Introduction
Lung cancer is the leading cause of cancer-related
deaths worldwide, and non-small cell lung cancer (NSCLC) accounts
for almost 80% of all lung cancers. Cisplatin (CDDP) is one of the
most important cytotoxic agents for the treatment of advanced
NSCLC. However, cancer cells often develop multiple mechanisms to
overcome CDDP-induced DNA damage and apoptosis, leading to CDDP
resistance (1), which is a major
limitation to its clinical effectiveness. Tumor-associated
angiogenesis after the development of acquired platinum resistance
is important for tumor growth and metastasis (2,3).
Angiotensin II (AngII), a biologically active
octapeptide in the renin-angiotensin system (RAS), mediates its
biological effect by binding to two subtypes of receptors, type 1
(AT1R) and type 2 (AT2R), that belong to the G-protein-coupled
receptor superfamily (4).
Angiotensin-converting enzyme 2 (ACE2) is a newly identified
component of RAS, which converts AngII to angiotensin-(1–7)
[Ang-(1–7)] (5), a
peptide with vasodilator and antiproliferative properties. The
tumor environment contains all RAS components necessary to produce
angiotensin locally, in addition to systemically produced
angiotensin. Previously, numerous prospective and retrospective
studies have reported that cancer patients who received
angiotensin-converting enzyme inhibitors (ACEIs) or angiotensin II
type 1 receptor blockers (ARBs) had a longer survival than
non-recipients (6–9). Studies have also shown that ACEIs and
ARBs reduced cancer growth and angiogenesis in vitro and
in vivo (10). Emerging data
suggest that platinum-resistant cancer cells show significantly
higher vascular endothelial growth factor (VEGF) expression than
their corresponding parental cells in vitro and in
vivo (2) and that acquired
platinum resistance enhanced tumor angiogenesis through AT1R in
bladder cancer (3). Our previous
studies demonstrated that ACE2 overexpression reduced
tumor-associated angiogenesis and tumor cell invasion in
vitro and in vivo (11,12).
However, the role of ACE2 in tumor associated angiogenesis induced
by acquired platinum resistance is still unknown.
Based on previous studies, we hypothesized that ACE2
plays an important role in tumor-associated angiogenesis induced by
acquired platinum resistance. In the present study, we investigated
the changes in VEGF and RAS component expression between
platinum-resistant NSCLC cells and their corresponding parent cells
in vitro and in vivo. The present study also examined
the effect and signaling roles of ACE2 overexpression in acquired
platinum resistance-induced tumor angiogenesis in NSCLC.
Materials and methods
Cell lines and culture
A549 lung cancer cells (Shanghai Institute of Cells)
were maintained in Ham's F12 medium with 10% fetal bovine serum
(FBS), 100 mg/ml penicillin and 100 U/ml streptomycin using media
and growth reagents from Gibco-BRL (Grand Island, NY). Lewis lung
carcinoma cells (LLC) (Shanghai Institute of Cells) were maintained
in RPMI-1640 medium. To develop platinum resistance, A549 and LLC
cells were supplemented with 10% FBS containing CDDP, at 37°C in a
humidified 5% CO2 atmosphere. Then, the concentration of
CDDP was increased to 3 µM. These cells were passaged upon
reaching confluency over a 6-month period. These new cell lines
were named A549-DDP and LLC-DDP (acquired platinum resistance for 6
months).
Retroviral vector construct and
transduction
The pcDNA3.1 vector containing human ACE2 cDNA was
kindly provided by Dr Paul McCray (University of Iowa, Iowa City,
IA, USA). The vectors were ligated into the
BglII/XhoI site in the pMSCV plasmid, which expresses
human ACE2. 293T cells were transfected with DNA (4 µg
pMD-gag-pol, 4 µg pMD-VSVG and 4 µg retroviral vector
pMSCV-ACE2 or pMSCV) using the Lipofectamine 2000 reagent
(Invitrogen Corp., Carlsbad, CA, USA). The replication retrovirus
was collected after 48 h and used to infect A549-DDP cells using
Polybrene (Sigma Corp., Cream Ridge, NJ, USA) followed by
centrifugation. Infected cells were selected with 4 µg/ml
puromycin for 2 weeks. The resistant clones were isolated by
limiting dilution and dispatched to new dishes. The obtained cells
were subsequently named vector and A549-DDP-ACE2 cells.
Tumor growth assays
Six-to eight-week-old BALB/c athymic nude mice
(Chinese Academy of Sciences, Shanghai, China) were used. Animal
experiments were performed in accordance with the institutional
guidelines of the University Committee on the Use and Care of
Animals. To investigate CDDP sensitivity, the mice were
administered CDDP (10 mg/kg) intraperitoneally on day 21 after
cancer cell implantation. The mice were divided into the following
5 groups: i) A549, ii) A549-CDDP, iii) A549-DDP, iv) A549-DDP-CDDP
and v) A549-DDP-ACE2. Each mouse was inoculated in the flank with
1×106 A549 cells (n=6), A549-DDP cells infected with
MSCV (n=6) or A549-DDP cells infected with ACE2 (n=6). Tumor growth
was monitored at 3-day intervals by measuring the tumor diameters
using a vernier caliper. Tumor volume was determined based on the
following formula: Tumor volume = [(major axis) × (minor
axis)2] × 1/2. Following a 30-day follow-up period, the
mice were euthanized and the tumors were removed. Harvested tumors
were cut into 2 pieces, one of which was placed in liquid nitrogen
and frozen at −80°C and the other was fixed using 4%
paraformaldehyde and then embedded in paraffin.
Real-time quantitative reverse
transcription analysis
ACE, ACE2, AT1R and VEGF mRNA expression in A549,
A549-DDP, LLC, LLC-DDP and A549-DDP-ACE2 cells were examined by
real-time quantitative reverse transcription-based polymerase chain
reaction (qRT-PCR) Total RNAs were extracted from the cells with
TRIzol reagent. RNA was treated with DNase (Promega, Madison, WI,
USA) and complementary DNA was synthesized using a cDNA synthesis
kit (Applied Biosystems, Foster City, CA, USA) according to the
manufacturer's instructions. Fluorescence qRT-PCR was performed
using the double-stranded DNA dye SYBR-Green (PCR Core Reagents, PE
Biosystems, Warrington, UK) using an ABI PRISM 7300 system (Applied
Biosystems). The SYBR-Green assay contained 1 µl 10X
SYBR-Green PCR buffer, 0.8 µl deoxynucleoside triphosphate
(dNTP) mixture, 0.1 µl AmpErase UNG (1 U/µl), 0.05
µl AmpliTaq Gold DNA Polymerase (5 U/µl), 1.2
µl MgCl2 (25 mM), 0.1 µl forward and
reverse primer (20 µM), 1 µl cDNA and 5.65 µl
double distilled H2O. The following PCR program was used
to determine gene expression: i) 50°C for 2 min, ii) 95°C for 10
min and iii) 45 cycles of 95°C for 30 sec for denaturation, 60°C
for 30 sec for annealing and 72°C for 30 sec for extension. All of
the data were analyzed using the ABI PRISM SDS 2.0 software
(Perkin-Elmer, Wellesley, MA, USA). GAPDH was co-amplified to
normalize for the RNA amount added to the reactions and the data
were subjected to cycling threshold analysis using the ΔCt method.
