Introduction
Hepatocellular carcinoma (HCC), one of the most
common malignancies, represents the third leading cause of
cancer-related death worldwide (1).
Although surgical resection and local ablation therapy, including
percutaneous ethanol injection and radiofrequency ablation, can be
considered as curative treatments for patients in the early stages,
intrahepatic and distant metastases are major obstacles in
obtaining long-term survival. More than 50% of patients have been
reported to relapse within 5 years and more than half of these
recurrences occur within 2 years after surgical resection due to
intrahepatic or distant metastases (2–5). Thus,
there is an urgent need for novel treatment strategies to prevent
metastasis.
Src, the archetypal member of a 9-gene family
(including Src, Yes, Fyn, Lyn, Lck, Hck, Fgr, Blk and Yrk), plays a
key role in cell adhesion, invasion, proliferation, survival and
angiogenesis during tumour development (6). To date, numerous studies have
indicated that Src expression and its activity are elevated in lung
(7), breast (8,9),
ovarian (10), pancreatic (11), colon (12,13)
and gastric cancers (14). Upon
activation, it recruits several signalling molecules to control
cell migration and cytoskeleton rearrangement, to increase growth
rates and invasion characteristics of tumours, and to resist
anoikis in tumour cells (6,15–17).
Conversely, reduced Src expression may potentially induce retarded
tumour growth and reduction of metastatic tendencies. Saracatinib
(AZD0530), a tyrosine kinase inhibitor (TKI) selective for Src, has
been associated with inhibition of cell migration and proliferation
in breast cancer (18);
additionally, it has been tested in a phase II trial to treat head
and neck squamous cell carcinoma (19) and advanced castration-resistant
prostate cancer (20). In HCC, Src
overexpression has been observed in 87.5% of the specimens, and
phosphorylated Src (tyr-416) (activated form) was found to be
closely associated with metastatic potential, such as microvessel
invasion or intrahepatic metastasis (unpublished data). Although
Src plays a role in tumorigenesis and in the development of various
human cancers, it remains largely unclear as to whether Src affects
tumour metastasis in HCC.
In the present study, we investigated the effect of
saracatinib on HCC. The results indicated that saracatinib reduced
lung metastasis, rather than inhibiting primary tumour growth. The
results revealed the promising effect of saracatinib in the
treatment of HCC metastasis and further studies suggest the use of
the Src inhibitor in combination with cytotoxic agents and other
anticancer treatments to improve HCC prognosis.
Materials and methods
Cell lines and reagents
HCC cell lines (MHCC97H and Hep3B) and a human
normal hepatic cell line (L02) were obtained from the Cell Bank of
the Chinese Academy of Sciences (Shanghai, China). All of these
cell lines were strictly maintained according to the supplier's
instructions and established procedures.
Polyclonal antibodies to Src, FAK and NADPH were
obtained from Santa Cruz Biotechnology (Santa Cruz, CA, USA).
Polyclonal antibodies to Stat3, p-Stat3 (Y705) and p-Src (Y416)
were obtained from Cell Signalling Technology (Danvers, MA, USA).
Polyclonal antibodies to p-FAK (Y861) were obtained from Invitrogen
by Life Technologies (Carlsbad, CA, USA). Saracatinib was obtained
from AstraZeneca International (Alderley Park, UK) and was
dissolved in dimethyl sulfoxide (DMSO; Fermentas, Pittsburgh, PA,
USA) to a stock solution of 10 mM and stored at −20°C.
Cell proliferation assay
Cell proliferation was measured using Cell Counting
Kit-8 (CCK-8), according to the manufacturer's protocols (Dojindo,
Kumamoto, Japan). Briefly, cells (MHCC97H, Hep3B and L02) were
triplicate plated onto 96-well plates (2.0×103
cells/well). Twelve hours later, saracatinib was added to the cells
at doses of 0-128.0 µM (including 0, 0.125, 0.25, 0.5, 1.0,
2.0, 8.0, 16.0, 32.0, 64.0 and 128.0 µM), and were incubated
for 0, 24, 48 or 72 h. The medium was then refreshed, followed by
the addition of 10 µl/well of CCK-8 solution and incubated
at 37°C for an additional 2 h. The absorbance at 450 nm represented
the number of viable cells and was measured using a Bio-Rad 680
microplate reader (Bio-Rad, Hercules, CA, USA).
Wound healing assay
The monolayer of cells (MHCC97H, Hep3B and L02) was
wounded by scraping with a 200-µl pipette tip and rinsed
several times with sterile phosphate-buffered saline (PBS) to
remove dislodged cells. Cells that had migrated into the wound area
were stained with crystal violet and their images were captured for
analysis.
Cell migration and invasion assays
Cell migration was assessed using a Transwell™
Permeable Supports system (Corning, Corning NY, USA) according to
the manufacturer's instructions. Briefly, cells (MHCC97H, Hep3B and
L02) were detached from the cell culture plates, washed with PBS
buffer, and resuspended to 1×103 cells/µl with
serum-free culture medium [Dulbecco's modified Eagle's medium
(DMEM), minimum essential medium (MEM) and RPMI-1640 medium,
respectively]. The cells were seeded onto the upper chambers of
Matrigel-coated (BD Biosciences, Franklin Lakes, NJ, USA) filter
inserts (8.0-µm pore size) and mixed with 0, 0.1 or 1.0
µM saracatinib (100 µl) per well. Culture medium (600
µl) containing 10% FBS was added to the lower chambers as a
chemoattractant. After being cultured for 48 h, filter inserts were
removed from the wells. The cells on the upper surface of the
filter were wiped with a cotton swab. The filter was fixed with 4%
paraformaldehyde for 10 min and stained with Giemsa dye (Sigma,
Munich, Germany) for 15 min. The cells invading the lower surface
were counted under an inverted microscope. The migration assay was
performed as described for the invasion assay, but without the
coating of Matrigel.
Colony formation assay
MHCC97H, Hep3B or L02 cells (1×103
cells/well) were mixed with 200 µl of 0.6% agarose, 150
µl of complete medium and 50 µl of a solution
containing 0, 0.1 or 1.0 µM saracatinib, and then seeded
into 6-well plates pre-coated with 0.6% low melting point agarose.
Photomicrographs of colonies were captured 2 weeks later and
quantified by counting the number of colonies formed.
Western blotting
Cells grown in 100-mm dishes were washed twice with
ice-cold PBS before lysis by incubation for 20 min in 1 ml of
ice-cold cell lysis RIPA buffer [10 mM Tris (pH 8.0), 150 mM NaCl,
1% sodium deoxycholate, 0.1% sodium dodecyl sulphate (SDS), 1%
Triton X-100, 10 mg/ml leupeptin, 10 mg/ml aprotinin and 1 mM
phenylmethylsulfonyl fluoride (PMSF)]. The protein concentrations
of the lysates were measured using a Bradford protein assay kit
(Bio-Rad). Equivalent amounts of protein were mixed with 6X
SDS-polyacrylamide gel electrophoresis (PAGE) sample buffer,
electrophoresed in a 4–20% linear gradient Tris-HCl-Ready Gel
(Bio-Rad), and then transferred to polyvinylidene fluoride (PVDF)
membranes. The membranes were blocked with 5% non-fat dry milk in
Tris-buffered saline, pH 7.4, containing 0.05% Tween-20 and were
incubated with specific primary antibodies and horseradish
peroxidase-labelled secondary antibodies (Rockland, Gilbertsville,
PA, USA) according to the manufacturer's instructions. The protein
of interest was visualized and quantitated using the LI-COR Odyssey
Infrared Imaging system (LI-COR Biosciences, Lincoln, NE, USA).
