Introduction
Lung cancer (LC) is currently the leading cause of
cancer-related mortality among both males and females worldwide and
was responsible for approximately 1.59 million deaths in 2012
(1,2). LC has also been the number one cause
of cancer-related deaths among patients with malignant tumors since
2008 in China (3). In addition, the
mortality rate of LC in China has increased dramatically during the
past three decades due to smoking and air pollution, imposing a
huge economic burden on patients, medical professionals, and
society (3).
LC is characterized by a series of hallmarks, such
as tumor-promoting inflammation, avoidance of immune destruction,
genomic instability, and induction of angiogenesis (4). Inflammatory responses contribute to
the initiation, progression, and metastases of malignancies as
proposed by Virchow in 1863, by promoting proliferative signaling,
destabilizing genomic integrity, and inducing the invasion of
cancer cells (5,6). Chronic inflammation triggered by
bacterial and viral infections, tobacco smoking, and chemicals
suppress wound healing and tissue regenerative responses, promoting
cancer development and progression, as described as 'wounds that do
not heal' (7,8). In addition, inflammation compromises
genomic maintenance and repair pathways, inducing genomic
instability (6,9). Furthermore, cancer cells modulate
inflammation by secreting soluble mediators and interfering with
innate and adaptive immune cells, such as, macrophages, dendritic
cells and lymphocytes (6). Multiple
inflammatory mediators may trigger and maintain tumorigenesis
individually or coordinately in the tumor microenvironment
(6). The crosstalk between the
inflammatory microenvironment and cancer cells controls and shapes
tumor growth and metastasis (5–7).
Interleukin (IL)-17A, a proinflammatory cytokine
discovered in 1993, induces tissue inflammation mainly by promoting
expression of various cytokines, chemokines, antimicrobial
peptides, and tissue-remodeling molecules (10–12).
IL-17A exerts complicated functions in allergic, autoimmune, and
malignant diseases, by targeting mesenchymal and myeloid cells
(10,13). Although originally linked to
IL-17-producing CD4+ T helper (Th17) cells, a distinct T
cell subtype different from Th1 and Th2 cells, IL-17A was
subsequently found to be produced by several other immune cells,
including IL-17A-producing γδT (γδT17) cells, IL-17A-producing
CD8+ T (Tc17) cells, natural killer T cells, and mast
cells (10,14,15).
IL-17A-producing T cells and associated cytokines, such as IL-17,
IL-23, and IL-1β, have been shown to be involved in both
inflammation and immune responses in various types of cancers
including gastric, breast, prostate and hepatocellular cancer
(10,16–19).
However, the roles of these T cells and associated cytokines are
conflicting in various animal models and patients (10,20–22).
IL-17-producing T cells have displayed both antitumor and protumor
functions, due to their plasticity and functions in the tumor
microenvironments (23). The
numbers of circulating Th17 and γδT17 cells were significantly
higher in patients with gastric cancer than those in healthy
controls (HCs) (16). On the
contrary, data have shown that Th17 cells elicit antitumor effects,
by promoting cytotoxic activities, enhancing Th1 response, and
augmenting the expression of MHC antigens (24–26).
In a murine model of LC, enhanced Th17 cells and overexpression of
IL-17A stimulated tumor growth in the lungs (27,28).
Similarly, an increased number of intratumoral IL-17-positive cells
in patients with LC was correlated with poor prognosis (29). However, a higher percentage of Tregs
but a lower frequency of Th17 cells was found in malignant pleural
effusion, as compared with those in parapneumonic effusion, and a
higher ratio of Treg/Th17 cells in malignant pleural effusion was
found to indicate a poor prognosis of patients with LC (30). Thus, the roles of IL-17A-producing T
cells, especially Tc17 and γδT17 cells, and associated cytokines in
the progression of LC remain to be defined.
In the present study, we investigated the
frequencies of IL-17-producing T cells (Th17, Tc17 and γδT17 cells)
and levels of IL-17A-associated cytokines (IL-17A, IL-23, IL-1β,
and TGF-β1) in patients with lung adenocarcinoma (LA) and HCs. We
found that the frequencies of both Th17 and γδT17 cells in the
peripheral blood of patients with LA were higher than those in HCs
and were positively associated with tumor invasion and metastasis,
whereas the frequency of Tc17 cells in patients with LA was
decreased. Furthermore, the major source of IL-17A production was
Th17 cells in peripheral blood from both patients with LA and HCs.
In addition, the protein and corresponding mRNA levels of IL-17A,
IL-23, IL-1β, and TGF-β1 were much higher in patients with LA than
those in HCs, and the levels of IL-17A were positively correlated
with numbers of both Th17 and γδT17 cells, but not Tc17 cells.
Finally, the frequencies of circulating Th17 and γδT17 cells, along
with levels of IL-17A, IL-23, IL-1β, and TGF-β1 were decreased in
the patients with LA after tumor resection, whereas the frequency
of circulating Tc17 cells was inversely increased in these
patients. This study provides further insight into the association
between IL-17A-producing T cells and progression of LA, and offers
promising targets for therapeutic strategies.
Materials and methods
Subjects
Forty patients, diagnosed with LA and admitted to
the Department of Respiratory Medicine at the First Affiliated
Hospital of Zhejiang University, and 35 HCs were enrolled in this
study from May 2014 to June 2015. All patients were histologically
confirmed with LA by two pathologists. The patients were excluded
if they had autoimmune diseases, immune compromised diseases, and
pulmonary infections, if they were taking any drugs which affect
immune responses, or if they had already received anticancer
therapies including chemotherapy, radiotherapy, targeted therapy,
surgery or immune therapy. The characteristics of the enrolled 40
patients and 35 HCs, are summarized in Table I. All cases with LA were staged
according to the 7th edition of the tumor, node, and metastasis
(TNM) classification for LC (31).
The study was approved by the Ethics Committee of the First
Affiliated Hospital of Zhejiang University, and informed consent
was obtained from all patients and HCs.
 | Table ICharacteristics of the patients with
lung adenocarcinoma and healthy controls. |
Table I
Characteristics of the patients with
lung adenocarcinoma and healthy controls.
|
Characteristics | Patients | Healthy
controls |
|---|
| Gender |
| Male | 27 | 24 |
| Female | 13 | 11 |
| Age (years) |
| Median | 53 | 54 |
| Range | 36–77 | 32–76 |
| Tumor grade |
| Well/moderate | 26 | |
| Poor | 14 | |
| Tumor stage |
| T1 | 19 | |
| T2 | 13 | |
| T3 | 0 | |
| T4 | 8 | |
| Node status |
| N0 | 21 | |
| N1 | 9 | |
| N2 | 7 | |
| N3 | 3 | |
| Metastasis | | |
| M0 | 34 | |
| M1a/b | 6 | |
| TNM stage |
| I | 16 | |
| II | 10 | |
| III | 8 | |
| IV | 6 | |
Sample collection and processing
Peripheral blood samples were collected from the
subjects before any regional or systemic anticancer treatments and
re-collected from 15 patients after thoracic surgery. The fresh
peripheral blood of all individuals was stored in heparin-coated
tubes (BD Biosciences, San Jose, CA, USA) and centrifuged at 4,000
rpm for 10 min at 4°C. Then, the cell-free supernatants, allocated
into 1.5-ml Eppendorf tubes, were frozen at −80°C for the detection
of cytokines (30). The cell
pellets were re-suspended in saline for further analyses of flow
cytometry and real-time quantitative reverse transcriptase
polymerase chain reaction (qRT-PCR) (30).
Flow cytometric analysis
Human peripheral blood mononuclear cells (PBMCs),
isolated from cell pellets using Ficoll-Hypaque density gradient
centrifugation, were re-suspended in RPMI-1640 medium supplemented
with 10% fetal bovine serum, 2 mM glutamine, 100 U/ml penicillin
and 100 µg/ml streptomycin (Invitrogen Life Technologies,
Carlsbad, CA, USA). Next, they were stimulated for 5 h with 50
ng/ml phorbol 12-myristate 13-acetate (PMA; BioVision, Mountain
View, CA, USA), 1 µg/ml ionomycin (Enzo Life Sciences, Inc.,
Farmingdale, NY, USA) and 500 ng/ml monensin (eBioscience, San
Diego, CA, USA) in 24-well plates (16). To analyze IL-17A-producing T cells,
stimulated PBMCs were stained with phycoerythrin (PE)-conjugated
anti-human CD3, fluorescein isothiocyanate (FITC)-conjugated
anti-human γδTCR, allophycocyanin (APC)-conjugated anti-human CD8,
and Pacific Blue-conjugated anti-human CD4 antibodies at 4°C for 30
min (16). Then, the cells, fixed
and permeabilized with IC fixation/permeabilization buffer
(eBioscience), were intracellularly stained with
PerCP-Cy5.5-conjugated anti-human IL-17 antibody according to the
manufacturer's instructions (16).
