Introduction
Gastric cancer (GC) is one of the most common
malignant tumors in China with a high incidence and mortality rate
(1). Local invasion and distant
metastasis are leading causes for the dismal outcome of GC
patients. Despite recent advances in surgical techniques and new
chemotherapy regimens, the 5-year survival rate of GC patients
remains low. Therefore, investigation of molecular mechanisms
underlying GC development and progression are urgently needed to
develop novel diagnostics and therapies for GC.
Aberrant expression of microRNAs (miRNAs) is assumed
to play an important role in cancer development by regulating cell
proliferation, differentiation, apoptosis and carcinogenesis
(2). Previous studies have shown
that many miRNAs are aberrantly expressed during GC progression
(3,4). Although the mechanisms underlying
miRNA dysregulation in GC remain a challenge, recent studies have
shown that promoter CpG island hypermethylation plays a key role in
the silencing of numerous miRNAs (5). For example, DNA methylation of miR-210
increases the proliferation of gastric epithelium during chronic
H. pylori infection (6).
Moreover, miR-10b functions as a tumor suppressor but its
hypermethylation occurs in GC (7).
Thus, aberrant hypermethylation events in the regulatory regions of
miRNAs may play a key role in cancer development. In contrast,
miRNAs can also modulate epigenetic regulatory mechanisms of gene
expression by targeting DNA methyltransferase enzymes (DNMT1,
DNMT3a and DNMT3b). For example, the miR-29 family directly targets
DNMT3a and DNMT3b in lung cancer, thereby protecting against
changes in the existing DNA methylation status and suppressing
tumorigenesis (8). Moreover,
miR-152 can negatively regulate DNMT1 directly by targeting the
3′UTR of DNMT1 in breast (9),
ovarian (10) and liver cancer
(11), respectively. These findings
indicate an important role for miRNAs which target DNMTs during
carcinogenesis. Collectively, the epigenetic silencing of both the
miRNAs and miRNAs targeting DNMTs have been described to play a
role in cancer.
To further explore the underlying molecular
mechanisms of this interaction in GC, we hypothesized that an
interaction between miRNA and DNMTs could be contributing to
gastric carcinogenesis. In a previous study, the miR-200 family was
proved to function as a tumor suppressor by suppressing other genes
(12), but frequently silenced by
hypermethylation in many types of cancers (13–16).
Intriguingly, the miR-200 family was predicted to target DNMT3a
directly, as predicted by several in silico methods for
target gene prediction (PicTAR, TargetScan and MiRanda). Given the
fact that the methylation-sensitive miRNAs and DNMT-targeting
miRNAs may have the ability to form an epigenetic-miRNA regulatory
circuit, we hypothesized that a negative feedback loop exists
between the miR-200 family and DNMT3a in GC.
To test this hypothesis, we specifically
investigated: i) the expression of miR-200 family and DNM3a in a
panel of GC tissues; ii) the probable associations between miR-200
family levels and the clinicopathological features of patients with
GC; iii) the probable negative feedback loop between the miR-200
family and DNA methyltransferase; and iv) the probable roles of
miR-200c and DNMT3a in cell proliferation and invasion. The answer
to these questions would provide new insights into the molecular
mechanisms involved in GC development and metastasis as well as
provide a new strategy for GC diagnostic and treatment.
Materials and methods
Cell culture, 5-Aza-dC treatment and
transfection
The human GC cell lines NCI-N87
(well-differentiated), SGC7901 and AGS (moderately differentiated),
HGC27 and MGC803 (poorly differentiated), GES-1 (the human normal
gastric cell line) were cultured in RPMI-1640 medium (HyClone,
Logan, UT, USA) supplemented with 10% fetal calf serum and
incubated at 37°C with 5% CO2 in a humidified
atmosphere. To examine the role of methylation in the regulation of
miRNA expression, GC cells at 5×105 cells/ml were
cultured with 5-aza-2′-deoxycytidine (5-Aza-dC) (Sigma-Aldrich, St.
Louis, MO, USA) at 2.5 µM for 72 h. Transfections were
performed using a Lipofectamine 2000 kit (Invitrogen, Carlsbad, CA,
USA) according to the manufacturer's instructions. A total of 100
nM of siRNA against DNMT3a (sc-37757; Santa Cruz Biotechnology,
Santa Cruz, CA, USA), double-stranded miR-200c mimics (50 nM),
single-stranded miR-200c inhibitor (100 nM) (RiboBio, Guangzhou,
China) or their relative negative control RNAs were introduced into
cells.
Primary tissue samples
Cancer and adjacent non-cancerous tissues were
obtained from 46 GC patients who underwent gastrectomy between
January 2005 and December 2012 at Guangzhou First People's
Hospital. Tissue samples were immediately embedded in liquid
nitrogen and then stored at −80°C until RNA, DNA and protein could
be extracted. Written consent was obtained from each patient before
operation. The study protocol was approved by the Medical Ethics
Committee of Guangzhou First People's Hospital.
Plasmids and luciferase activity
assays
For DNMT3a overexpression, the coding sequence of
DNMT3a was PCR-amplified (2,739 bp) and cloned into
pcDNA3.1+. The primers used were as follows: DNMT3a
sense, 5′-cgcggatccgccaccATGCCCGCCATGCCCTCCAG-3′ and antisense,
5′-ataagaatgcggccgc TTACACACACGCAAAATACTCCTT-3′. Insertion was
confirmed by Sanger sequencing. Expression plasmids were
transfected into MGC803 cells using Lipofectamine 2000 according to
the manufacturer's instructions. For luciferase reporter assays,
the 3′UTRs of DNMT3a containing an intact miR-200c recognition
sequence was PCR-amplified (1,303 bp) and cloned into psiCHECK™-2
(Promega, Madison, WI, USA). A psiCHECK™-2 construct containing the
DNMT3a 3′UTR with mutations in the seed sequence was synthesized
using a QuikChange Site-Directed Mutagenesis kit (Agilent
Technologies, Santa Clara, CA, USA). All constructs were confirmed
by Sanger sequencing. The primers used were as follows:
DNMT3a-wild-type sense, 5′-CCGCTCGAGGGG ACATGGGGGCAAACTG-3′ and
antisense, 5′-CATAAGAATGCGGCCGCTACGTTTTGTATGTTTTTTTATTTG-3′; mutant
sense, 5′-CCACACAAGACATTTTTCTAGTCATAATCAGGTGCCTACCACACAGG and
antisense, 5′-CCTGTGTGGTAGGCACCTGATTATGACTAGAAAAATGTC TTGTGTGG-3′.
SGC7901 cells at 50% confluency were transiently transfected with
0.5 µg of reporter plasmids alone or co-transfected with or
without miRNA mimics or inhibitors using Lipofectamine 2000
according to the protocol. The luciferase activities of both
firefly and Renilla luciferase were quantified by a
Dual-Luciferase Reporter Assay System and the relative luciferase
activity was obtained according to the manufacturer's instructions
(GloMax Detection System) (both from Promega). Independent
triplicate experiments were carried out for all samples.
