Introduction
Colorectal cancer (CRC) is one of the most common
cancers worldwide. Despite recent advances in cancer therapy
including chemotherapy, radiation and surgery, metastasis to other
organs such as the lymph nodes, the liver and the lungs remains on
ongoing issue (1). The survival
rate of CRC patients has increased, but only by a relatively short
period (20 months) when compared to other types of cancer (2,3). A
better understanding of the mechanisms involved in the pathways of
metastasis may improve the management of CRC.
The runt-related transcription factor (RUNX) family
consists of RUNX1, RUNX2 and RUNX3, which play an important role in
cell proliferation and tumorigenesis (4). Among them, RUNX3, in particular, has
been shown to play a tumor-suppressor role in several types of
cancers and its expression levels are downregulated in tumor
tissues (5,6). RUNX3 was first suggested to be a tumor
suppressor in gastric cancer (7).
Recent studies have shown that RUNX3 expression levels are
downregulated in CRC, either from hemizygous deletion or from
promoter methylation of the RUNX3 gene. Furthermore, a decrease in
RUNX3 protein expression was found to be significantly associated
with decreased survival of CRC patients (8,9). Mu
et al (10) showed that
RUNX3 expression was decreased in 70% of primary human colorectal
tumors; with over 90% of late stage tumors that represent highly
metastatic tumors, showing reduced RUNX3 levels.
Epithelial-mesenchymal transition (EMT) is a
morphogenetic process of the malignant transformation of epithelial
cells. The EMT phenotype is characterized by the loss of
cell-to-cell adhesion and remodeling of the actin cytoskeleton as
well as the loss of the epithelial molecule E-cadherin and the gain
of mesenchymal markers such as vimentin and Snail (11,12).
EMT is known to be necessary for cells to obtain migratory and
invasive capabilities (13). It is
necessary for tissue invasion, which is an essential step in
metastasis that requires breakdown of the extracellular matrix
(ECM) around the cancer cells. In the complicated multi-steps of
cancer metastasis, one of the first steps is the degradation of the
ECM by matrix metalloproteinases (MMPs). MMPs are mainly secreted
during tumor development, invasion and metastasis; the secretion of
MMPs promotes the invasion of malignant cancer cells into adjoining
tissue, blood and lymph vessels. MMPs, particularly MMP-2 and
MMP-9, have been postulated to promote invasion and lymph node
metastasis of gastrointestinal cancer cells. Suppression of MMP
activity impairs cancer cell migration and angiogenesis (14,15),
the recruitment of new blood vessels, which is required for
invasiveness and metastasis (16,17),
and is regulated by the angiogenic stimulator vascular endothelial
growth factor (VEGF) (18).
Therefore, tumor angiogenesis and metastasis are key targets for
anticancer therapy. It has been found that RUNX3 suppresses cell
motility and metastasis in renal cell carcinoma and prostate cancer
(19,20). This suggests that RUNX3 plays an
important role in tumorigenesis and progression. However, the role
of RUNX3 in CRC has not yet been well studied.
In the present study, we found that deficiency or
loss of RUNX3 expression was directly correlated with CRC
tumor-node-metastasis (TNM) stage. Furthermore, restoration of
RUNX3 expression led to the suppression of MMP-2 and MMP-9, and at
least in part, to the inhibition of tumor metastasis. We also
demonstrated that the reduction in VEGF release induced by RUNX3
reintroduction, inhibited CRC angiogenesis. Our clinical and
mechanistic data indicated that RUNX3 may be a tumor suppressor
involved in the progression of CRC.
Materials and methods
Reagents
RPMI-1640 medium, fetal bovine serum (FBS) and
antibiotics were purchased from Gibco (Carlsbad, CA, USA).
Antibodies obtained included RUNX3 (Abcam, Cambridge, MA, USA),
E-cadherin (BD Biosciences, San Jose, CA, USA), β-actin (Sigma, St.
Louis, MO, USA), vimentin (Dako, Carpinteria, CA, USA) and Snail
(Santa Cruz Biotechnology, Santa Cruz, CA, USA). MMP-2 inhibitor
was also purchased from Santa Cruz Biotechnology.
Cell lines and transfection
The human CRC cell lines (HCT116, DLD-1, SW480,
SW620 and Colo205) were purchased from the Korea Cell Line Bank
(KCLB) and maintained according to KCLB's instructions.
Transfection of 1 µg of plasmid DNA [pFlag-c1 (con) and
pFlag-c1 RUNX3] into CRC cells was performed using Lipofectamine
2000 transfection reagent (Invitrogen, Carlsbad, CA, USA).
Transfection of the RUNX3 siRNA and control siRNA (200 nM) was
carried out using Lipofectamine RNAiMAX transfection reagent
(Invitrogen). RUNX3 siRNA forward was 5′-GUG AUG GCA GGC AAU GAC
GAG AAC U-3′ and reverse, 5′-AGU UCU CGU CAU UGC CUG CCA UCA
C-3′.