PCR was repeated at least three times. The following primers were
used in the present study: i) ACE forward, 5′-CCGATCTGGCAGAACTTC-3′
and reverse, 5′-GTGTTCCAGATCGTCCTC-3′ and ii) ACE2 forward,
5′-CCACTGCTCAACTACTTTGAGCC-3′ and reverse,
5′-CTTATCCTCACTTTGATGCT.
Western blot analysis
ACE, ACE2, AT1R and VEGF expression levels were
measured in the A549, A549-DDP, LLC, LLC-DDP and A549-DDP-ACE2
cells. After being treated with protein assay reagent (Bio-Rad
Laboratories, Hercules, CA, USA), soluble protein was separated on
10% SDS-polyacrylamide gels and transferred to polyvinylidene
fluoride (PVDF) membranes (Millipore, Billerica, MA, USA). ACE2
(AF933; R&D Systems, Minneapolis, MN, USA), AT1R (1:1,000),
VEGF (1:1,000) (both from Santa Cruz Biotechnology, Santa Cruz, CA,
USA) and β-actin monoclonal antibodies (1:10,000, Sigma
Corporation, Frederick, MD, USA) were used to detect ACE2, AT1R,
VEGF and β-actin proteins, respectively. The immunoreactive bands
were visualized by ECL Plus reagent (Amersham Biosciences,
Piscataway, NJ, USA). VEGF expression protein in the A549 cells was
analyzed by Scion Image (Scion Corp., Frederick, MD, USA).
In vitro tube formation assay
Cellular angiogenesis was measured in vitro
based on tube formation on an extracellular Matrigel in a 24-well
plate as previously described (13). Briefly, 24-well plates were coated
with 40 ml cold liquid Matrigel (8 mg/ml)/well and incubated at
37°C for 30 min to promote solidification. A549 or A549-DDP or
A549-DDP-ACE2 cells were grown for 48 h in FCS-free media prior to
incubation with different fatty acids in the presence of 5% fetal
calf serum (FCS) media. Cells were seeded at a density of 70,000
cells/well. After a 16 h incubation, the wells were photographed
with an inverted microscope at a magnification of ×4 (Nikon TS100F;
Nikon Corporation, Tokyo, Japan). Images were captured from the
central view of at least five different fields/well and the extreme
edges were excluded due to gel meniscus formation. Adobe Photoshop
(version CS4) was used to quantify the tubule length from capillary
network formation (14). The length
of each tubule was determined by drawing a line over each tubule
and the mean length of the lines (pixels) drawn in each image was
calculated. Quantification of the tubular networks was measured by
counting the total length of tubes in pixels. The results are
expressed as a % over the control using the following formula: %
over the control = the mean length of total tubes × 100/mean length
of the tubes. The number of capillary connections or branch points
between cells was counted manually. Independent experiments were
performed in triplicate to reduce intra-assay variability. The
average number and length of the connections were calculated for
each fatty acid treatment.
Results
Establishment of the acquired
platinum-resistant NSCLC sublines in vitro
Using A549 and LLC cells, we generated NSCLC
sublines with an acquired resistance to platinum. These cells were
successfully cultured for 6 months in CDDP-conditioned medium and
were named A549-DDP and LLC-DDP. Following 3 months without
CDDP-exposure, further examinations were performed. A549, LLC,
A549-DDP and LLC-DDP cells were treated with various CDDP
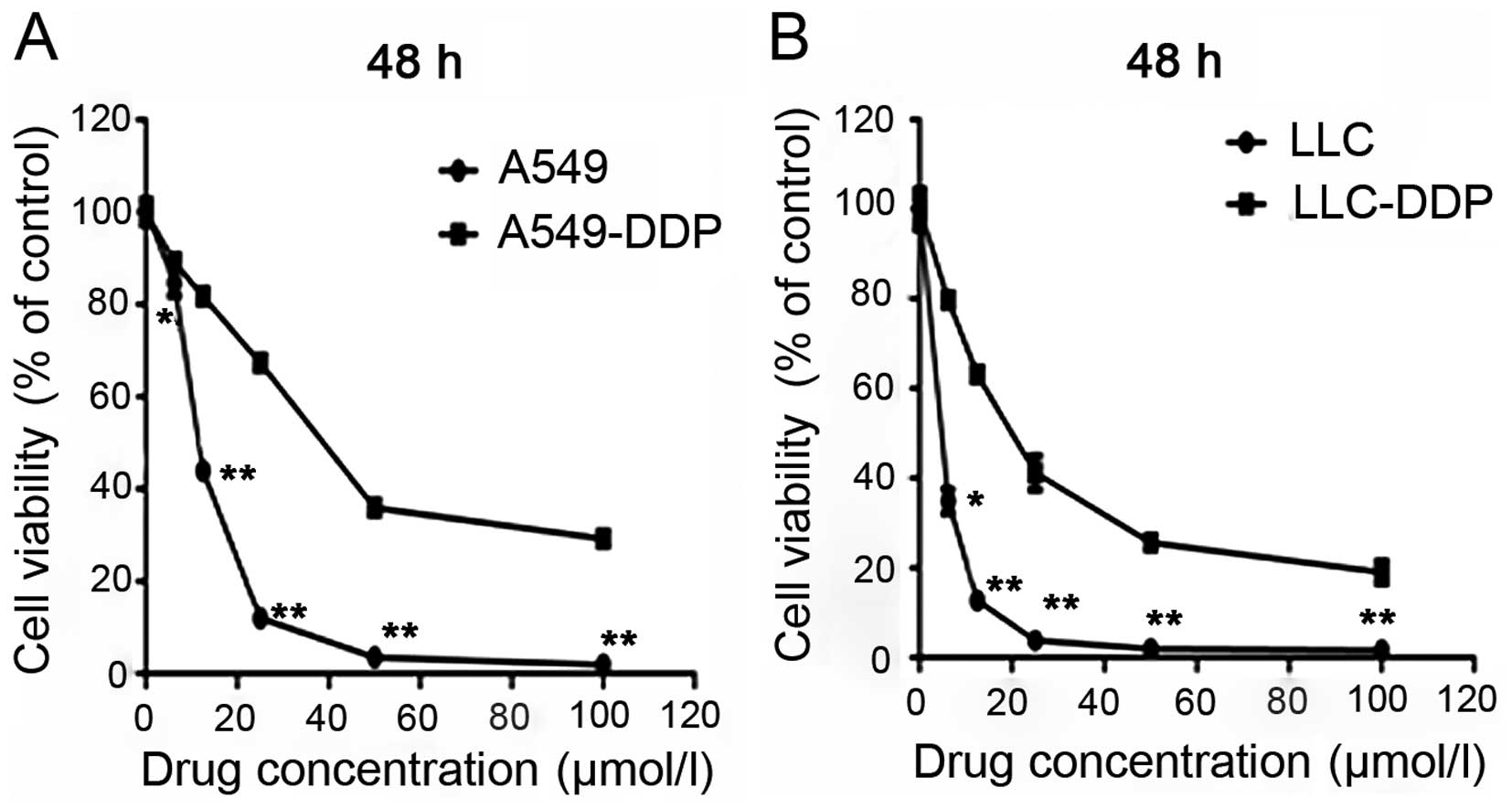
concentrations for 48 h (Fig. 1A and
B). The IC50 values for A549 and A549-DDP cells were
10.8±0.5 and 39.6±1.8 µmol/l, respectively. The resistance
factor (RF) was 3.67. The IC50 values for LLC and
LLC-DDP cells were 8.5±0.4 and 23.3±2.1 µmol/l,
respectively. The RF for these cells was 2.74.