RNA extraction and quantitative real-time
PCR
RNA was isolated using TRIzol (Invitrogen) according
to the manufacturer's protocol. The expression levels of miR-200b,
miR-200c and miR-141 were measured by reverse transcription-PCR
analysis according to the miRCURY LNA™ Universal RT microRNA PCR
manual (Exiqon, Vedbaek, Denmark). The expression of miR-200 family
member was normalized to that of RNU6B, small nuclear RNA
expression.
DNA methylation analysis
The CpG island of the miR-200c gene was searched in
the miRBase database (release 24 June, 2013), the UCSC Genome
Browser on Human Assembly (hg 19; release February 2009) and PubMed
(17). Genomic DNA from patient
samples and MGC803 cells were isolated by QIAamp DNA Mini kit
(Qiagen, Germantown, MD, USA). Bisulfite conversion of genomic DNA
was performed using the EpiTect Bisulfite kit (Qiagen) and then
amplified by PCR using methylation-specific primers. The primers
used were as follows: miR-200c sense, 5′-GGGGTAGGGGAAGGTGGTTTA-3′
and antisense, 5′-CACCACCCCAATCCCTAAAAACACT-3′; the primer used for
sequencing was as follows: GGGAAGGTGGTTTAGA. We investigated the
methylation status of GC tissues using pyrosequencing. DNA
methylation analysis of bisulfite PCR amplicons for all samples was
performed on the PyroMark Q24 instrument according to the
manufacturer's instructions (Qiagen). DNA methylation level was
defined as the average percent methylation of each individual
CpG-unit averaged across the regions. We evaluated the impact of
DNMT3a overexpression or invalidation on the methylation status of
the miR-200c gene using bisulfite sequencing PCR. The primers used
for bisulfite sequencing PCR were as follows: BSP-miR-200c sense,
5′-TTTTAGTATTTATTTTTTGGGGGTAG-3′ and antisense,
5′-CACCTTAAATCAAACAACTTCAAAC-3′. The products of interest (375 bp)
were purified by the DNA Gel Extraction kit, ligated into the
Pmd19-T vector (Takara, Dalian, China) and transformed into
Escherichia coli DH5α competent cells. Insertion was
confirmed by restriction enzyme digestion, and 10 clones were
chosen for sequencing.
Immunohistochemistry
For immunohistochemistry, paraffin sections were
deparaffinized and incubated with a primary rabbit polyclonal
DNMT3a antibody (Santa Cruz Biotechnology) at a dilution of 1:150
in a 4°C moist chamber overnight. Immunohistochemistry was
independently evaluated by two researchers who were blinded to the
patient outcome. The evaluation was based on the staining
intensity. Staining intensity for DNMT3a was scored as 0
(negative); 1 (weak); 2 (moderate); and 3 (strong). The specimens
were divided into two groups according to their scores: 0 and 1
were considered as the low expression group; 2 and 3 were
considered as the high expression group. In the event of a
discrepancy in scoring, the slides were re-examined by both
pathologists under a microscope.
Western blotting
To isolate the proteins, cells collected by 6-well
plates were washed once in phosphate-buffered saline and lysed in
lysis buffer. Each protein sample (15 µg) was resolved by
sodium dodecylsulfate-polyacrylamide gel electrophoresis,
transferred onto a polyvinylidene difluoride membrane, and
incubated with a monoclonal antibody against DNMT3a (1:1,000)
(Santa Cruz Biotechnology) or GAPDH (1:10,000) (KangCheng Bio-tech,
Shanghai, China). The signals were detected by incubation with
secondary antibodies (1:20,000 for goat anti-rabbit IgG, and
1:10,000 for rabbit anti-mouse IgG; Southern Biotech) labeled with
the ECL detection system.
Cell proliferation, invasion and
wound-healing assays
Cell proliferation assays were performed using Cell
Counting Kit-8 (Beyotime, Hangzhou, China) after transfection of
the miR-200c mimics or DNMT3a siRNA. Absorbance was measured at 450
nm using a microplate reader. Cell invasion assays were assessed at
48 h after transfection using Matrigel Invasion Chambers (BD
Biosciences, Franklin Lakes, NJ, USA). For the wound-healing
assays, cell monolayers transfected with miR-200c mimics or DNMT3a
siRNA were scratched with a clean pipette tip, and cell migration
was observed for up to 48 h.
Statistical analysis
All experiments were conducted in triplicate.
Differences between groups (continuous variables) were analyzed by
the Student's t-test. The relationship between the expression of
miR-200c and clinicopathological factors was analyzed by Chi-square