Wound-healing and Matrigel invasion
assays
After transfection, the cells were seeded at
5×105 cells/well in 12-well plates. At 100% confluence,
two parallel wounds were made using a plastic pipette tip. The
cells were then grown in culture medium with 5% FBS. Images of the
wound were collected at 0, 24 and 48 h using a microscope. The
migration rate was quantified by measuring the distance between the
wound edges. This assay was independently repeated three times. For
the Matrigel invasion assay, 3×105 cells/well were
seeded in the upper chamber, which was coated with Matrigel (BD
Biosciences). After 48 h at 37°C in 5% CO2, the cells
present on the lower surface of the insert were stained with
Diff-Quik Stain kit (Biochemical Sciences, Inc., Swedesboro, NJ,
USA). The cells invaded through the Matrigel-coated membrane were
counted by microscopy.
Cell viability assay
CRC cells were grown in a 96-well plate at
1×104 cells/well. The cells were transfected with
pFlag-cl RUNX3 or pFlag-cl (con). At 24 h after transfection, 25
µl of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium
bromide (MTT) (5 mg/ml) (Sigma-Aldrich Co. LLC, St. Louis, MO, USA)
was added to each well, and the plates were incubated for 4 h at
37°C. The MTT solution in the medium was then aspirated, and 150
µl of dimethyl sulfoxide (DMSO) was added before measurement
of the absorbance at 550 nm.
Colony formation assay
Stably transfected HCT116 and DLD-1 cells were
diluted and seeded into 6-well plates at a density of 500
cells/well, and then were cultured at 37°C. The medium was changed
every three days. After one week, the cells were washed with
phosphate-buffered saline (PBS), fixed with 4% paraformaldehyde for
30 min, and then stained with crystal violet for 30 min for
visualization and counting.
Immunofluorescence
Cells grown on a glass coverslip were fixed with
3.7% formaldehyde for 15 min, followed by permeabilization with
0.5% Triton X-100 for 15 min at room temperature. The cells were
then blocked for 1 h with 3% bovine serum albumin. Primary
antibodies were applied overnight at 4°C, followed by incubation
with secondary Alexa Fluor 594-conjugated secondary antibody
(Molecular Probes, Eugene, OR, USA) or the FITC-conjugated
secondary antibody (Sigma). The nuclei were costained with
4′,6-diamidino-2-phenylindole (DAPI) and visualized using
fluorescence microscopy.
Western blotting
Cells were lysed in RIPA buffer [50 mM Tris, 150 mM
NaCl, 1% Triton X-100, 0.1% SDS and 1% Na-deoxycholate (pH 7.4)]
with proteases and phosphatase inhibitor cocktails and were
subjected to SDS-PAGE. Cells were then transferred onto
nitrocellulose membranes (GE Healthcare Life Sciences, Logan, UT,
USA), blocked with TBS containing 0.2% Tween-20 and 5% skim milk,
incubated with the primary antibody and then incubated with the
horseradish peroxidase-labeled secondary antibody. The signals were
detected by X-ray film.
RT-PCR and real-time PCR
Total RNA was extracted using TRIzol reagent (Life
Technologies, Grand Island, NY, USA). Amplification of transcripts
was performed by reverse transcriptase polymerase chain reaction
kit (Life Technologies). RT-PCR primers for MMP-2 were: forward,
5′-TGA TGG TGT CTG CTG GAA AG-3′ and reverse, 5′-GAC ACG TGA AAA
GTG CCT TG-3′); and for MMP-9 forward, 5′-AAG ATG CTG CTG TTC AGC
GGG-3′ and reverse, 5′-GTC CTC AGG GCA CTG CAG GAT-3′). Real-time
PCR was performed on an Applied Biosystems 9700 real-time PCR using
gene specific oligonucleotide primers for TaqMan probes (Applied
Biosystems, Foster City, CA, USA). TaqMan probes were as follows:
GAPDH (Hs99999905_m1), RUNX3 (Hs00231709_m1) and MMP-2
(Hs01548727_m1). For expression of mRNA, gene expression was
normalized to GAPDH.
Tube formation assay
For the tube formation assay, a 48-well plate was
coated with Matrigel (BD Biosciences). Human umbilical vein
endothelial cells (HUVECs) (3×104) were suspended in 100
µl Medium 200PRF (Life Technologies) and applied to the
pre-coated 48-well plate. After 6 h, the number of capillary-like
tubes was counted using microscopy.
Enzyme-linked immunosorbent assay (ELISA)
for VEGF
HCT116 and DLD-1 cells were seeded in a 60-mm dish.
The supernatants were collected at 48 h. The levels of VEGF
secreted by the cells in the medium were detected using VEGF ELISA
kit (R&D Systems, Minneapolis, MN, USA).