Cells (5×106) were lysed on ice with RIPA buffer [10 mM
Tris (pH 8.0), 150 mM NaCl, 1% sodium deoxycholate, 0.1% SDS, 1%
Triton X-100, 10 mg/ml leupeptin, 10 mg/ml aprotinin and 1 mM
PMSF]. Next, 50 µg of whole cell protein extracts were
separated by 10% SDS-PAGE and transferred onto a PVDF membrane. The
membrane was subsequently blocked with 5% bovine serum albumin
(BSA; Sigma-Aldrich, Inc.) in Tris-buffered saline plus Tween-20
(TBST) for 1 h, and then incubated overnight at 4°C with specific
primary antibodies, followed by washing with TBST buffer and
incubation with horseradish peroxidase-conjugated secondary
antibodies for 1 h at room temperature. After intensive TBST
washing, the membrane was subjected to chemiluminescent detection
using the Bio-Rad Imaging Lab system.
HCC orthotopic xenograft model and
evaluation of lung metastasis
The HCC orthotopic xenograft model with high
metastatic potential was established with the MHCC-97 cell line.
Four-week-old nude mice (BALB/c nu/nu) were subcutaneously injected
with 1×107/0.2 ml of MHCC97H cells in the left upper
flank region to establish subcutaneous xenograft models. Four weeks
later, the tumour nodules with a diameter of 0.8 cm were removed
and cut into 1 mm3 pieces and implanted into the livers
of the nude mice to establish orthotopic xenograft models. The 24
mice were randomized into three groups, each comprised of eight
mice. Three days later, the mice were orally administered
saracatinib daily at doses of 0, 50 or 100 mg/kg/day for 35
consecutive days. Mouse weights were evaluated biweekly. On day 35,
all mice were sacrificed. The livers and lungs were collected, the
wet weights were determined, and then the livers and lungs were
immediately fixed in 10% neutral formalin solution for further
immunohistochemistry. The liver wet weights were substituted for
tumour weights for evaluation of tumour growth inhibition by
saracatinib. Lung metastasis was determined by examining
consecutively the serial sections of total lungs, using hematoxylin
and eosin staining under a microscope. Metastasis inhibition was
then evaluated determining the metastasis rate of all mice in the
groups and the number of metastatic foci for the whole lung of each
mouse (21). Animal care and
procedures were approved by our Institutional Animal Care and Use
Committee (Ethics Committee of Haikou People's Hospital; permit no.
2013–14).
Immunohistochemistry
Immunohistochemistry was performed as previously
described (22). Briefly, after
deparaffinization, hydration and blocking of endogenous peroxidase
(0.3% H2O2 for 20 min); antigen retrieval was
performed using Tris-ETDA (pH 9.0) by microwave heating for 5 min.
Sections were blocked with 5% BSA for 30 min at room temperature,
and then the primary antibodies, including rabbit polyclonal
anti-p-FAK (Y-861) (diluted 1:300), rabbit polyclonal anti-p-Src
(Y-416) (diluted 1:100), rabbit polyclonal anti-p-Stat3 (Y-705)
(diluted 1:100) or rabbit IgG serum (negative control), were
incubated overnight at 4°C. Sections were detected using the
EnVision detection system (Dako Denmark A/S, Glostrup, Denmark) and
DAB following counterstaining with hematoxylin, and then dehydrated
through alcohol and xylene before mounting under coverslips. Slides
were evaluated semi-quantitatively at a magnification of ×100 under
light microscopy in a blinded fashion on a scale of 0–4.
Statistical analysis
All statistical analysis was performed using
GraphPad Prism version 5.0. Quantitative variables are expressed as
the mean ± standard deviation (SD) and were analysed by ANOVA. The
χ2 or Fisher's exact tests were applied to compare
qualitative variables. Results were deemed statistically
significant at P<0.05.
Results
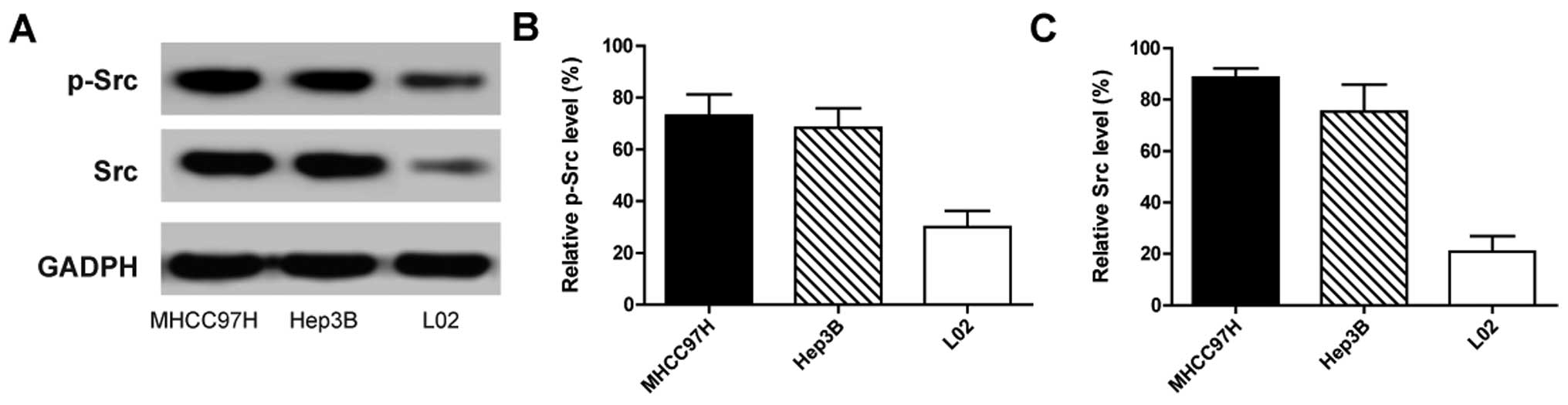
Src kinase activation correlates with the
metastatic potential of HCC in vivo and in vitro
To investigate Src expression and its correlation
with the metastasic potential in HCC, three cell lines (MHCC97H,
Hep3B and L02) were used. Among the three cell lines, a high lung
metastasic rate reaching 100% was found in mice injected with the
MHCC97H cells, whereas no lung metastasis was observed in mice
injected with Hep3B and L02 cells (data not shown). As
autophosphorylation of Src at tyrosine 416 (p-Src-Y416)
is a marker of its activity (23),
we next evaluated the expression of p-Src-Y416 and total
Src in the three cell lines. The results indicated that Src was
expressed in all three cell lines, but there were significantly
different activation levels in the three cell lines. The
p-Src-Y416 and Src expression in MHCC97H, a cell line
with high metastatic potential, was much higher than that in the
other two cell lines (Fig. 1). The
results indicated that the metastatic potential of HCC was closely
related to Src activation.