tests. All statistical analyses were two-sided and were performed
using SPSS software (version 16.0; SPSS, Inc., Chicago, IL, USA),
and P<0.05 was considered to indicate a statistically
significant result.
Results
miR-200c is downregulated in GC
tissues
The miR-200 family can be grouped into two
subfamilies in two ways. The first way is functionally, according
to the presence of two types of seed sequences: miR-200a/miR-141
with a common sequence of AAcACUG, and miR-200b/miR-200c/miR-429
with AAuACUG. The second way is genetically, according to the
location in two gene clusters on two different chromosomes:
miR-200b/miR-200a/miR-429 on chr. 1, and miR-200c/miR-141 on chr.
12. In the present study, we performed most experiments with three
miR-200 family members: miR-200b, miR-200c and miR-141, since they
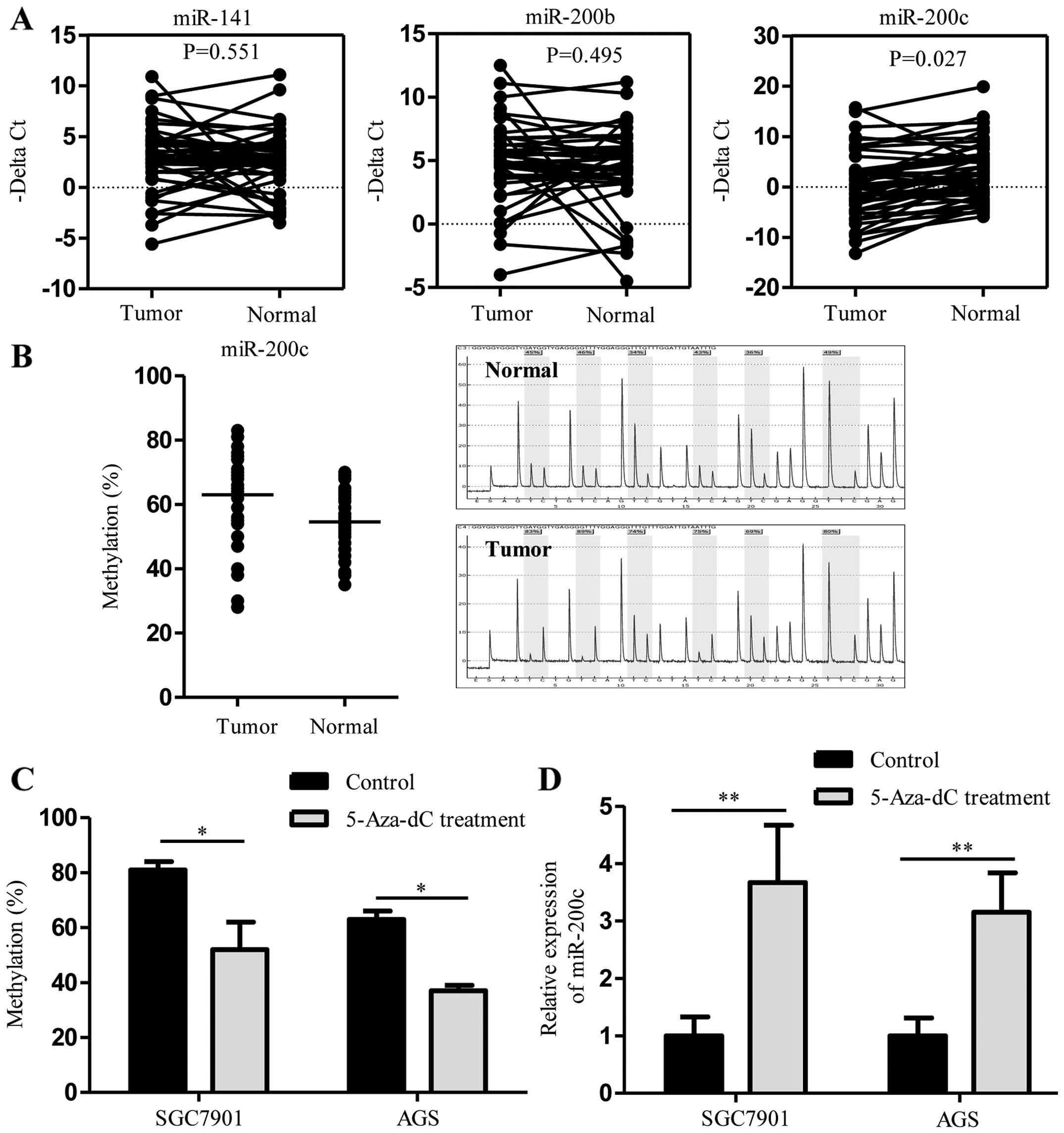
represent members of both subfamilies. As shown in Fig. 1A, we analyzed the expression levels
of all three miR-200 family members in 46 pairs of GC and their
corresponding non-tumor tissues. In qRT-PCR analysis, miR-200c was
significantly downregulated in the GC tissues compared with that
noted in the adjacent non-tumor tissues (P<0.05). There was no
significant difference in miR-200b (P=0.495) and miR-141 expression
levels (P=0.561) between the GC and non-tumor tissues. The
high-expression group was defined as cases with miR-200c expression
above the median value, while the remaining cases were included
into the low-expression group. We further compared the
clinicopathological parameters of GC according to their miR-200c
expression level. As shown in Table
I, the diffuse histologic type, deeper invasion and advanced
stage of GC were all associated with low expression of
miR-200c.
 | Table ImiR-200c expression and
clinicopathological characteristics of the gastric cancer
cases. |
Table I
miR-200c expression and
clinicopathological characteristics of the gastric cancer
cases.
| Characteristic | High expression
(n=23) | Low expression
(n=23) | P-value |
|---|
| Age, years (mean ±
SD) | 59.0±12.3 | 60.0±6.9 | 0.725 |
| Gender | | | 0.760 |
| Male | 15 | 14 | |
| Female | 8 | 9 | |
| Histological
type | | |
0.018a |
| Intestinal | 16 | 8 | |
| Diffuse | 7 | 15 | |
| Depth of
invasion | | |
0.010a |
| T1+T2 | 11 | 3 | |
| T3+T4 | 12 | 20 | |
| Lymph node
metastasis | | | 0.063 |
| Negative (N0) | 11 | 5 | |
| Positive
(N1-N3) | 12 | 18 | |
| Organ
metastasis | | | 0.710 |
| Negative (N0) | 19 | 18 | |
| Positive
(N1-N3) | 4 | 5 | |
| Stage | | |
0.038a |
| I+II | 14 | 7 | |
| III+IV | 9 | 16 | |
| Tumor site | | | 0.710 |
| Cardia cancer | 5 | 4 | |
| Non-cardia
cancer | 18 | 19 | |
Hypermethylation of the miR-200c CpG
island promoter causes downregulation of miR-200c
To determine the methylation status of the promoter
region in the miR-200c gene, we analyzed the methylation status at
six CpG dinucleotide sites of the miR-200c locus
(chr12:7072639-7072665) in 39 pairs of GC and matched adjacent
non-tumor tissues. Pyrosequencing analysis revealed that the
miR-200c promoter region was significantly hypermethylated in the
GC tissues when compared with that in the adjacent non-tumor
tissues (63.0% for GC tissues vs. 54.6% for non-tumor tissues;
P<0.01; Fig. 1B). In addition,
we treated GC cell lines with the demethylating agent, 5-aza-dC.
Pyrosequencing analysis showed that treatment with 5-aza-dC
decreased the methylation status of the miR-200c promoter region in
the SGC7901 cells (80.7% for untreated controls vs. 52.3% for
5-aza-dC; P<0.05) and in the AGS cells (63.0% for untreated
control vs. 37.3% for 5-aza-dC; P<0.05) (Fig. 1C). Demethylation of the miR-200c
promoter region resulted in the upregulation of miR-200c in the
SGC7901 and AGS cells as assayed by qRT-PCR (P<0.01 for all)
(Fig. 1D). Together, these data
indicate that DNA methylation may be involved in miR-200c
expression.
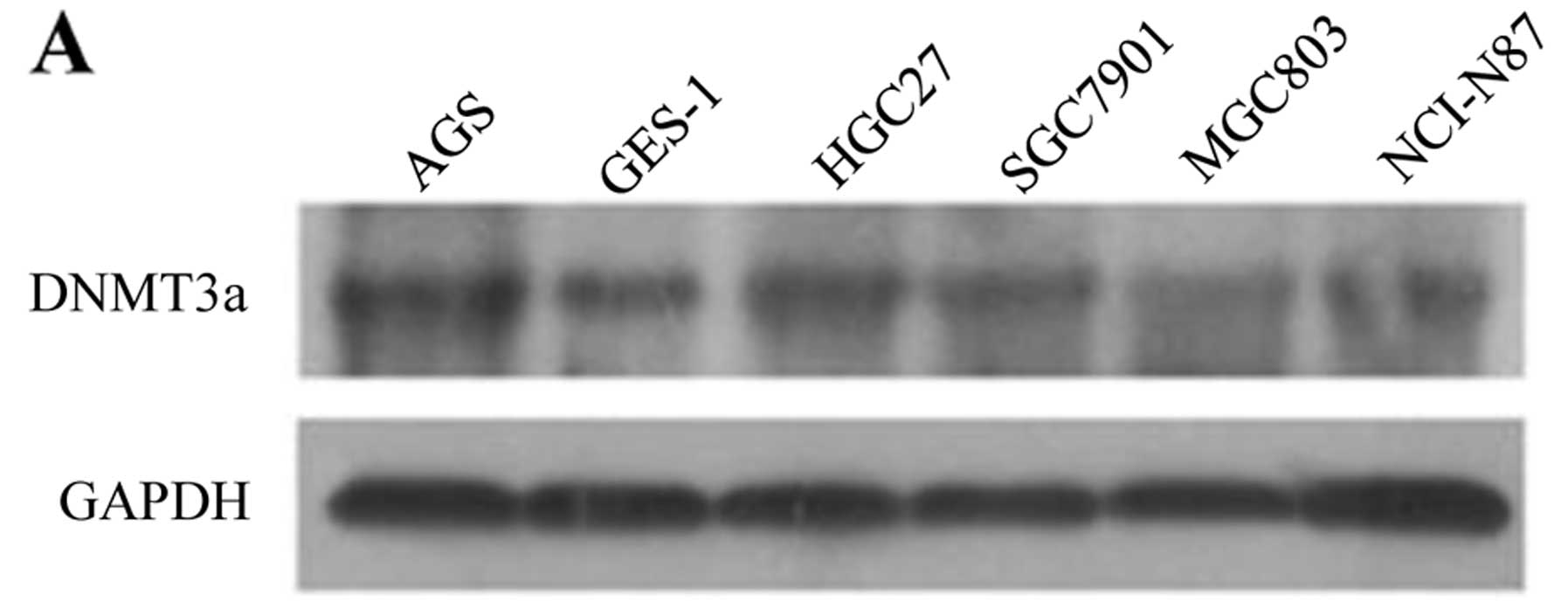
DNMT3a upregulation is responsible for
the hypermethylation of the miR-200c gene promoter
Previous studies have shown that DNMT3a
overexpression contributes to gene promoter hypermethylation and is
associated with the malignant potential and poor prognosis of human
cancer. In the present study, we found that the expression of
DNMT3a in five GC cell lines was higher than that in immortal
gastric epithelial GES-1 cells (Fig.