All antibodies used in the flow cytometric analysis were obtained
from Biolegend (San Diego, CA, USA) and isotype-matched antibody
controls were used in all procedures. Flow cytometric acquisition
was performed using a FACSCalibur (BD Biosciences), and data were
analyzed using FlowJo software, version 7.6.5 (TreeStar, Inc., San
Carlos, CA, USA).
ELISA measurement of serum cytokines
The cell-free supernatants of all individuals were
tested using ELISA kits according to the manufacturer's
instructions (eBioscience), for cytokines including IL-17A, IL-23,
IL-1β, and TGF-β1. All samples were tested in triplicate.
qRT-PCR
RNA samples were prepared from stimulated human
PBMCs using TRIzol (Invitrogen Life Technologies) (16). cDNA was synthesized using reverse
transcription reagent kits (Takara Biotechnolgy Co., Inc., Dalian,
China) and real-time PCR was performed in triplicate using the
QuantiFast™ SYBR Green PCR kit (Qiagen, Hilden, Germany)
in an ABI 7500 analysis system (Applied Biosystems, Foster City,
CA, USA) (16). The following
primer pairs were used: IL-1β forward,
5′-CCACAGACCTTCCAGGAGAATG-3′, and reverse,
5′-GTGCAGTTCAGTGATCGTACAGG-3′; IL-17A forward,
5′-CGGACTGTGATGGTCAACCTGA-3′, and reverse,
5′-GCACTTTGCCTCCCAGATCACA-3′; IL-23p19 forward,
5′-GAGCCTTCTCTGCTCCCTGATA-3′, and reverse,
5′-GACTGAGGCTTGGAATCTGCTG-3′; TGF-β1 forward,
5′-CAGAAATACAGCAACAATTCCTGG-3′, and reverse,
5′-TTGCAGTGTGTTATCCGTGCTGTC-3′; GAPDH forward,
5′-GGTCTCCTCTGACTTCAACA-3′, and reverse,
5′-GTGAGGGTCTCTCTCTTCCT-3′. The data were analyzed by ABI 7500
software (Applied Biosystems).
Statistical analysis
Values are presented as means ± SEM. Differences
among groups were tested by one-way ANOVA. Differences between two
groups were tested using non-paired Student's t-test. For
non-parametric data, the Mann-Whitney U test was performed between
groups. Correlations between values were determined using
Spearman's correlation coefficient. Analysis was performed with
SPSS statistical software (version 21.0; SPSS, Inc., Chicago, IL,
USA), and P<0.05 was considered statistically significant.
Results
Characteristics of the subjects
Clinical characteristic of the 40 patients with LA
and 35 HCs are summarized in Table
I. The median age was 53 years (range, 36–77 years) in the
patient group including 27 males and 13 females, and 54 years
(range, 32–76 years) in the control group including 24 males and 11
females. Baseline characteristics were balanced between the two
groups. In patients with LA, there were 26 cases with well/moderate
differentiation and 14 cases with poor differentiation; 32 cases in
tumor stage T1+T2 and 8 cases in T3+T4; 21 cases of node status N0
and 19 cases of node status N1+N2+N3; 34 cases in metastasis status
M0 and 6 cases in M1a/b; 26 cases in stages I+II and 14 cases in
TNM stages III+IV.
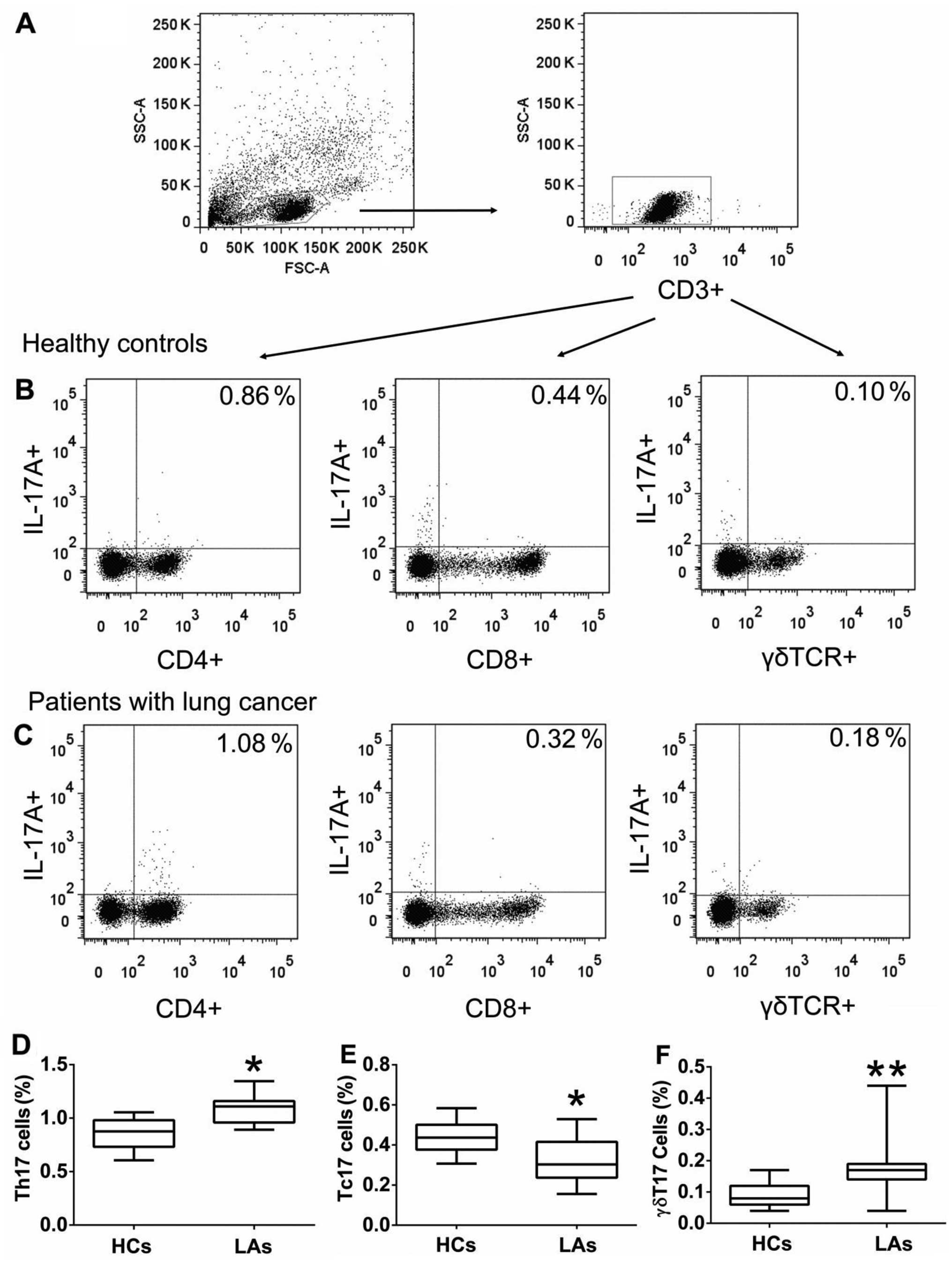
Frequency of circulating Th17, Tc17, and
γδT17 cells in peripheral blood
To investigate the roles of IL-17A-producing T cells