Effect of acquired platinum resistance on
VEGF production and RAS components in vitro
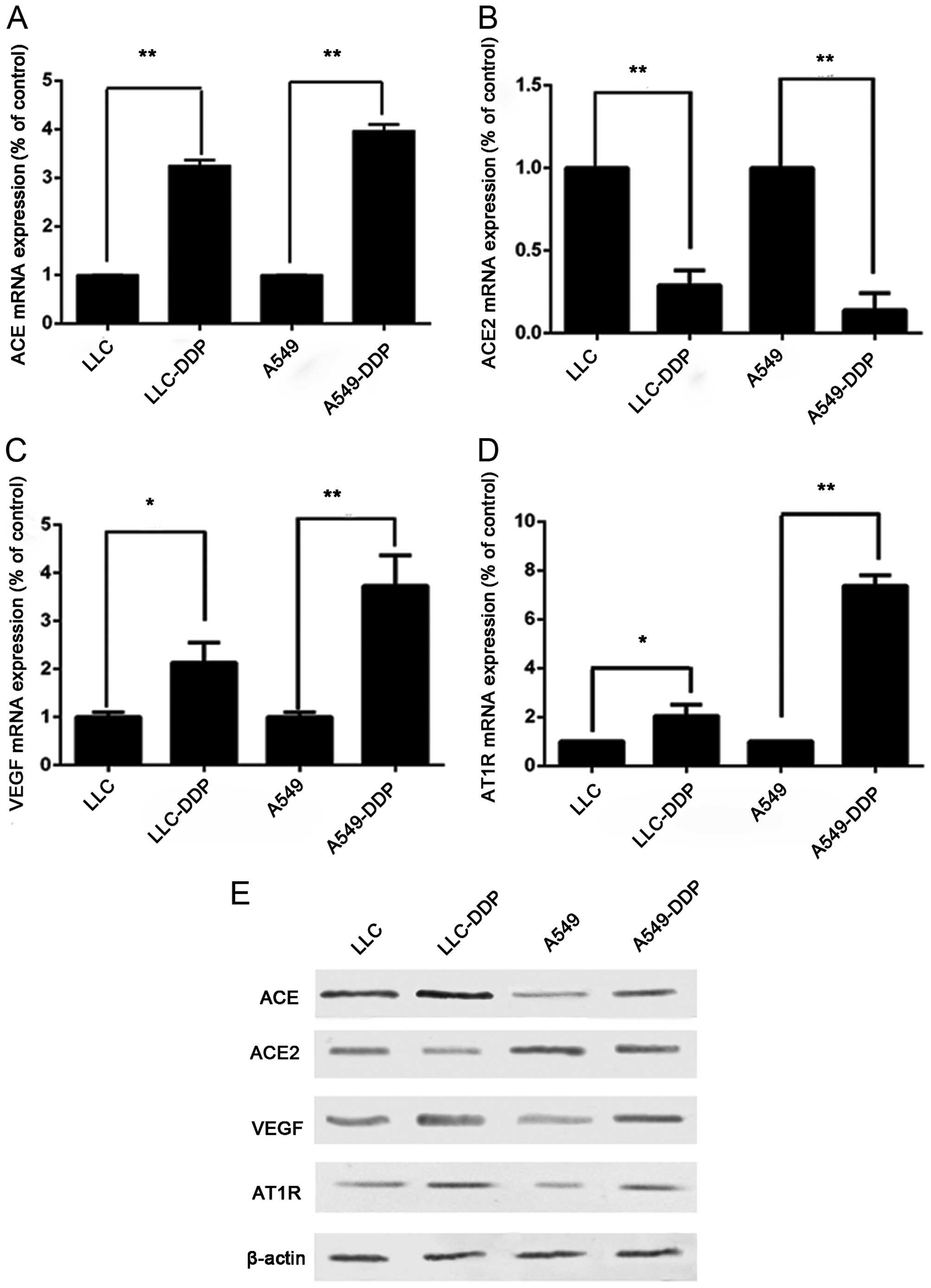
Analysis of the qRT-PCR data for ACE, ACE2, AT1R and
VEGF mRNAs, which were corrected with GAPDH as an internal control,
showed that both A549-DDP and LLC-DDP sublines showed significantly
higher ACE, AT1R and VEGF mRNA expression (Fig. 2A, C and D) and lower ACE2 mRNA
(Fig. 2B) expression than their
corresponding parent cells. Using western blot analysis, both the
A549-DDP and LLC-DDP sublines showed significantly higher ACE, AT1R
and VEGF expression and lower ACE2 (Fig. 2E) expression than their
corresponding parent cells.