2A). Consistent with the western blot results, the expression
of DNMT3a was strongly increased in the GC tissues than that noted
in the non-tumor tissues by immunohistochemistry. In GC, expression
of DNMT3a was mainly observed in the nucleus. Weak staining was
also observed in the cytoplasm (Fig.
2B). Among the 46 paired samples, DNMT3a high expression was
found in 31/46 of the GC samples. This was significantly higher
compared to that noted in the paired control samples (67.4 vs.
34.8%; P<0.01).
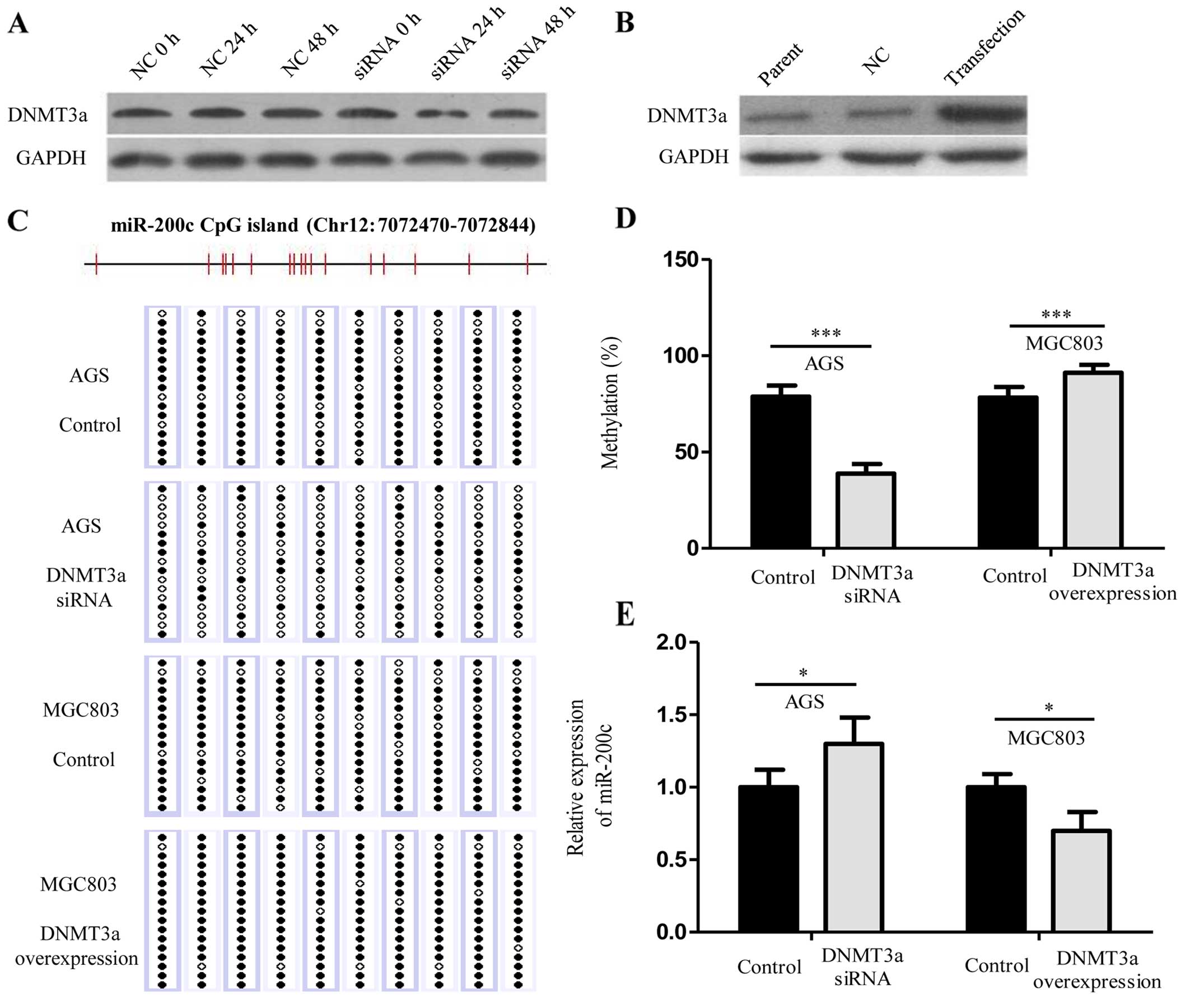
To further explore the role of DNMT3a in regulating
the expression of miR-200c, using siRNA against DNMT3a, we knocked
down de novo DNMT3a expression in the AGS cells which had
higher expression of DNMT3a when compared with the other GC cell
lines (Fig. 3A), and we enforced
expression of DNMT3a in the MGC803 cells which had lower expression
of DNMT3a when compared with the other GC cell lines (Fig. 3B). The results showed that DNMT3a
knockdown abolished the hypermethylation of the miR-200c gene
(78.9% for untreated controls vs. 38.8% for DNMT3a siRNA;
P<0.001) (Fig. 3C and D), and
induced upregulation of miR-200c expression (P<0.05) (Fig. 3E). Conversely, ectopic expression of
DNMT3a increased methylation levels in the miR-200c promoter (78.3%
for untreated controls vs. 91.2% for DNMT3a overexpression;
P<0.001) (Fig. 3C and D), and
decreased miR-200c expression (P<0.05) (Fig. 3E). Together, these data demonstrate
that DNMT3a regulates miR-200c expression via CpG island promoter
hypermethylation.