in the development of LC, we first determined the frequencies of
circulating Th17, Tc17, and γδT17 cells in the PBMCs obtained from
all individuals. The frequencies of circulating Th17 and γδT17
cells were considerably higher in patients with LA than those in
the HCs (Fig. 1), but the frequency
of circulating Tc17 cells was markedly lower in patients with LA
than that in HCs (Fig. 1). In
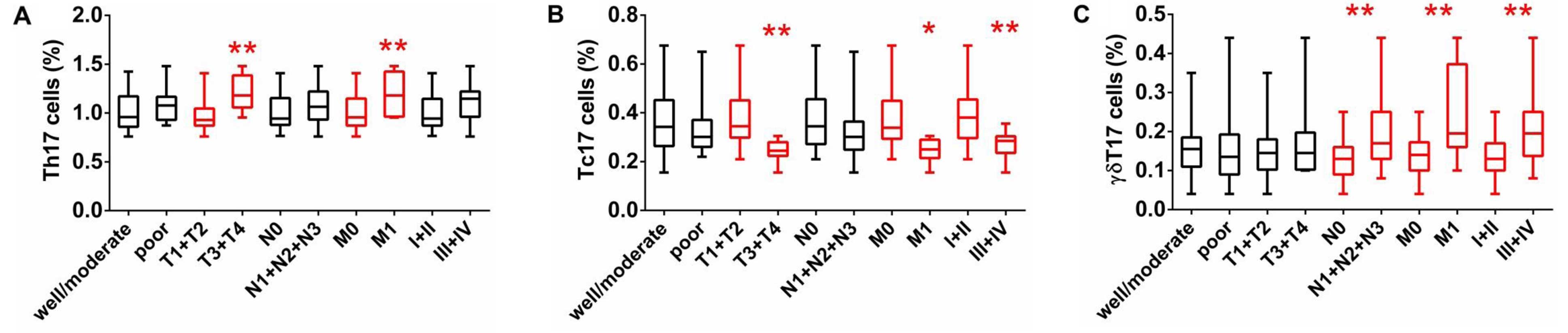
addition, there were no significant differences in the frequencies
of circulating Th17, Tc17, and γδT17 cells regarding tumor
differentiation (well-moderate vs. poor); however, there were
substantial differences between tumor invasion (T1+T2 vs. T3+T4),
distant metastasis (M0 vs. M1a/b), and TNM stage (I+II and III+IV)
(Fig. 2). The percentage of Th17
cells in patients with extensive tumor invasion (T3+T4) was higher
than that in patients with T1+T2, while the percentage of Tc17
cells was low in patients with T3+T4 and the percentage of γδT17
cells showed no difference in regards to tumor invasion (Fig. 2). The percentage of γδT17 cells in
patients with lymphatic metastasis (N1+N2+N3) was significantly
higher than that in patients with lymphatic metastasis (N0), while
there were no differences in Th17 and Tc17 cells regarding
lymphatic metastasis (Fig. 2). The
percentage of Th17 and γδT17 cells in patients with distant
metastasis (M1) were higher than that in patients without
metastasis (M0), while the percentage of Tc17 cells was low in
patients with distant metastasis (M1) (Fig. 2). The percentage of γδT17 cells was
higher in patients with III+IV stage than that in patients with
I+II stage, while the percentage of Tc17 cells was low in patients
with III+IV stage and there was no difference in the percentage of
Th17 cells regarding TNM staging (Fig.
2). These findings suggest that a high percentage of
circulating Th17 and γδT17 cells may contribute to the metastases
of LA and indicate poor prognosis.
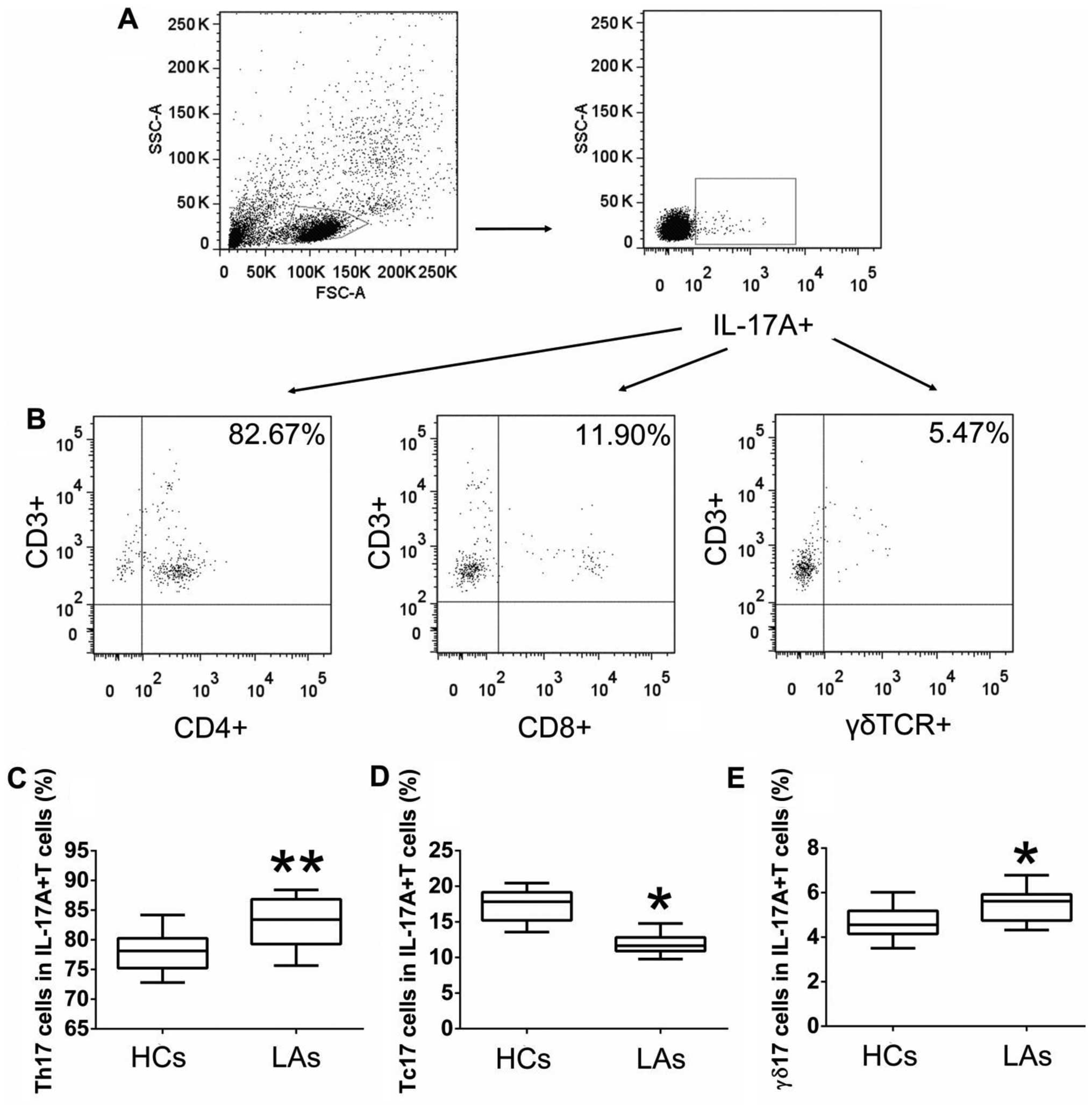
The main T cells secreting IL-17A in
peripheral blood
To further explore the main T cells secreting IL-17A
in the peripheral blood, we measured the frequencies of circulating
Th17, Tc17, and γδT17 cells in gated intracellular
IL-17A+ cells obtained from patients with LA and HCs by
flow cytometric analysis. The highest population among the
intracellular IL-17A+ T cells in both patients with LA
and HCs was circulating Th17 cells, followed by Tc17 and γδT17
cells (Fig. 3). Furthermore, the
percentages of Th17 and γδT17 cells in the total intracellular
IL-17A+ cells obtained from patients with LA were higher
than those from the HCs, while the percentage of intracellular
IL-17A+ cells represented by circulating Tc17 cells was
low in patients with LA compared to that in patients with LCs
(Fig. 3). These data indicated that
the main source of intracellular IL-17A in the peripheral blood was
Th17 cells for both the patients and the HCs, and that circulating
IL-17A+ T cells and IL-17A play key roles in the
initiation and progression of LC.