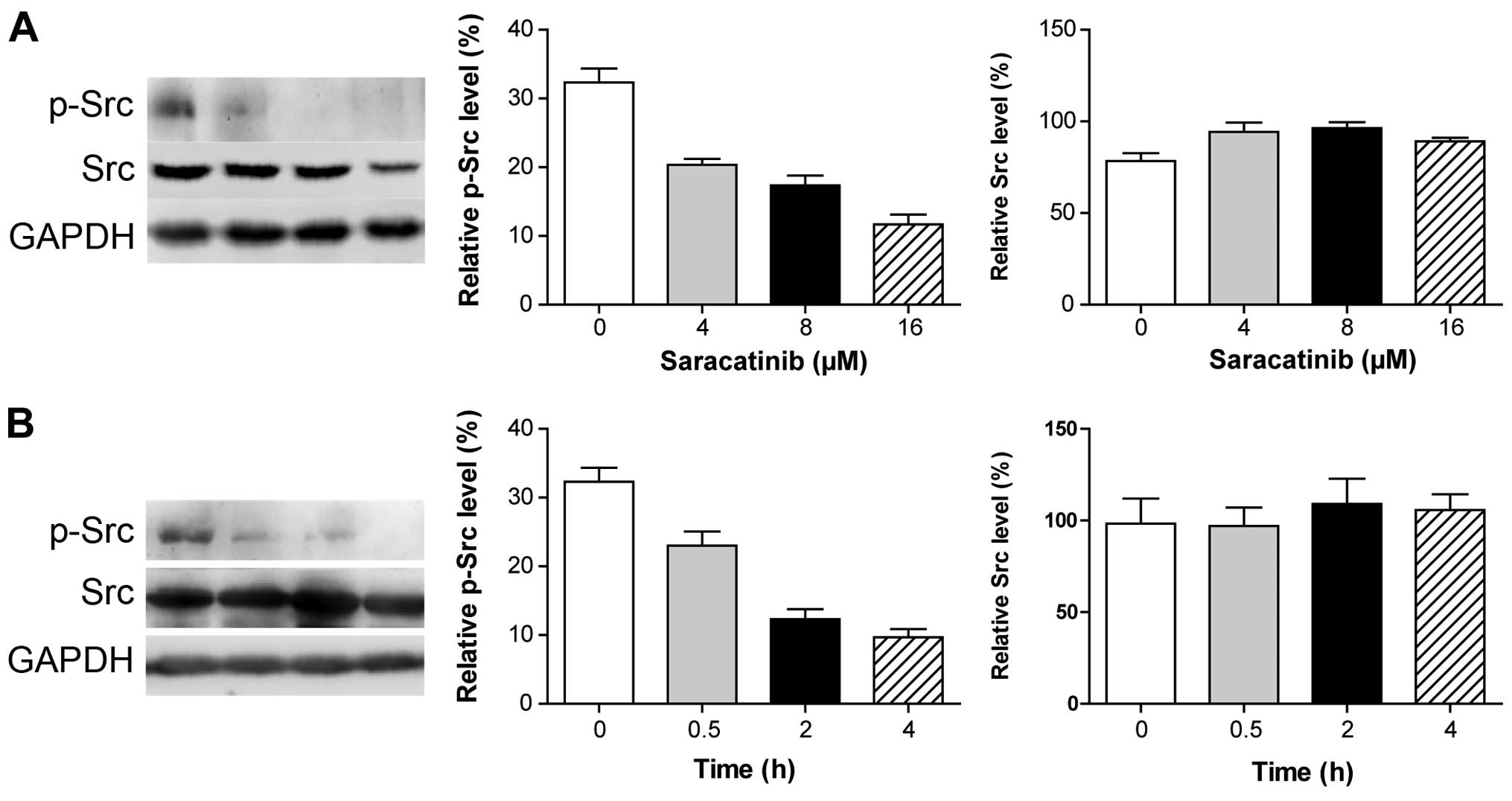
Saracatinib is involved in the inhibition
of Src phosphorylation in the MHCC97H cells
As saracatinib has been reported to inhibit Src
activation in prostate cancer cell lines (24), we determined whether saracatinib is
also an inhibitor of Src in HCC cell lines with high metastatic
potential. Treatment of the MHCC97H cells with saracatinib
partially prevented the phosphorylation of Src in a dose- and
time-dependent manner (Fig. 2).
This suggests that saracatinib is involved in inhibition of Src
activation in HCC cell lines with high metastatic potential.
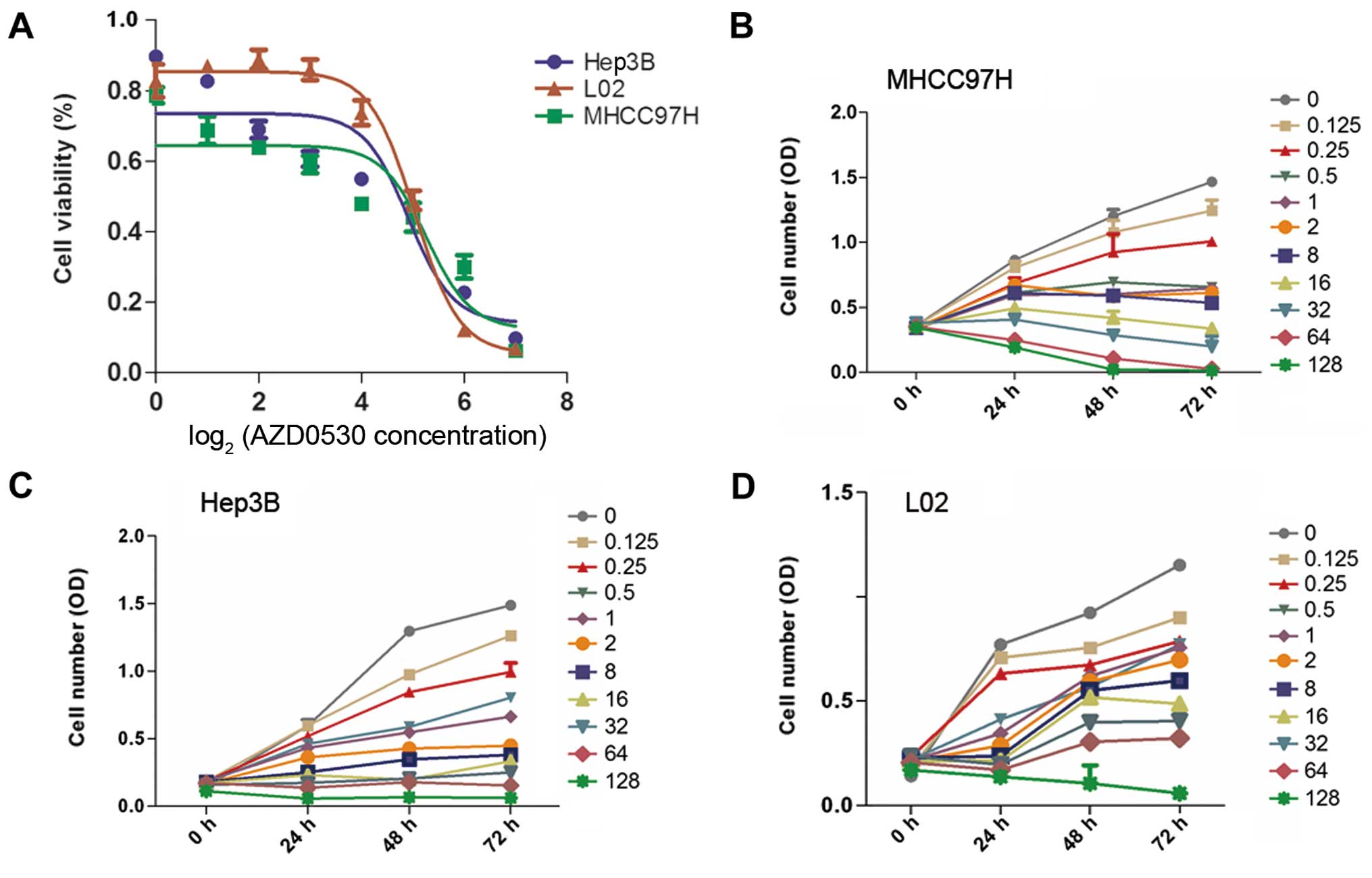
Saracatinib inhibits the invasive
potential of HCC cells
To evaluate the antiproliferative effect of
saracatinib on HCC, the MHCC97H, Hep3B and L02 cell lines treated
with saracatinib were used. The results showed that HCC
proliferation was inhibited significantly in a dose-dependent
manner in all three cell lines. The IC50 values in the
MHCC97H, Hep3B and L02 cell lines were 14.09, 22.75 and 31.06
µM, respectively (Fig.