Results
RUNX3 expression is decreased in human
CRC
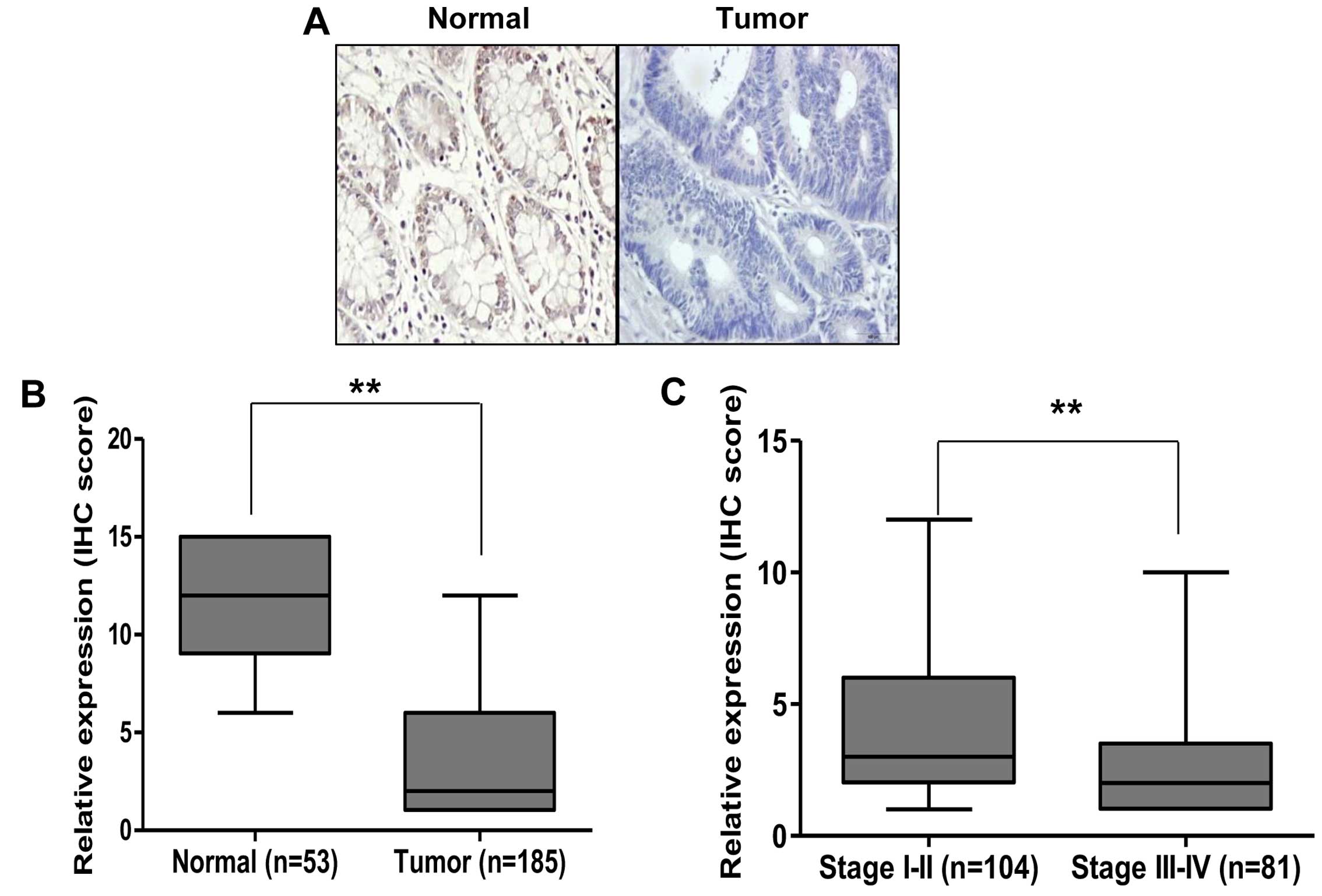
We first investigated whether expression of RUNX3 is
reduced in human CRC. Immunohistochemistry (IHC) staining was
performed on TMA slides containing normal and tumor cancer tissues.
As shown in Fig. 1A, from the
serial sections of paired normal colorectal and tumor tissues of
CRC patients, RUNX3 was highly expressed in the normal colorectal
tissues, but lower in the tumor tissues. Our IHC data showed that
there was a significantly lower level of RUNX3 in the tumor tissues
than that noted in the normal colon tissues (P<0.01, Fig. 1B). Since TNM stage is an important
prognostic and metastatic marker for patients, we confirmed
expression of RUNX3 in stage I–II (n=104) and stage III–IV (n=81)
CRC tissues. We found that expression of RUNX3 was markedly
decreased in stage III–IV compared with stage I–II tissues
(Fig. 1C). These results suggest
that RUNX3 may be an important prognostic marker in CRC
patients.
