Introduction
As well known potential carcinogens, reactive oxygen
species (ROS) are involved in both carcinogenesis and tumor
progression via ROS-induced DNA damage (1–4).
During normal metabolism, cells avoid oxidative damage by means of
antioxidant defense mechanisms that control the balance between the
generation and removal of oxygen radicals. The antioxidant system
includes various enzymes, such as superoxide dismutase and
catalase, as well as glutathione peroxidase (GPX) (5,6).
GPX3, a member of the GPX family, is an extra
cellular glycosylated enzyme that reduces both phospholipid
hydroperoxides and fatty acid hydroperoxides using various electron
donors, such as glutathione, thioredoxin, and glutaredoxin
(7–10). It is generally known that
hydroperoxides are involved in various biological behavior of
cancer cells such as proliferation, motility, and invasion
(11). Therefore, some
investigators expected that GPX3 might play a role in cancer
carcinogenesis by controlling the hydroperoxide levels inside cells
(11).
The human GPX3 gene, located on chromosome
5q32, is the only known extracellular antioxidant isoform
containing selenocysteine at the catalytic site (12–15).
GPX3 activity is regulated by the availability of selenium, a well
known cancer chemopreventive agent (16). Selenium uptake is related to
decreased cancer incidence, and although there is no direct
experimental evidence, this linkage may be related to the
anticancer effect of selenium (17).
GPX3 is mainly synthesized by the proximal tubules
of the kidney (18). GPX3 is
readily detectable in extracellular body fluids such as the aqueous
humor in the anterior and posterior chambers of the eye, blood
plasma, and thyroid colloid (19,20).
Moreover, transcripts of GPX3 are also detected in
epithelial cells of the oviduct (21). The mechanisms that regulate the
transcription of GPX3 remain largely unknown. Recent studies
have shown that oxidative stress can induce transcriptional
upregulation of GPX3 in inflammatory bowel disease and
experimental colitis, in patients with asthma, and in the diabetic
mouse heart (22–25). However, GPX3 expression is usually
downregulated in cancer tissues compared to its expression in
normal tissues (26). In contrast,
overexpression of GPX3 in prostate cancer cells has been found to
attenuate tumorigenic potential and spontaneous metastasis in
vivo (27). GPX3 may
serve as a potential tumor suppressor gene in cancer
progression.
Genetic and epigenetic mechanisms lead to
inactivation of tumor suppressor genes in cancers. As one of the
major modifications involved in epigenetic inactivation of genes,
abnormal methylation in CpG-rich promoter regions can induce
transcriptional silencing of tumor suppressor genes. Promoter
hypermethylation of GPX3 is frequently detected in various
cancer tissues, such as gastric, prostate, and Barrett's esophageal
cancer, and this may be a common and crucial cause of GPX3
downregulation in cancers.
Malignant melanoma is a common leading cause of
cancer-related death among patients with skin cancer. The incidence
of cutaneous melanoma has rapidly increased over the last 50 years
(28). However, the molecular basis
of melanoma is largely unknown. In the current study, for the first
time, we determined whether GPX3 is also downregulated in melanoma
and, if so, whether promoter hypermethylation is associated with
its repression. We further investigated the influence of GPX3 on
the biological behavior of melanoma cells and its
clinicopathological significance in melanoma patients.
Materials and methods
Patients
Two tissue microarray (TMA) sections of melanoma
samples were purchased from Pantomics, Inc. (Richmond, CA, USA) and
Super BioChips (Seoul, Korea), respectively, and used for
immunohistochemistry. The specimens included 70 primary melanoma
and 25 metastatic melanoma tissue samples. Three normal skin
tissues and 10 primary melanoma tissue samples were obtained from
Yanbian University Hospital and used for quantitative real-time PCR
(qRT-PCR) and methylation-specific PCR (MSP) analysis. The
clinicopathological characteristics of all patients are shown in
Table I. This study was approved by
the Institutional Review Board of Yanbian University Hospital.
 | Table ICharacteristics of the patients with
malignant melanoma. |
Table I
Characteristics of the patients with
malignant melanoma.
| Variables | No. of patients (%)
|
|---|
| Primary
melanoma | Metastatic
melanoma |
|---|
| Total cases | 80 | 25 |
| Age, years | | |
| Median age
(range) | 56 (11–84) | 55 (25–73) |
| Gender | | |
| Male | 47 (58.8) | 14 (56.0) |
| Female | 33 (41.2) | 11 (44.0) |
| Survival | | |
| Alive | 12 (15.0) | 4 (16.0) |
| Dead | 34 (42.5) | 9 (36.0) |
| Unknown | 34 (42.5) | 12 (48.0) |
Cell culture and establishment of GPX3
knockdown SK-MEL-24 cells
The human melanoma cell lines (SK-MEL-2 and
SK-MEL-24), and adult human epidermal melanocytes (HEMs) were
purchased from ATCC (Rockville, MD, USA). Melanoma cell lines were
cultured in microvascular endothelial cell medium (MEM)
supplemented with 10% fetal bovine serum (FBS), 100 U/ml
ampicillin, 100 µg/ml streptomycin, and 2 mM glutamine
(Gibco BRL, Gaithersburg, MD, USA). HEMs were cultured in MGM™-4
melanocyte cell basal medium (Lonza Group AG, Basel, Switzerland)
supplemented with MBM-4 plus SingleQuots™. Cells were grown in a
humidified atmosphere of 5% CO2 at 37°C. SK-MEL-24 cells
were positive for GPX3 on both the mRNA and protein levels.
