Introduction
Meningioma is a common type tumor of the central
nervous system, divided into three groups: grade I (benign
meningioma), II (Atypical meningioma), III (anaplastic/malignant
meningioma) (1). While grade I
meningioma with its benign clinical behavior with the possibility
of surgical and radiation cure, WHO grade II and III are correlated
with aggressive behavior with higher risk of recurrence, metastasis
and shorter survival times (2).
Moreover, according to the studies, lung, bone, liver, lymph node,
and kidney sites relapse frequently and occasionally undergo
malignant transformation or metastasis (3). Surgery is the mainstay of treatment
for patients with an atypical or malignant meningioma, these
lesions are usually treated with adjuvant external beam
radiotherapy (RT). However, the effect of the postoperative RT for
atypical meningiomas remains controversial, bringing us challenges
for the current treatment strategies of combined surgery and
radiotherapy (4). Therefore,
understanding the molecular mechanisms of invasion or metastasis
will pave the way to the discovery of new therapeutic agents for
decreasing recurrence rate and increasing prognostic accuracy in
human meningioma cells.
HER2 is affiliated with the erythoblastosis oncogene
B (ErbB) receptor tyrosine kinase (RTK) family, also known as
ErbB2, c-erbB2 or HER2/neu, coding a 185 kDa transmembrane tyrosine
kinase receptor protein. HER2 has a key role to form a heterodimer
with other HER members, mediating relevant activation of signal
transduction pathway that control cell growth, differentiation,
motility in normal cells as well as cell proliferation, survival,
invasion, angiogenesis in tumor cells. The two main signaling
pathways: phosphoinositide-3-kinase (PI3K)/AKT and rat
sarcoma/mitogen-activated protein kinase (RAS/MAPK) pathways
mediated by the dimerisation and transphosphorylation of RTKs
(5–7,14).
Molecular mechanisms of HER2-overexpression have been elucidated
both in breast cancers and non-breast cancers (8). Nevertheless, the role of HER2
associated with meningiomas have not been demonstrated completely
(9–12). In our previous study, we provided
evidence that expression of HER2 contributed to the cell
proliferation and invasion and inhibiting apoptosis in the human
malignant IOMM-Lee cells. Upregulation of HER2 impacted the protein
level of PI3K and AKT, thereby illustrating that the PI3K signal
pathway play a important role in mediating invasion in the IOMM-Lee
cells (13). Therefore, it is
speculated that HER-2-RAS-MAPK also plays a role on the
pathogenesis of malignant meningioma.
In the present study, the possible role of RAS-MAPK
signal pathway in cell cycle progression and tumor invasion of the
IOMM-Lee human malignant meningioma cell line was investigated. The
results may open a way to provide new ideas for further
investigation of the invasion and proliferation mechanism of
meningiomas and to find some promising molecular targeted
therapies.
Materials and methods
Cell lines and cell culture
The IOMM-Lee cell lines were incubated at a
humidified 37°C incubator containing 5% CO2 and grown in
Dulbecco's modified Eagle's medium (DMEM; GE Healthcare, Logan, UT,
USA), supplemented with 10% fetal bovine serum (FBS; Gibco-BRl,
Carlsbad, CA, USA), 100 U/ml streptomycin and 100 U/ml penicillin
(Beijing Solarbio Science & Technology Co., Ltd., Beijing,
China), The IOMM-Lee human meningioma cell line was kindly provided
by Dr ? Jensen and Dr ? Gillespie of the University
of Utah (Salt Lake City, UT, USA).
Plasmids and transfection
The acquisition of fragments of the short hairpin
(sh) HER-2 (HER-2-sh) and HER-2-overexpression sequence were
attributed to NCBI references from HanBio (Shanghai, China) and the
NM_004448 GenBank HER2 NCBI reference sequence, respectively,
thereby the HER-2-sh and HER-2-overexpression lentiviral vectors
were constructed. Nonsense sequence lentiviral vectors (NC-sh and
NC-overexpression) were used as negative controls. The HER-2-sh
lentiviral vectors were purchased from HanBio and the
HER-2-overxepression lentiviral vectors were purchased from
GeneChem (Shanghai, China), respectively. The IOMM-Lee cells were
seeded into a 6-well plate and cells at 30–50% confluence were
infected with NC-sh, HER-2-sh, NC-overexpression,
HER-2-overexpression, at a multiplicity of infection of 15 with
polybrene (5 µg/ml; GeneChem), then washed with fresh medium
8 h later. To obtain stable cell lines, selected with 2
µg/ml puromycin (Sigma-Aldrich, St. Louis, MO, USA) for 2
weeks. Stable transformants were examined with fluorescence
microscopy, real-time quantitative polymerase chain reaction
(q-PCR) and western blot analysis.
q-PCR analysis
Total RNAs were extracted from the transfected
IOMM-Lee cells using TRIzol reagent (Invitrogen Life Technologies,
Carlsbad, CA, USA). The RNA quality and concentration were analyzed
by measuring the absorbance at 260 and 280 nm with the spectrometer
(759S; Shanghai Lengguang Technology Co., Ltd., Shanghai, China),
and the A260/A280 ratios were between 1.8 and 2.0. For
single-stranded cDNA synthesis, the RT reaction was performed using
a RevertAid First Strand cDNA synthesis kit (Transgen, Beijing,
China). q-PCR was then performed with 1 µg RNA and 1
µl of the following primers from Invitrogen Life
Technologies: HER-2: forward, 5′-CGGACGCCTGATGGGTTAAT-3′ (120 bp)
and reverse, 5′-ACAGCAAAGGTTCTACCCCG-3′; and GAPDH: forward,
5′-CAGGGCTGCTTTTAACTCTGGT-3′ (203 bp) and reverse,
5′-GATTTTGGAGGGATCTCGCT-3′. The q-PCR procedure conducted in the
ABI PRISM 7500 System (Applied Biosystems, Waltham, MA, USA) was as
follows: denaturing at 95°C for 2 min and 40 cycles of annealing at
95°C for 15 sec and extension at 58°C for 30 sec.
Immunofluorescence (IF) staining
Cells were seeded on an 8-well Lab-Tek Chambered
Coverglass the day prior to experiments. The slides were washed
with phosphate-buffered saline (PBS) and cells were fixed with 4%
paraformaldehyde, followed by permeabilization with methanol. After
washing with PBS, the slides were blocked with 2% bovine serum
albumin (BSA), followed by incubation with primary and secondary
antibodies in 5% BSA. Subsequently, the slides were mounted in the
mounting solution (Invitrogen) containing DAPI for counterstaining
cell nucleus. The cell images were observed and photographed with
an immunofluorescence microscopy.
Western blot analysis
When the stable cells grew in the exponential growth
phase, they were seeded into 6-well plates and allowed to grow
until 80–90% confluence, following which they were lysed in lysis
buffer (Beyotime Institute of Biotechnology, Beijing, China) on
ice. The cells were harvested, washed twice with 1X
phosphate-buffered saline (PBS) and lysed in 100 µl
radioimmunoprecipitation assay lysis buffer [Vazyme Biotech
(Nanjing) Co., Ltd., Nanjing, China]. Protein concentrations were
determined using a bicinchoninic acid kit [Vazyme Biotech (Nanjing)
Co., Ltd.]. The proteins were separated using 10% SDS-PAGE and were
then blotted onto nitro cellulose membranes by wet electroblotting
at a constant current 200 mA for 2 h.
The membranes were blocked with 5% non-fat milk
powder at room temperature for 1 h and incubated overnight with the
following primary antibodies: monoclonal mouse HER-2 (3B5; 1:500;
Abcam, Cambridge, MA, USA), polyclonal rabbit ERK5 (D315V; 1:1,000;
Cell Signaling Technology, Inc., Danvers, MA, USA), polyclonal
rabbit phosphorylated (p)-ERK5 (Thr218/Tyr220; 1:1,000; Cell
Signaling Technology, Inc.), polyclonal rabbit JNK (1:1,000; Cell
Signaling Technology, Inc.), polyclonal rabbit p-JNK
(Thr183/Tyr185; 1:1,000; Cell Signaling Technology, Inc.),
polyclonal rabbit ERK1/2 (137F5; 1:1,000; Cell Signaling
Technology, Inc.), polyclonal rabbit p-ERK1/2 (Thr202/Tyr204;
1:1,000; Cell Signaling Technology, Inc.), polyclonal rabbit P38
(D13E1; 1:1,000; Cell Signaling Technology, Inc.), polyclonal
rabbit p-P38 (Thr180/Tyr182; 1:1,000; Cell Signaling Technology,
Inc.), polyclonal rabbit Ras (27H5; 1:1,000; Cell Signaling
Technology, Inc.), and monoclonal mouse β-actin (T0022; 1:5,000;
Cell Signaling Technology, Inc.) at 4°C. Following incubation, the
membrane was rinsed with Tris-buffered saline with Tween-20 (TBST)
for 15 min three times, and incubated with secondary antibody. The
membrane was agitated for 1 h at room temperature, washed again in
TBST, and were developed using an ECL Plus Western Blotting
Detection System (Bio-Rad Laboratories, Inc., Hercules, CA,
USA).