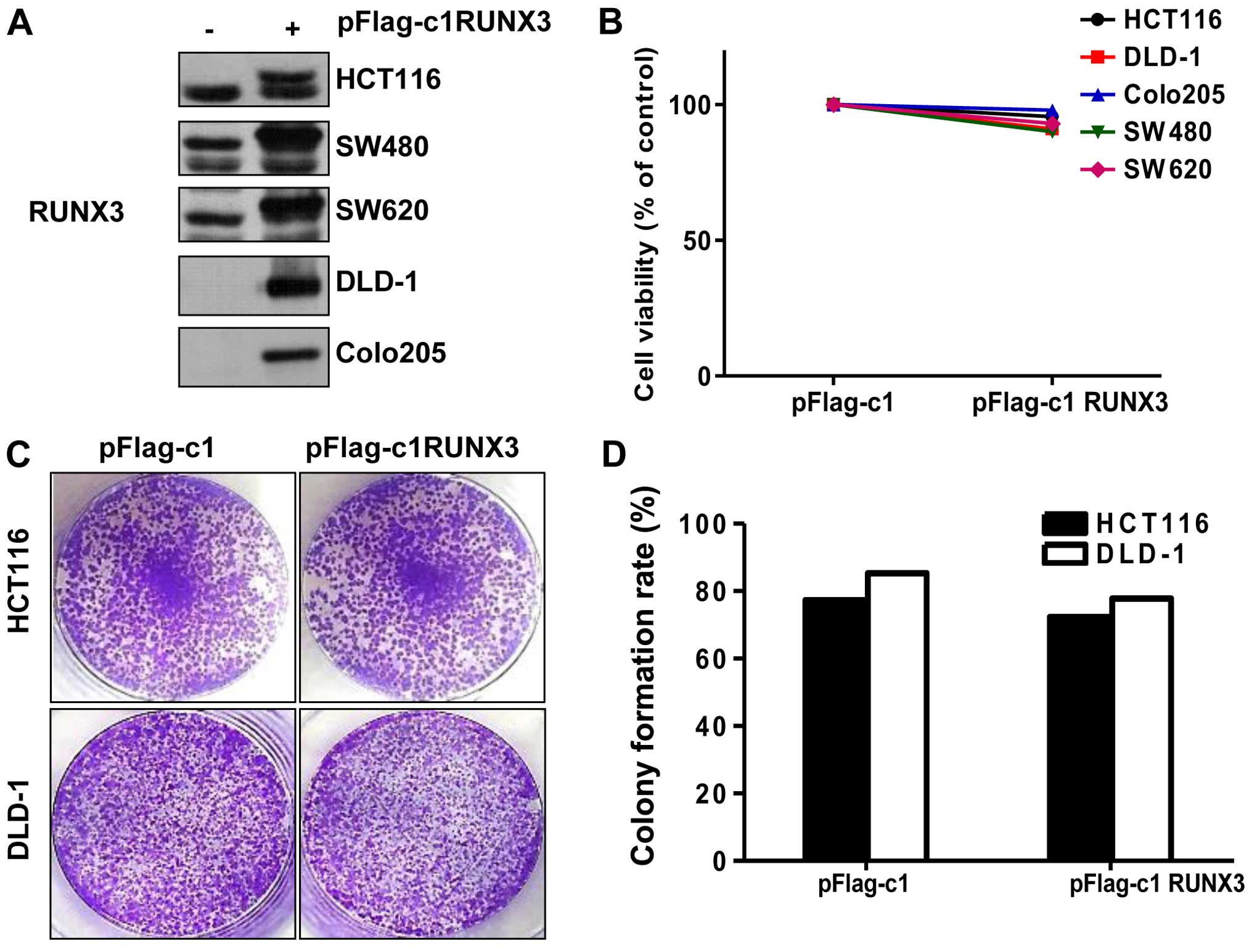
RUNX3 does not affect cell viability and
proliferation in CRC cells
A previous study reported that loss of RUNX3 induces
proliferative effect in gastric cancer cells (21). To investigate the function of RUNX3
in CRC, we tested whether RUNX3 affects the viability of CRC cells.
We first determined the endogenous protein level of RUNX3 in CRC
cell lines. As shown in Fig. 2A,
HCT116, SW480 and SW620 cells expressed a higher RUNX3 level than
corresponding DLD-1 and Colo205 cells. Then, after pFlag-c1 RUNX3
or pFlag-c1 (con) transfection, the cell viability of the CRC cells
was performed by MTT assay, but there was no change in the
RUNX3-overexpressing cells (Fig.
2B). In order to further confirm cell proliferation, we
performed a colony formation assay. As shown in Fig. 2C and D, there was no significant
difference in colony formation ability in RUNX3-overexpressing
HCT116 and DLD-1 cells compared with this ability in cells
transfected with pFlag-c1 (control).
