Introduction
Ovarian cancer is the second leading gynecological
malignancy after cervical cancer. However, it is the most lethal
gynecological malignancy, accounting for more deaths than
endometrial and cervical cancers combined (1). Ovarian cancer is typically
asymptomatic during the early stages, and most ovarian cancers are
diagnosed at an advanced III or IV stage due to its deep pelvic
location, multiple morphologies and characteristic genetic factors
(2). The current standard of care
includes surgical cytoreduction and platinum-based chemotherapy.
Although cytoreductive surgery with chemotherapy can significantly
prolong patient survival, most cancers eventually relapse and
metastasize, becoming resistant and refractory to standard
chemotherapy; the 5-year survival rate is ~30% (3).
Growing evidence suggests that microRNAs (miRNAs)
play an important role in tumor development, progression and
metastasis and may offer a promising therapeutic strategy to
improve the management of cancer patients. miRNAs are a class of
small non-coding RNAs (18–22 nt in length) which regulate the
expression of target genes by binding to the 3′-untranslated
regions (3′-UTRs) resulting in the degradation of mRNAs or the
translational inhibition of functional proteins (4). Emerging evidence strongly suggests the
critical roles of miRNAs in the pathogenesis of ovarian cancer,
including miR-106a. To date, miR-106a has been found to be
upregulated or downregulated in many types of cancers, such as
renal carcinoma (5), colon cancer
(6), esophageal carcinoma (7), gastric (8) and lung cancer (9). However, the mechanism of miR-106a in
ovarian cancer is not clear, thus, we aimed to reveal the role of
miR-106a in ovarian cancer.
Recent convincing data support the involvement of
the inflammatory stromal microenvironment, caused by overexpression
of cytokines or chemokines, in promoting ovarian tumorigenesis,
cancer progression and resistance to chemotherapies (10). Among the cytokines reported to date,
interleukin-6 (IL-6) is one of the pivotal immunoregulatory
cytokines present in the ovarian cancer microenvironment; it
induces several pathways leading to tumor proliferation,
angiogenesis and chemoresistance (11). Higher serum and ascites levels of
IL-6 have been found in patients with ovarian cancer than levels in
patients with other malignancies, and levels have been shown to
correlate with the extent of disease and poor clinical outcome
(12–14). Tocilizumab is a humanized anti-human
IL-6R antibody and binds to the IL-6-binding site of human IL-6R.
It is known to competitively inhibit IL-6/IL-6R signaling and
completely neutralizes IL-6 activities (15,16). A
series of clinical studies has successfully shown that the
suppression of IL-6/IL-6R signaling by tocilizumab is
therapeutically effective in alleviating Castleman's disease and
rheumatoid arthritis (17,18).
Hence, the present study was aimed to identify the
role of miR-106a in ovarian cancer. miR-106a expression was
analyzed in ovarian cancer tissues and cell lines. After that, we
investigated the effects of miR-106a and tocilizumab on ovarian
cancer cell proliferation, migration and invasion and further
discuss the mechanisms of action of miR-106a by identifying its
potential target gene.
Materials and methods
Antibodies and reagents
The antibodies for STAT3, and phospho-STAT3 were
purchased from Abcam (Shanghai, China). The antibodies for
phosphatase and tensin homolog (PTEN), N-cadherin, E-cadherin,
SOCS6, vimentin and α-tubulin were purchased from Proteintech
(Wuhan, China). RIPA lysis buffer was purchased from CW Biotech
(Beijing, China). Tocilizumab was purchased from Chugai
Pharmaceutical (Shizuoka, Japan).
Clinical sample collection
Paired ovarian cancer tissues and normal
contralateral ovary tissues were obtained from 15 patients who
underwent primary surgical resection for ovarian cancer at Shandong
Cancer Hospital affiliated to Shangdong University (Shandong,
China). None of the patients had received pre-operative adjuvant
therapy. These samples were snap-frozen in liquid nitrogen after
resection. Prior patient consent and approval from the Ethics
Committee of Shandong Cancer Hospital were obtained for the use of
these clinical materials for research purposes.
Cell culture
SKOV3 and OVCAR3 cell lines were routinely
maintained in Dulbecco's modified Eagle's medium (DMEM) (HyClone,
Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS)
(CLARK), 100 U/ml penicillin sodium and 100 mg/ml streptomycin
sulfate (Solarbio, Beijing, China) at 37°C in a humidified air
atmosphere containing 5% CO2. Cells were used in the
logarithmic growth phase.
Transfection of the miR-106a
inhibitor
The inhibitor of miR-106a was purchased from RiboBio
(Guangzhou, China). Transfection was performed when cells were
grown to 80% confluency, using the NanoFectin Transfection Reagent
(ExCell Biology, Shanghai, China) according to the manufacturer's
instructions.
RNA isolation and quantitative real-time
PCR
Total RNA and miRNA were isolated using the
Ultrapure RNA kit (CWBio, Beijing, China) according to the
manufacturer's protocol. cDNA was reverse transcribed from total
RNA samples using the miRNA cDNA kit (CWBio). miR-106a expression
was detected by quantitative real-time PCR (qRT-PCR) using the
miRNA real-time PCR assay kit. The small nuclear RNA U6 was used
for normalization. The relative amount of miR-106a was calculated
using the cycle threshold (CT) value as the relative miRNA level.
Each sample was performed in triplicate. The miR-106a and U6
primers were synthesized by Genewiz (Beijing, China). The following
primers were used: miR-106a-F, AAA AGT GCT TAC AGT GCA GGT AG; and
human U6-F, CTC GCT TCG GCA GCA CA.