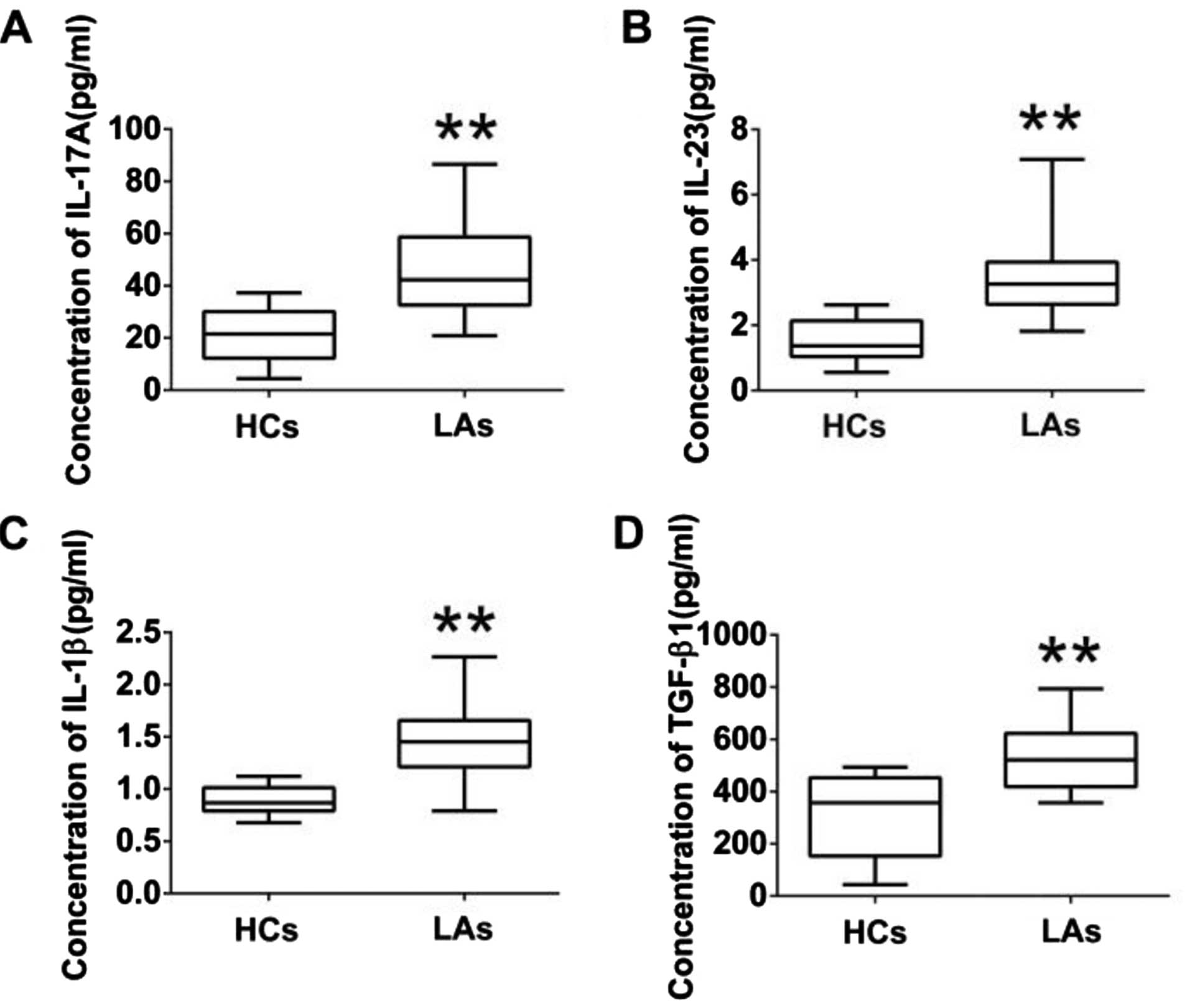
Analysis of IL-17A and associated
cytokines in peripheral blood
To further explore the functions of IL-17A-producing
T cells, the protein levels of IL-17A and associated cytokines
(IL-23, IL-1β, and TGF-β1) in serum and their mRNA levels in PBMCs
from both the patients and the HCs were measured by ELISA and
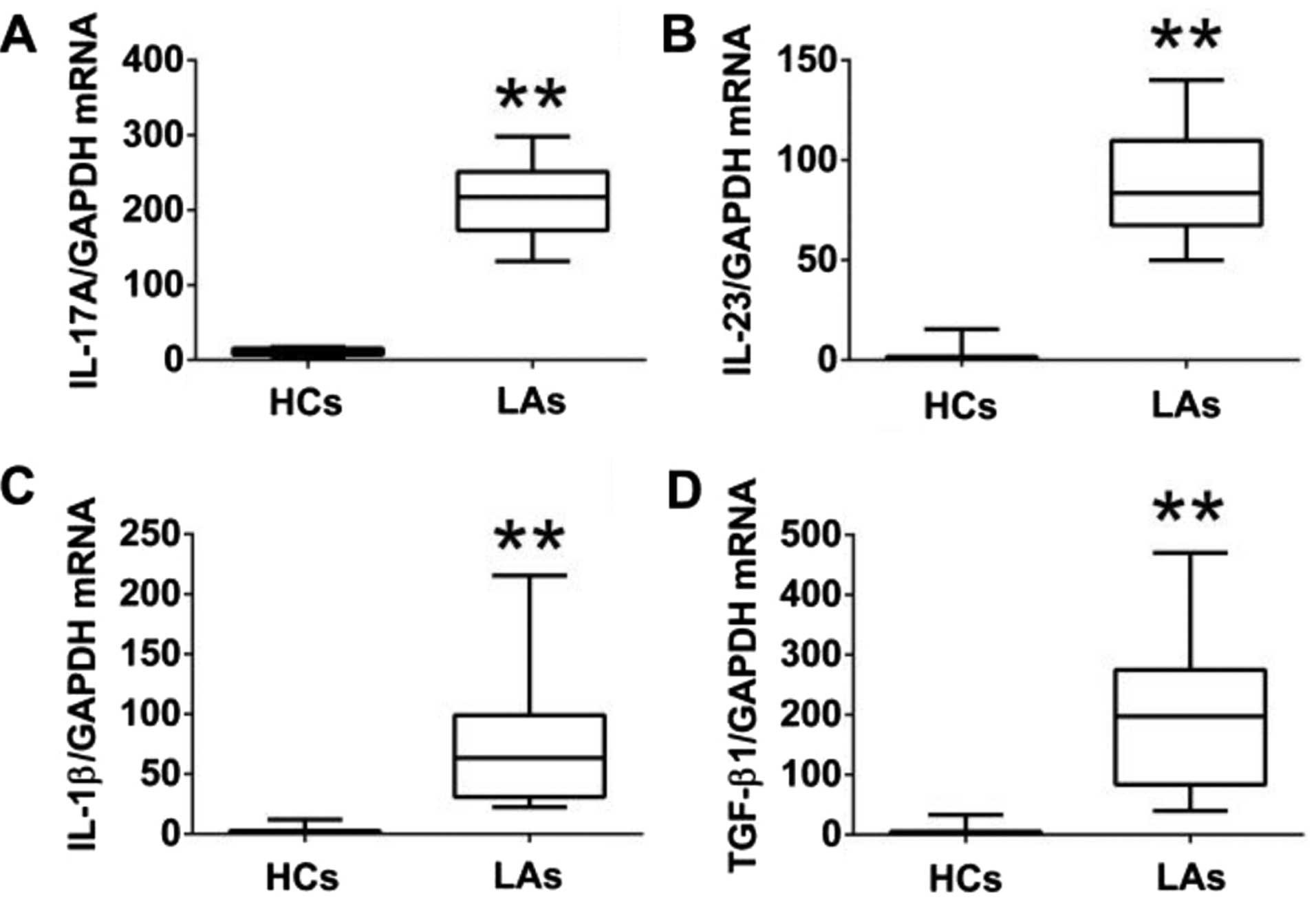
qRT-PCR. The protein levels of IL-17A, IL-23, IL-1β, and TGF-β1 in
the serum of patients with LA were much higher than those in the
HCs (Fig. 4). Similarly, the mRNA
levels of IL-17A IL-23, IL-1β, and TGF-β1 in PBMCs of patients with
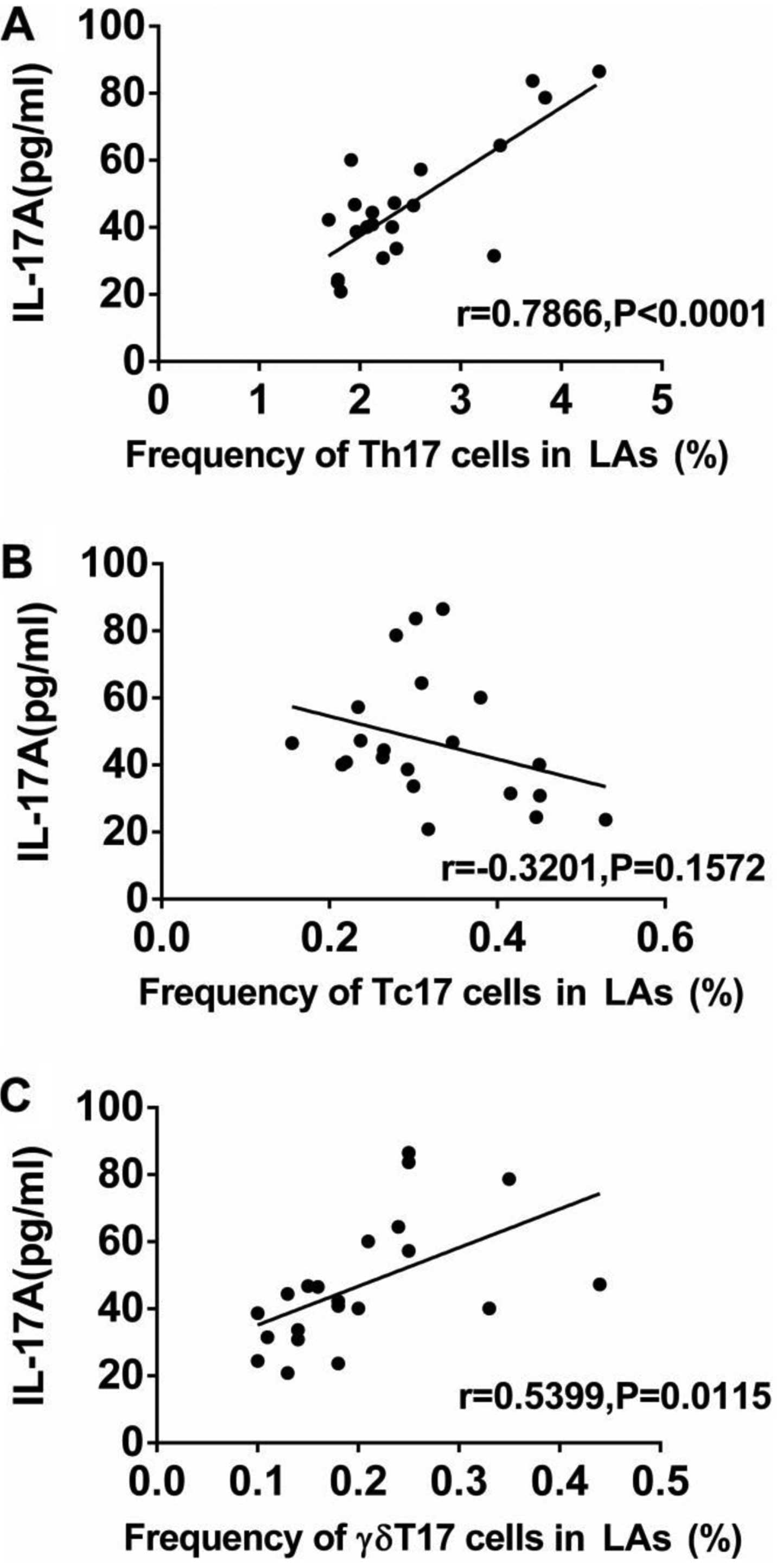
LA were markedly higher than those in the HCs (Fig. 5). In addition, we analyzed the
association between the levels of IL-17A and the frequencies of