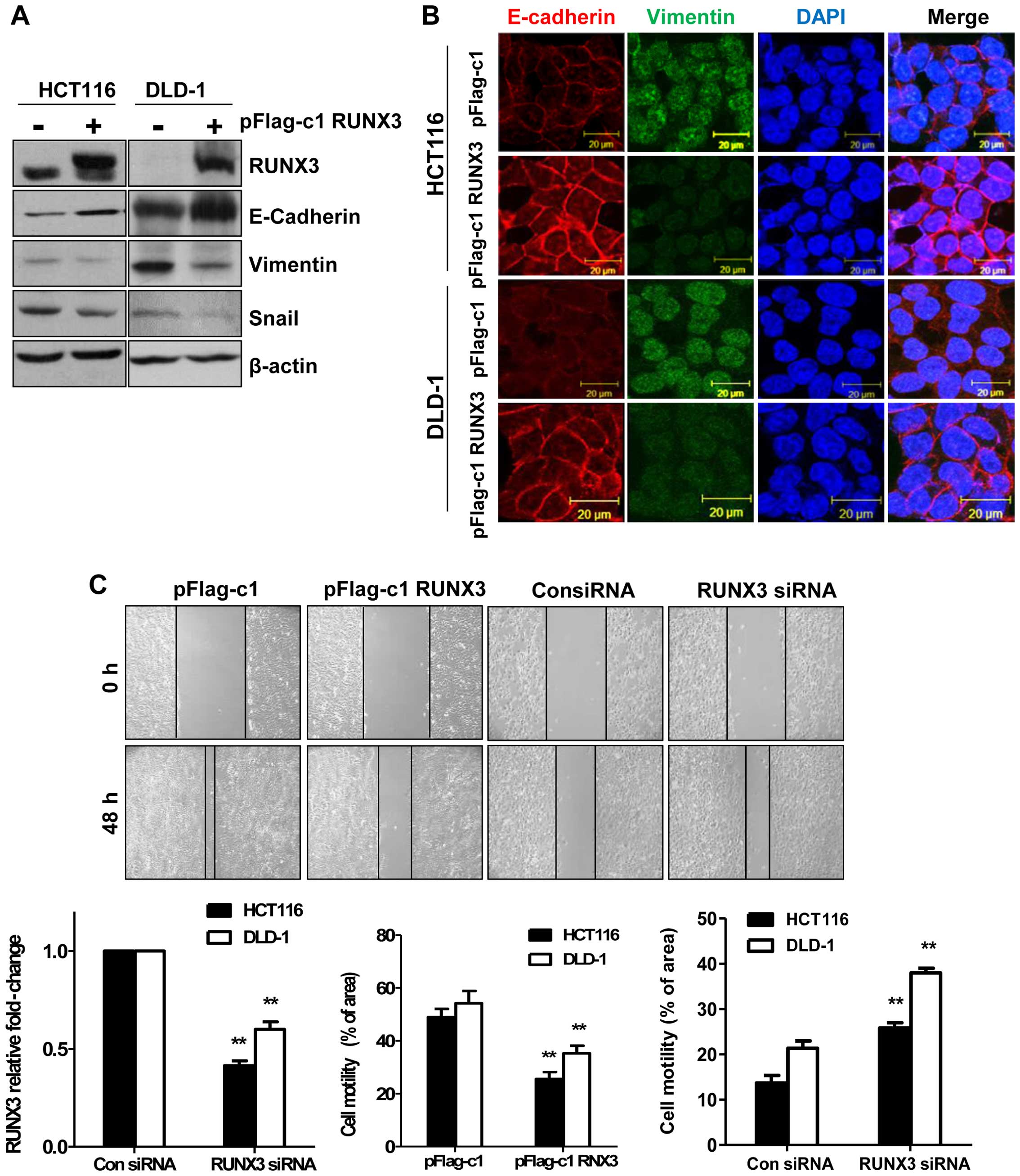
RUNX3 plays an important role in motility
and invasiveness
Since Fig. 1A
demonstrates that expression of RUNX3 is associated with TNM stage,
RUNX3 may play an important role in CRC metastasis. Thus, we
performed an experiment to test whether overexpression of RUNX3
could prevent EMT. First, we examined EMT markers at the protein
levels in the RUNX3-overexpressing cells. As shown in Fig. 3A, epithelial marker E-cadherin was
significantly increased while mesenchymal markers vimentin and
Snail were decreased in the RUNX3-overexpressing cells compared
with these levels in pFlag-c1 (con) cells. These observations were
also confirmed by immunofluorescence. E-cadherin expression in the
RUNX3-overexpressing cells was increased and vimentin expression in
the RUNX3-overexpressing cells was decreased (Fig. 3B). We next investigated the effect
of RUNX3 on the cell motility of the CRC cells. Cell motility was
significantly decreased in the RUNX3-overexpressing cells. In
contrast, cell migration was also increased in the RUNX3
siRNA-transfected HCT116 and DLD-1 cells (Fig. 3C). In addition, to examine the
effect of RUNX3 expression on the metastatic ability of CRC cells,
HCT116 and DLD-1 cells were transiently transfected before
performing Matrigel invasion assay. The RUNX3-overexpressing cells
had decreased cell invasive ability while knockdown of RUNX3
increased the cell invasive ability compared with that of the cells
transfected with control siRNA (Fig.
3D). The results suggest that RUNX3 acts as a key regulator of
metastasis and EMT.