3).
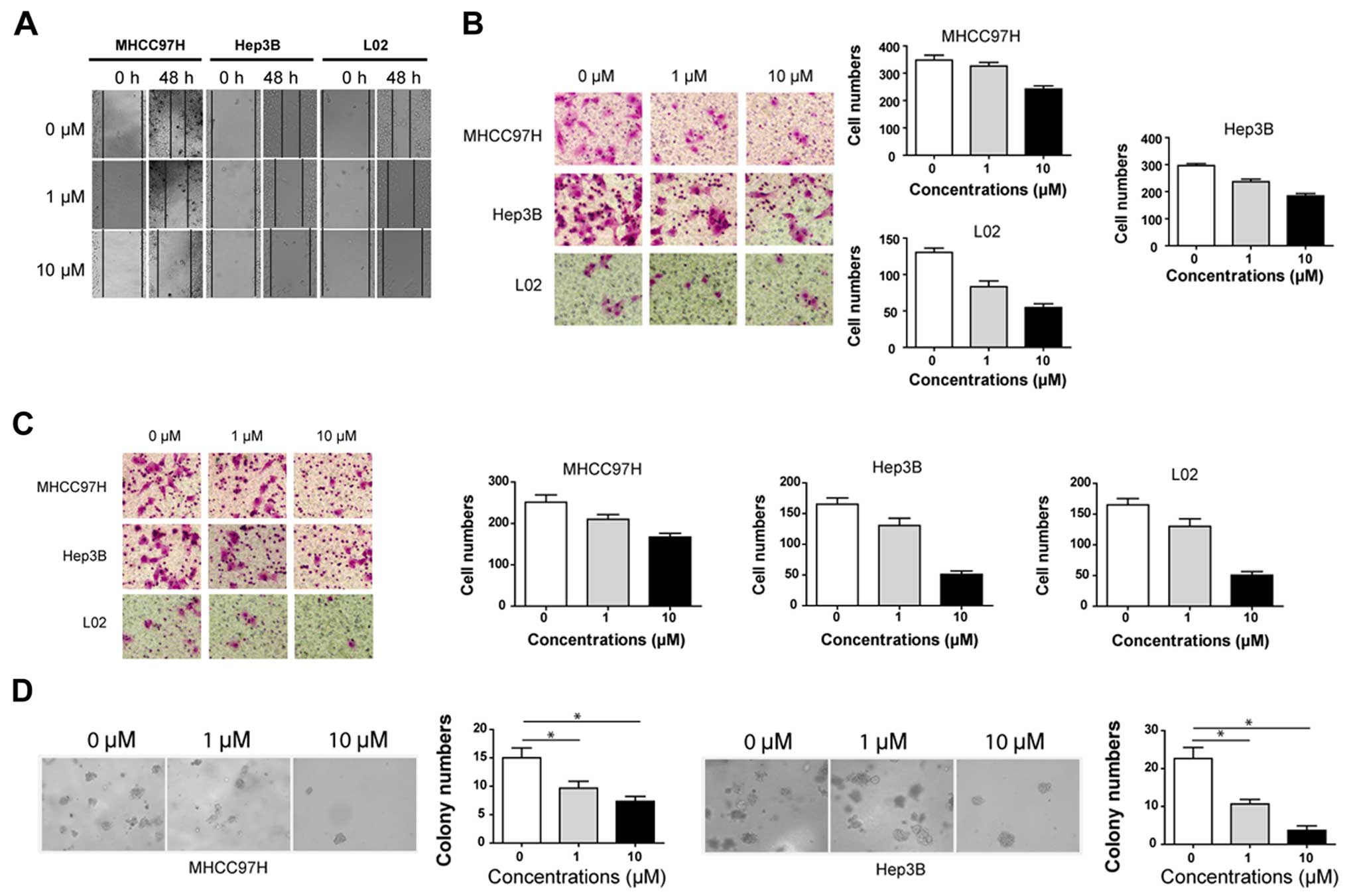
Subsequently, the wound healing and Transwell
migration assays were used to evaluate cell migration capacity.
Cell migration was significantly inhibited by saracatinib in all
three cell lines in a dose-dependent manner both in the wound
healing assay (Fig. 4A) and in the
Transwell migration assay (Fig.
4B). For the Transwell migration assay, the average numbers of
migrated cells in the MHCC97H cell line treated with 0, 1.0, and
10.0 µM saracatinib were 348.0±31.4, 326.7±21.8 and
242.0±20.3, respectively. Inhibitory effects were also observed in
the Hep3B and L02 cell lines. The numbers of migrated cells in the
Hep3B cell line treated with 0, 1.0 and 10.0 µM saracatinib
were 296.7±11.6, 237.3±15.2 and 184.3±15.6, respectively; while in
the L02 cell line they were 130.3±10.0, 83.3±13.3 and 54.7±9.0,
respectively.
The Matrigel invasion chamber assay revealed that
saracatinib significantly inhibited the invasion of MHCC97H, Hep3B
and L02 cells. After being treated with 0, 1.0 and 10.0 µM
saracatinib, the average numbers of invaded cells were 251.3±31.0,
210.0±20.0 and 167.0±16.1 in the MHCC97H cell line; 165.0±18.0,
130.3±21.0 and 50.7±10.1 in the Hep3B cell line; and 120.3±10.5,
83.3±13.3 and 47.0±16.1 in the L02 cell line, respectively
(Fig. 4C).
As colony formation is a vital feature in the high
metastasic potential of carcinomas, the impact of saracatinib on
colony formation in the MHCC97H and Hep3B cell lines was observed.