Kinase inhibitors and MTT assay
PD98059, a special inhibitor of the ERK1/2, was from
(Sigma Company), SP600125, the inhibitor of JNK, was from (Sigma
Company), XMD8-92, an effective inhibitor of the ERK5, from (Santa
Cruz Biotechnology). Treated with different concentration of ERK1/2
inhibitor, JNK inhibitor, ERK5 inhibitor, cell proliferation
ability of each group were assessed using MTT assay. Briefly, the
exponential growth phase, were trypinized, centrifuged at 400 × g
for 5 min at room temperature, resuspend in complete medium (10%
FBS) and were counted. The cells were then seeded into five 96-well
plates (1×103 cells/well), with five parallel wells for
each cell group. The medium was replaced with 120 µl MTT
solution, containing 100 µl medium and 20 µl MTT
(Beijing Solarbio Science & Technology Co., Ltd.), at different
time-points (24, 48, 72 h). After 4 h, the medium was aspirated and
150 µl DMSO was added to each well, then the plate was
agitated for 45 sec at 27°C. The optical density value at a
wavelength of 490 nm was determined using a Multiskan FC Microplate
photometer (Thermo Fisher Scientific, Waltham, MA, USA).
Cell invasion analysis
The invasion assays were performed using Transwell
inserts (Merck Millipore, Billerica, MA, USA; 8 µm pore
size) in 24-well plates. Approximately 1×105 cells in
200 µl of serum-free DMEM-F12 medium were placed in the
upper chamber, and 500 µl of the medium containing 15% FBS
were placed in the lower chamber. For the invasion assay, Transwell
membranes were pre-coated with Matrigel (BD Biosciences, Bedford,
MA, USA). The cells were incubated for 36 h at 37°C in 5%
CO2. The cells were incubated in the uncoated Transwells
for 24 h. Then, the cells were fixed in 10% paraformaldehyde for 15
min and stained with 0.05% crystal violet in PBS for 30 min. The
cells on the upper side of the filters were removed with
cotton-tipped swabs, and the filters were washed with PBS. The
cells invaded through the filter were dried for 20 min, fixed in
absolute alcohol and stained with 8 g/l hematoxylin and eosin. The
cells on the underside of the filters were examined and counted
with a total ×100 magnification under a Leica DMI 4000 microscope.
Each experiment was performed in triplicate and repeated at least
three times.
Effects of ERK1/2, ERK5, JNK inhibitors
on the proliferation and invasion of IOMM-Lee cells transfected
with HER-2 overexpression lentiviral vector
The IOMM-Lee cells were treated with 10 µg/l,
20 µg/l, 40 µg/l PD98059 (ERK1/2 inhibitor), 10
µg/l, 20 µg/l, 40 µg/l SP600125 (JNK
inhibitor), 5 µg/l, 10 µg/l, 20 µg/l XMD8-92
(ERK5 inhibitor) for 30 min, respectively, and then with
HER-2-overexpression cells or HER-2-NC cells for indicated periods.
The ERK1/2, JNK and ERK5 expression was detected by western blot
assay 30 min later, the cell invasion was detected 24 h later, and
the cell proliferation was measured by MTT assay, then observed
after 24, 48, 72 h, respectively.
Animal models
A total of 30 female infant (4–6 weeks) BALB/c nude
mice, of specific pathogen-free grade, weighing 18–20 g, were used
in this study. Rats were obtained from the SJA laboratory Animal
Co., Ltd. (Hunan, China). All nude mice were housed in the
Laboratory Animal Center of the First Affiliated Hospital of
Nanchang University (Nanchang, Jiangxi, China). All nude mice were
maintained in an air-conditioned room with a 12-hour light/dark
cycle, standard diet and water were available ad libitum.
Ethical approval for this study was obtained from the Ethics
Committee of Nanchang University. First, the nude mice were
randomly separated into five groups (n=6/group) (blank, NC-sh,
HER-2-sh, NC-ox, HER-2-ox), After five days, the IOMM-Lee malignant
cells were seeded for each groups. IOMM-Lee cells, in the
exponential growth phase, were trypinized, at 400 × g for 5 min at
room temperature, resuspending in complete medium (10% FBS) and
were counted for 5×107/ml. Then IOMM-Lee cells
(1×107/mice) were injected subcutaneously into the right
armpit of six-week-old nude mice, five mice in each group, and the
blank control was nude mice injected equivalent serum-free DMEM
medium. Close observation after injection, recorded the nude mice
in vivo. The tumors were measured once every 4 days for the
longest diameter a and shortest path b, according to the formula V
= 0.52 ab2 calculation into the tumor size, rendering time-tumor
volume growth curve. All the mice were sacrificed at day 30, the
tumors were removed and calculated, the average tumor weight to
calculate the inhibitory rate. Tumor inhibitory rate = (control
group − experimental tumor) / control tumor weight by 100%.
Statistical analysis
Data were expressed as mean ± standard error of the
mean (SEM), either Student's t-test or one-way analysis of variance
(ANOVA) was used for statistical analysis. All analyses were
performed using the Statistical Package for Social Sciences (SPSS)
17.0 software (SPSS, Chicago, IL, USA) and P<0.05 was considered
to be statistically significant.
Results
Expression of HER-2 in human malignant
meningioma cell lines
To analyze the role of HER-2 in the IOMM-Lee cell
lines, the present study assessed the expression of HER-2 in the
IOMM-Lee cell lines applying q-PCR following transfection for 96 h
(Fig. 1A and B). Compared with the
mock or the cells transfected with NC-sh in the levels of HER-2,
the HER-2-expression of transfected with HER-2-sh1, HER-2-sh2,
HER-2-sh3 all in the IOMM-Lee cells decreased, but HER-2-sh2
decreased 75.3% (P<0.01). The HER-2-expression in the
HER-2-overexpression IOMM-Lee cells was significantly increased
(8.7-fold), compared with the blank or NC-overexpression cells
(P<0.01). Next, we also found similar effects on the protein
levels of HER-2, 72 h post-infection in the western blotting. As
shown in Fig. 1C, it revealed that
the protein level of HER-2 in the HER-2-sh2 group significantly
decreased, by 60.67%. Compared with the blank, NC cells or
HER-2-sh1, HER-2-sh3 (P<0.01). As shown in Fig. 1D the expression of HER-2 in the
HER-2-overexpression group increased (2.6-fold), compared with mock
or NC cells (P<0.01). In the next experiments, we elected the
HER-2-sh2 as the HER-2-sh group because of its high efficiency of
transfection with the silence lentiviral vector.