Western blotting
Cells were harvested and lysed in cold RIPA lysis
buffer containing 1% Halt Protease (CWBio) for 30 min. The
supernatant was collected after 10 min of centrifugation at 12,000
rpm; the protein concentration of which was measured using the
bicinchoninic acid method, then denatured with sample loading
buffer for 5 min at 95°C and stored at −20°C for future use. Equal
amounts of proteins were separated by 10% SDS-polyacrylamide gel
electrophoresis and transferred onto polyvinylidene fluoride
membranes. After blocking with 5% skim milk for 1 h, the membranes
were incubated with primary antibodies overnight at 4°C followed by
secondary antibodies for 1 h at room temperature. The bands were
subsequently detected by an enhanced chemiluminescence system (EMD
Millipore, Billerica, MA, USA) and analyzed by Quantity One
software.
Proliferation, invasion and migration
assays
The cells were cultured in a 6-well plate. When the
cells were grown to 80% confluency, the cells were transfected with
160 pmol of miR-106a, negative control (NC) or treated with
tocilizumab (Chugai Pharmaceutical, Shizuoka, Japan) (10
µg/ml) for 48 h.
For the Cell Counting kit-8 (CCK-8) assay, the cells
(1,000/well) were seeded into a 96-well plate in 100 ml complete
DMEM supplemented with 10% FBS, and cell growth was monitored at
indicated time points using the CCK-8 assay.
For the cell Matrigel Transwell invasion assays,
24-well Transwell containing polycarbonate filters with 8-mm pores
(Corning Costar, Corning, NY, USA) and the inserts were precoated
with 50 µl Matrigel matrix (dilution at 1:3; BD Biosciences,
Franklin Lakes, NJ, USA) according to the manufacturer's protocol,
and 200 ml cell suspension (2×105) obtained from the
primary step in serum-free medium was placed into the upper
chamber. The lower chamber was filled with 600 ml 10% FBS-DMEM. The
plates were subsequently incubated for 24 h under normal
conditions. The cells, which had invaded the lower surface of the
membrane, were fixed and stained with 0.1% crystal violet, The
number of invading and migrating cells was calculated using a
microscope at a magnification of ×200 in 5 random fields. Three
independent experiments were performed.
For the wound-healing assay, the cells obtained from
primary step one were seeded in a 6-well plate. When the cells grew
to a confluency of 90–95%, the cell monolayer was scratched using a
sterile 200-µl pipette tip. After washing and removal of the
detached cells, the plates were incubated at 37°C with FBS-free
DMEM and the wounds were photographed every 24 h. At least 5
different wounds were performed, and the experiments were
independently repeated 3 times.
Statistical analysis
Statistical testing was conducted with the
assistance of SPSS 17.0 software. All data are expressed as means ±
SD. Student's t-test and one-way analysis of variance (ANOVA) were
used to analyze data. Results were considered significant at a
P-value <0.05.
Results
miR-106a is upregulated in ovarian cancer
samples and cell lines
In the present study, the levels of miR-106a were
measured by quantitative real-time PCR in normal ovarian tissues
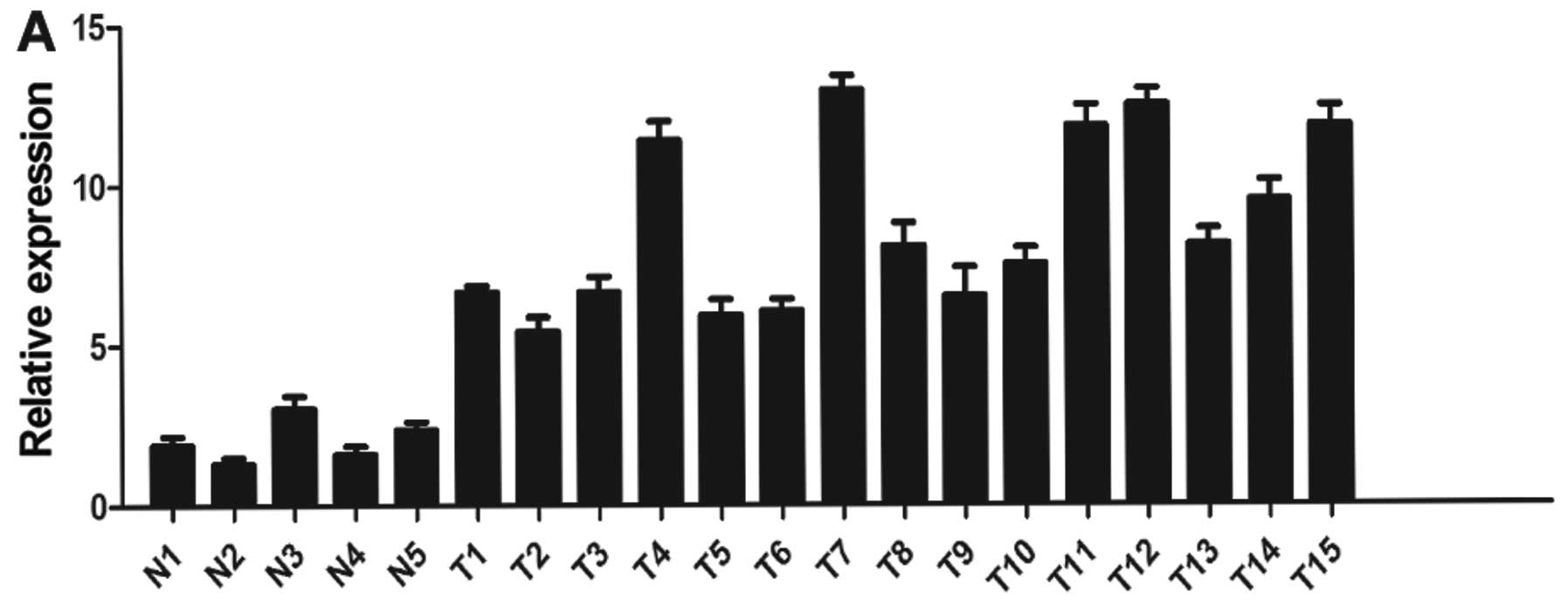
and primary ovarian cancer samples. As shown in Fig. 1A, we found that miR-106a expression
was significantly increased in the primary ovarian cancer tissues
compared with that noted in the normal tissues (P<0.05). In
addition, miR-106a expression levels were high in two ovarian
cancer cell lines compared with the level in the normal ovarian
tissues (Fig. 1A). We used SKOV3
cells in the cell function assay, due to higher IL-6 and miR-106a
expression levels (Fig. 1B). These
findings suggest that upregulation of miR-106a may play a role in
ovarian cancer development.