Considering L02 is a normal hepatocyte cell line, we did not use
this cell line in this experiment. The results showed that colony
formation was significantly inhibited with saracatinib. After
treatment with 0, 1.0 or 10.0 µM saracatinib for 14 days,
the numbers of formed spheres were 15.0±3.0, 9.7±2.1 and 7.3±1.5 in
the MHCC97H cell line, respectively (P<0.05; Fig. 4D left); and 22.7±5.0, 10.7±2.1 and
3.7±2.1 in the Hep3B cell line, respectively (P<0.05; Fig. 4D right). These findings suggest that
Src promotes migration and invasion of HCC cells in vitro,
and these capacities are inhibited by saracatinib.
Saracatinib inhibits the metastasic
potential in the HCC orthotopic xenograft model
To verify the inhibitory effect of saracatinib on
the HCC metastasic potential in vivo, the effect of
saracatinib on HCC metastasis in an orthotopic xenograft model was
then tested.
Following administration of 50 or 100 mg/kg/day
saracatinib for 5 weeks, tumour growth and lung metastasis were
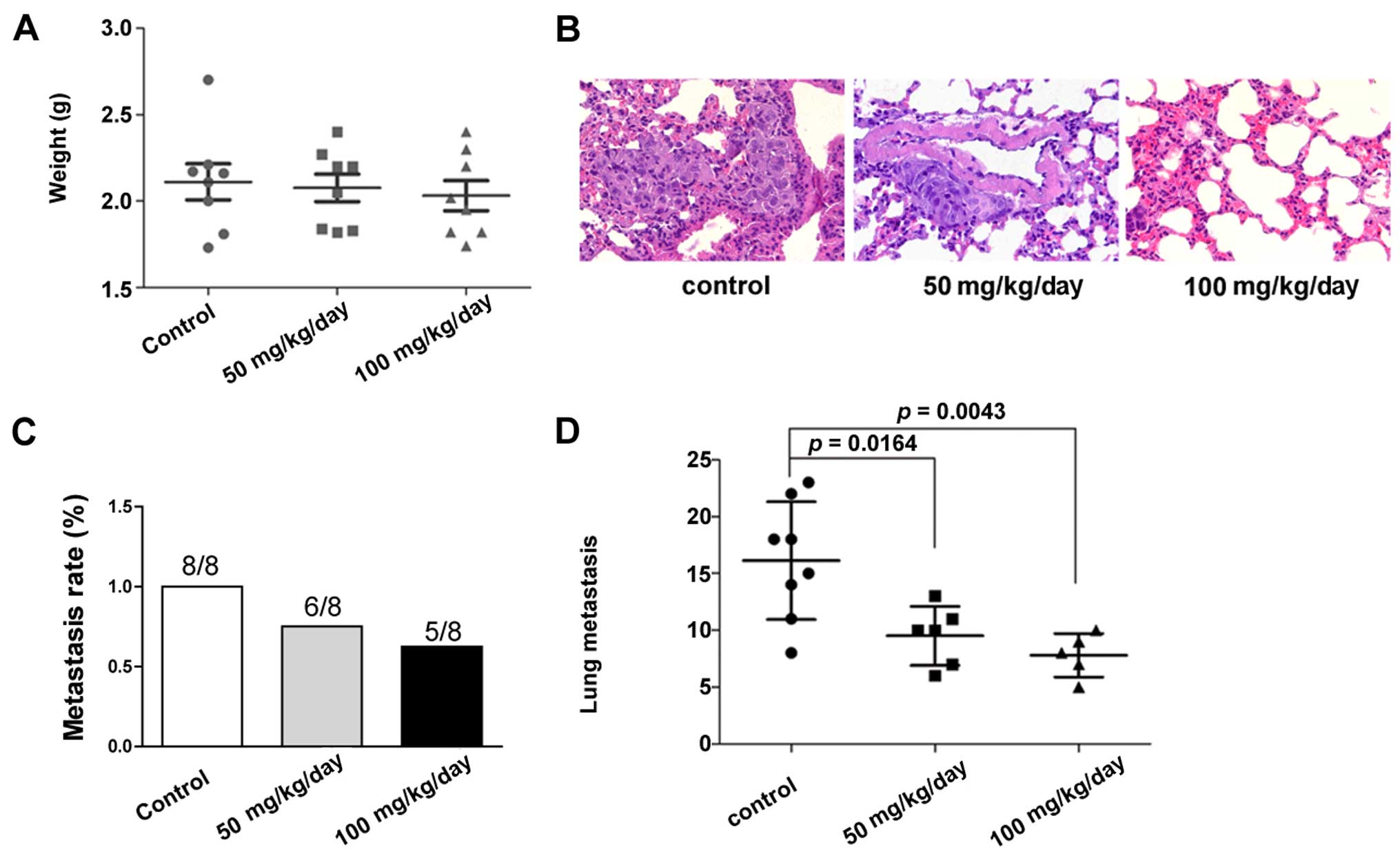
evaluated. The results showed that the liver weight (represented as
the tumour weight) was slightly different between the groups
(Fig. 5A). The results indicated
that growth inhibition of tumour was not achieved at the present
dose. We next compared the effect of saracatinib on the lung
metastasis of the model. The results showed that 8/8, 6/8 and 5/8
nude mice developed lung metastasis in the control, 50 and 100
mg/kg/day group, respectively (Fig. 5B
and C). Although no statistically significant difference was
found in the lung metastasis rates among the groups, inhibitory
effects were identified regarding the number of metastatic foci
after saracatinib treatment. The numbers of lung metastatic foci
were 15.63±4.98, 9.17±2.56 and 7.00±1.58 in the control group, 50
and 100 mg/kg/day group, respectively. The results revealed
saracatinib inhibition as evaluated by the number of metastasis
foci either with 50 mg/kg/day (P=0.0164) or 100 mg/kg/day
saracanitib (P=0.0043; Fig.
5D).
Saracatinib inhibits Src phosphorylation
and downstream signals of FAK and Stat3 phosphorylation
Src plays a vital role in the modulation of cellular
signalling pathway components relevant to cell proliferation,
migration, invasion and angiogenesis, including receptor tyrosine
kinase, FAK, Stat3 and adaptor protein P130Cas (25–28).
Therefore, Src activity and its downstream signals were evaluated
in vitro and in vivo to further understand the
underlying mechanisms involved in the anti-metastatic effect of
saracatinib. Phospho-Src (Y416), which is known to
activate Src, was significantly inhibited after treatment with
saracatinib in the MHCC97H cells. FAK and Stat3 are known to be
downstream signals of Src and their phosphorylation was also
evaluated following Src inhibition. The results revealed that Y816
phosphorylation of FAK and Y705 phosphorylation of Stat3 were
inhibited as well, indicating that the anti-metastasic effect of
saracatinib involved inhibition of FAK and Stat3. These results
support that the signalling axis Src/FAK/Stat3 plays a crucial role
in promotion of metastasis of HCC (29–31).