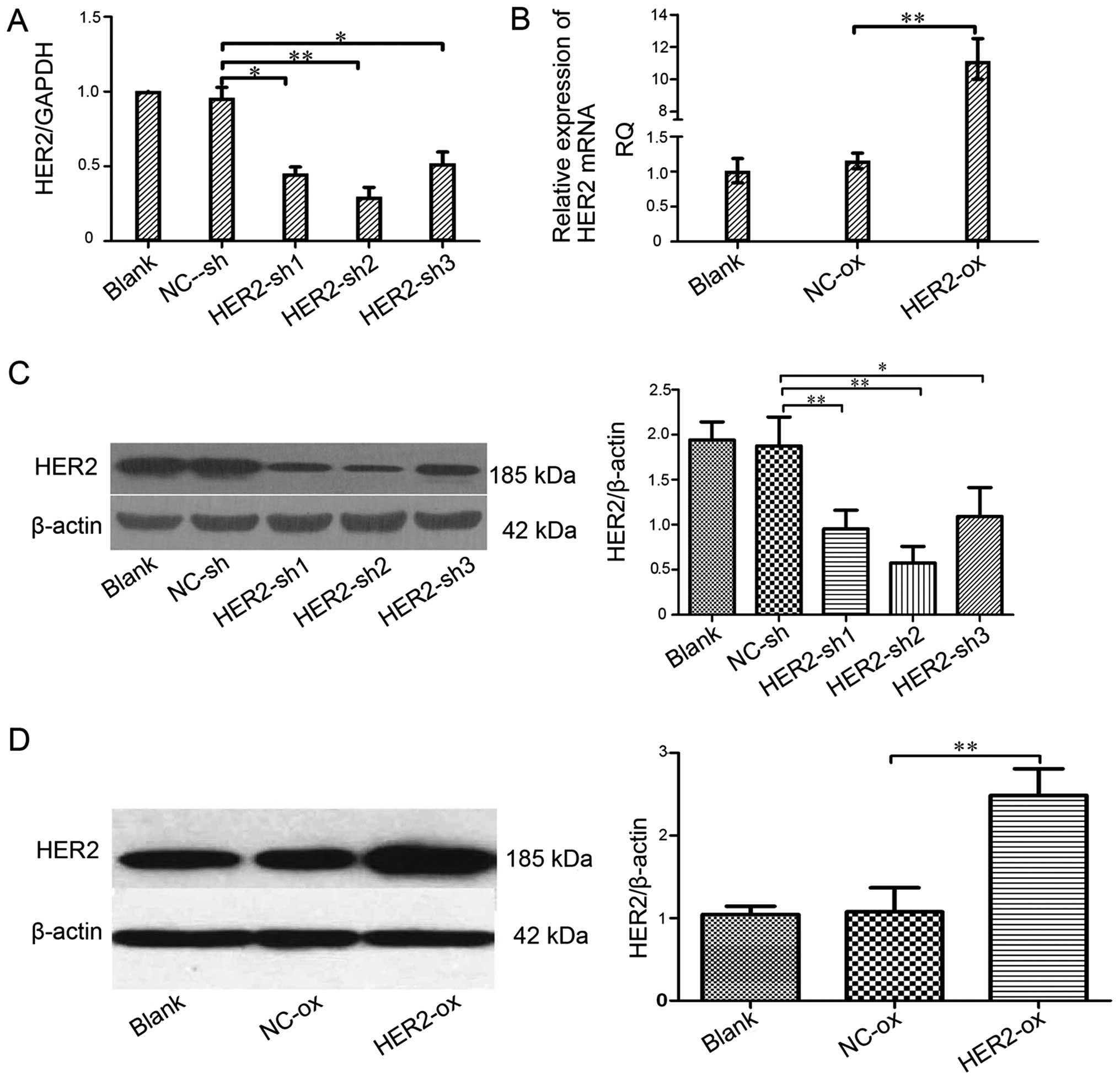 | Figure 1Effect of the HER-2 gene on the
expression of HER-2 in IOMM-Lee cells. (A) The levels of HER-2 in
different cell lines. Compared with NC-sh, the levels of HER-2 in
the IOMM-Lee cell lines transfected with HER2-sh1, HER2-sh2,
HER2-sh3 were decreased, obviously HER2-sh2
(**P<0.01). (B) The mRNA levels of HER-2 in the
IOMM-Lee malignant meningioma cells. Compared with the
NC-overexpression, the expression of HER-2 in the IOMM-Lee cells
transfected with HER-2-over was significantly increased
(**P<0.01). (C and D) The protein expression of HER-2
in IOMM-Lee cells transfected with HER-2-sh and HER-2-over were
analyzed by western blotting. (C) The protein expression levels of
HER-2 in the IOMM-Lee cell lines transfected with HER-2-sh2
obviously decreased (**P<0.01). The data are
expressed as the mean standard deviation from three independent
experiments. (D) The protein expression levels of HER-2 in the
IOMM-Lee cell lines transfected with HER-2-over obviously increased
(**P<0.01). The data are expressed as the mean
standard deviation from three independent experiments. Blank, blank
control; NC, negative; sh, short hairpin, over, overexpression. |
IOMM-Lee cells transfected with the
HER-2-overexpression lentiviral reflected by
immunofluorescence
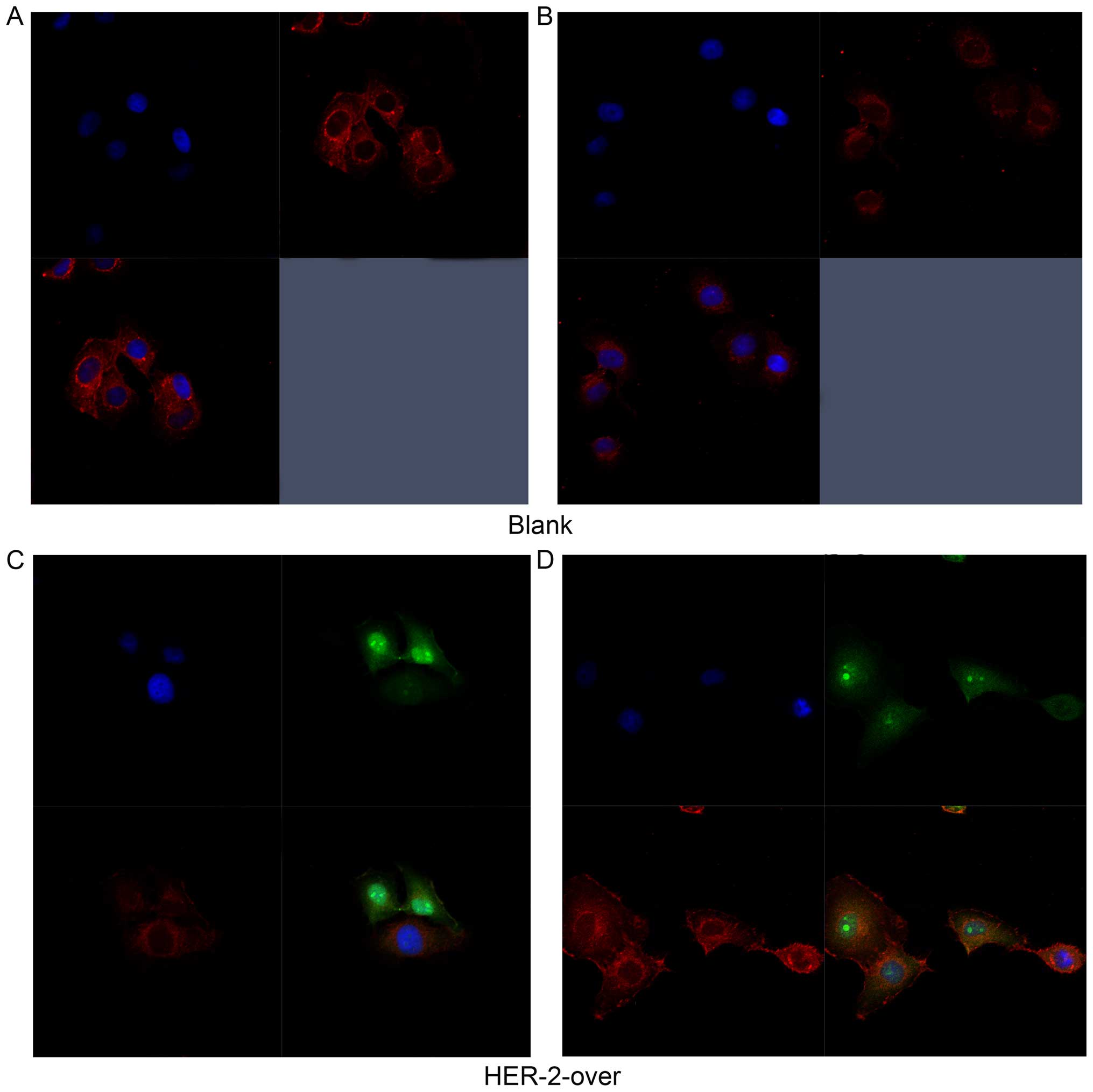
In Fig. 2A and B,
the immunofluoresence results revealed the expression of HER-2 both
cell surface and part of cytoplasm reflected by the red
fluorescence in the blank group. As shown in Fig. 2C, the results suggested that HER-2
express in the cell surface in the IOMM-Lee cells transfected with
HER-2-overexpression lentiviral vector refected by the green
fluorescence, but the positive intensity of red fluoresence
decreased, compared with the blank group. However, in Fig. 2D, the red fluorescence intensity
significantly heighten suggested that the expression of HER-2
increased in the HER-2-overexpression IOMM-Lee cells, compared with
blank group. Thus, we chose the HER-2-overexpression cells for the
further experiments.