Effect of ACE2 gene transfection on
A549-DDP cells
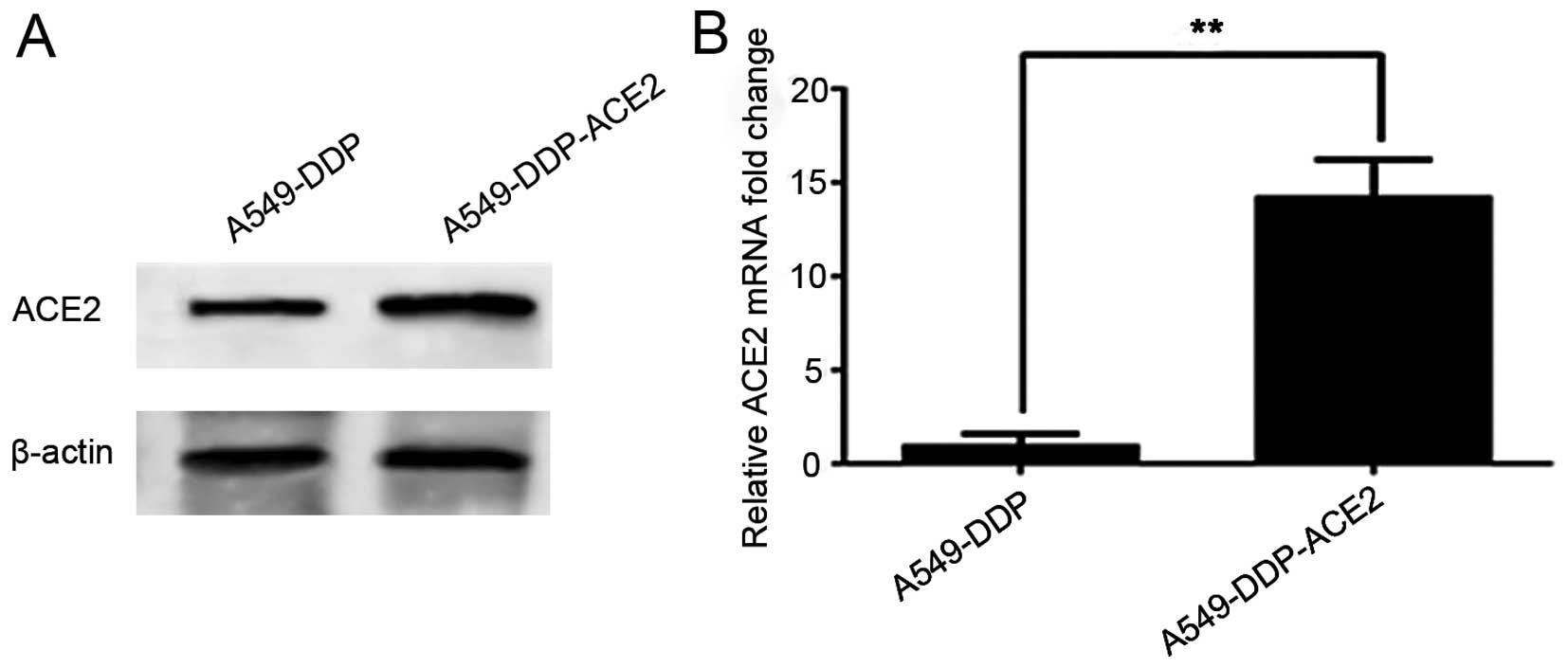
Our objective was to determine the efficacy of the
MSCV-ACE2 transfection. A549-DDP cells were infected with MSCV-ACE2
and selected in the presence of puromycin. Infection of A549-DDP
cells with MSCV-ACE2 resulted in robust ACE2 expression by western
blot analysis at 72 h. Fig. 3A
shows that ACE2 was overexpressed in the A549-DDP-ACE2 group in
comparison with the A549-DDP group. As expected, the expression of
ACE2 mRNA (Fig. 3B) was notably
higher in the A549-DDP-ACE2 group when compared to the A549-DDP
group.
Effect of ACE2 overexpression on VEGF
production and RAS components in vitro
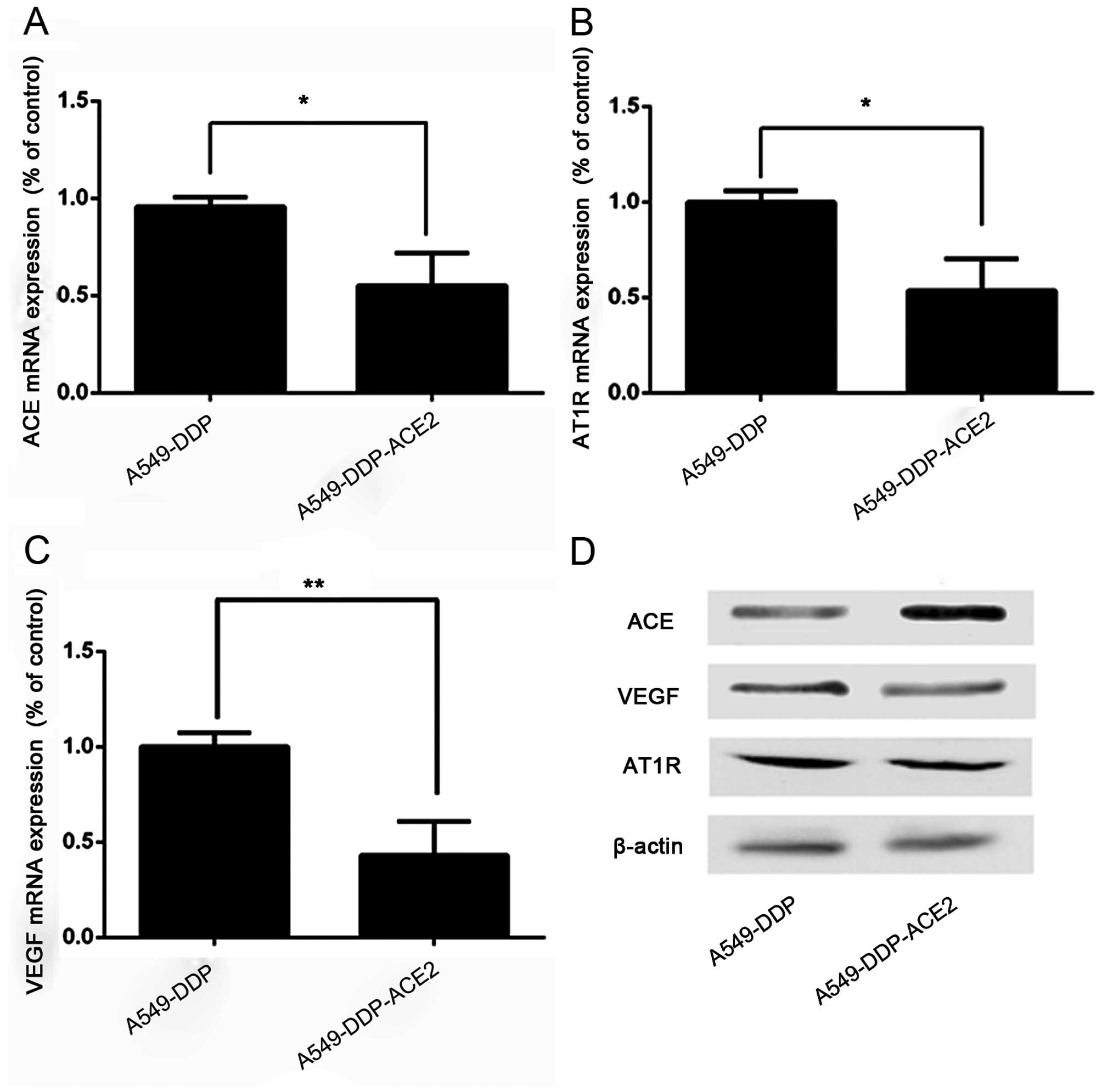
We investigated whether the ACE2 overexpression
inhibited VEGF production in acquired platinum-resistant NSCLC
sublines. ACE, AT1R and VEGF mRNA expression was decreased in the
A549-DDP-ACE2 group when compared to the A549-DDP group (Fig. 4A–C). Western blot analysis also
demonstrated that the ACE, AT1R and VEGF productions was decreased
in the A549-DDP-ACE2 group compared to the A549-DDP group (Fig. 4D).