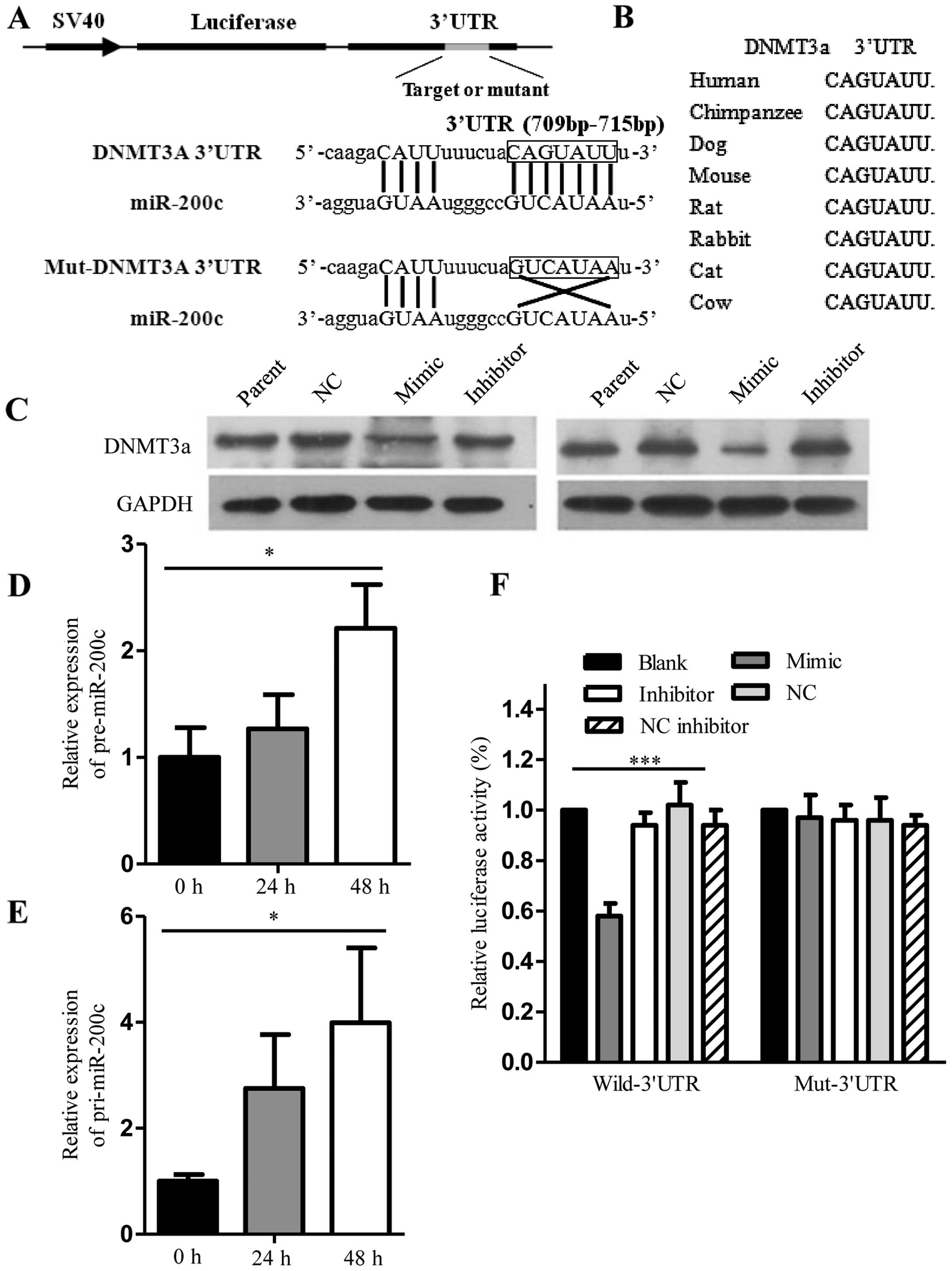
miR-200c directly targets DNMT3a and
induces endogenous pre-miR-200c and pri-miR-200c re-expression
As shown in Fig. 4A,
one potential miR-200c targeting site was found in the 3′UTR of
DNMT3a. Comparative sequence analysis showed that the target
sequences were evolutionarily conserved in different species
(Fig. 4B). Total RNA and protein
was isolated from the SGC7901 and AGS cells collected 48 h after
transfection. DNMT3a protein expression as demonstrated by western
blot analysis was decreased in the SGC7901 and AGS cells
transfected with miR-200c mimics (Fig.
4C). Additionally, pre-miR-200c and pri-miR-200c expression
demonstrated by qRT-PCR was increased in the AGS cells transfected
with the miR-200c mimics (Fig. 4D and
E). By means of a luciferase reporter assay in the SGC7901
cells, we found that the luminescence intensity was significantly
decreased in the DNMT3a 3′UTR reporter and miR-200c
mimic-transfected cells than the others, suggesting that miR-200c
has a target site in the 3′UTR of DNMT3a mRNA (Fig. 4F).
miR-200c mimics and DNMT3a siRNA suppress
cell growth and invasion
The effects of miR-200c upregulation and DNMT3a
downregulation on cell growth and invasion were determined via
several assays in vitro. Transfection of miR-200c mimics or
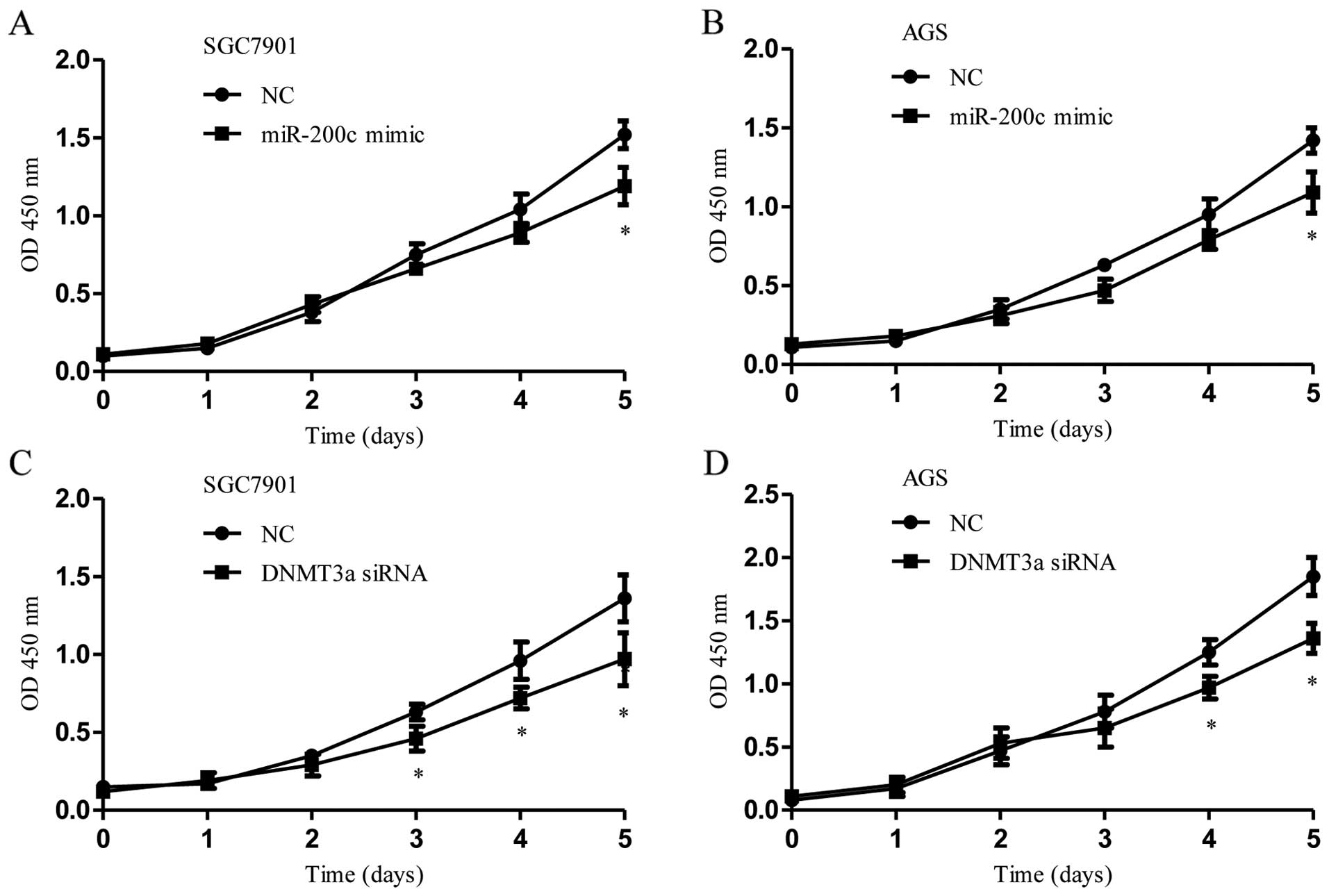
DNMT3a siRNA significantly reduced cell proliferation in the
SGC7901 and AGS cells compared to the proliferation noted in the
control transfections (Fig. 5A–D).