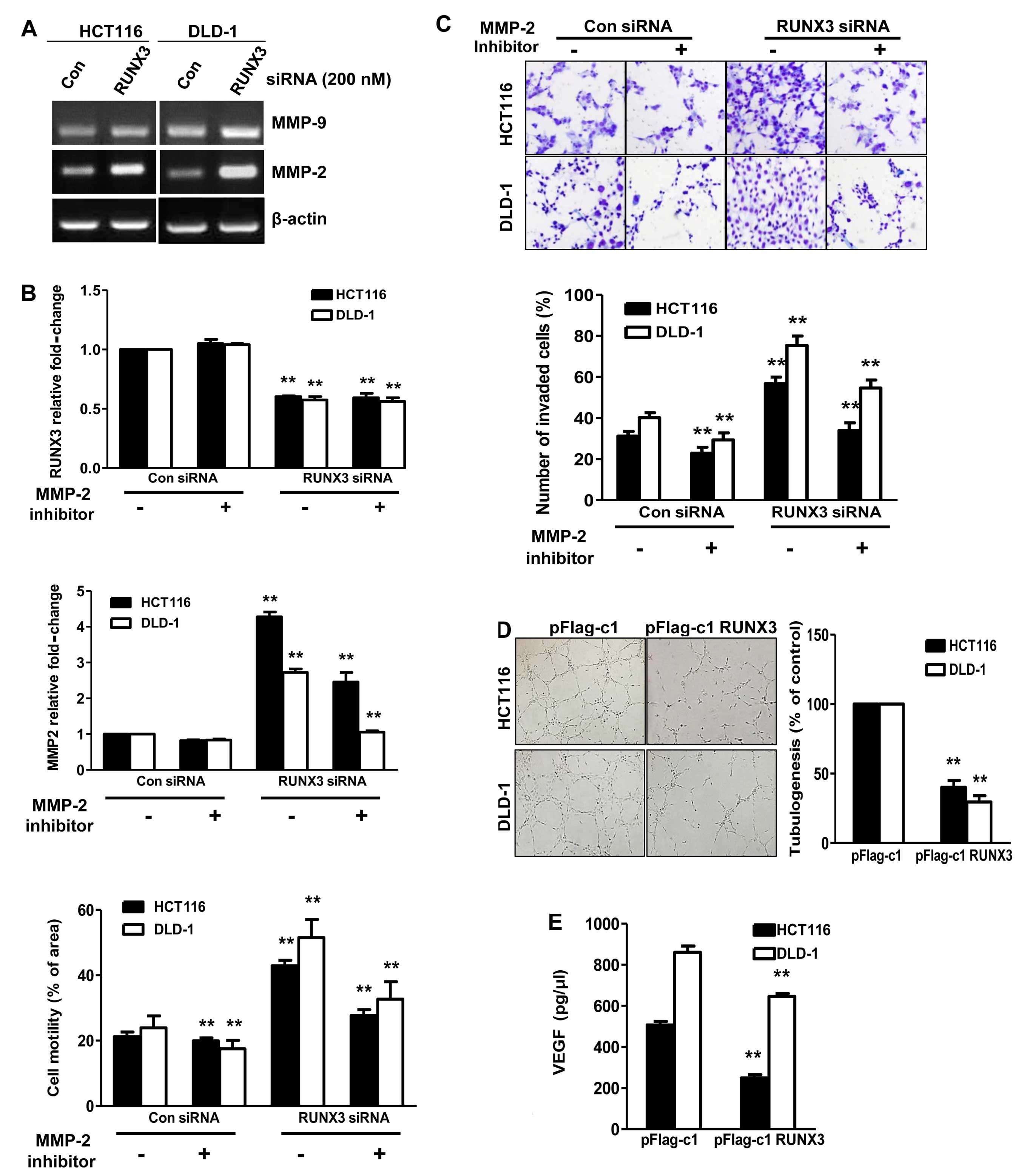
RUNX3 is a regulator of MMPs for
invasiveness and angiogenesis
To study the role of RUNX3 in cell invasion, we
measured the activities of MMP-2 and MMP-9 by RT-PCR. As shown in
Fig. 4A, the activity of MMP-2 was
increased, but not that of MMP-9, in the RUNX3 siRNA-transfected
HCT116 and DLD-1 cells compared with the activities in the con
siRNA cells. Hence, we hypothesized that MMP-2 is affected by the
invasive effects of cells transfected with RUNX3 siRNA. To test
this hypothesis, we treated the cells with the MMP-2 inhibitor (5
µM) for 24 h after transfection of RUNX3 siRNA. Cell
motility promoted by RUNX3 siRNA was significantly decreased by the
MMP-2 inhibitor (Fig. 4B). We then
assessed the effects of the MMP-2 inhibitor on the invasion of
RUNX3 siRNA-transfected HCT116 and DLD-1 cells. As shown in
Fig. 4C, the cells treated with the
MMP-2 inhibitor showed significantly lower invasiveness. Together,
these data demonstrated that RUNX3 inhibits metastasis through
MMP-2. To further examine the effect of RUNX3 on angiogenesis, we
performed an endothelial cell tube formation assay. The degree of
tube formation was assessed as the percentage of cell surface area
vs. total surface area. As shown in Fig. 4D, the average number of complete
tubular structures formed by HUVECs was significantly reduced in
the conditioned medium from the RUNX3-overexpressing HCT116 and
DLD-1 cells compared with the vector controls. To evaluate the
mechanism involved in the regulation of angiogenesis by RUNX3,
ELISA was performed to detect VEGF secretion into the conditioned
culture medium of CRC cells. As shown in Fig. 4E, a significant reduction in VEGF
secretion was observed in the conditioned medium from the HCT116
and DLD-1 cells transfected with pFlag-RUNX3 compared with the
control cells (P<0.05).