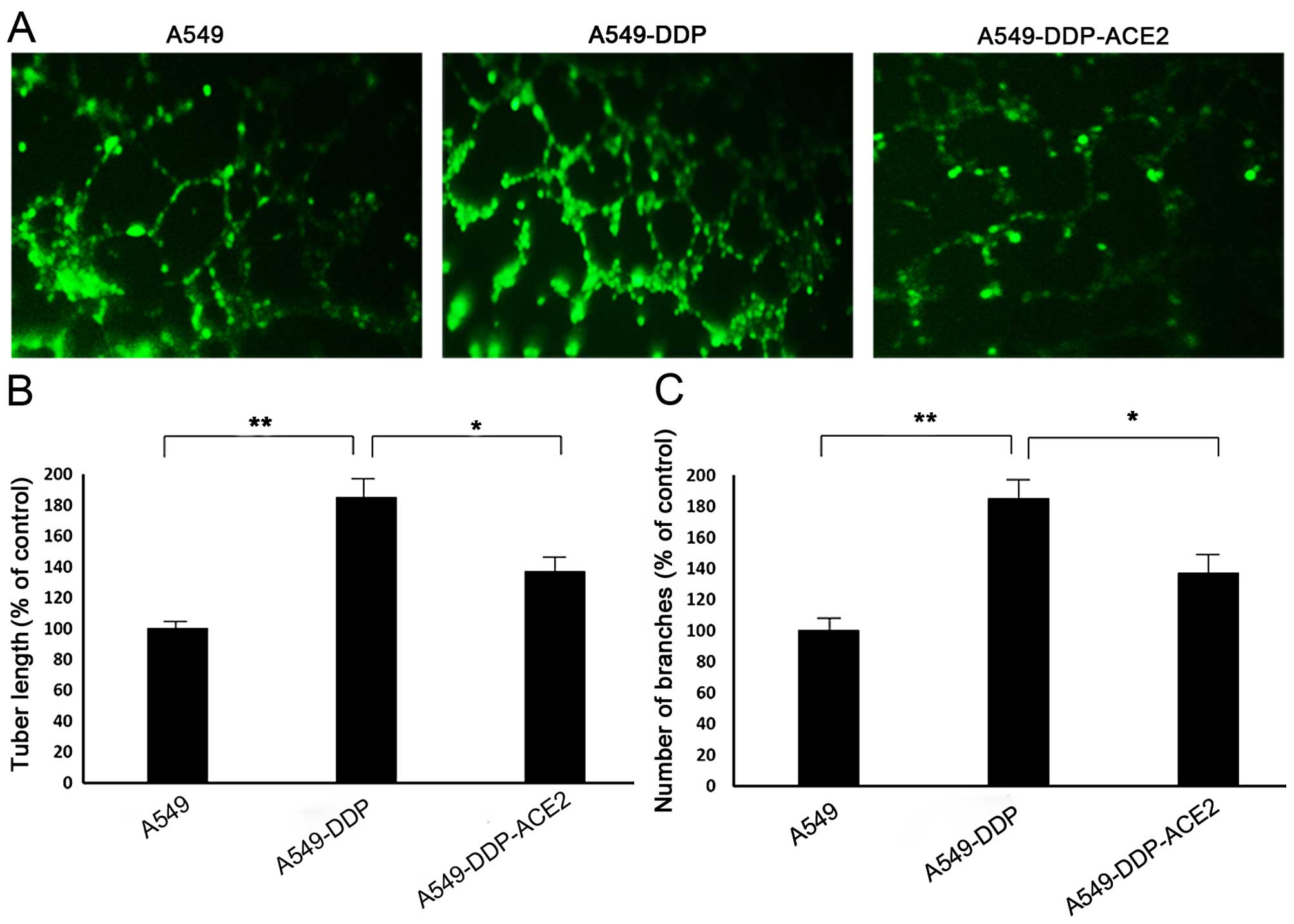
Effect of ACE2 overexpression on tube
formation in acquired platinum-resistant NSCLC sublines
A tube formation assay was performed by Matrigel to
evaluate the angiogenic response of acquired platinum-resistant
NSCLC sublines. A549-DDP cells significantly increased capillary
network complexities and induced more tubes in the network compared
with A549 cells. The A549-DDP-ACE2 cells showed a reduced ability
to form capillary-like structures (i.e. decreased tube length and
numbers of branches) compared to the A549-DDP cells (Fig. 5).