miR-106a inhibitor inhibits ovarian
cancer cell proliferation and invasion
To investigate the potential biological function of
miR-106a in ovarian cancer, we modulated the miR-106a expression by
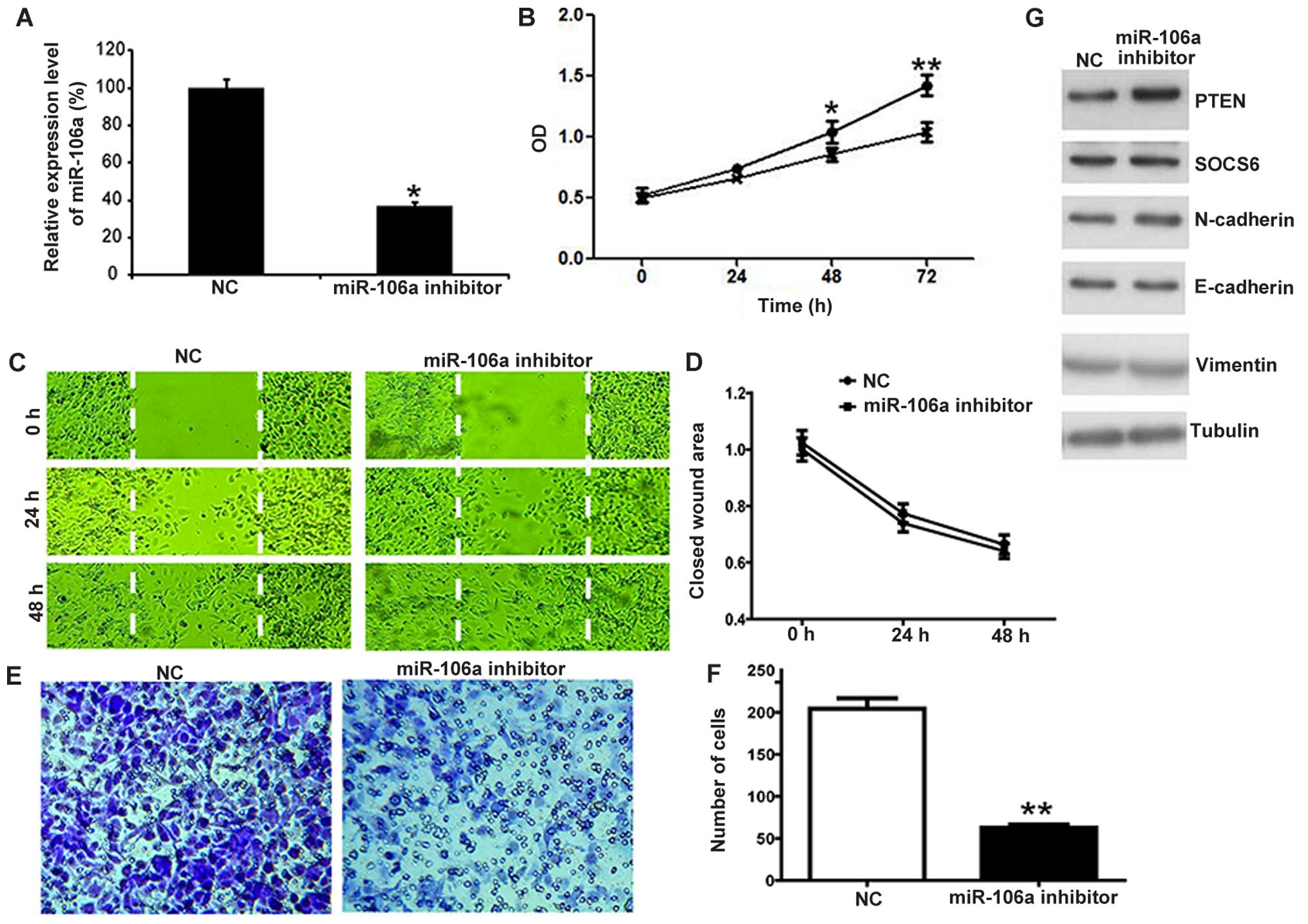
transfection with the miR-106a inhibitor. Using qPCR, we found that
the expression of miR-106a was significantly decreased in the cells
transfected with the miR-106a inhibitor compared with that noted in
the control group (NC) (Fig. 2A;
P<0.05). In order to observe the impact of miR-106a on the cell
proliferation of SKOV3 cells, the proliferation rates of SKOV3
cells treated with miR-106a inhibitor were determined by CCK-8
assay. As shown in Fig. 2B, growth
inhibition was noted when the cells were transfected with the
miR-106a inhibitor. To test the effect of miR-106a on the motility
of SKOV3 cells, in vitro migration and invasion assays were
performed. We assessed the effect of miR-106b on the migratory
capacity of SKOV3 cells using a wound-healing assay. As shown in
Fig. 2C and D, the miR-106a
inhibitor had no effect on SKOV3 cell migration. In contrast,
miR-106a inhibitor transfection suppressed SKOV3 cell invasion as
detected by the Matrigel invasion assay (Fig. 2E and F). These observations revealed
that miR-106a significantly promoted the proliferation and invasion
of SKOV3 cells. To investigate the molecular mechanism by which
miR-106a suppresses the growth and invasion of SKOV3 cells, we
detected putative target genes of miR-106a by western blotting.