HER-2 affects protein expression levels
of ERK5, JNK, ERK1/2, P38 and Ras in IOMM-Lee cells without any
inhibitors
In order to investigate the association between
HER-2 and the activity of the Ras/MAPK signaling pahway, the
present study measured the protein levels of ERK5, p-ERK5, JNK,
p-JNK, ERK1/2, p-ERK1/2, P38, p-P38, Ras in the IOMM-Lee cells
following transfection. The results shown that, compared with NC-sh
group, the protein levels of ERK5, p-ERK5, JNK, p-JNK, ERK1/2,
p-ERK1/2, Ras in the downregulated HER-2 group were all reduced,
with significant difference between the two groups, with
statistical significance (P<0.05; Fig. 3A and B). By contrast, in the
upregulated HER-2 group, the protein levels of ERK5, p-ERK5, JNK,
p-JNK, ERK1/2, p-ERK1/2, Ras all increased (P<0.05; Fig. 3A and B). However, no difference was
observed in the protein expression of P38, p-P38 in these two
groups, compared with NC group.
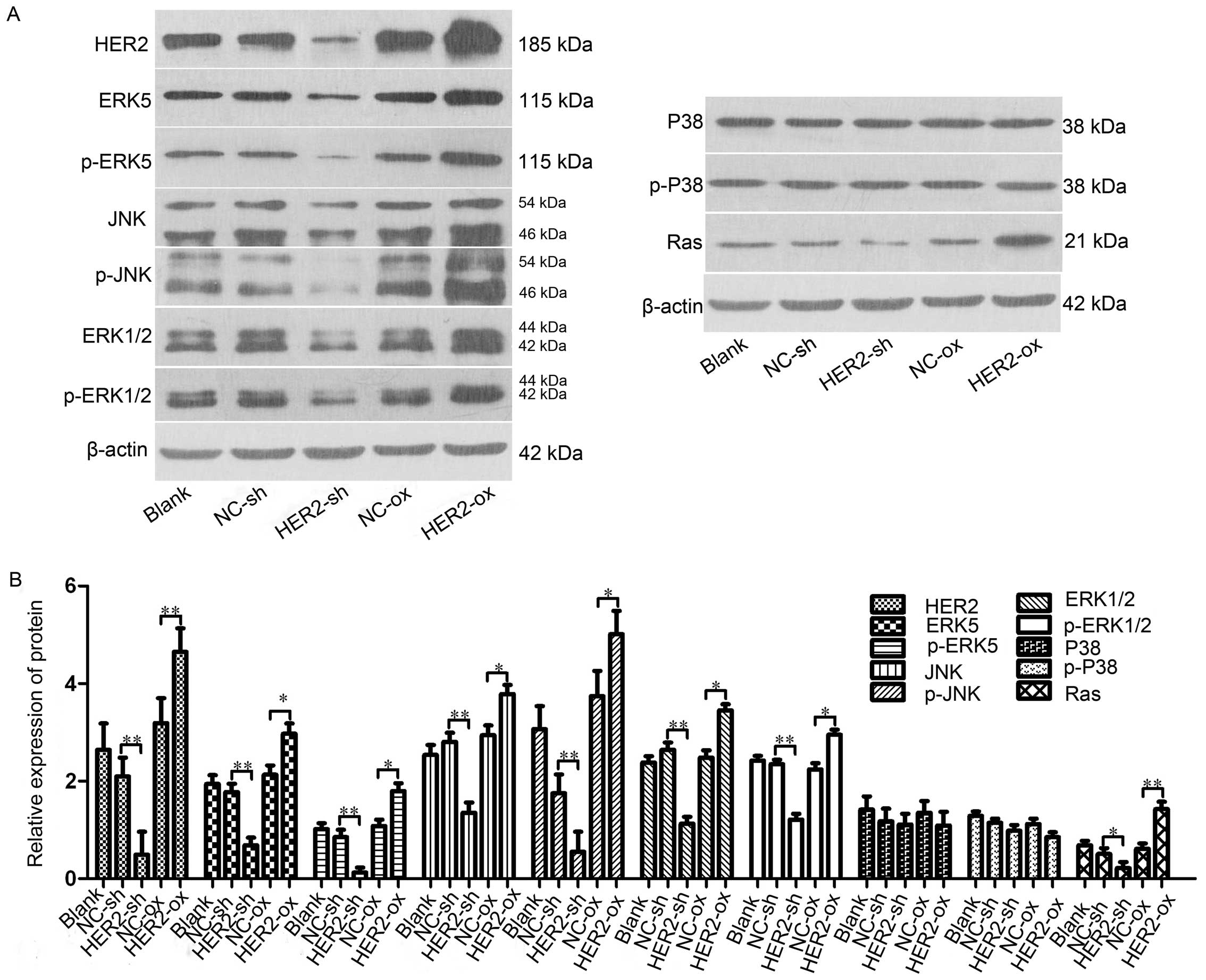 | Figure 3HER-2 affects protein expression
levels of ERK5, JNK, ERK1/2, P38 and Ras in IOMM-Lee cells. (A)
Protein levels of ERK5, p-ERK5, JNK, p-JNK, ERK1/2, p-ERK1/2, P38,
p-P38, Ras in the IOMM-Lee cells were determined using western blot
analysis 72 h post-transfection. β-actin was used as an internal
loading control. (B) Protein levels of ERK5, p-ERK5, JNK, p-JNK,
ERK1/2, p-ERK1/2, Ras in the HER-2-sh group were significantly
decreased. However, in the HER-2-over group, the protein levels of
ERK5, p-ERK5, JNK, p-JNK, ERK1/2, p-ERK1/2, Ras were increased,
compared with NC control and following normalization against
β-actin (P<0.05). No difference was observed between the protein
expression of P38, p-P38 in the HER-2-sh and HER-2-over cells with
that in the NC control. The data are expressed as the mean standard
deviation from three independent experiments. NC, negative control;
sh, short hairpin; over, overexpression; ERK5, extracellular
signal-regulated kinase 5; JNK, Jun N-terminal kinase; ERK1/2,
extracellular regulated protein kinases 1/2; p-,
phosphorylated. |
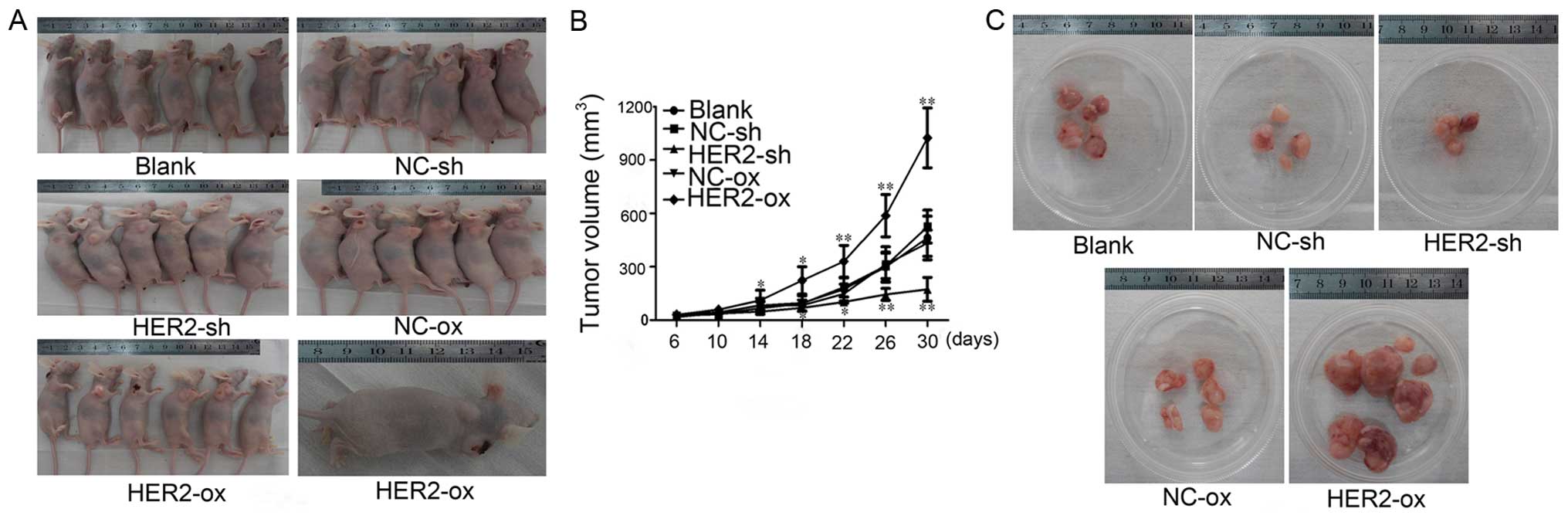
Tumorigenesis of HER-2-overexpression
IOMM-Lee cells in vivo
Our results suggested that HER-2 can increase cell
proliferation in vitro. To confirm this effect in
vivo, the cells (blank, NC-sh, HER-2-sh, NC-ox, HER-2-ox) were
subcutaneously inoculated into nude mice. The tumor was observed in
the blank group, NC-sh group, HER-2-sh group, NC-ox group and
HER-2-ox group on days four after subcutaneous inoculation showed
there were 2, 3, 2, 2, 4 nude mice with solid mass, respectively,
but liquidity bag piece of the blank group had disappeared. The
tumor was observed in the blank group, NC-sh group, HER-2-sh group,
NC-ox group and HER-2-ox group on days six after subcutaneously
inoculation showed there were 4, 4, 3, 5, 5 nude mice with solid
mass, respectively. On day 22, and 26 days after subcutaneous
inoculation, in the HER-2-ox group one nude mouse had macroscopic
transfer. All of the mice were sacrificed at days 30, the tumor at
formation rate of 84% with on natural death (Fig. 4A). Close observation after
injection, on days six, recorded the nude mice in vivo with
the tumor, measured once every 4 days for the longest diameter and