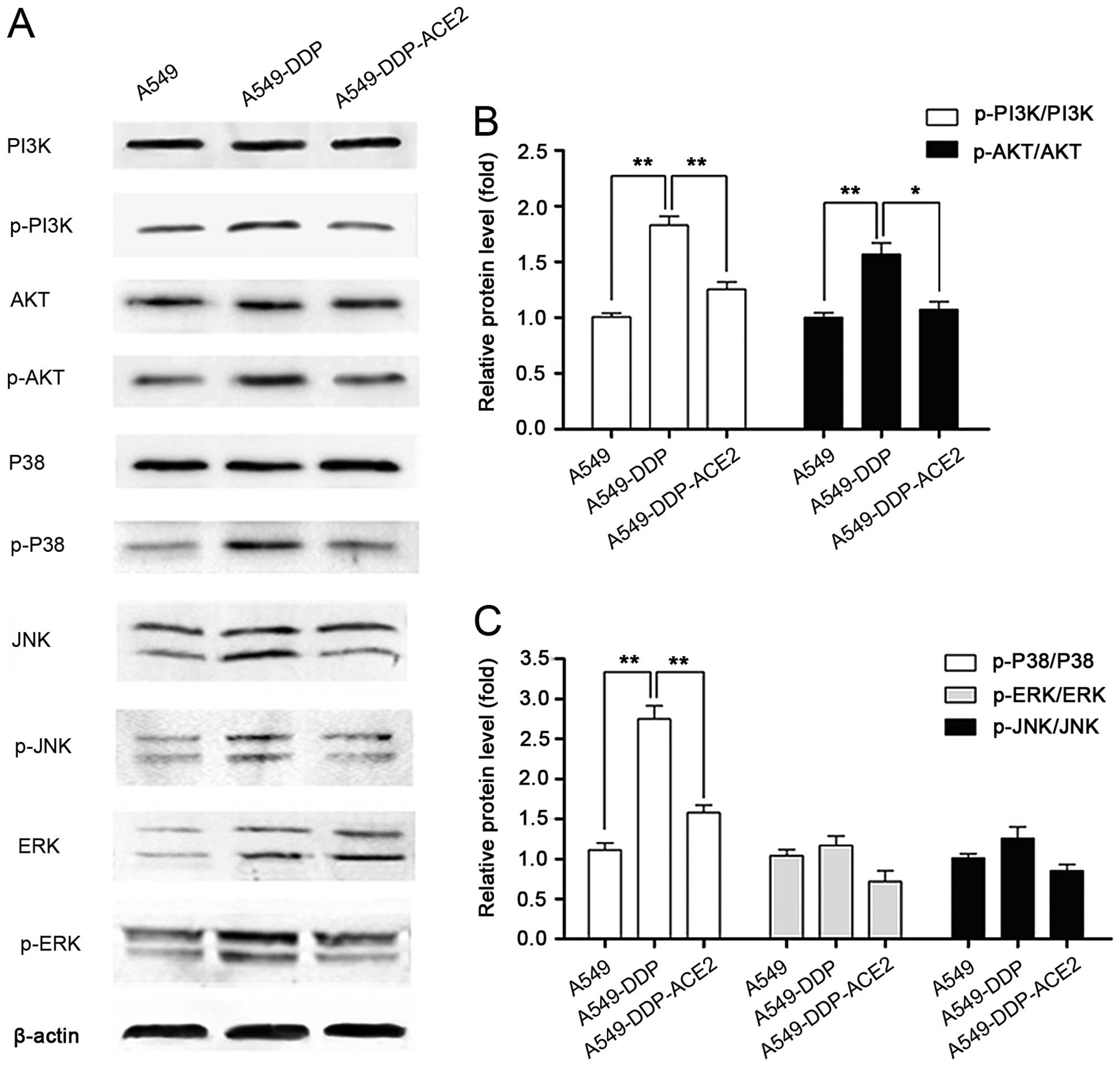
Effect of ACE2 overexpression on the
PI3K/Akt and MAPK signaling pathways in the A549-DDP cells
The PI3K/Akt and MAPK signaling pathways are
involved in the expression of VEGF and angiogenesis. To investigate
the anti-angiogenic mechanism of ACE2 in the acquired
platinum-resistant NSCLC sublines, the effects of ACE2
overexpression on the expression and phosphorylated status of
PI3K/Akt and MAPK signaling proteins in A549-DDP cells were
investigated by western blot analysis (Fig. 6A). The levels of PI3K/Akt and MAPK
signaling proteins were not altered in the A549-DDP cells, while
the phosphorylation levels in PI3K/Akt and p38 MAPK signaling were
significantly increased in A549-DDP cells compared with A549 cells
(Figs. 6B and C). In contrast,
these active effects were blocked by the ACE2 overexpression in
A549-DDP-ACE2 cells. The data revealed that inhibition of VEGF
expression and angiogenesis in acquired platinum-resistant NSCLC
could partially occur through PI3K/Akt and p38 MAPK
inactivation.
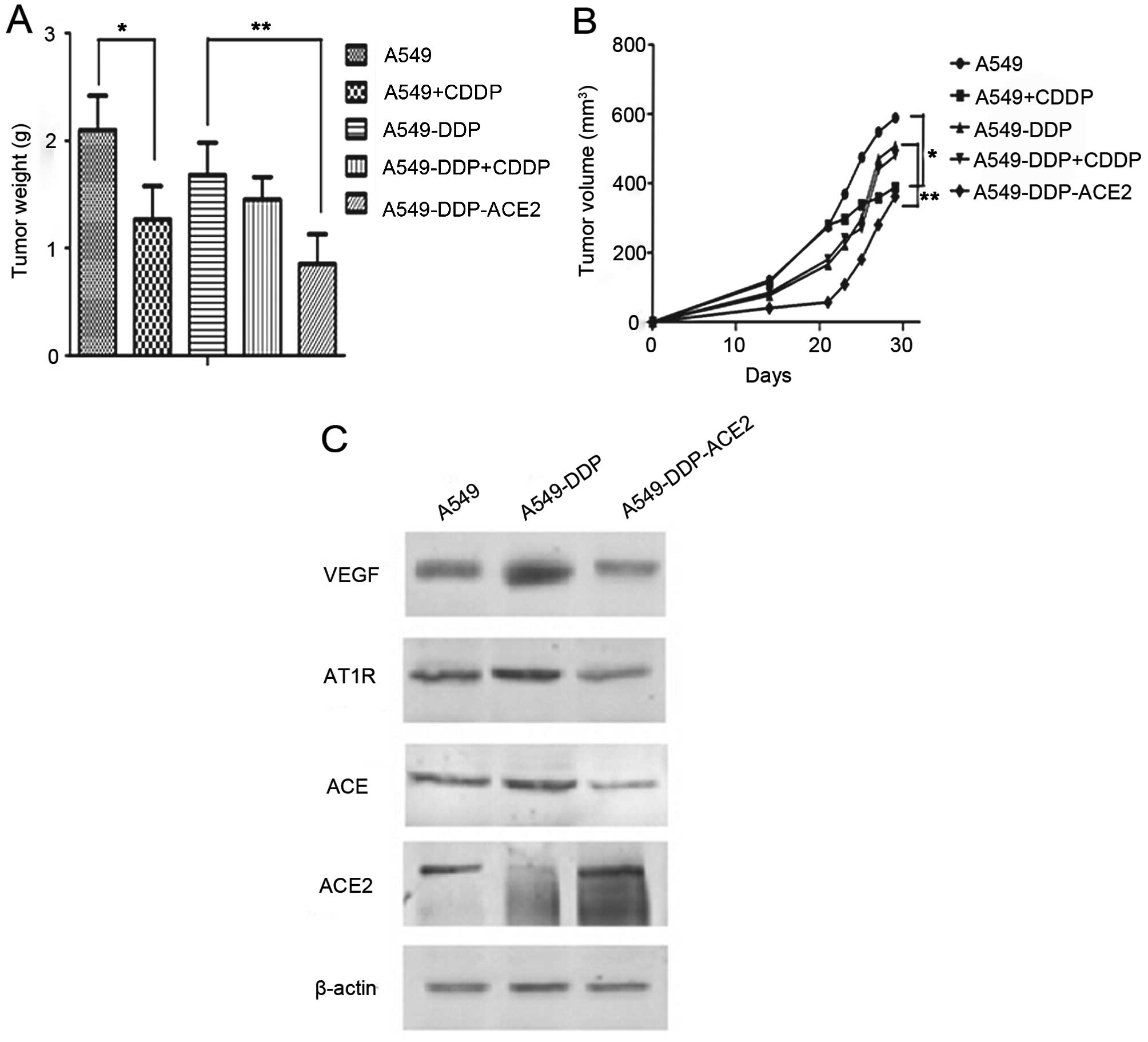
ACE2 overexpression inhibits the growth
of A549-DDP tumor cells in vivo
To investigate whether the ACE2 overexpression
inhibits pre-established tumor growth in nude mice, mice were
divided into 5 groups (A549, A549-CDDP, A549-DDP, A549-DDP-CDDP and
A549-DDP-ACE2). All of the animals were observed and tumors were
measured during a 31-day follow-up period. Tumor weights in the
A549-CDDP group were significantly reduced compared to the A549
group, while there were no significant changes in the A549-DDP
group compared with the A549-DDP-CDDP group (Fig. 7A). The graph also showed that the
tumor weight in the A549-DDP-ACE2 group was significantly reduced
compared to the A549-DDP group. The data from the tumor growth
curves are in agreement with the results for tumor weight and
indicated that tumor growth in the A549-DDP-ACE2 group was
significantly reduced when compared with the A549-DDP group
(Fig. 7B). The results demonstrate
that ACE2 overexpression significantly inhibited tumor growth in
the acquired platinum-resistant A549-DDP nude mouse model. We next
investigated whether ACE2 overexpression inhibits VEGF production
in acquired platinum-resistant NSCLC sublines in vivo. By
western blot analysis, A549-DDP cells showed significantly higher
ACE, AT1R and VEGF expression and decreased ACE2 production than
the A549 cells. The results also demonstrated that ACE, AT1R and
VEGF production decreased in the A549-DDP-ACE2 group when compared
to the A549-DDP group (Fig.
7C).
Discussion
In the present study, we analyzed RAS expression and
VEGF production following the development of acquired platinum
resistance in NSCLC. We also demonstrated the efficacy of ACE2 in
acquired platinum resistance sublines in vitro and in
vivo. To the best of our knowledge, this is the first study
that shows the expression and effect of ACE2 on platinum-resistant
cancer cells.
Current evidence supports the idea that both host
and tumor RAS are important for tumor growth and angiogenesis in
cancer (10,15). As a new part of RAS, the
ACE2/Ang-(1–7)/Mas axis also plays an important role in
cancer (16,17). Recent studies have shown that low
ACE2 expression may be a useful indicator of poor prognosis in HCC
and gallbladder cancer (18,19).
Our previous study also demonstrated the same results (11).
In the present study, we established acquired
platinum-resistant NSCLC sublines and found significantly higher