Among the candidates, PTEN was found to be regulated by miR-106a
(Fig. 2G). On the contrary, SOCS6,
N-cadherin, E-cadherin and vimentin were not significantly affected
upon miR-106a inhibitor transfection (Fig. 2G). These findings suggest that
miR-106a may promote the growth and invasion of SKOV3 cells by
upregulating PTEN.
miR-106a is regulated by IL-6 in SKOV3
cells
Furthermore, we investigated the signaling that
triggers the upregulation of miR-106a. Interleukin-6 (IL-6) is one
of the important immunoregulatory cytokines present in the ovarian
cancer microenvironment (11);
IL6-R expression is highly expressed in ovarian cancer tissues
compared with levels in normal tissues or benign diseases, and the
IL-6 receptor pathway is believed to be a new therapeutic target in
ovarian cancer (11,19). Tocilizumab is a humanized anti-human
IL-6R antibody and competitively inhibits IL-6/IL-6R signaling
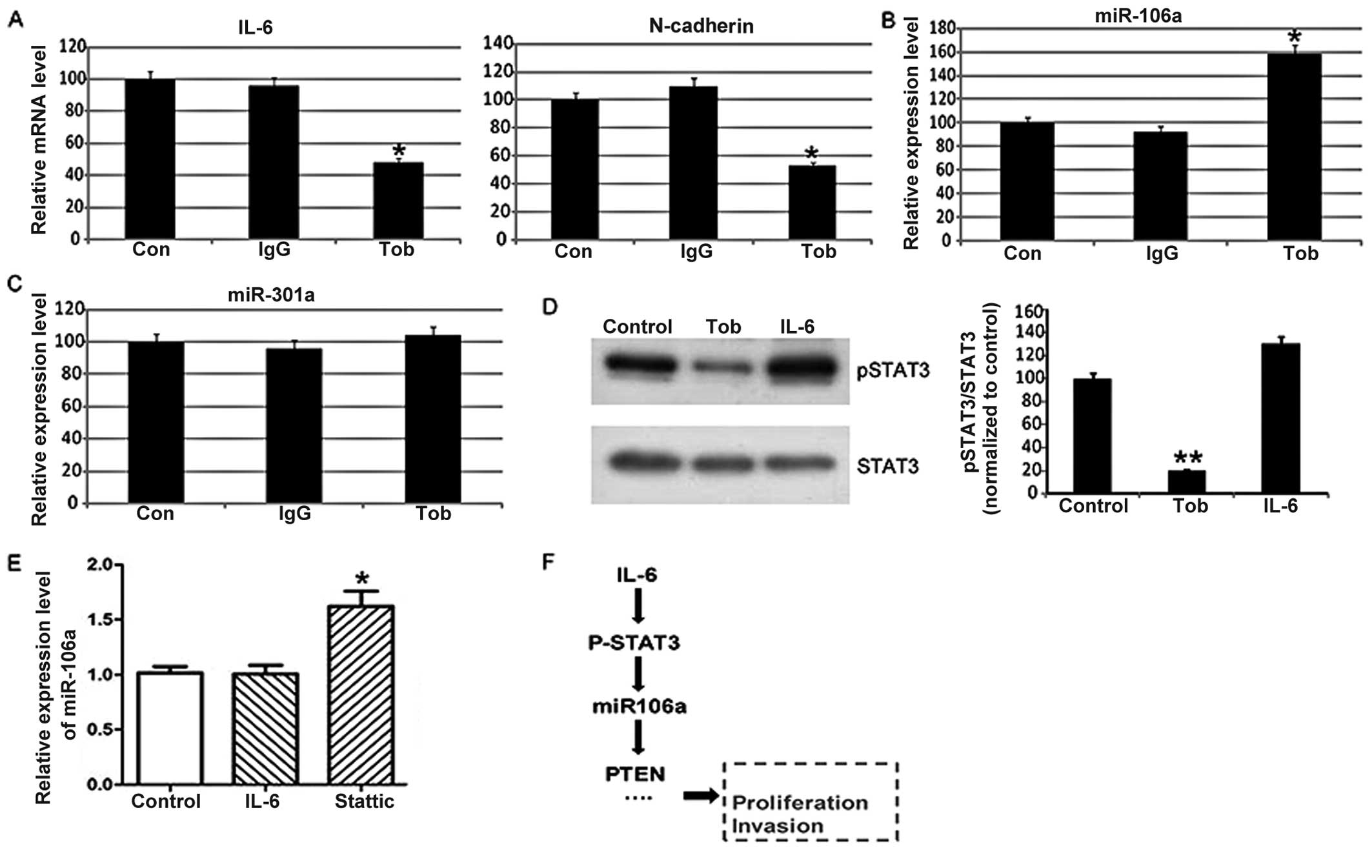
(19). As shown in Fig. 3A, tocilizumab decreased the mRNA
level of IL-6 and its downstream target-N-cadherin herein. Notably,
we found that tocilizumab significantly enhanced the expression of
miR-106a (Fig. 3B), while the
expression level of miR-301a was not affected (Fig. 3C). These findings suggest that IL-6
is a regulator of miR-106a in SKOV3 cells.
IL-6 inhibits miR-106a expression by
activating STAT3
To investigate the molecular mechanism by which IL-6
suppresses miR-106a expression, we detected the activation level of
STAT3, a target gene of IL-6. We found that tocilizumab
significantly decreased the phosphoryation level of STAT3 (Fig. 3D). Furthermore, we found that STAT3
inhibitor Stattic increased the expression level of miR-106a
(Fig. 3E). Thus, the present study
indicated that IL-6 is a regulator of miR-106a. IL-6 inhibits
miR-106a expression by activating STAT3 in SKOV3 cells. Then,
miR-106a may downregulate PTEN, and promote the growth and invasion
of SKOV3 cells.