shortest path, calculated the tumor size, rendering time-tumor
volume growth curve. Our result revealed tumor volume change at
days 14 after subcutaneous inoculation. Volume growth of HER-2-sh
decreased by 28.36% compared with NC-sh group, While the volume
growth of HER-2-ox increased by 32.14% compared with the NC-ox
group (P<0.05). After 14 days the volume significantly
increased, the differences were statistically significant
(P<0.01). Time-tumor volume growth result revealed that HER-2
increased cell growth and proliferation in meningioma cells
(Fig. 4B). Tumor volume was
measured once every 4 days using a vernier caliper and the tumors
were collected on day 30. The mean volume of tumors in HER-2-sh
group, NC-sh group, HER-2-ox group, NC-ox group was 139.33±13.89
mg, 236.34±54.18 mg, 357.33±42.24 mg and 223±36.16 mg,
respectively. Tumor inhibitory rate of HER-2-sh group was 41.05%
compared with NC-sh group, HER-2-ox group was −55.64%, the
differences were statistically significant (P<0.01; Fig. 4C). The result also illustrated HER-2
increased the cell proliferation of malignant meningioma in
vivo.
The role of MAPK pathway inhibitions
PD98059 (ERK1/2 inhibitor), SP600125 (JNK inhibitor), XMD8-92 (ERK5
inhibitor) in inhibiting the proliferation and metastasis of
IOMM-Lee cells through the MAPK pathway in vitro
Cell invasion plays a crucial role in the tumor
metastasis. MTT assay result present that at 24, 48, 72 h, the cell
proliferation of the blank group and HER-2-ox group with the
different concentration inhibition of PD98059, XMD8-92, SP600125,
respectively. As shown in Fig. 5,
The present study observed a significant decrease in proliferation
following adding PD98059 in MTT assay. At 72 h, with the
concentration of 10, 20, 40 µg/l PD98059 cell resistance was
observed. However, HER-2-over with concentration of 40 µg/l
ERK1/2 inhibition (PD98059) cell resistance was 1.54-fold higher
than that of HER-2-over without inhibition (P<0.01). Whereas,
the resistance of the HER-2-over with concentration of 40
µg/l of PD98059(ERK1/2 inhibition) cells decrease
significantly 45%, compared with the blank group with the same
concentration PD98059 (P<0.01). Therefore, we elected 40
µg/l PD98059 as the best inhibition concentration for
further experiments.
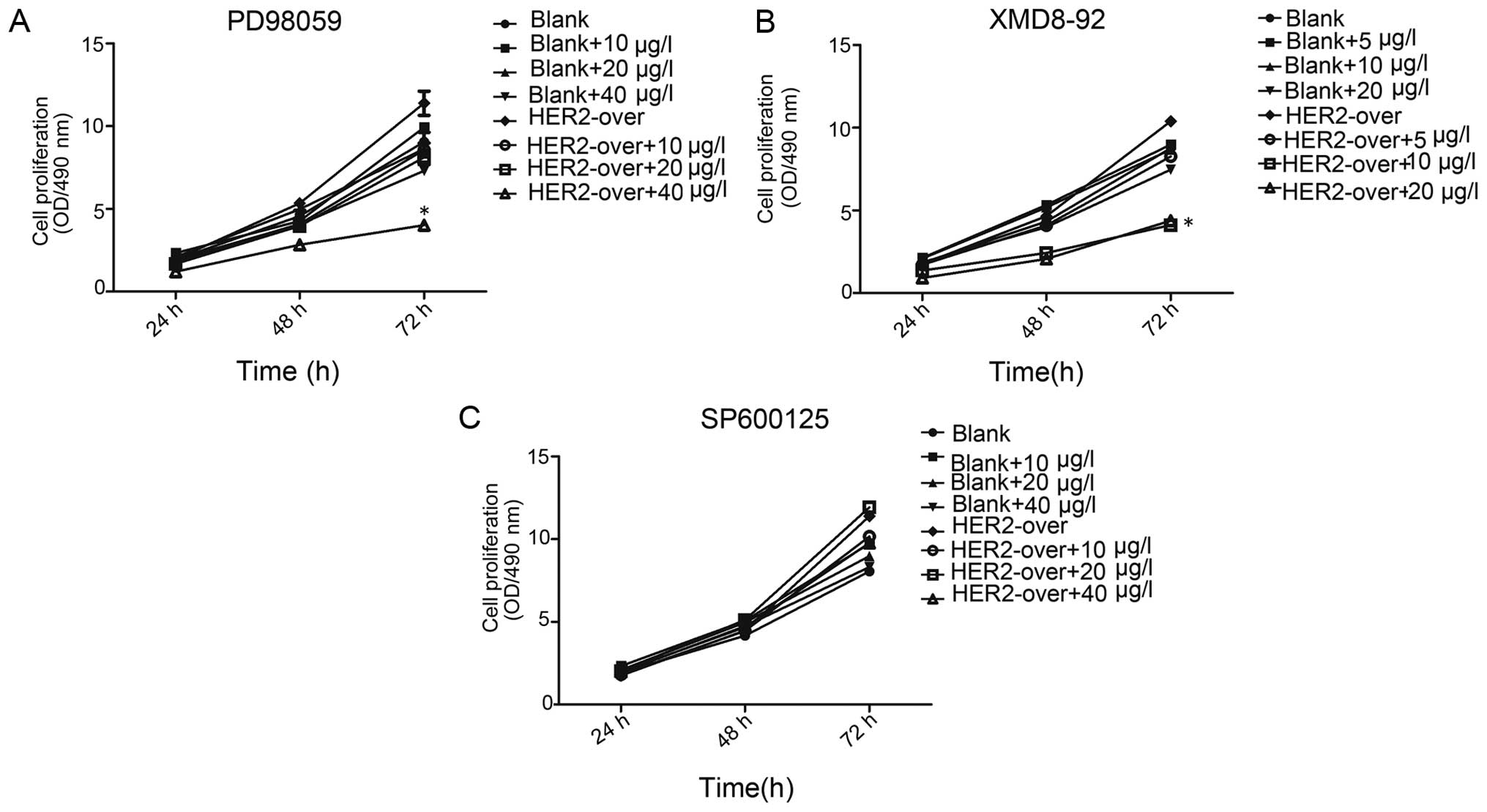 | Figure 5MTT assay present the inhibition of
the cell proliferation. (A) MTT assay at 72 h, in HER-2-over with
concentration of 40 µg/l PD98059 (ERK1/2 inhibition) cell
resistance was 1.54-fold higher than that of HER-2-over without
inhibition (P<0.01), whereas the resistance of the HER-2-over
with concentration of 40 µg/l of PD98059 (ERK1/2 inhibition)
cells significantly decreased 45%, compared with the blank group
with the same concentration of PD98059 (P<0.01). (B) At 72 h,
HER-2-over with concentration of 10 µg/l XMD8-92 (ERK5
inhibition) cell resistance was decreased 73.8% compared with the
HER-2-over without inhibition (P<0.01), whereas the resistance
of the HER-2-over with concentration of 10 µg/l XMD8-92
(ERK5 inhibition) cells significantly decreased 24%, compared with
the blank group with the same concentration PD98059 (P<0.01).
