF-1α allows cells to adapt to a hypoxic
microenvironment and stimulates tumor growth and metastasis by
promoting cell EMT, the regulation of angiogenesis (via VEGF) and
increased cell adhesion (via ITGβ1) (4,5).
Therefore, ITGβ1 is important in the prevention and treatment of
tumor recurrence along with HIF-1α.
Here, we exposed gastric cancer BGC-823 cells to
hypoxia to stimulate HIF-1α expression and to observe whether
HIF-1α expression was affected by DS. We found that HIF-1α mRNA and
protein expression levels in the experimental and control groups
were positively correlated with time in hypoxia and that DS reduced
the expression of HIF-1α in a hypoxic environment. DS prevented
cell adaptation to a hypoxic environment and reduced tumor growth
and metastasis by inhibiting the expression of HIF-1α. This finding
requires further experimental and clinical validation.
In addition, the increased expression of HIF-1α can
increase cell adhesion (through ITGβ1) as cells adapt to a hypoxic
microenvironment, promoting tumor growth and metastasis. This
process can occur through the transcriptional regulatory mechanism
to promote ITGβ1 expression and function. Subsequent studies
identified functional hypoxia-inducible activity, mediated by HIF,
in the distal 5′-region of the ITGB1 promoter (11). Adherence plays an important role in
the process of gastric cancer metastasis. Integrins are a family of
adhesion factors, and ITGβ1 is an important β subunit of the
integrin family. ITGβ1 can combine with different α subunits to
constitute important receptors in the extracellular matrix.
Integrin plays a primary role in the occurrence and development of
tumors: i) by mediating the adhesion of tumor cells and
extracellular matrix, thereby promoting tumor cell invasion and
metastasis. ii) The differentiation. Abnormal integrin signaling
promotes tumor cell growth and stimulates differentiation and
distant metastasis (12). Research
has shown that when tumor cells enter the circulation in a free
state, the enhanced expression of ITGβ1 facilitates cancer cell
adhesion, implantation and transfer (12,13).
Research has shown that dysregulated ITGβ1
expression is related to cancer celiac implantation (7) and plays a key role in gastric cancer
cell adherence to the peritoneum. The hTERT/MDM2-FOXO3a-ITGB1
pathway markedly contributes to gastric cancer invasion (14), suggesting that ITGβ1 may be a novel
target for the prevention and treatment of gastric cancer
metastasis. Other studies have suggested that an intra-peritoneal
injection of the ITGβ1 monoclonal antibody can significantly reduce
adhesion, inhibit cancer cell peritoneal implants, and reduce the
number of peritoneal implanting nodules (15). We observed ITGβ1 expression in
gastric cancer cells and found that DS reduced ITGβ1 expression and
inhibited gastric cancer cell adhesion, thus inhibiting gastric
cancer metastasis.
The above in vitro experimental results
indicated that DS can inhibit HIF-1α and ITGβ1 expression in
gastric cancer cells and cell adhesion. Because of the close
relationship of HIF-1α and ITGβ1 and their important roles in tumor
cell metastasis, we analyzed the correlation between the two
proteins. Both HIF-1α and ITGβ1 expression were strongly related to
intervention time, and HIF-1α expression was positively correlated
with ITGβ1 expression in both the experimental group and the
control group. We hypothesized that HIF-1α promotes tumor cell
adaptation to a hypoxic microenvironment, thereby promoting tumor
cell survival, growth and metastasis, by regulating the downstream
factor ITGβ1.
Animal experiments have shown that DS can prevent
B-16 melanoma cell growth in the greater omentum and peritoneal
planting and can prolong the survival of mice with carcinoma of
peritonitis (8). To further verify
the impact of DS on gastric cancer celiac implantation, we used a
series of nude mice that were administered a celiac injection of
gastric cancer cells. The results showed that the number of tumor
nodules markedly increased with the extension of the time since
cancer cell injection. Nodules appear in the greater omentum first
and most frequently, but tumor nodules can also be found in the
liver, spleen, stomach wall and mesentery. However, this study
focused on the greater omentum tumor nodules. At each time point,
the tumor nodules on the greater omentum in the experimental group
were significantly reduced in number and volume compared with those
of the control group. The tumor nodule number was significantly and
positively correlated with the time since injection.
Immunohistochemistry, RT-PCR and western blot test results showed
that the mRNA and protein expression levels of HIF-1α and ITGβ1 in
the experimental group were significantly lower than those of the
control group. In addition, ITGβ1 expression was positively
correlated with HIF-1α expression, and the expression of both ITGβ1
and HIF-1α was positively correlated with injection time and tumor
nodule number. We hypothesized that DS likely acts through the
inhibition of HIF-1α expression, which reduced ITGβ1 expression and
inhibited cancer cell adhesion. Thus, the local tumor hypoxia state
cannot be improved in the presence of DS and the tumor growth is
restrained, eventually reducing celiac implantation and gastric
cancer metastasis.