Tocilizumab does not inhibit the
proliferation and invasion of SKOV3 cells
Tocilizumab has proven useful in treating
IL-6-related cancers (20–25). We aimed to ascertain whether
tocilizumab increases miR-106a expression, thus it may not be
effective in ovarian cancers. Thus, we detected the impact of
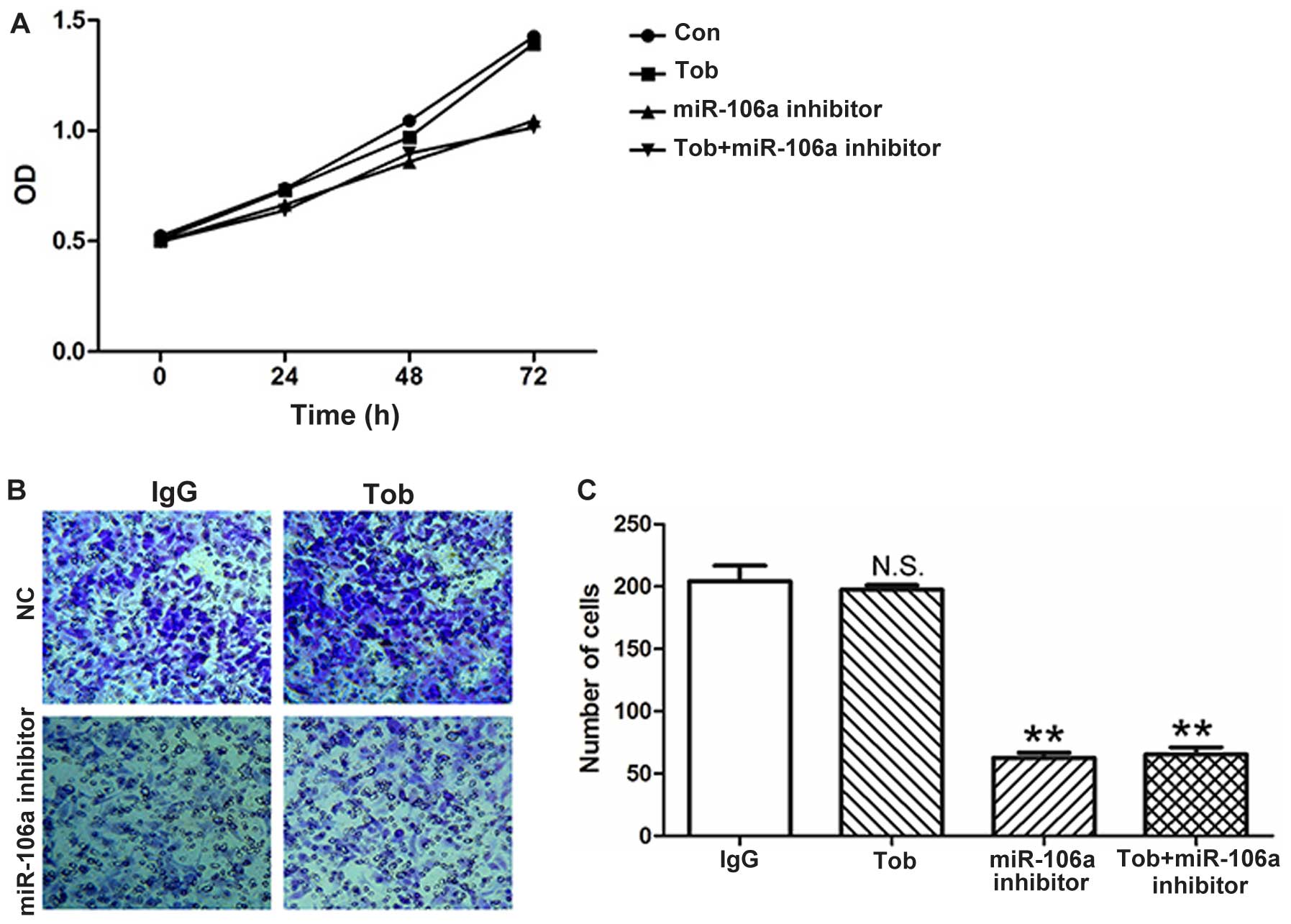
miR-106a on the cell proliferation and invasion of SKOV3 cells. As
shown in Fig. 4A, tocilizumab did
not inhibit the cell proliferation in SKOV3 cells. Co-treatment
with the miR-106a inhibitor slowed down the cell proliferation. In
addition, tocilizumab did not affect the invasion of the SKOV3
cells (Fig. 4B and C). The results
suggest that tocilizumab may increase miR-106a expression, and the
overexpression of miR-106a may impair the effect of tocilizumab on
ovarian cancer cells.
Discussion
Ovarian cancer is a leading cause of malignant
gynecological tumor-related deaths among women. The treatment of
ovarian cancer patients continues to be challenging. MicroRNAs
(miRNAs) have recently been described as important players in human
cancer, and some are believed to be potential therapeutic targets.
Expression of microRNAs is markedly deregulated in ovarian cancer,
which strongly suggests that miRNAs are involved in the
pathogenesis of ovarian cancer (26–42).
In the present study, we identified the deregulation of miR-106a in
ovarian cancer.
miR-106a is widely expressed in diverse human
tumors, including gastric, non-small cell lung, pancreatic,
colorectal and ovarian cancer (6,8,9,43–53).