Th17, Tc17 and γδT17 cells in patients with LA. The expression of
IL-17A in serum from patients was positively associated with the
frequencies of Th17 and γδT17 cells, but was not related to the
frequency of Tc17 cells (Fig.
6).
Alterations of IL-17A-producing T cells
and IL-17A-associated cytokines in patients with LA after thoracic
surgery
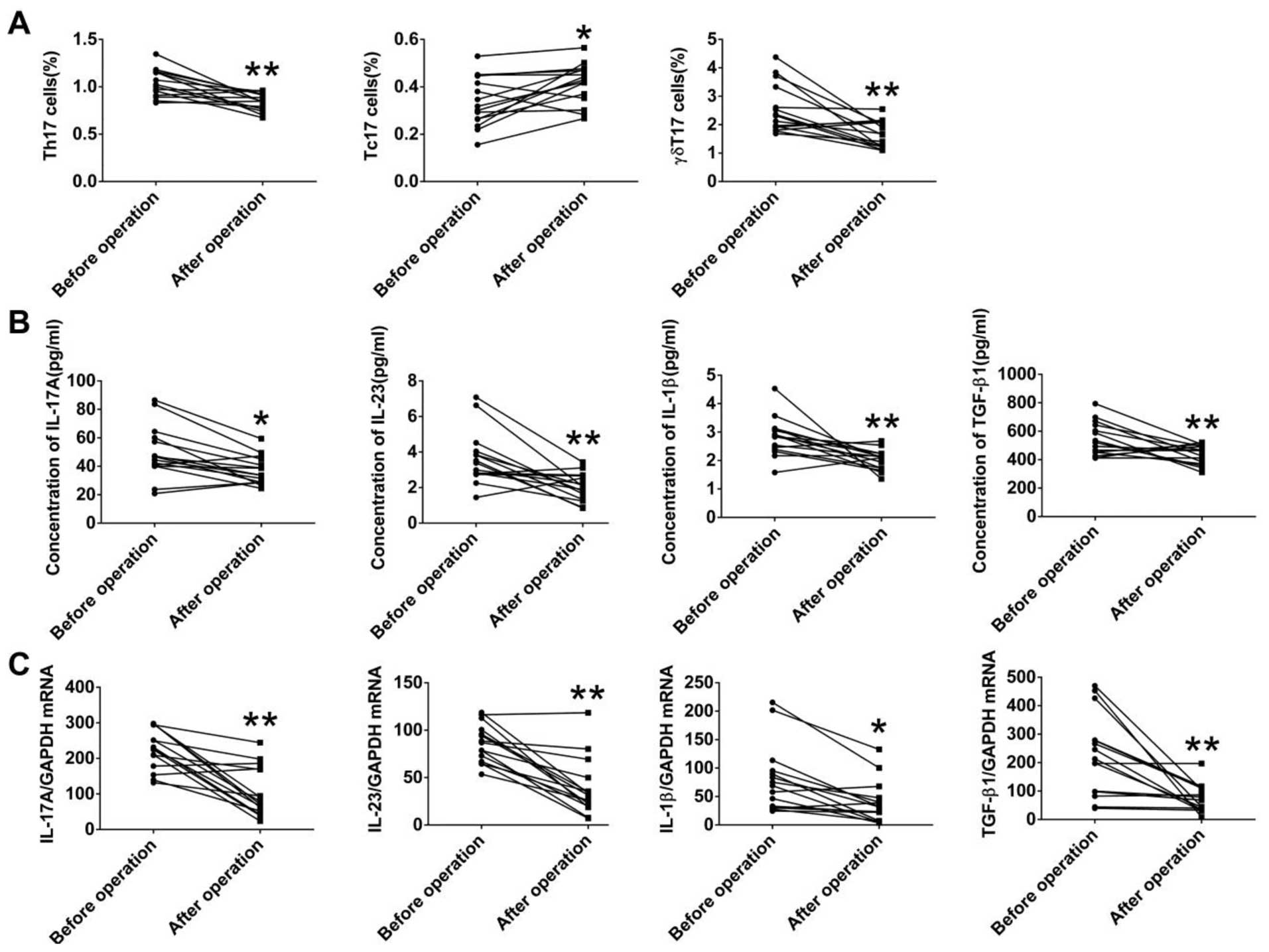
To explore the effects of tumor burden on
IL-17A-producing T cells, we also measured both the frequencies of
circulating Th17, Tc17, and γδT17 cells and levels of IL-17A and
associated cytokines in 15 patients with LA, who received surgery,
at 1 month after the resection. The frequencies of Th17 and γδT17
cells in the patients with LA were markedly decreased after
surgery, while the frequency of Tc17 was significantly increased
after surgery (Fig. 7A).