(C) At different time-point (24, 48, 72 h), MTT assay revealed
that, no difference was observed in the groups with the different
concentration of SP600125 (10, 20, 40 µg/l) (P>0.05).
Blank, blank control; over, overexpression. |
In the blank group and HER-2-over group with the
different concentration of XMD8-92 (ERK5 inhibition), in each group
was observed a certain degree of inhibition (P<0.01). However at
72 h, HER-2-over with concentration of 10 µg/l of XMD8-92
(ERK5 inhibition) cell resistance was significantly decreased
73.8%, compared with the HER-2-over without inhibition (P<0.01).
Whereas, the resistance of the HER-2-over with concentration of 10
µg/l XMD8-92 (ERK5 inhibition) cells significantly decreased
24%, compared with the blank group with the same concentration
PD98059 (P<0.01). Thus, in the next experiments we elected the
concentration of 10 µg/l of XMD8-92 as the ideal inhibition
concentration. In addition, we next used the SP600125 (JNK
inhibition) in a similar way to investigate the cell proliferation.
At different time-point (24, 48, 72 h), MTT assay revealed that, no
difference was observed in the groups with the different
concentration of XMD8-92 (10, 20, 40 µg/l) (P>0.05).
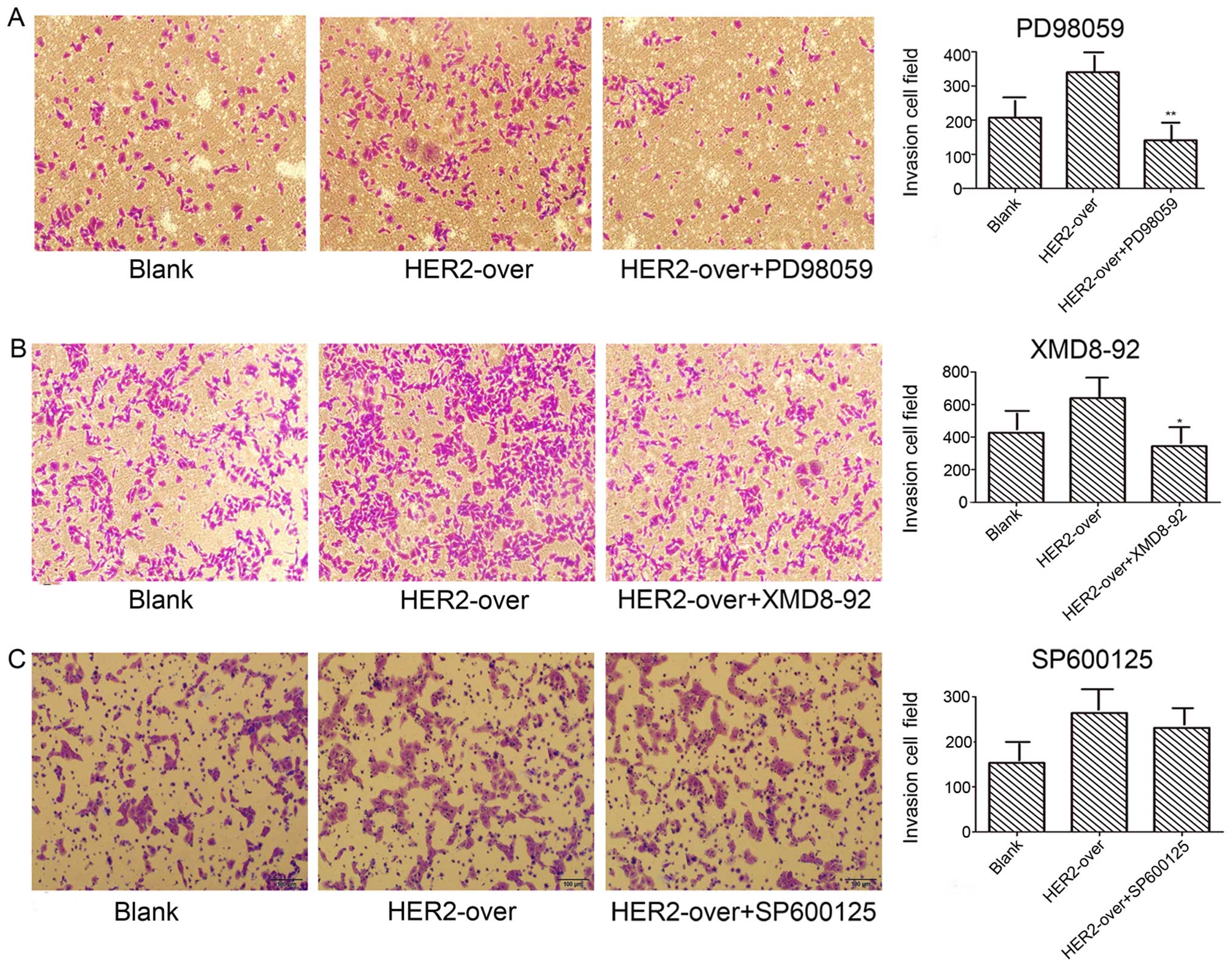
Transwell invasion assays were employed to detect
the invasion abilities mediated by PD98059, SP600125, XMD8-92
inhibitions, respectively. Results as shown in Fig. 6A demonstrate the invasion ability of
PD98059 (ERK1/2 inhibition) group used with concentration of 40
µg/l significantly decreased 58.5%, compared with the
HER-2-over group (P<0.05). In addition, XMD8-92 (ERK5 inhibitor)
group with concentration of 10 µg/l decreased 46.1% when
compared with the HER-2-over group (Fig. 6B; P<0.05). However, no difference
was observed in the invasion ability of SP600125 group contrast to
the HER-2-over group (Fig. 6C).