Previous studies showed that miR-106a acts as a tumor suppressor or
an oncogene in different cancers, depending on the different
cellular context. In gastric cancer, expression of miR-106a was
increased, and downregulation of the expression of miR-106a
inhibited gastric cancer cell proliferation and caused apoptosis by
targeting FAS (52). In non-small
cell lung cancers, miR-106a inhibited the growth and metastasis of
NSCLC cells by decreasing phosphatase and tensin homolog (PTEN)
expression (9). In ovarian cancer,
upregulation of miR-106a was found to be associated with paclitaxel
and cisplatin resistance (47,50).
In the present study, we investigated the biological
and phthological roles of miR-106a in ovarian cancers. We found
that miR-106a expression was significantly increased in primary
ovarian cancer tissues and ovarian cancer cells compared with
levels noted in normal tissues. Ectopic expression of the miR-106a
inhibitor attenuated ovarian cancer cell line proliferation and
invasion. These results demonstrated that miR-106a can promote
ovarian cancer progression. A recent study reported that miR-106a
promotes the growth and metastasis of non-small cell lung cancer by
targeting PTEN (9). In line with
this finding, we found that PTEN is also a direct target of
miR-106a, and overexpression of miR-106a suppressed PTEN
expression.
In addition, at the molecular level, the present
study indicated that IL-6 is a regulator of miR-106a. IL-6 inhibits
miR-106a expression by activating STAT3. Interleukin-6 (IL-6) is
one of the important immunoregulatory cytokines present in the
ovarian cancer microenvironment (11); IL6-R expression is highly expressed
in ovarian cancer tissues compared with that noted in normal
tissues or benign diseases and the IL-6 receptor pathway is
believed to be a new therapeutic target in ovarian cancer (11,19).
Tocilizumab is a humanized anti-human IL-6R antibody and binds to
the IL-6-binding site of human IL-6R which competitively inhibits
IL-6/IL-6R signaling (19).
Tocilizumab has proven useful for treating IL-6-related cancers
(20–25). The therapeutic potential of
tocilizumab against ovarian cancer is still not well investigated.
Our findings indicated that tocilizumab did not inhibit the
proliferation and invasion of SKOV3 cells. This may be due to the
upregulation of miR-106a which was triggered by IL-6 activity
inhibition. Thus, the miR-106a level must be considered before
tocilizumab treatment.
In conclusion, the present study revealed that
miR-106a was significantly increased in ovarian cancer tissues and
cell lines. Downregulation of miR-106a expression inhibited cell
growth and metastasis of ovarian cancer cells by increasing PTEN
expression. IL-6 is a regulator of miR-106a by activating STAT3.
Together, the present study suggests that miR-106a acts as an
oncogene in ovarian cancers.
Acknowledgments
This study was supported by the Natural Science
Foundation of Shandong Province (grant no. ZR2015YL047), Projects
of Medical and Health Technology Development Program in Shandong
Province (grant no. 2015WS0157) and the Science Foundation of
Shandong Academy of Medical Sciences (grant no. 2014-50).
References
|
1
|
Siegel R, Naishadham D and Jemal A: Cancer
statistics, 2013. CA Cancer J Clin. 63:11–30. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Gao L, Ye X, Ma RQ, Cheng HY, Han HJ, Cui
H, Wei LH and Chang XH: Low programmed cell death 5 expression is a
prognostic factor in ovarian cancer. Chin Med J. 128:1084–1090.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kim MK, James J and Annunziata CM:
Topotecan synergizes with CHEK1 (CHK1) inhibitor to induce
apoptosis in ovarian cancer cells. BMC Cancer. 15:1962015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ma R, Jiang T and Kang X: Circulating
microRNAs in cancer: Origin, function and application. J Exp Clin
Cancer Res. 31:382012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ma Y, Zhang H, He X, Song H, Qiang Y, Li
Y, Gao J and Wang Z: miR-106a* inhibits the proliferation of renal
carcinoma cells by targeting IRS-2. Tumour Biol. 36:8389–8398.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yue B, Sun B, Liu C, Zhao S, Zhang D, Yu F
and Yan D: Long non-coding RNA Fer-1-like protein 4 suppresses
oncogenesis and exhibits prognostic value by associating with
miR-106a-5p in colon cancer. Cancer Sci. 106:1323–1332. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ma HL, Wen XP, Zhang XZ, Wang XL, Zhao DL,
Che SM and Dang CX: miR-106a* inhibits the proliferation of
esophageal carcinoma cells by targeting CDK2-associated Cullin 1
(CACUL1). Cell Mol Biol. 61:56–62. 2015.
|
|
8
|
Yuan R, Wang G, Zhi Q, Chen H, Han Y, Wang
B, Kou Z, Hu H, Guo Z, Xue X, et al: Up-regulated circulating
miR-106a by DNA methylation promised a potential diagnostic and
prognostic marker for gastric cancer. Anticancer Agents Med Chem.
15:12015.
|
|
9
|
Xie X, Liu HT, Mei J, Ding FB, Xiao HB, Hu
FQ, Hu R and Wang MS: miR-106a promotes growth and metastasis of
non-small cell lung cancer by targeting PTEN. Int J Clin Exp
Pathol. 8:3827–3834. 2015.PubMed/NCBI
|
|
10
|
Macciò A and Madeddu C: Inflammation and
ovarian cancer. Cytokine. 58:133–147. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dijkgraaf EM, Welters MJ, Nortier JW, van
der Burg SH and Kroep JR: Interleukin-6/interleukin-6 receptor
pathway as a new therapy target in epithelial ovarian cancer. Curr
Pharm Des. 18:3816–3827. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Plante M, Rubin SC, Wong GY, Federici MG,
Finstad CL and Gastl GA: Interleukin-6 level in serum and ascites
as a prognostic factor in patients with epithelial ovarian cancer.