In summary, the above results demonstrate that the
expression of ITGβ1 is related to HIF-1α expression and that DS can
reduce the expression of HIF-1α and ITGβ1 in gastric cancer cells
and the greater omentum, effectively restraining the growth of
nodules in the omenta of nude mice and, to some extent, may inhibit
the celiac implantation and metastasis of gastric cancer cells. The
mechanism may involve the inhibition of HIF-1α, which reduces ITGβ1
expression and inhibits cancer cell adhesion; thus, the local tumor
hypoxic condition persists, thereby reducing tumor metastasis. This
study may help the development of new effective drugs for the
prevention and treatment of gastric cancer celiac metastasis.
Acknowledgments
An English Language Service from American Journal
Experts was used to help prepare the manuscript. This study was
supported by the Ningxia Natural Science Foundation (NZ1085),
National Natural Science Foundation of China (no. 81460370) and
Ningxia Natural Science Foundation (no. NZ13143), Ningxia Science
and Technology Support Projects (2002310201).
References
|
1
|
Yamaguchi H, Kitayama J, Emoto S, Ishigami
H, Ito T, Hanafusa N and Watanabe T: Cell-free and concentrated
ascites reinfusion therapy (CART) for management of massive
malignant ascites in gastric cancer patients with peritoneal
metastasis treated with intravenous and intraperitoneal paclitaxel
with oral S-1. Eur J Surg Oncol. 41:875–880. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jing SW, Wang YD, Kuroda M, Su JW, Sun GG,
Liu Q, Cheng YJ and Yang CR: HIF-1α contributes to hypoxia-induced
invasion and metastasis of esophageal carcinoma via inhibiting
E-cadherin and promoting MMP-2 expression. Acta Med Okayama.
66:399–407. 2012.
|
|
3
|
Li H, Chen J, Zen W, Xu X, Xu Y, Chen Q
and Yang T: Effect of hypoxia inducible factor-1 antisense
oligonucleotide on liver cancer. Int J Clin Exp Med. 8:12650–12655.
2015.PubMed/NCBI
|
|
4
|
Bao B, Wang Z, Ali S, Kong D, Li Y, Ahmad
A, Banerjee S, Azmi AS, Miele L and Sarkar FH: Notch-1 induces
epithelial-mesenchymal transition consistent with cancer stem cell
phenotype in pancreatic cancer cells. Cancer Lett. 307:26–36. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lee SH, Lee YJ and Han HJ: Role of
hypoxia-induced fibronectin-integrin β1 expression in embryonic
stem cell proliferation and migration: Involvement of PI3K/Akt and
FAK. J Cell Physiol. 226:484–493. 2011. View Article : Google Scholar
|
|
6
|
Goggins BJ, Chaney C, Radford-Smith GL,
Horvat JC and Keely S: Hypoxia and integrin-mediated epithelial
restitution during mucosal inflammation. Front Immunol. 4:2722013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Niu G and Chen X: Why integrin as a
primary target for imaging and therapy. Theranostics. 1:30–47.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hagiwara A, Sakakura C, Yamasaki J, Togawa
T, Sonoyama Y, Fujiyama J and Yamagishi H: Dextran sulfate inhibits
injured abdominal wall-specific tumor implantation in mice.
Anticancer Drugs. 11:873–877. 2000. View Article : Google Scholar
|
|
9
|
Takagi T, Sakakura C, Kin S, Nakase Y,
Fukuda K, Shimomura K, Ito T, Fujiyama J, Yamasaki J, Tsujimoto H,
et al: Dextran sulfate suppresses cell adhesion and cell cycle
progression of melanoma cells. Anticancer Res. 25:895–902.
2005.PubMed/NCBI
|
|
10
|
Kitajima Y and Miyazaki K: The critical
impact of HIF-1a on gastric cancer biology. Cancers (Basel).
5:15–26. 2013. View Article : Google Scholar
|
|
11
|
Keely S, Glover LE, MacManus CF, Campbell
EL, Scully MM, Furuta GT and Colgan SP: Selective induction of
integrin beta1 by hypoxia-inducible factor: Implications for wound
healing. FASEB J. 23:1338–1346. 2009. View Article : Google Scholar :
|
|
12
|
Yeh YC, Lin HH and Tang MJ: A tale of two
collagen receptors, integrin β1 and discoidin domain receptor 1, in
epithelial cell differentiation. Am J Physiol Cell Physiol.
303:C1207–C1217. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang L and Zou W: Inhibition of integrin
β1 decreases the malignancy of ovarian cancer cells and potentiates
anticancer therapy via the FAK/STAT1 signaling pathway. Mol Med
Rep. 12:7869–7876. 2015.PubMed/NCBI
|
|
14
|
Hu C, Ni Z, Li BS, Yong X, Yang X, Zhang
JW, Zhang D, Qin Y, Jie MM, Dong H, et al: hTERT promotes the
invasion of gastric cancer cells by enhancing FOXO3a ubiquitination
and subsequent ITGB1 upregulation. Gut. Sep 14–2015.Epub ahead of
print. View Article : Google Scholar
|
|
15
|
Lynch L, Vodyanik PI, Boettiger D and
Guvakova MA: Insulin-like growth factor I controls adhesion
strength mediated by alpha5beta1 integrins in motile carcinoma
cells. Mol Biol Cell. 16:51–63. 2005. View Article : Google Scholar :
|