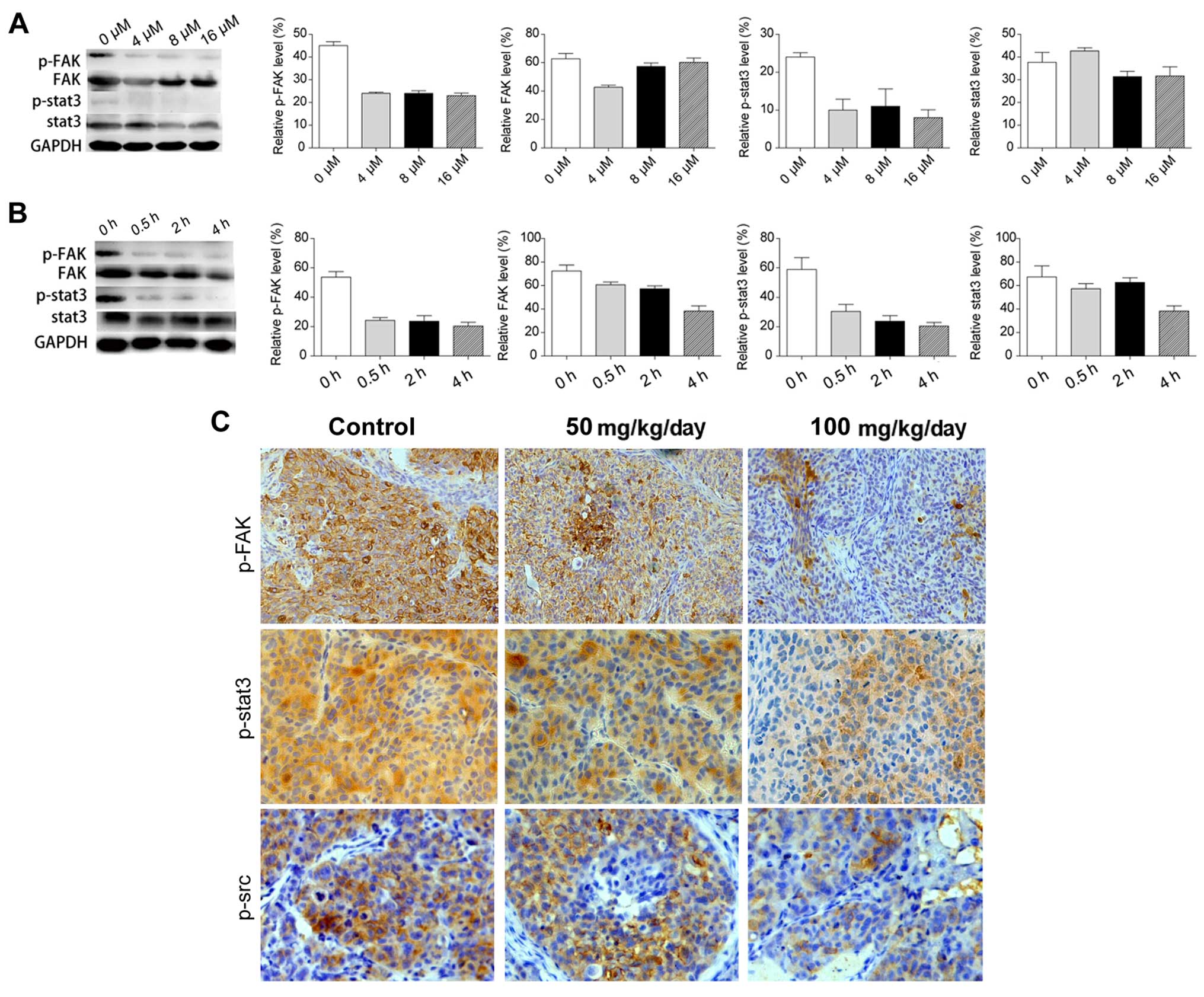
The effect of saracatinib on the inhibition of the Src/FAK/Stat3
axis in a dose- and time-dependent manner was confirmed (Fig. 6A and B). In accordance to in
vivo metastasis inhibition of saracatinib in the orthotopic
xenograft model, phosphorylation of Src, FAK and Stat3 were
inhibited in HCC tissues using immunohistochemistry. However,
phosphorylation was not completely inhibited in vivo at
doses of 50 and 100 mg/kg/day (Fig.
6C). This result may explain the limited anti-metastasic
effects of saracatinib in nude mice.
Discussion
Intrahepatic and distant metastases represent a key
challenge to achieve long-term survival for patients with HCC.
Studies concerning anti-metastasis of HCC have attracted
considerable attention in recent years. Various achievements have
been obtained in studies from the exploration of possible targets
to prevent metastasis, such as miR-124 (32), erythroblastic leukemia viral
oncogene homolog 3 (ERBB3) (33),
matrix metalloproteinase (MMP), c-Met, CD151, PDGFRα, transcription
factor late SV40 factor (LSF) (34)
and p38MAP kinase (35). However,
promising targets are not yet available for clinical use in
HCC.
The role of Src kinases has recently been well
defined in cancer tumorigenesis, tumour progression and metastasis.
Dysregulation of Src has been considered to lead malignant
transformation and tumour progression, and to promote metastasis in
various types of cancers (36–38). A
small amount of reagents have been developed to inhibit Src, and
also have been applied in phase I or II trials (39–41).
Although phase II clinical trials for Src inhibitors have shown a
modest effect against tumour growth in breast cancer, as well as
head and neck squamous carcinoma, clinical significance is being
overlooked for its metastatic role in cancers, including HCC
(19,41,42).
In HCC, 36/45 (55.4%) patients were identified with overexpressed
activated Src, and its overexpression was closely associated with
proliferation and metastatic potential (43–45);
however, the significance of inhibition of Src in HCC metastasis
has not been elucidated.
In the present study, we tested the effects of
saracatinib, a specific Src inhibitor, on HCC cell proliferation,
migration, invasion and colony formation in vitro. We found
that Src activation was correlated with the metastasis potential of
HCC cell lines. MHCC97H, a cell line with high metastatic
potential, showed a higher expression level of total Src and
activated Src compared with the other two cell lines. Saracatinib
effectively inhibited the activation of Src and its downstream
signals (FAK and Stat3), as well as the metastatic potential
(including cell migration, invasion and colony formation) in the
two HCC cell lines. In the orthotopic xenograft HCC model,
saracatinib inhibited lung metastasis, without influencing primary
tumour inhibition. These results indicated that Src is mainly
involved in the process of cancer metastasis, and suggest that, for
anticancer therapy, the use of saracatinib alone is not sufficient
to effectively control cancer growth. Thus, saracatinib combined
with other antiproliferatory agents may be used to block metastasis
formation, and to reduce primary tumour growth. Furthermore,
although inhibition of metastasis was observed in the orthotopic
xenograft model, saracatinib was not capable of completely blocking
lung metastasis and activation of the Src/FAK/Stat3 axis,
indicating that a more complex mechanism exists for Src activation
such as crosstalk regulation or feedback regulation (46). For example, in breast cancer, Src
was activated by tyrosine kinase receptor and the estrogen
receptor; although, saracatinib alone did not effectively inhibit
tumour growth and related signalling molecules, including
phosphorylated Src (Y416) and its downstream signalling
molecules (Akt and MAPK). However, saracatinib combined with
anastrozole (an estrogen inhibitor) significantly inhibited tumour
growth and Src (Y416), Akt (T473) and MAPK
(T202/Y204) phosphorylation (41).
Our present results also indicate that the anti-metastatic effect
of saracatinib should not be overlooked, even if it fails to
completely inhibit lung metastasis in the orthotopic model.
Notably, the Hep3B cells, with non-metastatic
potential were also found to possess relatively high levels of
p-Src and Src. Despite this, there is evidence of a prominent role
of Src in metastasis and in other tumour events such as
epithelial-mesenchymal transition (EMT) and in the development of
invasion in various tumours (47,48).