Cancer. 73:1882–1888. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Scambia G, Testa U, Benedetti Panici P,
Foti E, Martucci R, Gadducci A, Perillo A, Facchini V, Peschle C
and Mancuso S: Prognostic significance of interleukin 6 serum
levels in patients with ovarian cancer. Br J Cancer. 71:354–356.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Duan Z, Foster R, Bell DA, Mahoney J,
Wolak K, Vaidya A, Hampel C, Lee H and Seiden MV: Signal
transducers and activators of transcription 3 pathway activation in
drug-resistant ovarian cancer. Clin Cancer Res. 12:5055–5063. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mihara M, Kasutani K, Okazaki M, Nakamura
A, Kawai S, Sugimoto M, Matsumoto Y and Ohsugi Y: Tocilizumab
inhibits signal transduction mediated by both mIL-6R and sIL-6R,
but not by the receptors of other members of IL-6 cytokine family.
Int Immunopharmacol. 5:1731–1740. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shinriki S, Jono H, Ota K, Ueda M, Kudo M,
Ota T, Oike Y, Endo M, Ibusuki M, Hiraki A, et al: Humanized
anti-interleukin-6 receptor antibody suppresses tumor angiogenesis
and in vivo growth of human oral squamous cell carcinoma. Clin
Cancer Res. 15:5426–5434. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nishimoto N, Kanakura Y, Aozasa K, Johkoh
T, Nakamura M, Nakano S, Nakano N, Ikeda Y, Sasaki T, Nishioka K,
et al: Humanized anti-interleukin-6 receptor antibody treatment of
multicentric Castleman disease. Blood. 106:2627–2632. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Nishimoto N, Hashimoto J, Miyasaka N,
Yamamoto K, Kawai S, Takeuchi T, Murata N, van der Heijde D and
Kishimoto T: Study of active controlled monotherapy used for
rheumatoid arthritis, an IL-6 inhibitor (SAMURAI): Evidence of
clinical and radiographic benefit from an x ray reader-blinded
randomised controlled trial of tocilizumab. Ann Rheum Dis.
66:1162–1167. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Isobe A, Sawada K, Kinose Y, Ohyagi-Hara
C, Nakatsuka E, Makino H, Ogura T, Mizuno T, Suzuki N, Morii E, et
al: Interleukin 6 receptor is an independent prognostic factor and
a potential therapeutic target of ovarian cancer. PLoS One.
10:e01180802015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ando K, Takahashi F, Kato M, Kaneko N, Doi
T, Ohe Y, Koizumi F, Nishio K and Takahashi K: Tocilizumab, a
proposed therapy for the cachexia of Interleukin6-expressing lung
cancer. PLoS One. 9:e1024362014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ando K, Takahashi F, Motojima S, Nakashima
K, Kaneko N, Hoshi K and Takahashi K: Possible role for
tocilizumab, an anti-interleukin-6 receptor antibody, in treating
cancer cachexia. J Clin Oncol. 31:e69–e72. 2013. View Article : Google Scholar
|
|
22
|
Berti A, Boccalatte F, Sabbadini MG and
Dagna L: Assessment of tocilizumab in the treatment of cancer
cachexia. J Clin Oncol. 31:29702013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hirata H, Tetsumoto S, Kijima T, Kida H,
Kumagai T, Takahashi R, Otani Y, Inoue K, Kuhara H, Shimada K, et
al: Favorable responses to tocilizumab in two patients with
cancer-related cachexia. J Pain Symptom Manage. 46:e9–e13. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kim NH, Kim SK, Kim DS, Zhang D, Park JA,
Yi H, Kim JS and Shin HC: Anti-proliferative action of
IL-6R-targeted antibody tocilizumab for non-small cell lung cancer
cells. Oncol Lett. 9:2283–2288. 2015.PubMed/NCBI
|
|
25
|
Yamagata K, Shimojo N, Ito H, Ijima J,
Hasegawa S, Yanagawa T, Mizutani T and Bukawa H: Severe pulmonary
suppuration with infection-induced systemic inflammatory response
syndrome following tongue cancer surgery in a patient undergoing
tocilizumab therapy for rheumatoid arthritis. Case Rep Dent.
2014:6490862014.PubMed/NCBI
|
|
26
|
Calin GA and Croce CM: MicroRNA-cancer
connection: The beginning of a new tale. Cancer Res. 66:7390–7394.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen W, Huang L, Hao C, Zeng W, Luo X, Li
X, Zhou L, Jiang S, Chen Z and He Y: MicroRNA-155 promotes
apoptosis in SKOV3, A2780, and primary cultured ovarian cancer
cells. Tumour Biol. Jan 15–2016.Epub ahead of print.
|
|
28
|
Chen W, Zeng W, Li X, Xiong W, Zhang M,
Huang Y, Zhou L and Jiang S: MicroRNA-509-3p increases the
sensitivity of epithelial ovarian cancer cells to cisplatin-induced
apoptosis. Pharmacogenomics. 17:187–197. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fan JY, Yang Y, Xie JY, Lu YL, Shi K and
Huang YQ: MicroRNA-144 mediates metabolic shift in ovarian cancer
cells by directly targeting Glut1. Tumour Biol. 37:6855–6860. 2016.
View Article : Google Scholar
|
|
30
|
Fu X, Cui Y, Yang S, Xu Y and Zhang Z:
MicroRNA-613 inhibited ovarian cancer cell proliferation and
invasion by regulating KRAS. Tumour Biol. 37:6477–6483. 2016.In
Chinese. View Article : Google Scholar
|
|
31
|
Guo P, Peng D, Xiong X and Zhang S:
Expression of microRNA-100 and its correlation with drug resistance
in human ovarian cancer SKOV3/DDP cells. Nan Fang Yi Ke Da Xue Xue
Bao. 35:1624–1627. 2015.In Chinese. PubMed/NCBI
|
|
32
|
Langhe R: microRNA and ovarian cancer. Adv
Exp Med Biol. 889:119–151. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Liang H, Jiang Z, Xie G and Lu Y: Serum
microRNA-145 as a novel biomarker in human ovarian cancer. Tumour
Biol. 36:5305–5313. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Liu X and Li G: MicroRNA-133b inhibits
proliferation and invasion of ovarian cancer cells through Akt and
Erk1/2 inactivation by targeting epidermal growth factor receptor.