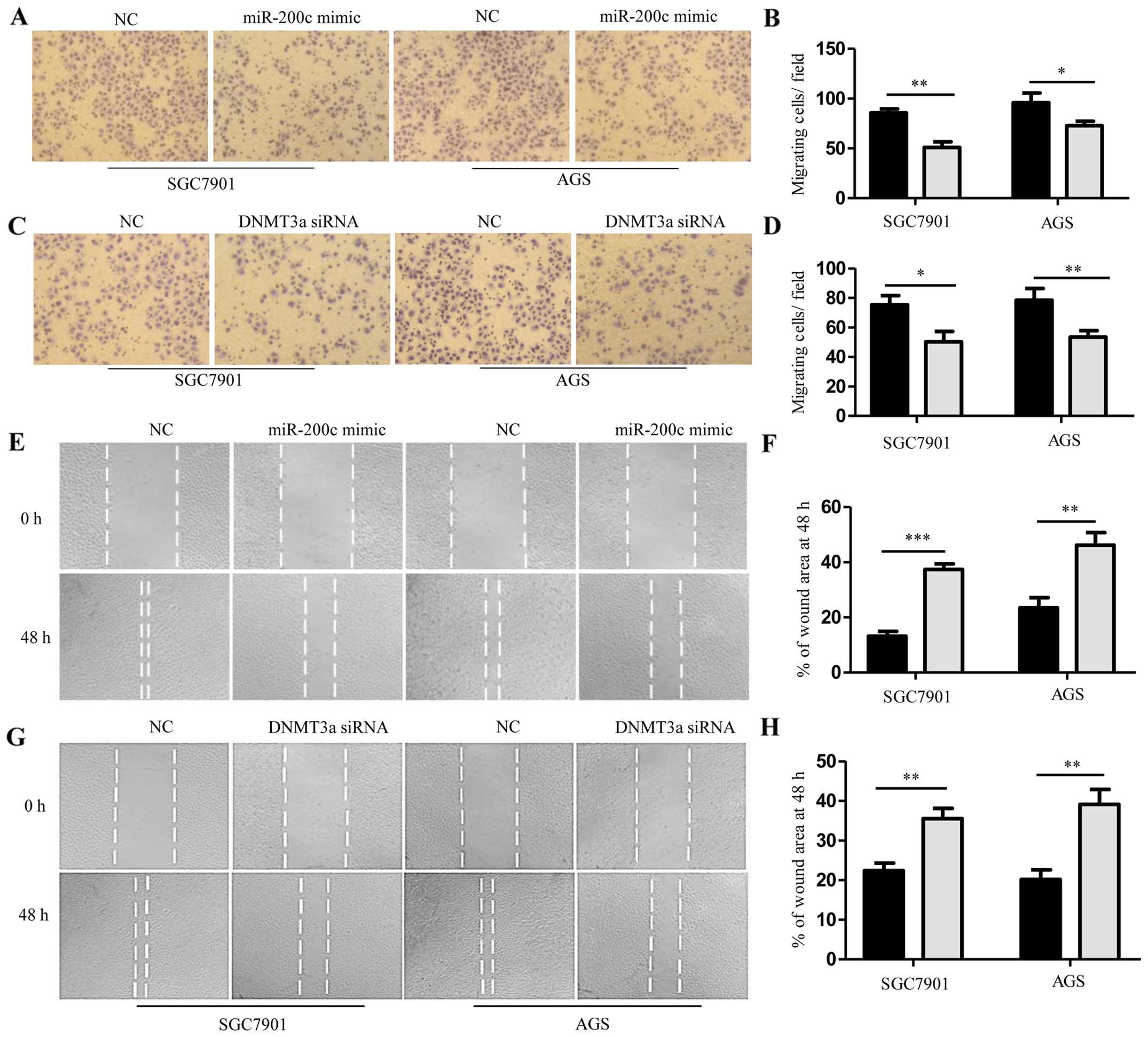
Upregulation of miR-200c or knockdown of DNMT3a in the SGC7901 and
AGS cells significantly inhibited cell invasion (Fig. 6A–D). The effects of miR-200c on cell
migration were determined by wound-healing assays. Both SGC7901 and
AGS cells treated with miR-200c mimics or DNMT3a siRNA were
distinctively less migratory than that noted in the negative
control cells (Fig. 6E–H). These
results indicate that upregulation of miR-200c or knockdown of
DNMT3a suppressed cell growth, migration and invasion of GC cells
in vitro.
Discussion
Gastric carcinogenesis is a multistep process
characterized by accumulation of molecular alterations (18). A complicated reciprocal repression
mechanism between miRNAs and epigenetic pathways appears to form an
epigenetic-miRNA network contributing to gastric carcinogenesis
(19). In the present study, we
found that miR-200c was downregulated in GC and was associated with
the histologic type in GC. Dysregulation of miR-200c in GC has also
been observed in other studies (20,21),
but little is known concerning its expression in diffuse-type GC.
This is the first study to show that miR-200c is downregulated in
diffuse-type GC.
It is well recognized that miR-200c regulates the
epithelial-mesenchymal transition (EMT) and cancer cell invasion
(22,23). Ectopic expression of miR-200 hinders
EMT by suppressing ZEB1 and ZEB2 expression and enhancing
E-cadherin expression (24,25). Notably, EMT is an important process
to form diffuse histology and initiate metastasis by enhancing the
motility of cancer cells (18).
Therefore, reduced expression of miR-200c may participate in the
genesis of diffuse histology of GC by reducing E-cadherin
expression through ZEB1 and ZEB2.