Furthermore, both the protein levels of IL-17A IL-23, IL-1β, and
TGF-β1 in serum and the corresponding mRNA levels in PBMCs were
markedly decreased in the patients with LA after surgery (Fig. 7B and C). These data suggested that
tumor resection resulted in the alteration of IL-17A-producing
cells, IL-17A-associated cytokines, and the tumor-related
microenvironment.
Discussion
Evidence has shown the dual roles of T cells and
associated cytokines in the initiation, progression, and metastasis
of LC. The present study investigated the frequencies of
IL-17A-producing cells (Th17, Tc17, and γδT17 cells), and
expression levels of IL-17-associated cytokines in patients with LA
and HCs. We found that the frequencies of circulating Th17 and
γδT17 cells, and the protein and corresponding mRNA levels of
IL-17A, IL-23, IL-1β, and TGF-β1 in the peripheral blood of
patients with LA were higher than those in the HCs, whereas the
frequency of Tc17 cells in patients with LA was decreased.
Moreover, the frequencies of circulating Th17 and γδT17 cells,
along with the levels of IL-17A, IL-23, IL-1β, and TGF-β1 were
decreased in patients with LA after tumor resection. Our findings
suggest that Th17, γδT17, Tc17 cells, and IL-17A-associated
cytokines contribute to the development of LA and thus represent
promising targets for therapeutic strategies.
Th17 cells were originally discovered to promote
inflammatory responses in autoimmune diseases and act in host
defenses against microbes (32,33).
Recently, their functions in the initiation and development of
cancers have been extensively studied (34). However, the roles of Th17 cells and
its main cytokine, IL-17A, in carcinogenesis are still
controversial (35). Accumulation
of tumor-infiltrating Th17 cells have been found in a variety of
cancers, including gastric, hepatocellular, breast, and LCs, and
contributed to poor patient prognosis (28,36–38).
Cytokines secreted by cancer cells, such as RANTES, MCP1, IL-6,
IL-1β, and CCL 20, contribute to the differentiation and expansion
of Th17 cells, in the tumor microenvironment (35,39).
Moreover, Th17 cells and IL-17 resulted in the recruitment of
Gr-1+CD11b+ myeloid cells, promoting the
inflammation and tumor growth in a mouse model with oncogenic K-ras
mutation expressed only in the lungs (28). Smoking-mediated Th17 inflammation by
induction of osteopontin and IL-17A deficiency attenuated
smoking-induced emphysema in mice (40). In contrast, elevated Th17 cells or a
high ratio of Th17/Treg, in malignant pleural effusion, partially
promoted by chemokines, predicted a good prognosis in patients with
LC (30,41). Th17 cells may promote the
recruitment of dendritic cells and subsequent activation of
CD8+ T cells in tumors, exerting antitumor immunity
(26). In IL-17-deficient mice,
tumor growth and lung metastasis were augmented, accompanied by
decreased natural killer cells and T cells, suggesting the
protective roles of endogenous IL-17 and Th17 in tumors (25). Consistent with previous studies, our
results demonstrated that the frequency of circulating Th17 cells
was increased in the patients with LA, and was associated with
tumor invasion and metastasis, indicating the pro-tumoral roles of
Th17 cells in the initiation and progression of LA (42). It was also confirmed that Th17 cells
were the main source of IL-17A in human peripheral blood in both
patients with LA and HCs. However, the mechanisms involved in the
accumulation of Th17 cells in the tumor microenvironment are still
unknown, and Th17 cells could be induced, expanded, or converted
from other T cells.
Tc17 cells, a minor subset of CD8+ T
cells characterized by the production of IL-17, play various roles
in infection, cancers and autoimmune diseases (43,44).
Although most of the knowledge concerning the differentiation and
plasticity of Tc17 cells stems from autoimmune diseases, emerging
evidence has also shown that Tc17 cells are associated with the
development of cancers in both animal models and humans (43). In patients with hepatocellular
carcinoma, tumor-activated monocytes triggered the proliferation of
Tc17 cells at the edge of invading tumors, by a set of
proinflammatory cytokines (45). In
patients with uterine cervical cancer, high levels of Tc17 cells
were found in both the peripheral blood and in cervical tissues,
and were involved in the metastases of pelvic lymph nodes and tumor
vasculogenesis (46). Notably,
adoptive transfer of tumor-specific Tc17 cells led to the
regression of melanoma in mice by the recruitment of neutrophils
and induction of chemokines, and elicited antitumor immunity, which
was reversed by the blocking of the inducible costimulator pathway
(47–49). The numbers of both CD4+
and CD8+ T cells were significantly elevated in lung
tumor tissues compared with HCs, and were positively related to the
staging of cancers (50). In spite
of some functional similarities shared with Th17 cells, it has
become gradually clear that Tc17 cells are a distinct subset of T
cells regarding differentiation, development, and plasticity.
Contrary to Th17 cells, the frequency of circulating Tc17 cells was
reduced in the patients with LA, especially those with tumor
invasion and distant metastasis. These results are consistent with
data from patients with thyroid tumors or gastric carcinoma,
suggesting that Tc17 cells may exhibit some antitumor activities in
lung carcinoma and Th17 cells may suppress the development of Tc17
in the tumor microenvironment (16,51).
γδT cells, compared with conventional αβT cells,
exhibit distinct and versatile functions by recognition of
non-peptide antigens, production of proinflammatory cytokines
(IL-17, interferon-γ, and TNF-α), and interaction with activation
of adaptive immune cells (21,52).
γδT17 cells, the primary source of IL-17 in the early stage of some
diseases and one of the pivotal players in immune surveillance,
display diverse responses to tumors (20,53).
In patients with colorectal cancers, γδT17 cells were found to be a
major source of IL-17, related to the expansion of myeloid-derived
suppressor cells, and positively associated with tumor stages
(54). Murine γδT17 cells also
mobilized small peripheral macrophages, which expressed
proangiogenic and proinflammatory mediators, and promoted ovarian
cancer growth in vivo (55).
Recently, in a murine model of breast cancer, γδT17 cells resulted
in expansion and polarization of specific neutrophils which
subsequently inhibited cytotoxic CD8+ lymphocytes, and
led to pulmonary and lymph nodal metastases, indicating a
cooperative mechanism among γδT17 cells, cytotoxic T cells and
neutrophils in the metastatic microenvironment (18). In our study, γδT17 cells were the
third source of IL-17A, which were consistent with gastric patients
but not with colorectal cancers (16,54).
In addition, increased frequency of γδT17 cells was found in
patients with LA and was positively related to the metastasis and
staging of cancers, and was markedly decreased after the resection
of the tumor. The prevalence and variety of γδT17 cells in patients
with LA were very similar with those of Th17 cells, suggesting
these two IL17-producting T cells may collaboratively promote
pulmonary carcinogenesis.
In response to stress, injury, and pathogenic
stimuli, IL-17-associated cytokines, including IL-23, IL-1β, and
TGF-β1, drive the differentiation of naïve T cells into
IL-17-producing T cells (14,43,56).
IL-23 further induces the production of IL-17 by Th17 and γδT
cells, and promotes tumor growth (22,57).
IL-17 targets myeloid and mesenchymal cells, and induces tissue
inflammation by promoting the expression of proinflammatory
cytokines, chemokines, and antimicrobial peptides (10). In addition, IL-17 resulted in the
infiltration of myeloid-derived suppressor cells and angiogenesis
in tumors, and contributes to the tumor-promoting microenvironments
in mice (58,59). Elevated levels of IL-17 were found
in patients with gastric, colorectal and prostatic cancers, and are
associated with poor prognosis (60). Recently, increased levels of IL-23,
IL-1β, and IL-17A were found in gastric patients and were
positively related to tumor invasion and metastasis (24). In experimental silicosis, IL-17A
produced by both Th17 and γδT17 cells was required for acute
pulmonary inflammation and injury, but not chronic responses and
fibrosis (61). Our study showed
that both the mRNA and protein levels of IL-17A IL-23, IL-1β, and
TGF-β1 in PBMCs of patients with LA were markedly higher than those
in the HCs. In addition, the expression of IL-17A in serum was
positively associated with the number of Th17 and γδT17 cells, but
not Tc17 cells. Results indicated that these inflammatory cytokines
contribute to the proliferation of Th17 and γδT17 cells, and the
progression of LC in the tumor microenvironment.