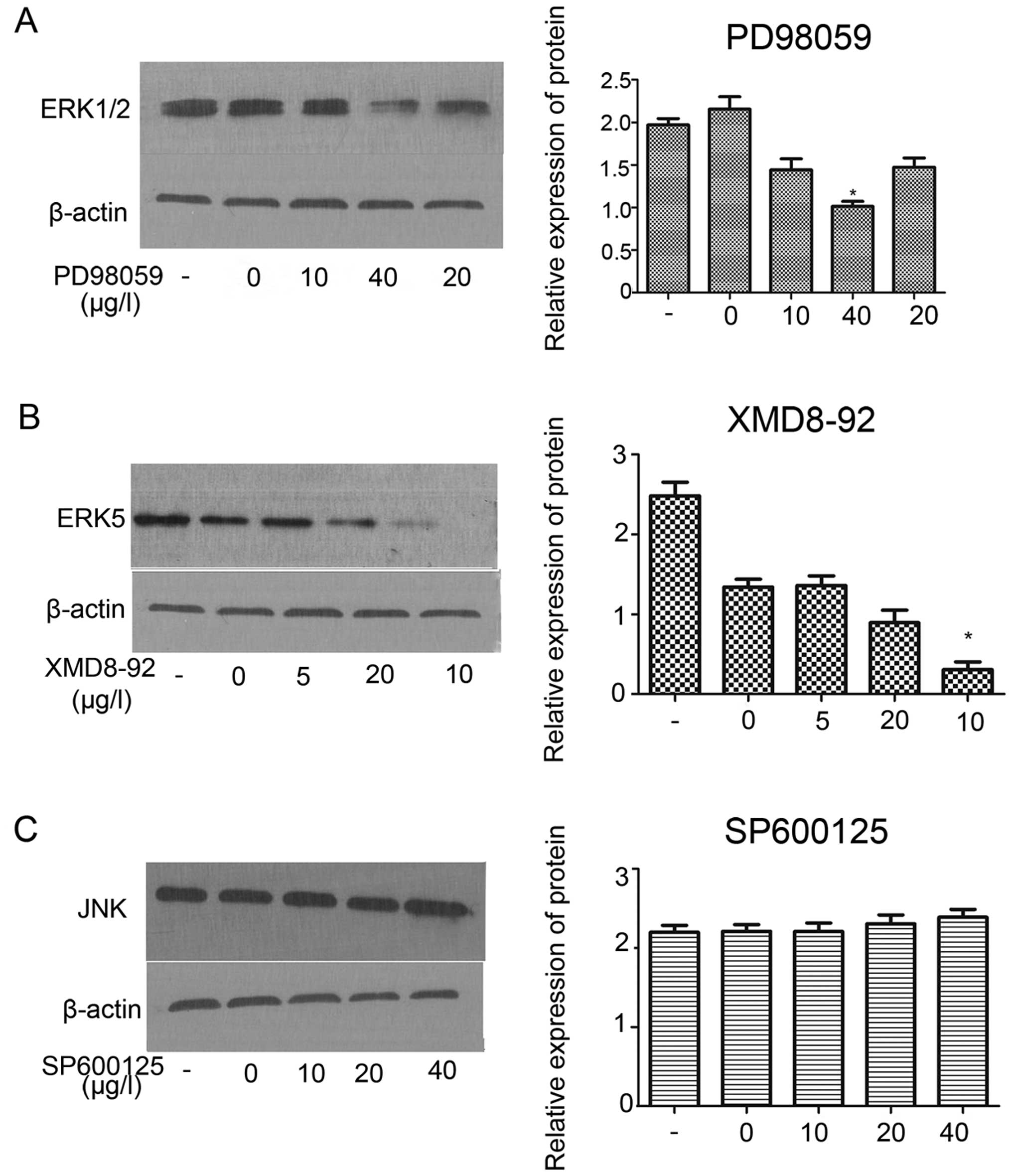
MAPK inhibitors (PD98059, XMD8-92,
SP600125) inhibited protein expression of ERK1/2 and ERK5 in
HER-2-overexpression IOMM-Lee meningioma cells
In order to investigate the effect of PD98059,
XMD8-92, SP600125 on the protein expression of MAPK (ERK1/2)
signaling pathway. The result revealed that, PD98059 (ERK1/2
inhibition) inhibited the protein expression of ERK1/2 at the best
inhibition concentration of 40 µg/l (Fig. 7A; P<0.05). Western blot assay
demonstrated XMD8-92 (ERK5 inhibition) significantly inhibited the
protein expression of ERK5 the component of ERK1/2 signaling at the
best inhibition concentration of 10 µg/l (Fig. 7B; P<0.01). However, no effect was
observed on the JNK at different inhibition concentration of
SP600125 (Fig. 7C).
Discussion
HER-2 is a type of oncogene in human carcinoma, and
many studies have indicated HER-2 overexpression in several types
of cancer and is associated with a particularly aggressive form of
the disease (15,16). Our previous study demonstrated that
overexpression of HER-2 and its mediated PI3K/AKT signaling pathway
in patients with meningioma simulate human meningioma cell
proliferation and invasion, which may contribute to its poor
prognosis and development (13,17).
In the present study, we constructed stable
transformants cell lines (HER-2-overexpression and HER-2-sh) to
investigate the expression level of HER-2 in human malignant
meningioma cells, then examined with fluorescence microscopy,
reverse transcription-quantitative polymerase chain reaction
(q-PCR), Immunofluorescence (IF) staining and western blot
analysis. The results reveal that the HER-2 mRNA and protein levels
of HER-2-sh group cell significantly decreased over 50%, while the
HER-2-over group increased over 2-fold higher than of the blank
group. In our previous study we demonstrated when the gene
expression of HER-2 was downregulated, the proliferative ability of
cells declined in vitro, determined using MTT assay
(13).
Thus, in the present study we further investigated
the proliferative ability of cells in vivo, determined using
animal models. The result was consistent with the experiment in
vitro that HER-2 gene downregulation declined the proliferative
ability of cells. Next we analyzed the effect of PD98059, XMD8-92,
SP600125 on the proliferation, metastasis and MAKP(ERK) signal
pathway relevant protein expression in HER-2-overexpression in
human malignant meningioma cells, determined using MTT assay,
Transwell invasion assay and western blotting. Results showed that
increased PD98059 inhibition concentration inhibited the cell
proliferation and invasion of HER-2-overexpression meningioma
cells, the effect of XMD8-92 on the inhibition of cell
proliferation ability of HER-2-overexpression meningioma cells
compared with PD98059 was more potent and the inhibition effect of
cell invasion was observed. However, no effect was observed in the
cell proliferation and invasion of HER-2-overexpression meningioma
cells of SP600125. In terms of western blotting, our results showed
that PD98059 and XMD8-92 decreased the protein expression of ERK1/2
and ERK5, whereas SP600125 had no effect on the JNK. Therefore, the
present study demonstrated that HER-2 promoted cell proliferation
and invasion in the human malignant meningioma IOMM-Lee cells and
provided some evidences for a functional linkage between HER-2
signaling and the activity of MAPK (ERK) in cell proliferation and
invasion.
According to a previous study, HER-2 plays a role by
homo- or heterodimerization with an extracellular domain (ECD) of
other ErbB family members, which close proximity of the receptors
leads to phosphorylation of the C-terminal tyrosines. Some
phosphorylation sites are the tyrosines residues on the receptor
molecule serving as recognition and docking sites for
SH2-containing protein which consist of the components to activate
the RAS/MAPK pathway and PI3K/AKT pathway (4,18,19).
The generic MAPK signaling pathway is shared by at
least four distinct cascades, which are named according to their
MAPK tier component: the extracellular signal-related kinase
(ERK1/2), Jun amino-terminal kinases (JNK1/2/3), p38-MAPK and ERK5.
MAPK pathway is an essential pathway in the cell proliferation,
differentiation, migration, senescence and apoptosis (20). Therefore, based on the present study
we speculate that HER-2 can affect the protein synthesis or
activities of MAPK pathway, promote the cell proliferation and
invasion. To assess this hypothesis, the present study used western
blot analysis to determine the expression levels of MAPK pathway.
Upregulation of the expression of HER-2 lead to increased levels of
ERK1/2, ERK5 and JNK. ERK1/2 is pivotal in further signaling of the
pathway, as it is reported that the GRB2 interacts with the guanine
nucleotide exchange factor, SOS. SOS can then cause the exchange of
guanosine diphosphate (GDP) to guanosine triphosphate (GTP) on
RAS.
Activated RAS then initiates the activation of a
kinase cascade culminating in the phosphorylation and activation of
extracellular signal-regulated kinases 1 and 2 (ERK1, ERK2), where
the ERK phosphorylates and activates various transcription factors,
regulating various cellular processes, including proliferation,
migration and differentiation (21,22).
Thus, the addition of PD98059 (ERK1/2) in the HER-2-overexpression
meningioma cells suppressed cell proliferation and invasion. Next,
ERK 5 is the effector kinase of a canonical three-tiered MAPK
signalling cascade comprising MEK (MAPK/ERK kinase) 5, MEKK (MEK
kinase) 2/3 and ERK5 itself. The 444-amino-acid ERK5 protein,
contains an N-terminal kinase domain with 40% homology to the ERK2,
and a large C-terminal extension that contains a transactivation
domain.
ERK5 is activated via the dual phosphorylation of
its Serine 311 and Threonine 315 by MEKK2, 3/Tpl2, phosphorylates
substrates including MEF2, c-Fos, Fra 1, Sap-1, c-Myc and NF-κB
that regulate the cell proliferation, survival, motility and
angiogenesis. ERK5 and ERK1/2 present the cooperative effect in
regulating some kinases (23–25).
In other words, ERK5 and ERK1/2 signaling pathway may cross-talk on
the regulating tumor cell proliferation and motility. As the
present study results showed, inhibition of ERK5 also contributes
to the impact on the cell proliferation and invasion abilities.