We next demonstrated that miR-200c expression is
regulated by DNA methylation. It is widely accepted that miRNAs
undergo the same regulatory mechanisms of the protein coding gene,
including methylation regulation. Generally, DNA methylation is
accomplished by three enzymes: DNMT1, DNMT3a and DNMT3b. DNMT1 is
responsible for maintaining preexisting methylation patterns during
DNA replication, whereas DNMT3a and DNMT3b are considered to be
de novo DNA methyltransferase (26). In breast, miR-200c was repressed by
DNA methylation, whereas the miR-200b-200a-429 cluster was silenced
primarily through polycomb group-mediated histone modifications
(27), suggesting that epigenetic
control of miRNAs may be miRNA-specific and epigenetic
effector-specific. The increased expression of DNMT3a may play an
important role in the malignant progression of cancer, leading to
aberrant methylation in many important tumor suppressor genes
(28). More recently, Cao et
al (29) reported that the
expression of DNMT3a is an independent poor prognostic indicator in
GC. Yang et al (30)
reported that overexpression of DNMT3a in GC tissues was observed
in 70.4% of 54 cases, and DNMT3a was associated with TNM stage and
lymph node metastasis. Aberrant expression of DNMT3a may cause
upregulation of global DNA methylation, decreasing expression of
tumor suppressor genes that may contribute to aggressive poorly
differentiated cancer. However, the mechanisms underlying aberrant
DNMT expression have not been fully elucidated.
Our results showed that miR-200c can target 3′UTR of
DNMT3a directly; in contrast, DNMT3a can establish and maintain
miR-200c methylation to suppress its expression. Thus, DNMT3a and
miR-200c form a negative feedback loop in GC cells, and this may
plays a key role in gastric carcinogenesis. Most investigations
which focused on the mutual regulation between DNA methylation and
miRNAs were frequently confirmed in fibrotic disease and cancer
research. For example, a mutual regulation of miRNAs and DNA
methylation may be involved in the pathogenesis of lung fibrosis
(31). A regulatory circuit of
miR-148a/152 and DNMT1 was found to modulate breast cancer cell
transformation and tumor angiogenesis through IGF-IR and IRS1
(9). Another study also
demonstrated that miR-152 and DNMT1 interacted with each other via
a feedback loop involved in NiS-induced malignant transformation
(32). In esophageal squamous cell
carcinoma, the DNMT1-miRNA-126 epigenetic circuit was contributed
to cancer growth via ADAM9/EGFR/AKT signaling (33). Thus, epigenetic-miRNA loops are
widely present in various diseases including cancers. Given that
both miRNA and epigenetic modulations strongly regulate the
expression of various disease-associated genes (34), controlling the dynamic balance of
epigenetic-miRNA loops may be of great clinical significance.
Regulation of DNMT3a by miR-200c has also been
described in GC by Tang et al (20), but the mutual regulation between
DNMT3a and miR-200c has not been confirmed in previous studies. The
results indicate that DNMT3a is a crucial regulator, as its
aberrant expression in cancer may start a self-enhancing feed
forward loop by downregulating its own inhibitor miR-200c. It has
been reported that a reciprocal repression between ZEB1 and the
miR-200 family promotes EMT and invasion in cancer cells (35), thus, it is plausible that aberrant
expression of DNMT3a may interact with the reciprocal repression
between ZEB1 and miR-200c, thereby inducing downregulation of
miR-200c and upregulation of ZEB1, subsequently promoting EMT and
invasion in cancer cells. Indeed, in vitro invasion and
migration assays implicate that the loop between miR-200c and
DNMT3a is involved in promoting EMT and metastasis of GC cells.
Future experiments are needed to determine whether both DNMT3a and
ZEB1 act separately or synergistically as a complex in this
setting.
In conclusion, miR-200c directly suppressed the
expression levels of DNMT3a. In contrast, overexpression of DNMT3a
was responsible for the promoter hypermethylation of miR-200c.
Downregulated expression of miR-200c further increased DNMT3a
expression by less targeting DNMT3a 3′UTR. A novel miR-200c-DNMT3a
regulatory circuit may exist in GC. The discovery of this
miRNA-epigenetic regulatory circuit would be highly beneficial for
deepening our understanding of gastric carcinogenesis.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (no. 81372633), the National
Natural Science Foundation of China (no. 81502040), the Projects of
Medical and Health Technology Program in Guangzhou City (no.
20161A010009), and the Key Programs of the Health Bureau of
Guangzhou City (20121A021004).
Abbreviations:
|
miRNAs
|
microRNAs
|
|
GC
|
gastric cancer
|
|
DNMT3a
|
DNA (cytosine-5-)-methyltransferase 3
α
|
|
EMT
|
epithelial-mesenchymal transition
|
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Lin S and Gregory RI: MicroRNA biogenesis
pathways in cancer. Nat Rev Cancer. 15:321–333. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ueda T, Volinia S, Okumura H, Shimizu M,
Taccioli C, Rossi S, Alder H, Liu CG, Oue N, Yasui W, et al:
Relation between microRNA expression and progression and prognosis
of gastric cancer: A microRNA expression analysis. Lancet Oncol.
11:136–146. 2010. View Article : Google Scholar
|
|
4
|
Song S and Ajani JA: The role of microRNAs
in cancers of the upper gastrointestinal tract. Nat Rev
Gastroenterol Hepatol. 10:109–118. 2013. View Article : Google Scholar
|
|
5
|
Ando T, Yoshida T, Enomoto S, Asada K,
Tatematsu M, Ichinose M, Sugiyama T and Ushijima T: DNA methylation
of microRNA genes in gastric mucosae of gastric cancer patients:
Its possible involvement in the formation of epigenetic field
defect. Int J Cancer. 124:2367–2374. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kiga K, Mimuro H, Suzuki M,
Shinozaki-Ushiku A, Kobayashi T, Sanada T, Kim M, Ogawa M, Iwasaki
YW, Kayo H, et al: Epigenetic silencing of miR-210 increases the
proliferation of gastric epithelium during chronic Helicobacter
pylori infection. Nat Commun. 5:44972014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Li Z, Lei H, Luo M, Wang Y, Dong L, Ma Y,
Liu C, Song W, Wang F, Zhang J, et al: DNA methylation
downregulated mir-10b acts as a tumor suppressor in gastric cancer.
Gastric Cancer. 18:43–54. 2015. View Article : Google Scholar
|
|
8
|
Morita S, Horii T, Kimura M, Ochiya T,
Tajima S and Hatada I: miR-29 represses the activities of DNA
methyltransferases and DNA demethylases. Int J Mol Sci.
14:14647–14658. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Xu Q, Jiang Y, Yin Y, Li Q, He J, Jing Y,
Qi YT, Xu Q, Li W, Lu B, et al: A regulatory circuit of
miR-148a/152 and DNMT1 in modulating cell transformation and tumor
angiogenesis through IGF-IR and IRS1. J Mol Cell Biol. 5:3–13.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Xiang Y, Ma N, Wang D, Zhang Y, Zhou J, Wu
G, Zhao R, Huang H, Wang X, Qiao Y, et al: MiR-152 and miR-185
co-contribute to ovarian cancer cells cisplatin sensitivity by
targeting DNMT1 directly: A novel epigenetic therapy independent of
decitabine. Oncogene. 33:378–386. 2014. View Article : Google Scholar
|
|
11
|
Huang J, Wang Y, Guo Y and Sun S:
Down-regulated microRNA-152 induces aberrant DNA methylation in
hepatitis B virus-related hepatocellular carcinoma by targeting DNA
meth-yltransferase 1. Hepatology. 52:60–70. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Song F, Yang D, Liu B, Guo Y, Zheng H, Li
L, Wang T, Yu J, Zhao Y, Niu R, et al: Integrated microRNA network
analyses identify a poor-prognosis subtype of gastric cancer
characterized by the miR-200 family. Clin Cancer Res. 20:878–889.