GPX3-depleted stable SK-MEL-24 cell lines were created using GPX3
shRNA-encoding plasmid (Origin Pharmaceutical Services Ltd.,
Abingdon, Oxfordshire, UK). Retroviruses were generated in 293T
cells and the transduced cells were selected with 1 µg/ml
puromycin. Silencing of GPX3 in SK-MEL-24 cell lines was
confirmed by reverse transcription polymerase chain reaction
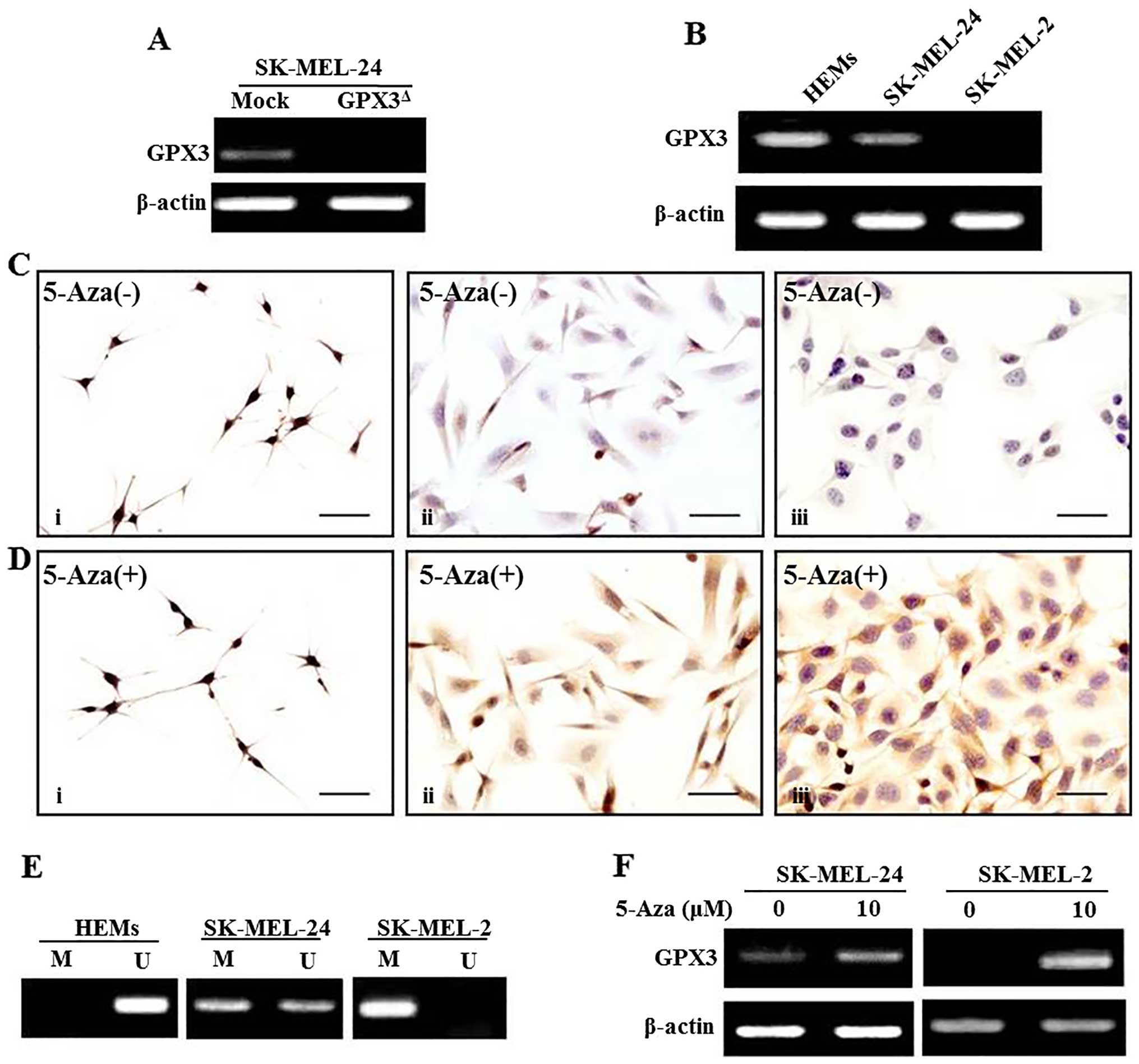
(RT-PCR) analysis (Fig. 1A).