Another component of the MAPK, the c-Jun NH2-terminal kinase (JNKs)
is encoded by 2 ubiquitously expressed genes (Jnk1 and Jnk2) and by
a third gene (Jnk3) can phosphorylate the c-Jun transcription
factor at serine (Ser) 63 and -73, resulting in the robust
induction of c-Jun trans-activation. Previous studies demonstrated
JNK is not required for proliferation and cell motility of mammary
epithelial cells, but contributed to cell apoptosis. In addition,
in individual tumor types, JNK may have an affect on tumor
development or may contribute (positively or negatively) to tumor
pathology. Further, owing to the poor selectivity of SP600125 for
JNK, it is therefore unclear whether JNK inhibition mediates the
effects of SP600125 on the proliferation and invasion of IOMM-Lee
meningioma cells (26,27). There are also other signal pathway,
which infect the components of MAPK signal pathway (14). Thus, in the present study, ERK1/2,
ERK5 and JNK may not be changed completely. We should provide
insight into the exact molecular mechanisms in further study.
In summary, the present results suggested that
over-expression of HER-2 promoted human meningioma cell
proliferation and invasion in vivo and in vitro which
may affect the meningioma development and progression. These
results may explain, in part, the increased HER-2 in human
meningioma cells which is clinically associated with the high
recurrence potential and poor prognosis. Furthermore, the present
study recorded the correlation between HER-2 signaling and the
activity of MAPK (ERK) in cell proliferation and invasion. These
data indicated that the HER-2-RAS-MAPK pathway to a certain extent
may be available for further clinical development of human
malignant meningioma for novel therapeutic approaches.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (grant no. 81260372) and the
Science and Technology of Jiangxi Province (grant no.
20151BBG70217).
References
|
1
|
Louis DN, Ohgaki H, Wiestler OD, Cavenee
WK, Burger PC, Jouvet A, Scheithauer BW and Kleihues P: WHO
classification of tumours of the central nervous system. Acta
Neuropathol. 114:97–109. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shibuya M: Pathology and molecular
genetics of meningioma: Recent advances. Neurol Med Chir (Tokyo).
55:14–27. 2015. View Article : Google Scholar
|
|
3
|
Jennifer M, Cope WP, Vartanian ED, Reiner
AS, Kellen R, Ogilvie SQ, Huse JT and Gutin PH: Survival in
patients treated for anaplastic meningioma. J Neurosurg. 123:23–30.
2015. View Article : Google Scholar
|
|
4
|
Cain SA, Smoll NR, Van Heerden J, Tsui A
and Drummond KJ: Atypical and malignant meningiomas: Considerations
for treatment and efficacy of radiotherapy. J Clin Neurosci.
22:1742–1748. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Dittrich A, Gautrey H, Browell D and
Tyson-Capper A: The HER2 signaling network in breast cancer - Like
a spider in its web. J Mammary Gland Biol Neoplasia. 19:253–270.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tai W, Mahato R and Cheng K: The role of
HER2 in cancer therapy and targeted drug delivery. J Control
Release. 146:264–275. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ménard S, Casalini P, Campiglio M, Pupa SM
and Tagliabue E: Role of HER2/neu in tumor progression and therapy.
Cell Mol Life Sci. 61:2965–2978. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Vernimmen D, Gueders M, Pisvin S, Delvenne
P and Winkler R: Different mechanisms are implicated in ERBB2 gene
over-expression in breast and in other cancers. Br J Cancer.
89:899–906. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mahzouni P and Movahedipour M: An
immunohistochemical study of HER2 expression in meningioma and its
correlation with tumor grade. Pathol Res Pract. 208:221–224. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Schwechheimer K, Läufle RM, Schmahl W,
Knödlseder M, Fischer H and Höfler H: Expression of neu/c-erbB-2 in
human brain tumors. Hum Pathol. 25:772–780. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Schlegel J, Ullrich B, Stumm G, Gass P,
Harwerth IM, Hynes NE and Kiessling M: Expression of the
c-erbB-2-encoded oncoprotein and progesterone receptor in human
meningiomas. Acta Neuropathol. 86:473–479. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chozick BS, Benzil DL, Stopa EG, Pezzullo
JC, Knuckey NW, Epstein MH, Finkelstein SD and Finch PW:
Immunohistochemical evaluation of erbB-2 and p53 protein expression
in benign and atypical human meningiomas. J Neurooncol. 27:117–126.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wang W, Tu Y, Wang S, Xu S, Xu L, Xiong Y,
Mei J and Wang C: Role of HER-2 activity in the regulation of
malignant meningioma cell proliferation and motility. Mol Med Rep.
12:3575–3582. 2015.PubMed/NCBI
|
|
14
|
De Luca A, Maiello MR, D'Alessio A,
Pergameno M and Normanno N: The RAS/RAF/MEK/ERK and the PI3K/AKT
signalling pathways: role in cancer pathogenesis and implications
for therapeutic approaches. Expert Opin Ther Targets. 16(Suppl 2):
S17–S27. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Richman SD, Southward K, Chambers P, Cross
D, Barrett J, Hemmings G, Taylor M, Wood H, Hutchins G, Foster JM,
et al: HER2 overexpression and amplification as a potential
therapeutic target in colorectal cancer: Analysis of 3256 patients
enrolled in the QUASAR, FOCUS and PICCOLO colorectal cancer trials.
J Pathol. 238:562–570. 2016. View Article : Google Scholar :
|
|
16
|
Ren W, Liu Y, Wan S, Fei C, Wang W, Chen
Y, Zhang Z, Wang T, Wang J, Zhou L, et al: BMP9 inhibits
proliferation and metastasis of HER2-positive SK-BR-3 breast cancer
cells through ERK1/2 and PI3K/AKT pathways. PloS One. 9:e968162014.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wang CL, Mei JH, Wang SS, Xu S, Xu LL and
Xiong YF: Expression of HER2/neu in meningiomas: An
immunohistochemistry and fluorescence in situ hybridization study.
Zhonghua Bing Li Xue Za Zhi. 39:156–160. 2010.In Chinese.
PubMed/NCBI
|
|
18
|
Yang L, Li Y and Zhang Y: Identification
of prolidase as a high affinity ligand of the ErbB2 receptor and
its regulation of ErbB2 signaling and cell growth. Cell Death Dis.
5:e12112014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Elster N, Collins DM, Toomey S, Crown J,
Eustace AJ and Hennessy BT: HER2-family signalling mechanisms,
clinical implications and targeting in breast cancer. Breast Cancer
Res Treat. 149:5–15. 2015. View Article : Google Scholar
|
|
20
|
Sun Y, Liu WZ, Liu T, Feng X, Yang N and
Zhou HF: Signaling pathway of MAPK/ERK in cell proliferation,
differentiation, migration, senescence and apoptosis. J Recept
Signal Transduct Res. 35:600–604. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Pearson G, Robinson F, Beers Gibson T, Xu
BE, Karandikar M, Berman K and Cobb MH: Mitogen-activated protein
(MAP) kinase pathways: Regulation and physiological functions.
Endocr Rev. 22:153–183. 2001.PubMed/NCBI
|
|
22
|
Rubinfeld H and Seger R: The ERK cascade:
A prototype of MAPK signaling. Mol Biotechnol. 31:151–174. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lochhead PA, Gilley R and Cook SJ: ERK5
and its role in tumour development. Biochem Soc Trans. 40:251–256.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hu B, Ren D, Su D, Lin H, Xian Z, Wan X,
Zhang J, Fu X, Jiang L, Diao D, et al: Expression of the
phosphorylated MEK5 protein is associated with TNM staging of
colorectal cancer. BMC Cancer. 12:1272012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang X and Tournier C: Regulation of
cellular functions by the ERK5 signalling pathway. Cell Signal.
18:753–760. 2006. View Article : Google Scholar
|
|
26
|
Cellurale C, Girnius N, Jiang F,
Cavanagh-Kyros J, Lu S, Garlick DS, Mercurio AM and Davis RJ: Role
of JNK in mammary gland development and breast cancer. Cancer Res.
72:472–481. 2012. View Article : Google Scholar :
|
|
27
|
Li W, Wen C, Bai H, Wang X, Zhang X, Huang
L, Yang X, Iwamoto A and Liu H: JNK signaling pathway is involved
in piperlongumine-mediated apoptosis in human colorectal cancer
HCT116 cells. Oncol Lett. 10:709–715. 2015.PubMed/NCBI
|