ACE, AT1R and VEGF expression and lower ACE2 expression in both the
A549-DDP and LLC-DDP sublines compared with their corresponding
parent cells. A previous study also showed changes in AT1R and VEGF
production after the development of acquired platinum resistance in
bladder cancer (3). However, the
present study focused more on the dysregulation of ACE/ACE2 in
acquired platinum-resistant NSCLC sublines. Notably, the ratio of
ACE/ACE2 is important in RAS. Recent research has shown that an
imbalance in this ratio leads to various diseases. Numerous studies
have also shown that ACE2 appears to be a negative regulator of RAS
and counterbalances the functions of ACE. ACE2 converts AngII to
Ang-(1–7), which has the opposite function of
AngII in cancer development. We previously found that Ang-(1–7)
inhibits the migration and invasion of NSCLC cells (20).
We then overexpressed ACE2 in acquired
platinum-resistant A549 cells and found that ACE2 overexpression
altered the expression of other RAS components. We also found that
ACE2 inhibited VEGF production and tumor-associated angiogenesis in
NSCLC. To the best of our knowledge, this is the first study to
show the effects of ACE2 and tumor-associated angiogenesis in
cancer. Identifying the key proteins involved in these processes is
vital for understanding carcinogenesis and for devising new
therapies. The ACE2/Ang-(1–7)/Mas axis, a new axis in RAS, has the
opposite effect of the classical ACE/AngII/AT1R axis. ACE2,
Ang-(1–7) and Mas have the same signaling
pathways. A recent study showed that Ang-(1–7)
suppressed hepatocellular carcinoma growth and angiogenesis through
inactivation of the p38 MAPK phosphorylation signaling pathway
(21). Our previous study also
demonstrated that Ang-(1–7) inhibited migration and invasion through
the PI3K/Akt and MAPK signaling pathways (20). In agreement with these studies, we
observed that the effect of ACE2 on the inhibition of VEGF
expression and angiogenesis in acquired platinum-resistant NSCLC
partly occured through PI3K/Akt and p38 MAPK inactivation.
Our previous studies showed that the ACE2
overexpression inhibited cell growth and VEGF production in
vivo (11,12). A recent study also showed that ACE2
suppressed tumor growth in gallbladder cancer (22). In the present study, we found that
ACE2 overexpression inhibited the growth of A549-DDP tumor cells
in vivo. Consistent with our data, Tanaka et al found
that ARB olmesartan suppressed tumor growth in acquired
platinum-resistant tumors (3).
Another previous study showed that Ang-(1–7)
inhibited tumor angiogenesis in human lung cancer xenografts with a
reduction in VEGF (23). We also
demonstrated that ACE2 overexpression inhibited VEGF production in
acquired platinum-resistant A549 cells. These data demonstrated
that the effect of ACE2 on cell growth inhibition may be through
the inhibition of VEGF production and tumor angiogenesis after the
development of acquired platinum resistance.
In summary, acquired platinum resistance may induce
tumor-associated angiogenesis and dysregulation of the local RAS
system. Our studies showed that ACE2 overexpression could suppress
VEGF expression in NSCLC and acquired-platinum resistant NSCLC
in vitro and in vivo. These results suggest the
therapeutic potential of ACE2 for controlling tumor growth and
tumor-associated angiogenesis after the development of acquired
platinum resistance.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (nos. 81201837 and
81370130).