2014. View Article : Google Scholar
|
|
13
|
Kurashige J, Mima K, Sawada G, Takahashi
Y, Eguchi H, Sugimachi K, Mori M, Yanagihara K, Yashiro M, Hirakawa
K, et al: Epigenetic modulation and repression of miR-200b by
cancer-associated fibroblasts contribute to cancer invasion and
peritoneal dissemination in gastric cancer. Carcinogenesis.
36:133–141. 2015. View Article : Google Scholar
|
|
14
|
Ning X, Shi Z, Liu X, Zhang A, Han L,
Jiang K, Kang C and Zhang Q: DNMT1 and EZH2 mediated methylation
silences the microRNA-200b/a/429 gene and promotes tumor
progression. Cancer Lett. 359:198–205. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Davalos V, Moutinho C, Villanueva A, Boque
R, Silva P, Carneiro F and Esteller M: Dynamic epigenetic
regulation of the microRNA-200 family mediates epithelial and
mesenchymal transitions in human tumorigenesis. Oncogene.
31:2062–2074. 2012. View Article : Google Scholar :
|
|
16
|
Hur K, Toiyama Y, Takahashi M, Balaguer F,
Nagasaka T, Koike J, Hemmi H, Koi M, Boland CR and Goel A:
MicroRNA-200c modulates epithelial-to-mesenchymal transition (EMT)
in human colorectal cancer metastasis. Gut. 62:1315–1326. 2013.
View Article : Google Scholar :
|
|
17
|
Neves R, Scheel C, Weinhold S, Honisch E,
Iwaniuk KM, Trompeter HI, Niederacher D, Wernet P, Santourlidis S
and Uhrberg M: Role of DNA methylation in miR-200c/141 cluster
silencing in invasive breast cancer cells. BMC Res Notes.
3:2192010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yasui W, Sentani K, Sakamoto N, Anami K,
Naito Y and Oue N: Molecular pathology of gastric cancer: Research
and practice. Pathol Res Pract. 207:608–612. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Gigek CO, Chen ES, Calcagno DQ, Wisnieski
F, Burbano RR and Smith MA: Epigenetic mechanisms in gastric
cancer. Epigenomics. 4:279–294. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tang H, Deng M, Tang Y and Xie X, Guo J,
Kong Y, Ye F, Su Q and Xie X: miR-200b and miR-200c as prognostic
factors and mediators of gastric cancer cell progression. Clin
Cancer Res. 19:5602–5612. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhou X, Wang Y, Shan B, Han J, Zhu H, Lv
Y, Fan X, Sang M, Liu XD and Liu W: The downregulation of
miR-200c/141 promotes ZEB1/2 expression and gastric cancer
progression. Med Oncol. 32:4282015. View Article : Google Scholar
|
|
22
|
Mongroo PS and Rustgi AK: The role of the
miR-200 family in epithelial-mesenchymal transition. Cancer Biol
Ther. 10:219–222. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Feng X, Wang Z, Fillmore R and Xi Y:
MiR-200, a new star miRNA in human cancer. Cancer Lett.
344:166–173. 2014. View Article : Google Scholar :
|
|
24
|
Paterson EL, Kazenwadel J, Bert AG,
Khew-Goodall Y, Ruszkiewicz A and Goodall GJ: Down-regulation of
the miRNA-200 family at the invasive front of colorectal cancers
with degraded basement membrane indicates EMT is involved in cancer
progression. Neoplasia. 15:180–191. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ahn SM, Cha JY, Kim J, Kim D, Trang HT,
Kim YM, Cho YH, Park D and Hong S: Smad3 regulates E-cadherin via
miRNA-200 pathway. Oncogene. 31:3051–3059. 2012. View Article : Google Scholar
|
|
26
|
Jin B and Robertson KD: DNA
methyltransferases, DNA damage repair, and cancer. Adv Exp Med
Biol. 754:3–29. 2013. View Article : Google Scholar
|
|
27
|
Lim YY, Wright JA, Attema JL, Gregory PA,
Bert AG, Smith E, Thomas D, Lopez AF, Drew PA, Khew-Goodall Y, et
al: Epigenetic modulation of the miR-200 family is associated with
transition to a breast cancer stem-cell-like state. J Cell Sci.
126:2256–2266. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Daniel FI, Cherubini K, Yurgel LS, de
Figueiredo MA and Salum FG: The role of epigenetic transcription
repression and DNA methyltransferases in cancer. Cancer.
117:677–687. 2011. View Article : Google Scholar
|
|
29
|
Cao XY, Ma HX, Shang YH, Jin MS, Kong F,
Jia ZF, Cao DH, Wang YP, Suo J and Jiang J: DNA methyltransferase3a
expression is an independent poor prognostic indicator in gastric
cancer. World J Gastroenterol. 20:8201–8208. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Yang J, Wei X, Wu Q, Xu Z, Gu D, Jin Y,
Shen Y, Huang H, Fan H and Chen J: Clinical significance of the
expression of DNA methyltransferase proteins in gastric cancer. Mol
Med Rep. 4:1139–1143. 2011.PubMed/NCBI
|
|
31
|
Dakhlallah D, Batte K, Wang Y,
Cantemir-Stone CZ, Yan P, Nuovo G, Mikhail A, Hitchcock CL, Wright
VP, Nana-Sinkam SP, et al: Epigenetic regulation of miR-17~92
contributes to the pathogenesis of pulmonary fibrosis. Am J Respir
Crit Care Med. 187:397–405. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ji W, Yang L, Yuan J, Yang L, Zhang M, Qi
D, Duan X, Xuan A, Zhang W, Lu J, et al: MicroRNA-152 targets DNA
methyltrans-ferase 1 in NiS-transformed cells via a feedback
mechanism. Carcinogenesis. 34:446–453. 2013. View Article : Google Scholar
|
|
33
|
Liu R, Gu J, Jiang P, Zheng Y, Liu X,
Jiang X, Huang E, Xiong S, Xu F, Liu G, et al: DNMT1-microRNA126
epigenetic circuit contributes to esophageal squamous cell
carcinoma growth via ADAM9-EGFR-AKT signaling. Clin Cancer Res.
21:854–863. 2015. View Article : Google Scholar
|
|
34
|
Jones PA and Baylin SB: The fundamental
role of epigenetic events in cancer. Nat Rev Genet. 3:415–428.
2002.PubMed/NCBI
|
|
35
|
Burk U, Schubert J, Wellner U, Schmalhofer
O, Vincan E, Spaderna S and Brabletz T: A reciprocal repression
between ZEB1 and members of the miR-200 family promotes EMT and
invasion in cancer cells. EMBO Rep. 9:582–589. 2008. View Article : Google Scholar : PubMed/NCBI
|