We further explored the effects of the resection of
lung tumors on the alterations of IL-17-producing T cells and
inflammatory cytokines. Notably, after surgery in patients with LA,
the frequencies of Th17 and γδT17 cells, and cytokines including
IL-17A IL-23, IL-1β, and TGF-β1 were markedly reduced, whereas the
frequency of Tc17 cells recovered, suggesting that removal of
tumors may restore immune hemostasis and surveillance, and
IL-17-producing cells may be critical to tumor progression.
In conclusion, our data demonstrated that the
frequencies of circulating Th17 and γδT17 cells, and the protein
and corresponding mRNA levels of IL-17A, IL-23, IL-1β, and TGF-β1
in the peripheral blood of patients with LA were higher than those
in HCs, whereas the frequency of Tc17 cells in patients with LA was
decreased. In addition, Th17 cells were the major source of IL-17A
in patients with LA and HCs. Moreover, the frequencies of
circulating Th17 and γδT17 cells, along with levels of IL-17A,
IL-23, IL-1β, and TGF-β1 were decreased in the patients with LA
after tumor resection. Our data suggest that Th17, γδT17 and Tc17
cells, and IL-17A-associated cytokines play pivotal roles in the
crosstalk between tumor-related inflammation and immunity, and
contribute to the development of LC. In the future, a better
understanding of the distribution and cooperation among
IL-17-producing T cells would provide us with a rationale for novel
anticancer strategies.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (nos. 81472171, and 81300009), the
Major Project of the Science Technology Department of Zhejiang
Province, China (no. 2012C13022-2), the Zhejiang Provincial Natural
Science Foundation of China (no. LY14H010002), the Key Personnel
Grant of Zhejiang Medicine and Health Platform (no. 2012RCA025),
and the Grant for Returned Overseas Chinese Scholars of the
Personnel Department of Zhejiang Province (no. J20120565).
References
|
1
|
Islami F, Torre LA and Jemal A: Global
trends of lung cancer mortality and smoking prevalence. Transl Lung
Cancer Res. 4:327–338. 2015.PubMed/NCBI
|
|
2
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2016. CA Cancer J Clin. 66:7–30. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
She J, Yang P, Hong Q and Bai C: Lung
cancer in China: Challenges and interventions. Chest.
143:1117–1126. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Balkwill F and Mantovani A: Inflammation
and cancer: Back to Virchow? Lancet. 357:539–545. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Elinav E, Nowarski R, Thaiss CA, Hu B, Jin
C and Flavell RA: Inflammation-induced cancer: Crosstalk between
tumours, immune cells and microorganisms. Nat Rev Cancer.
13:759–771. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Grivennikov SI, Greten FR and Karin M:
Immunity, inflammation, and cancer. Cell. 140:883–899. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dvorak HF: Tumors: Wounds that do not
heal. Similarities between tumor stroma generation and wound
healing. N Engl J Med. 315:1650–1659. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Pálmai-Pallag T and Bachrati CZ:
Inflammation-induced DNA damage and damage-induced inflammation: A
vicious cycle. Microbes Infect. 16:822–832. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xu S and Cao X: Interleukin-17 and its
expanding biological functions. Cell Mol Immunol. 7:164–174. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yao Z, Fanslow WC, Seldin MF, Rousseau AM,
Painter SL, Comeau MR, Cohen JI and Spriggs MK: Herpesvirus Saimiri
encodes a new cytokine, IL-17, which binds to a novel cytokine
receptor. Immunity. 3:811–821. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Rouvier E, Luciani MF, Mattéi MG, Denizot
F and Golstein P: CTLA-8, cloned from an activated T cell, bearing
AU-rich messenger RNA instability sequences, and homologous to a
herpesvirus saimiri gene. J Immunol. 150:5445–5456. 1993.PubMed/NCBI
|
|
13
|
Iwakura Y, Ishigame H, Saijo S and Nakae
S: Functional specialization of interleukin-17 family members.
Immunity. 34:149–162. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Cua DJ and Tato CM: Innate IL-17-producing
cells: The sentinels of the immune system. Nat Rev Immunol.
10:479–489. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Aggarwal S, Ghilardi N, Xie MH, de Sauvage
FJ and Gurney AL: Interleukin-23 promotes a distinct CD4 T cell
activation state characterized by the production of interleukin-17.
J Biol Chem. 278:1910–1914. 2003. View Article : Google Scholar
|
|
16
|
Zhong F, Cui D, Tao H, Du H and Xing C:
IL-17A-producing T cells and associated cytokines are involved in
the progression of gastric cancer. Oncol Rep. 34:2365–2374.
2015.PubMed/NCBI
|
|
17
|
Ma S, Cheng Q, Cai Y, Gong H, Wu Y, Yu X,
Shi L, Wu D, Dong C and Liu H: IL-17A produced by γδ T cells
promotes tumor growth in hepatocellular carcinoma. Cancer Res.
74:1969–1982. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Coffelt SB, Kersten K, Doornebal CW,
Weiden J, Vrijland K, Hau CS, Verstegen NJ, Ciampricotti M,
Hawinkels LJ, Jonkers J, et al: IL-17-producing γδ T cells and
neutrophils conspire to promote breast cancer metastasis. Nature.
522:345–348. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang Q, Liu S, Ge D, Zhang Q, Xue Y,
Xiong Z, Abdel-Mageed AB, Myers L, Hill SM, Rowan BG, et al:
Interleukin-17 promotes formation and growth of prostate
adenocarcinoma in mouse models. Cancer Res. 72:2589–2599. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Silva-Santos B, Serre K and Norell H: γδ T
cells in cancer. Nat Rev Immunol. 15:683–691. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Patil RS, Bhat SA, Dar AA and Chiplunkar
SV: The Jekyll and Hyde story of IL17-producing γδT cells. Front
Immunol. 6:372015. View Article : Google Scholar
|
|
22
|
Wang K and Karin M: The IL-23 to IL-17
cascade inflammation-related cancers. Clin Exp Rheumatol. 33(Suppl
92): S87–S90. 2015.PubMed/NCBI
|
|
23
|
Shalapour S and Karin M: Immunity,
inflammation, and cancer: An eternal fight between good and evil. J
Clin Invest. 125:3347–3355. 2015. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hemdan NY: Anti-cancer versus
cancer-promoting effects of the interleukin-17-producing T helper
cells. Immunol Lett. 149:123–133. 2013. View Article : Google Scholar
|
|
25
|
Kryczek I, Wei S, Szeliga W, Vatan L and
Zou W: Endogenous IL-17 contributes to reduced tumor growth and
metastasis. Blood. 114:357–359. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Martin-Orozco N, Muranski P, Chung Y, Yang
XO, Yamazaki T, Lu S, Hwu P, Restifo NP, Overwijk WW and Dong C: T
helper 17 cells promote cytotoxic T cell activation in tumor
immunity. Immunity. 31:787–798. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu B, Guenther JF, Pociask DA, Wang Y,
Kolls JK, You Z, Chandrasekar B, Shan B, Sullivan DE and Morris GF:
Promotion of lung tumor growth by interleukin-17. Am J Physiol Lung
Cell Mol Physiol. 307:L497–L508. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Chang SH, Mirabolfathinejad SG, Katta H,
Cumpian AM, Gong L, Caetano MS, Moghaddam SJ and Dong C: T helper
17 cells play a critical pathogenic role in lung cancer. Proc Natl
Acad Sci USA. 111:5664–5669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chen X, Wan J, Liu J, Xie W, Diao X, Xu J,
Zhu B and Chen Z: Increased IL-17-producing cells correlate with
poor survival and lymphangiogenesis in NSCLC patients. Lung Cancer.