In the future, we may prove more evidence of the role of Src in
other HCC progression-related events, such as cell adhesion,
invasion, proliferation, survival and angiogenesis.
Moreover, the emerging role of Src has also been
demonstrated in mediating resistant tumours (46,49).
Application of saracatinib or its combination with other targeted
reagents (eg. trastuzumab) is encouraged based on more detailed
activation mechanisms of Src in overcoming various resistance
mechanisms, which warrant further investigation, particularly in
systemic chemotherapy-resistant HCC.
Abbreviations:
|
HCC
|
hepatocellular carcinoma
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
MEM
|
minimum essential medium
|
|
CCK-8
|
Cell Counting Kit-8
|
|
PVDF
|
polyvinylidene fluoride
|
|
ERBB3
|
erythroblastic leukemia viral oncogene
homolog 3
|
|
LSF
|
transcription factor late
|
Acknowledgments
The present study was supported by grants from the
International Cooperative Projects of Hainan Province (no.
KJHZ2014-14).
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Shah SA, Cleary SP, Wei AC, Yang I, Taylor
BR, Hemming AW, Langer B, Grant DR, Greig PD and Gallinger S:
Recurrence after liver resection for hepatocellular carcinoma: Risk
factors, treatment, and outcomes. Surgery. 141:330–339. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Regimbeau JM, Abdalla EK, Vauthey JN,
Lauwers GY, Durand F, Nagorney DM, Ikai I, Yamaoka Y and Belghiti
J: Risk factors for early death due to recurrence after liver
resection for hepatocellular carcinoma: Results of a multicenter
study. J Surg Oncol. 85:36–41. 2004. View Article : Google Scholar
|
|
4
|
Cha C, Fong Y, Jarnagin WR, Blumgart LH
and DeMatteo RP: Predictors and patterns of recurrence after
resection of hepatocellular carcinoma. J Am Coll Surg. 197:753–758.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cucchetti A, Piscaglia F, Caturelli E,
Benvegnù L, Vivarelli M, Ercolani G, Cescon M, Ravaioli M, Grazi
GL, Bolondi L, et al: Comparison of recurrence of hepatocellular
carcinoma after resection in patients with cirrhosis to its
occurrence in a surveilled cirrhotic population. Ann Surg Oncol.
16:413–422. 2009. View Article : Google Scholar
|
|
6
|
Yeatman TJ: A renaissance for SRC. Nat Rev
Cancer. 4:470–480. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Haura EB, Tanvetyanon T, Chiappori A,
Williams C, Simon G, Antonia S, Gray J, Litschauer S, Tetteh L,
Neuger A, et al: Phase I/II study of the Src inhibitor dasatinib in
combination with erlotinib in advanced non-small-cell lung cancer.
J Clin Oncol. 28:1387–1394. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hiscox S, Jordan NJ, Smith C, James M,
Morgan L, Taylor KM, Green TP and Nicholson RI: Dual targeting of
Src and ER prevents acquired antihormone resistance in breast
cancer cells. Breast Cancer Res Treat. 115:57–67. 2009. View Article : Google Scholar
|
|
9
|
Finn RS: Targeting Src in breast cancer.
Ann Oncol. 19:1379–1386. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wiener JR, Nakano K, Kruzelock RP, Bucana
CD, Bast RC Jr and Gallick GE: Decreased Src tyrosine kinase
activity inhibits malignant human ovarian cancer tumor growth in a
nude mouse model. Clin Cancer Res. 5:2164–2170. 1999.PubMed/NCBI
|
|
11
|
Ito H, Gardner-Thorpe J, Zinner MJ, Ashley
SW and Whang EE: Inhibition of tyrosine kinase Src suppresses
pancreatic cancer invasiveness. Surgery. 134:221–226. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bolen JB, Veillette A, Schwartz AM, DeSeau
V and Rosen N: Activation of pp60c-src protein kinase activity in
human colon carcinoma. Proc Natl Acad Sci USA. 84:2251–2255. 1987.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cartwright CA, Kamps MP, Meisler AI, Pipas
JM and Eckhart W: pp60c-src activation in human colon carcinoma. J
Clin Invest. 83:2025–2033. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Humar B, Fukuzawa R, Blair V, Dunbier A,
More H, Charlton A, Yang HK, Kim WH, Reeve AE, Martin I, et al:
Destabilized adhesion in the gastric proliferative zone and c-Src
kinase activation mark the development of early diffuse gastric
cancer. Cancer Res. 67:2480–2489. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Summy JM and Gallick GE: Src family
kinases in tumor progression and metastasis. Cancer Metastasis Rev.
22:337–358. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Biscardi JS, Ishizawar RC, Silva CM and
Parsons SJ: Tyrosine kinase signalling in breast cancer: Epidermal
growth factor receptor and c-Src interactions in breast cancer.
Breast Cancer Res. 2:203–210. 2000. View
Article : Google Scholar
|
|
17
|
Rajeshkumar NV, Tan AC, De Oliveira E,
Womack C, Wombwell H, Morgan S, Warren MV, Walker J, Green TP,
Jimeno A, et al: Antitumor effects and biomarkers of activity of
AZD0530, a Src inhibitor, in pancreatic cancer. Clin Cancer Res.
15:4138–4146. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hiscox S, Morgan L, Green TP, Barrow D,
Gee J and Nicholson RI: Elevated Src activity promotes cellular
invasion and motility in tamoxifen resistant breast cancer cells.
Breast Cancer Res Treat. 97:263–274. 2006. View Article : Google Scholar
|
|
19
|
Fury MG, Baxi S, Shen R, Kelly KW, Lipson
BL, Carlson D, Stambuk H, Haque S and Pfister DG: Phase II study of
saracatinib (AZD0530) for patients with recurrent or metastatic
head and neck squamous cell carcinoma (HNSCC). Anticancer Res.
31:249–253. 2011.PubMed/NCBI
|
|
20
|
Edwards J: Src kinase inhibitors: An
emerging therapeutic treatment option for prostate cancer. Expert
Opin Investig Drugs. 19:605–614. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Li Y, Tang Z, Ye S, Liu Y, Chen J, Xue Q,
Huang X, Chen J, Bao W, Yang J, et al: Establishment of human
hepatocellular carcinoma cell line with spontaneous pulmonary
metastasis through in vivo selection. Zhonghua Yi Xue Za Zhi.
82:601–605. 2002.In Chinese. PubMed/NCBI
|
|
22
|
Qian YB, Zhang JB, Wu WZ, Fang HB, Jia WD,
Zhuang PY, Zhang BH, Pan Q, Xu Y, Wang L, et al: P48 is a
predictive marker for outcome of postoperative interferon-alpha
treatment in patients with hepatitis B virus infection-related
hepatocellular carcinoma. Cancer. 107:1562–1569. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bjelfman C, Hedborg F, Johansson I,
Nordenskjöld M and Påhlman S: Expression of the neuronal form of
pp60c-src in neuroblastoma in relation to clinical stage and
prognosis. Cancer Res. 50:6908–6914. 1990.PubMed/NCBI
|
|
24
|
Chang YM, Bai L, Liu S, Yang JC, Kung HJ
and Evans CP: Src family kinase oncogenic potential and pathways in
prostate cancer as revealed by AZD0530. Oncogene. 27:6365–6375.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Frame MC: Src in cancer: Deregulation and
consequences for cell behaviour. Biochim Biophys Acta.