Int J Clin Exp Pathol. 8:10605–10614. 2015.PubMed/NCBI
|
|
35
|
Luo P, Fei J, Zhou J and Zhang W:
microRNA-126 suppresses PAK4 expression in ovarian cancer SKOV3
cells. Oncol Lett. 9:2225–2229. 2015.PubMed/NCBI
|
|
36
|
Pal MK, Jaiswar SP, Dwivedi VN, Tripathi
AK, Dwivedi A and Sankhwar P: MicroRNA: A new and promising
potential biomarker for diagnosis and prognosis of ovarian cancer.
Cancer Biol Med. 12:328–341. 2015.
|
|
37
|
Quitadamo A, Tian L, Hall B and Shi X: An
integrated network of microRNA and gene expression in ovarian
cancer. BMC Bioinformatics. 16(Suppl 5): S52015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shariati-Kohbanani M, Zare-Bidaki M,
Taghavi MM, Taghipour Z, Shabanizadeh A, Kennedy D, Dahim H,
Salahshoor MR, Jalili C and Kazemi Arababadi M: DNA Methylation and
microRNA patterns are in association with the expression of BRCA1
in ovarian cancer. Cell Mol Biol. 62:16–23. 2016.PubMed/NCBI
|
|
39
|
Sun KX, Jiao JW, Chen S, Liu BL and Zhao
Y: MicroRNA-186 induces sensitivity of ovarian cancer cells to
paclitaxel and cisplatin by targeting ABCB1. J Ovarian Res.
8:802015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang ZH and Xu CJ: Research progress of
microRNA in early detection of ovarian cancer. Chin Med J.
128:3363–3370. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wu DD, Li XS, Meng XN, Yan J and Zong ZH:
MicroRNA-873 mediates multidrug resistance in ovarian cancer cells
by targeting ABCB1. Tumour Biol. Feb 5–2016.Epub ahead of
print.
|
|
42
|
Ying HC, Xu HY, Lv J, Ying TS and Yang Q:
MicroRNA signatures of platinum-resistance in ovarian cancer. Eur J
Gynaecol Oncol. 36:16–20. 2015.PubMed/NCBI
|
|
43
|
Catela Ivkovic T, Aralica G, Cacev T,
Loncar B and Kapitanovic S: miR-106a overexpression and pRB
downregulation in sporadic colorectal cancer. Exp Mol Pathol.
94:148–154. 2013. View Article : Google Scholar
|
|
44
|
Díaz R, Silva J, García JM, Lorenzo Y,
García V, Peña C, Rodríguez R, Muñoz C, García F, Bonilla F, et al:
Deregulated expression of miR-106a predicts survival in human colon
cancer patients. Genes Chromosomes Cancer. 47:794–802. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Fang Y, Shen H, Li H, Cao Y, Qin R, Long
L, Zhu X, Xie C and Xu W: miR-106a confers cisplatin resistance by
regulating PTEN/Akt pathway in gastric cancer cells. Acta Biochim
Biophys Sin. 45:963–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hou X, Zhang M and Qiao H: Diagnostic
significance of miR-106a in gastric cancer. Int J Clin Exp Pathol.
8:13096–13101. 2015.
|
|
47
|
Huh JH, Kim TH, Kim K, Song JA, Jung YJ,
Jeong JY, Lee MJ, Kim YK, Lee DH and An HJ: Dysregulation of
miR-106a and miR-591 confers paclitaxel resistance to ovarian
cancer. Br J Cancer. 109:452–461. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Koga Y, Yamazaki N, Yamamoto Y, Yamamoto
S, Saito N, Kakugawa Y, Otake Y, Matsumoto M and Matsumura Y: Fecal
miR-106a is a useful marker for colorectal cancer patients with
false-negative results in immunochemical fecal occult blood test.
Cancer Epidemiol Biomarkers Prev. 22:1844–1852. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Li P, Xu Q, Zhang D, Li X, Han L, Lei J,
Duan W, Ma Q, Wu Z and Wang Z: Upregulated miR-106a plays an
oncogenic role in pancreatic cancer. FEBS Lett. 588:705–712. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Rao YM, Shi HR, Ji M and Chen CH: MiR-106a
targets Mcl-1 to suppress cisplatin resistance of ovarian cancer
A2780 cells. J Huazhong Univ Sci Technolog Med Sci. 33:567–572.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Schee K, Boye K, Abrahamsen TW, Fodstad Ø
and Flatmark K: Clinical relevance of microRNA miR-21, miR-31,
miR-92a, miR-101, miR-106a and miR-145 in colorectal cancer. BMC
Cancer. 12:5052012. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Wang Z, Liu M, Zhu H, Zhang W, He S, Hu C,
Quan L, Bai J and Xu N: miR-106a is frequently upregulated in
gastric cancer and inhibits the extrinsic apoptotic pathway by
targeting FAS. Mol Carcinog. 52:634–646. 2013. View Article : Google Scholar
|
|
53
|
Zhang L, Meng L, Fan Z, Liu B, Pei Y and
Zhao Z: Expression of plasma miR-106a in colorectal cancer and its
clinical significance. Nan Fang Yi Ke Da Xue Xue Bao. 34:354–357.
2014.In Chinese. PubMed/NCBI
|