References
|
1
|
Galluzzi L, Senovilla L, Vitale I, Michels
J, Martins I, Kepp O, Castedo M and Kroemer G: Molecular mechanisms
of cisplatin resistance. Oncogene. 31:1869–1883. 2012. View Article : Google Scholar
|
|
2
|
Pyaskovskaya ON, Dasyukevich OI, Kolesnik
DL, Garmanchouk LV, Todor IN and Solyanik GI: Changes in VEGF level
and tumor growth characteristics during Lewis lung carcinoma
progression towards cis-DDP resistance. Exp Oncol. 29:197–202.
2007.PubMed/NCBI
|
|
3
|
Tanaka N, Miyajima A, Kosaka T, Miyazaki
Y, Shirotake S, Shirakawa H, Kikuchi E and Oya M: Acquired platinum
resistance enhances tumour angiogenesis through angiotensin II type
1 receptor in bladder cancer. Br J Cancer. 105:1331–1337. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Fleming I, Kohlstedt K and Busse R: The
tissue renin-angiotensin system and intracellular signalling. Curr
Opin Nephrol Hypertens. 15:8–13. 2006. View Article : Google Scholar
|
|
5
|
Arici M and Erdem Y: Dual blockade of the
renin-angiotensin system for cardiorenal protection: An update. Am
J Kidney Dis. 53:332–345. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yoshiji H, Noguchi R, Toyohara M, Ikenaka
Y, Kitade M, Kaji K, Yamazaki M, Yamao J, Mitoro A, Sawai M, et al:
Combination of vitamin K2 and angiotensin-converting enzyme
inhibitor ameliorates cumulative recurrence of hepatocellular
carcinoma. J Hepatol. 51:315–321. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Nakai Y, Isayama H, Ijichi H, Sasaki T,
Sasahira N, Hirano K, Kogure H, Kawakubo K, Yagioka H, Yashima Y,
et al: Inhibition of renin-angiotensin system affects prognosis of
advanced pancreatic cancer receiving gemcitabine. Br J Cancer.
103:1644–1648. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Wilop S, von Hobe S, Crysandt M, Esser A,
Osieka R and Jost E: Impact of angiotensin I converting enzyme
inhibitors and angiotensin II type 1 receptor blockers on survival
in patients with advanced non-small-cell lung cancer undergoing
first-line platinum-based chemotherapy. J Cancer Res Clin Oncol.
135:1429–1435. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Tatokoro M, Fujii Y, Kawakami S, Saito K,
Koga F, Matsuoka Y, Iimura Y, Masuda H and Kihara K: Phase-II trial
of combination treatment of interferon-α, cimetidine,
cyclooxygenase-2 inhibitor and renin-angiotensin-system inhibitor
(I-CCA therapy) for advanced renal cell carcinoma. Cancer Sci.
102:137–143. 2011. View Article : Google Scholar
|
|
10
|
Imai N, Hashimoto T, Kihara M, Yoshida S,
Kawana I, Yazawa T, Kitamura H and Umemura S: Roles for host and
tumor angiotensin II type 1 receptor in tumor growth and
tumor-associated angiogenesis. Lab Invest. 87:189–198. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Feng Y, Wan H, Liu J, Zhang R, Ma Q, Han
B, Xiang Y, Che J, Cao H, Fei X, et al: The angiotensin-converting
enzyme 2 in tumor growth and tumor-associated angiogenesis in
non-small cell lung cancer. Oncol Rep. 23:941–948. 2010.PubMed/NCBI
|
|
12
|
Feng Y, Ni L, Wan H, Fan L, Fei X, Ma Q,
Gao B, Xiang Y, Che J and Li Q: Overexpression of ACE2 produces
antitumor effects via inhibition of angiogenesis and tumor cell
invasion in vivo and in vitro. Oncol Rep. 26:1157–1164.
2011.PubMed/NCBI
|
|
13
|
Arnaoutova I, George J, Kleinman HK and
Benton G: The endothelial cell tube formation assay on basement
membrane turns 20: State of the science and the art. Angiogenesis.
12:267–274. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Moyes AJ, Maldonado-Pérez D, Gray GA and
Denison FC: Enhanced angiogenic capacity of human umbilical vein
endothelial cells from women with preeclampsia. Reprod Sci.
18:374–382. 2011. View Article : Google Scholar
|
|
15
|
Ino K, Shibata K, Kajiyama H, Nawa A,
Nomura S and Kikkawa F: Manipulating the angiotensin system - new
approaches to the treatment of solid tumours. Expert Opin Biol
Ther. 6:243–255. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ender SA, Dallmer A, Lässig F, Lendeckel U
and Wolke C: Expression and function of the
ACE2/angiotensin(1–7)/Mas axis in osteosarcoma cell lines U-2 OS
and MNNG-HOS. Mol Med Rep. 10:804–810. 2014.PubMed/NCBI
|
|
17
|
Fan L, Feng Y, Wan HY, Ni L, Qian YR, Guo
Y, Xiang Y and Li QY: Hypoxia induces dysregulation of local
renin-angiotensin system in mouse Lewis lung carcinoma cells. Genet
Mol Res. 13:10562–10573. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li J, Yang ZL, Ren X, Zou Q, Yuan Y, Liang
L, Chen M and Chen S: ACE2 and FZD1 are prognosis markers in
squamous cell/adenosquamous carcinoma and adenocarcinoma of
gallbladder. J Mol Histol. 45:47–57. 2014. View Article : Google Scholar
|
|
19
|
Ye G, Qin Y, Lu X, Xu X, Xu S, Wu C, Wang
X, Wang S and Pan D: The association of renin-angiotensin system
genes with the progression of hepatocellular carcinoma. Biochem
Biophys Res Commun. 459:18–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ni L, Feng Y, Wan H, Ma Q, Fan L, Qian Y,
Li Q, Xiang Y and Gao B: Angiotensin-(1–7) inhibits the migration
and invasion of A549 human lung adenocarcinoma cells through
inactivation of the PI3K/Akt and MAPK signaling pathways. Oncol
Rep. 27:783–790. 2012.
|
|
21
|
Liu Y, Li B, Wang X, Li G, Shang R, Yang
J, Wang J, Zhang M, Chen Y, Zhang Y, et al: Angiotensin-(1–7)
suppresses hepatocellular carcinoma growth and angiogenesis via
complex interactions of angiotensin II type 1 receptor, angiotensin
II type 2 receptor and Mas receptor. Mol Med. 21:626–636. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zong H, Yin B, Zhou H, Cai D, Ma B and
Xiang Y: Loss of angiotensin-converting enzyme 2 promotes growth of
gallbladder cancer. Tumour Biol. 36:5171–5177. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Soto-Pantoja DR, Menon J, Gallagher PE and
Tallant EA: Angiotensin-(1–7) inhibits tumor angiogenesis in human
lung cancer xenografts with a reduction in vascular endothelial
growth factor. Mol Cancer Ther. 8:1676–1683. 2009. View Article : Google Scholar : PubMed/NCBI
|