1602:114–130. 2002.PubMed/NCBI
|
|
26
|
Roy S, Ruest PJ and Hanks SK: FAK
regulates tyrosine phosphorylation of CAS, paxillin, and PYK2 in
cells expressing v-Src, but is not a critical determinant of v-Src
transformation. J Cell Biochem. 84:377–388. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Frame MC: Newest findings on the oldest
oncogene; How activated src does it. J Cell Sci. 117:989–998. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mitra SK and Schlaepfer DD:
Integrin-regulated FAK-Src signaling in normal and cancer cells.
Curr Opin Cell Biol. 18:516–523. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Brunton VG and Frame MC: Src and focal
adhesion kinase as therapeutic targets in cancer. Curr Opin
Pharmacol. 8:427–432. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Serrels A, McLeod K, Canel M, Kinnaird A,
Graham K, Frame MC and Brunton VG: The role of focal adhesion
kinase catalytic activity on the proliferation and migration of
squamous cell carcinoma cells. Int J Cancer. 131:287–297. 2012.
View Article : Google Scholar
|
|
31
|
Rajendran P, Li F, Shanmugam MK, Vali S,
Abbasi T, Kapoor S, Ahn KS, Kumar AP and Sethi G: Honokiol inhibits
signal transducer and activator of transcription-3 signaling,
proliferation, and survival of hepatocellular carcinoma cells via
the protein tyrosine phosphatase SHP-1. J Cell Physiol.
227:2184–2195. 2012. View Article : Google Scholar
|
|
32
|
Zheng F, Liao YJ, Cai MY, Liu YH, Liu TH,
Chen SP, Bian XW, Guan XY, Lin MC, Zeng YX, et al: The putative
tumour suppressor microRNA-124 modulates hepatocellular carcinoma
cell aggressiveness by repressing ROCK2 and EZH2. Gut. 61:278–289.
2012. View Article : Google Scholar
|
|
33
|
Hsieh SY, He JR, Hsu CY, Chen WJ, Bera R,
Lin KY, Shih TC, Yu MC, Lin YJ, Chang CJ, et al:
Neuregulin/erythroblastic leukemia viral oncogene homolog 3
autocrine loop contributes to invasion and early recurrence of
human hepatoma. Hepatology. 53:504–516. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yoo BK, Emdad L, Gredler R, Fuller C,
Dumur CI, Jones KH, Jackson-Cook C, Su ZZ, Chen D, Saxena UH, et
al: Transcription factor Late SV40 Factor (LSF) functions as an
oncogene in hepatocellular carcinoma. Proc Natl Acad Sci USA.
107:8357–8362. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Tan FL, Ooi A, Huang D, Wong JC, Qian CN,
Chao C, Ooi L, Tan YM, Chung A, Cheow PC, et al: p38delta/MAPK13 as
a diagnostic marker for cholangiocarcinoma and its involvement in
cell motility and invasion. Int J Cancer. 126:2353–2361. 2010.
|
|
36
|
Carretero J, Shimamura T, Rikova K,
Jackson AL, Wilkerson MD, Borgman CL, Buttarazzi MS, Sanofsky BA,
McNamara KL, Brandstetter KA, et al: Integrative genomic and
proteomic analyses identify targets for Lkb1-deficient metastatic
lung tumors. Cancer Cell. 17:547–559. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Byers LA, Sen B, Saigal B, Diao L, Wang J,
Nanjundan M, Cascone T, Mills GB, Heymach JV and Johnson FM:
Reciprocal regulation of c-Src and STAT3 in non-small cell lung
cancer. Clin Cancer Res. 15:6852–6861. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Tryfonopoulos D, Walsh S, Collins DM,
Flanagan L, Quinn C, Corkery B, McDermott EW, Evoy D, Pierce A,
O'Donovan N, et al: Src: A potential target for the treatment of
triple-negative breast cancer. Ann Oncol. 22:2234–2240. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Puls LN, Eadens M and Messersmith W:
Current status of SRC inhibitors in solid tumor malignancies.
Oncologist. 16:566–578. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Hannon RA, Finkelman RD, Clack G, Iacona
RB, Rimmer M, Gossiel F, Baselga J and Eastell R: Effects of Src
kinase inhibition by saracatinib (AZD0530) on bone turnover in
advanced malignancy in a Phase I study. Bone. 50:885–892. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Gucalp A, Sparano JA, Caravelli J,
Santamauro J, Patil S, Abbruzzi A, Pellegrino C, Bromberg J, Dang
C, Theodoulou M, et al: Phase II trial of saracatinib (AZD0530), an
oral SRC-inhibitor for the treatment of patients with hormone
receptor-negative metastatic breast cancer. Clin Breast Cancer.
11:306–311. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Guo C, Liu QG, Yang W, Zhang ZL and Yao
YM: Relation among p130Cas, E-cadherin and beta-catenin expression,
clinicopathologic significance and prognosis in human
hepatocellular carcinoma. Hepatobiliary Pancreat Dis Int.
7:490–496. 2008.PubMed/NCBI
|
|
43
|
Ito Y, Kawakatsu H, Takeda T, Sakon M,
Nagano H, Sakai T, Miyoshi E, Noda K, Tsujimoto M, Wakasa K, et al:
Activation of c-Src gene product in hepatocellular carcinoma is
highly correlated with the indices of early stage phenotype. J
Hepatol. 35:68–73. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Lau GM, Lau GM, Yu GL, Gelman IH, Gutowski
A, Hangauer D and Fang JW: Expression of Src and FAK in
hepatocellular carcinoma and the effect of Src inhibitors on
hepatocellular carcinoma in vitro. Dig Dis Sci. 54:1465–1474. 2009.
View Article : Google Scholar
|
|
45
|
Rajendran P, Ong TH, Chen L, Li F,
Shanmugam MK, Vali S, Abbasi T, Kapoor S, Sharma A, Kumar AP, et
al: Suppression of signal transducer and activator of transcription
3 activation by butein inhibits growth of human hepatocellular
carcinoma in vivo. Clin Cancer Res. 17:1425–1439. 2011. View Article : Google Scholar
|
|
46
|
Zhang S, Huang WC, Li P, Guo H, Poh SB,
Brady SW, Xiong Y, Tseng LM, Li SH, Ding Z, et al: Combating
trastuzumab resistance by targeting SRC, a common node downstream
of multiple resistance pathways. Nat Med. 17:461–469. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Guarino M: Src signaling in cancer
invasion. J Cell Physiol. 223:14–26. 2010.PubMed/NCBI
|
|
48
|
Kim LC, Song L and Haura EB: Src kinases
as therapeutic targets for cancer. Nat Rev Clin Oncol. 6:587–595.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Simpkins F, Hevia-Paez P, Sun J, Ullmer W,
Gilbert CA, da Silva T, Pedram A, Levin ER, Reis IM, Rabinovich B,
et al: Src Inhibition with saracatinib reverses fulvestrant
resistance in ER-positive ovarian cancer models in vitro and in
vivo. Clin Cancer Res. 18:5911–5923. 2012. View Article : Google Scholar : PubMed/NCBI
|