69:348–354. 2010. View Article : Google Scholar
|
|
30
|
Yang G, Li H, Yao Y, Xu F, Bao Z and Zhou
J: Treg/Th17 imbalance in malignant pleural effusion partially
predicts poor prognosis. Oncol Rep. 33:478–484. 2015.
|
|
31
|
Goldstraw P, Crowley J, Chansky K, Giroux
DJ, Groome PA, Rami-Porta R, Postmus PE, Rusch V, Sobin L;
International Association for the Study of Lung Cancer
International Staging Committee; et al: The IASLC Lung Cancer
Staging Project: proposals for the revision of the TNM stage
groupings in the forthcoming (seventh) edition of the TNM
Classification of malignant tumours. J Thorac Oncol. 2:706–714.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Muranski P and Restifo NP: Essentials of
Th17 cell commitment and plasticity. Blood. 121:2402–2414. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Harrington LE, Hatton RD, Mangan PR,
Turner H, Murphy TL, Murphy KM and Weaver CT: Interleukin
17-producing CD4+ effector T cells develop via a lineage
distinct from the T helper type 1 and 2 lineages. Nat Immunol.
6:1123–1132. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ye J, Livergood RS and Peng G: The role
and regulation of human Th17 cells in tumor immunity. Am J Pathol.
182:10–20. 2013. View Article : Google Scholar :
|
|
35
|
Guéry L and Hugues S: Th17 Cell plasticity
and functions in cancer Immunity. Biomed Res Int. 2015:3146202015.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhang JP, Yan J, Xu J, Pang XH, Chen MS,
Li L, Wu C, Li SP and Zheng L: Increased intratumoral
IL-17-producing cells correlate with poor survival in
hepatocellular carcinoma patients. J Hepatol. 50:980–989. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li Q, Li Q, Chen J, Liu Y, Zhao X, Tan B,
Ai J, Zhang Z, Song J and Shan B: Prevalence of Th17 and Treg cells
in gastric cancer patients and its correlation with clinical
parameters. Oncol Rep. 30:1215–1222. 2013.PubMed/NCBI
|
|
38
|
Chen WC, Lai YH, Chen HY, Guo HR, Su IJ
and Chen HH: Interleukin-17-producing cell infiltration in the
breast cancer tumour microenvironment is a poor prognostic factor.
Histopathology. 63:225–233. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Su X, Ye J, Hsueh EC, Zhang Y, Hoft DF and
Peng G: Tumor microenvironments direct the recruitment and
expansion of human Th17 cells. J Immunol. 184:1630–1641. 2010.
View Article : Google Scholar
|
|
40
|
Shan M, Yuan X, Song LZ, Roberts L,
Zarinkamar N, Seryshev A, Zhang Y, Hilsenbeck S, Chang SH, Dong C,
et al: Cigarette smoke induction of osteopontin (SPP1) mediates
TH17 inflammation in human and experimental emphysema. Sci Transl
Med. 4:117ra92012. View Article : Google Scholar
|
|
41
|
Ye ZJ, Zhou Q, Gu YY, Qin SM, Ma WL, Xin
JB, Tao XN and Shi HZ: Generation and differentiation of
IL-17-producing CD4+ T cells in malignant pleural
effusion. J Immunol. 185:6348–6354. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li S, Li Y, Qu X, Liu X and Liang J:
Detection and significance of TregFoxP3+ and Th17 cells
in peripheral blood of non-small cell lung cancer patients. Arch
Med Sci. 10:232–239. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liang Y, Pan HF and Ye DQ: Tc17 cells in
immunity and systemic autoimmunity. Int Rev Immunol. 34:318–331.
2015. View Article : Google Scholar
|
|
44
|
Kondo T, Takata H, Matsuki F and Takiguchi
M: Cutting edge: Phenotypic characterization and differentiation of
human CD8+ T cells producing IL-17. J Immunol.
182:1794–1798. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Kuang DM, Peng C, Zhao Q, Wu Y, Zhu LY,
Wang J, Yin XY, Li L and Zheng L: Tumor-activated monocytes promote
expansion of IL-17-producing CD8+ T cells in
hepatocellular carcinoma patients. J Immunol. 185:1544–1549. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Zhang Y, Hou F, Liu X, Ma D, Zhang Y, Kong
B and Cui B: Tc17 cells in patients with uterine cervical cancer.
PLoS One. 9:e868122014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yu Y, Cho HI, Wang D, Kaosaard K, Anasetti
C, Celis E and Yu XZ: Adoptive transfer of Tc1 or Tc17 cells
elicits antitumor immunity against established melanoma through
distinct mechanisms. J Immunol. 190:1873–1881. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Garcia-Hernandez ML, Hamada H, Reome JB,
Misra SK, Tighe MP and Dutton RW: Adoptive transfer of
tumor-specific Tc17 effector T cells controls the growth of B16
melanoma in mice. J Immunol. 184:4215–4227. 2010. View Article : Google Scholar
|
|
49
|
Nelson MH, Kundimi S, Bowers JS, Rogers
CE, Huff LW, Schwartz KM, Thyagarajan K, Little EC, Mehrotra S,
Cole DJ, et al: The inducible costimulator augments Tc17 cell
responses to self and tumor tissue. J Immunol. 194:1737–1747. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Banat GA, Tretyn A, Pullamsetti SS,
Wilhelm J, Weigert A, Olesch C, Ebel K, Stiewe T, Grimminger F,
Seeger W, et al: Immune and inflammatory cell composition of human
lung cancer stroma. PLoS One. 10:e01390732015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Jiang G, Ma S, Wei Y, Wu Y, Yu X and Liu
H: The prevalence and distribution of Th17 and Tc17 cells in
patients with thyroid tumor. Immunol Lett. 162:68–73. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vantourout P and Hayday A:
Six-of-the-best: Unique contributions of γδ T cells to immunology.
Nat Rev Immunol. 13:88–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Rei M, Pennington DJ and Silva-Santos B:
The emerging protumor role of γδ T lymphocytes: Implications for
cancer immunotherapy. Cancer Res. 75:798–802. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wu P, Wu D, Ni C, Ye J, Chen W, Hu G, Wang
Z, Wang C, Zhang Z, Xia W, et al: γδT17 cells promote the
accumulation and expansion of myeloid-derived suppressor cells in
human colorectal cancer. Immunity. 40:785–800. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Rei M, Gonçalves-Sousa N, Lança T,
Thompson RG, Mensurado S, Balkwill FR, Kulbe H, Pennington DJ and
Silva-Santos B: Murine CD27− Vγ6+ γδ T cells
producing IL-17A promote ovarian cancer growth via mobilization of
protumor small peritoneal macrophages. Proc Natl Acad Sci USA.
111:E3562–E3570. 2014. View Article : Google Scholar
|
|
56
|
Bystrom J, Taher TE, Muhyaddin MS, Clanchy
FI, Mangat P, Jawad AS, Williams RO and Mageed RA: Harnessing the
therapeutic potential of Th17 cells. Mediators Inflamm.
2015:2051562015. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Langowski JL, Zhang X, Wu L, Mattson JD,
Chen T, Smith K, Basham B, McClanahan T, Kastelein RA and Oft M:
IL-23 promotes tumour incidence and growth. Nature. 442:461–465.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
He D, Li H, Yusuf N, Elmets CA, Li J,
Mountz JD and Xu H: IL-17 promotes tumor development through the
induction of tumor promoting microenvironments at tumor sites and
myeloid-derived suppressor cells. J Immunol. 184:2281–2288. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Chung AS, Wu X, Zhuang G, Ngu H, Kasman I,
Zhang J, Vernes JM, Jiang Z, Meng YG, Peale FV, et al: An
interleukin-17-mediated paracrine network promotes tumor resistance
to anti-angiogenic therapy. Nat Med. 19:1114–1123. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Yang B, Kang H, Fung A, Zhao H, Wang T and
Ma D: The role of interleukin 17 in tumour proliferation,
angiogenesis, and metastasis. Mediators Inflamm. 2014:6237592014.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Lo Re S, Dumoutier L, Couillin I, Van Vyve
C, Yakoub Y, Uwambayinema F, Marien B, van den Brûle S, Van Snick
J, Uyttenhove C, et al: IL-17A-producing gammadelta T and Th17
lymphocytes mediate lung inflammation but not fibrosis in
experimental silicosis. J Immunol. 184:6367–6377. 2010. View Article : Google Scholar : PubMed/NCBI
|