Discussion
In previous studies, RUNX3 in colorectal cancer
(CRC) has been considered to be associated with cell apoptosis and
hypermethylation (22,23). However, the effect of RUNX3
alteration on tumor angiogenesis and metastasis has not been
examined in CRC. In the present study, we found that RUNX3 was
frequently silenced or downregulated in CRC cell lines, as well as
in primary CRC compared to normal colorectal mucosa tissues.
Specifically, we found that the expression of RUNX3 was decreased
in the late stage III–IV compared to the early stage I–II tumor
samples (Fig. 1C). These clinical
data clearly demonstrated that expression of RUNX3 affects cancer
metastasis in CRC. Finally, functional experiments revealed that
restoration of RUNX3 in CRC cells inhibited cell proliferation,
suppressed cell invasion and migration and regulated EMT-related
proteins, E-cadherin, vimentin and Snail. All things considered,
our data suggest that RUNX3 is a tumor-suppressor gene, which is
downregulated through promoter hypermethylation in CRC.
The major cause of mortality of cancer patients is
metastasis (24). The EMT process
in tumor cells is an essential step in metastasis. Thus, we
detected changes in expression of epithelial marker E-cadherin and
mesenchymal markers Snail and vimentin corresponding to changes in
RUNX3 expression. In the present study, knockdown of RUNX3
expression markedly weakened the migration and invasion ability of
the HCT116 and DLD-1 cells compared with the negative control
(Fig. 3). It has also been found
that downregulation of the epithelial marker E-cadherin induced the
expression of various mesenchymal markers, such as Snail and
vimentin, during EMT (25).
Consistent with these observations, we found that overexpression of
RUNX3 significantly inhibited motility and invasiveness in CRC
cells. Whereas, knockdown of RUNX3 promoted motility and
invasiveness in the HCT116 and DLD-1 cells (Fig. 3). These results indicated that RUNX3
may inhibit EMT progression in CRC cells.
It is possible that MMPs degrade components of the
basement membrane and ECM, allowing cancer cells to invade and
migrate (26,27). In addition, several studies have
demonstrated that MMP-2 and MMP-9 promote invasion and metastasis
in CRC (28). To determine how
RUNX3 inhibits CRC cell migration and invasion, we focused on
clarifying the relationship between RUNX3 and MMPs, which have been
reported to participate in tumor progression. In the present study,
we found that RUNX3 overexpression significantly inhibited the
expression of MMP-2 and MMP-9 in HCT116 and DLD-1 cells (Fig. 4A–C). We found that knockdown of
RUNX3 induced expression of MMP-2 but not of MMP-9. Our data imply
that RUNX3 may suppress CRC cell invasion and migration by
decreasing MMP-2 and MMP-9 protein expression. However, it remains
to be elucidated how RUNX3 regulates MMP-2 and MMP-9 expression and
how the signaling pathway of RUNX3 regulates CRC cell invasion.
VEGF, an angiogenic factor, has been identified as a
key mediator of tumor angiogenesis involved in the development of
tumor blood supply in the progression of solid tumors (29). Previous studies have demonstrated
that RUNX3 downregulated VEGF expression via transcriptional
repression in human gastric cancer and renal cancer cells (19,30).
We detected the expression and secretion of VEGF after RUNX3
transfection. Our data showed that VEGF secretion was reduced by
restoration of RUNX3 (Fig. 4D and
E). These results suggested that RUNX3 suppresses blood vessel
formation by regulating VEGF secretion.
In summary, the present study revealed that the
tumor-suppressor gene RUNX3 was inactivated by promoter methylation
in CRC cell lines and primary CRC tissues. Restoration of RUNX3
enhanced cell invasion and migration, but had no effect on cell
proliferation. Consequently, these findings show that the knockdown
or restoration of RUNX3 presents a possible means for inhibiting
angiogenesis or metastasis and for inhibiting CRC progression.
Acknowledgments
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education, Science and Technology
(no. NRF-2013R1A1A2064367) and the Hanmi Pharm. Co., Ltd. (no.
11500841).
References
|
1
|
Geoghegan JG and Scheele J: Treatment of
colorectal liver metastases. Br J Surg. 86:158–169. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Yokota J: Tumor progression and
metastasis. Carcinogenesis. 21:497–503. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Fidler IJ: The pathogenesis of cancer
metastasis: The 'seed and soil' hypothesis revisited. Nat Rev
Cancer. 3:453–458. 2003. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lund AH and van Lohuizen M: RUNX: A
trilogy of cancer genes. Cancer Cell. 1:213–215. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bae SC and Choi JK: Tumor suppressor
activity of RUNX3. Oncogene. 23:4336–4340. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Subramaniam MM, Chan JY, Yeoh KG, Quek T,
Ito K and Salto-Tellez M: Molecular pathology of RUNX3 in human
carcinogenesis. Biochim Biophys Acta. 1796:315–331. 2009.PubMed/NCBI
|
|
7
|
Li QL, Ito K, Sakakura C, Fukamachi H,
Inoue K, Chi XZ, Lee KY, Nomura S, Lee CW, Han SB, et al: Causal
relationship between the loss of RUNX3 expression and gastric
cancer. Cell. 109:113–124. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Soong R, Shah N, Peh BK, Chong PY, Ng SS,
Zeps N, Joseph D, Salto-Tellez M, Iacopetta B and Ito Y: The
expression of RUNX3 in colorectal cancer is associated with disease
stage and patient outcome. Br J Cancer. 100:676–679. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Silva TD, Vidigal VM, Felipe AV, DE Lima
JM, Neto RA, Saad SS and Forones NM: DNA methylation as an
epigenetic biomarker in colorectal cancer. Oncol Lett. 6:1687–1692.
2013.PubMed/NCBI
|
|
10
|
Mu WP, Wang J, Niu Q, Shi N and Lian HF:
Clinical significance and association of RUNX3 hypermethylation
frequency with colorectal cancer: A meta-analysis. Onco Targets
Ther. 7:1237–1245. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thiery JP: Epithelial-mesenchymal
transitions in development and pathologies. Curr Opin Cell Biol.
15:740–746. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bedi S, Vidyasagar A and Djamali A:
Epithelial-to-mesenchymal transition and chronic allograft
tubulointerstitial fibrosis. Transplant Rev. 22:1–5. 2008.
View Article : Google Scholar
|
|
13
|
Tsai JH and Yang J: Epithelial-mesenchymal
plasticity in carcinoma metastasis. Genes Dev. 27:2192–2206. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wagenaar-Miller RA, Gorden L and Matrisian
LM: Matrix metalloproteinases in colorectal cancer: Is it worth
talking about? Cancer Metastasis Rev. 23:119–135. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zucker S and Vacirca J: Role of matrix
metalloproteinases (MMPs) in colorectal cancer. Cancer Metastasis
Rev. 23:101–117. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zetter BR: Angiogenesis and tumor
metastasis. Annu Rev Med. 49:407–424. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Folkman J: Role of angiogenesis in tumor
growth and metastasis. Semin Oncol. 29(Suppl 16): S15–S18. 2002.
View Article : Google Scholar
|
|
18
|
Leung DW, Cachianes G, Kuang WJ, Goeddel
DV and Ferrara N: Vascular endothelial growth factor is a secreted
angiogenic mitogen. Science. 246:1306–1309. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Chen F, Bai J, Li W, Mei P, Liu H, Li L,
Pan Z, Wu Y and Zheng J: RUNX3 suppresses migration, invasion and
angiogenesis of human renal cell carcinoma. PLoS One. 8:e562412013.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Chen F, Wang M, Bai J, Liu Q, Xi Y, Li W
and Zheng J: Role of RUNX3 in suppressing metastasis and
angiogenesis of human prostate cancer. PLoS One. 9:e869172014.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Cheng HC, Liu YP, Shan YS, Huang CY, Lin
FC, Lin LC, Lee L, Tsai CH, Hsiao M and Lu PJ: Loss of RUNX3
increases osteopontin expression and promotes cell migration in
gastric cancer. Carcinogenesis. 34:2452–2459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tong DD, Jiang Y, Li M, Kong D, Meng XN,
Zhao YZ, Jin Y, Bai J, Fu SB and Geng JS: RUNX3 inhibits cell
proliferation and induces apoptosis by TGF-beta-dependent and
-independent mechanisms in human colon carcinoma cells.
Pathobiology. 76:163–169. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Nishio M, Sakakura C, Nagata T, Komiyama
S, Miyashita A, Hamada T, Kuryu Y, Ikoma H, Kubota T, Kimura A, et
al: RUNX3 promoter methylation in colorectal cancer: Its
relationship with microsatellite instability and its suitability as
a novel serum tumor marker. Anticancer Res. 30:2673–2682.
2010.PubMed/NCBI
|
|
24
|
Tam WL and Weinberg RA: The epigenetics of
epithelial-mesenchymal plasticity in cancer. Nat Med. 19:1438–1449.
2013. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang M, Zhao F, Li S, Chang AK, Jia Z,
Chen Y, Xu F, Pan H and Wu H: AIB1 cooperates with ERα to promote
epithelial mesenchymal transition in breast cancer through SNAI1
activation. PLoS One. 8:e655562013. View Article : Google Scholar
|
|
26
|
Rundhaug JE: Matrix metalloproteinases and
angiogenesis. J Cell Mol Med. 9:267–285. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Yang B, Tang F, Zhang B, Zhao Y, Feng J
and Rao Z: Matrix metalloproteinase-9 overexpression is closely
related to poor prognosis in patients with colon cancer. World J
Surg Oncol. 12:242014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cao Y, e G, Wang E, Pal K, Dutta SK,
Bar-Sagi D and Mukhopadhyay D: VEGF exerts an
angiogenesis-independent function in cancer cells to promote their
malignant progression. Cancer Res. 72:3912–3918. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Peng Z, Wei D, Wang L, Tang H, Zhang J, Le
X, Jia Z, Li Q and Xie K: RUNX3 inhibits the expression of vascular
endothelial growth factor and reduces the angiogenesis, growth, and
metastasis of human gastric cancer. Clin Cancer Res. 12:6386–6394.
2006. View Article : Google Scholar : PubMed/NCBI
|