Immunofluorescence in TMA sections
Melanoma tissue sections were deparaffinized in
xylene and rehydrated in graded alcohol. Antigen retrieval was
performed by a 2 min autoclave treatment using antigen retrieval
buffer (Dako North America, Inc., Carpinteria, CA, USA). Sections
were treated with primary antibody against GPX3 (working dilution:
1:100; Abcam plc, Cambridge, UK) at room temperature (RT) for 1 h,
and the Donkey Anti-Mouse IgG H&L (Alexa Fluor® 488)
(Abcam) was used as the secondary antibody. Counterstaining was
performed using 4′,6-diamidino-2-phenylindole (DAPI). In this
study, sections of cell blocks from HeLa cells were used as a
positive control for staining of GPX3. Rabbit IgG (R&D Systems,
Wiesbaden-Nordenstadt, Germany) was used as a negative control in
this study.
Immunocytochemistry in human melanoma
cell lines and adult HEMs
GPX3 expression was determined in both HEMs and
melanoma cell lines by immunocytochemistry. Cell lines were plated
on glass coverslips containing 6-well plates on the day before
staining. After fixation with 95% ethanol, the cells were blocked
with 5% bovine serum antigen (BSA) at RT for 1 h. Endogenous
peroxidase activity was inactivated with a mixture of
H2O2 and methanol, in a ratio of 1:40. The
cells were then treated with primary antibody against GPX3 (working
dilution: 1:200; Abcam) at RT for 1 h, and the Real™EnVision™HRP
Rabbit/Mouse detection system (Dako North America, Inc.) was used
as the secondary antibody. After visualization with
3,3′-diaminobenzidine, counterstaining was performed using
hematoxylin. HeLa cells were used as a positive control for
GPX3.
Total RNA extraction and analysis for
GPX3 mRNA expression
Total RNA was isolated with TRIzol reagent (for cell
line samples; Invitrogen, Carlsbad, CA, USA) and the RNeasy FFPE
kit (from paraffin tissue samples; Qiagen GmbH, Hilden, Germany),
and cDNA was synthesized using total RNA and AccuPower®
RT PreMix (Bioneer, Seoul, Korea) according to the manufacturer's
protocol. qRT-PCR was performed as previously described using 1X
SYBR-Green Master Mix (Applied Biosystems, Foster City, CA, USA)
and synthesized cDNA in an ABI7500 Real-time PCR system (Applied
Biosystems) under the following conditions: initial denaturation
for 10 min at 95°C, followed by 40 cycles of 95°C for 20 sec, 50°C
for 30 sec, and 72°C for 45 sec. Conventional RT-PCR analysis was
performed using AccuPower PCRPreMix (Bioneer) and the synthesized
cDNA with an annealing temperature of 58°C. β-actin was used as the
housekeeping gene. Oligonucleotide primers used for the PCR were:
5′-CAACCAATTTGGAAAACAGG-3′ and 5′-GTGGGAGGACAGGAGTTCTT-3′ for GPX3;
and 5′-ATAGCACAGCCTGGATAGCAACGTAC-3′ and
5′-CACCTTCTACAATGAGCTGCGTGTG-3′ for β-actin (29).
Treatments with 5-aza-2′-deoxycytidine
(5-Aza)
Two melanoma cell lines (SK-MEL-24 and SK-MEL-2)
were treated with 5-Aza (Sigma-Aldrich, Inc., St. Louis, MO, USA),
1 or 10 µM, for 72 h, with drug replacement every 24 h.
Cells were then harvested and mRNA expression of GPX3 was
detected in each group of the cells.
DNA extraction and MSP
Genomic DNA extraction was performed in HEMs and
melanoma cell lines, three normal skin samples, and 10 melanoma
tissue samples using the QIAamp DNA mini kit (Qiagen GmbH).
Modification of DNA was performed with the EZ DNA Methylation kit
(Zymo Research Corp., Irvine, CA, USA), and the
methylation-specific or unmethylation-specific amplification was
performed with an annealing temperature of 59°C using the following
primers: 5′-TATGTTATTGTCGTTTCGGGAC-3′ and
5′-GTCCGTCTAAAATATCCGACG-3′ for methylation-specific amplification;
and 5′-TTTATGTTATTGTTTTGGGATG-3′ and
5′-ATCCATCTAAAATATCCAACACTCC-3′ for unmethylation-specific
amplification. The amplified DNA products were loaded on 2% agarose
gels for electrophoresis and stained with ethidium bromide.
Cell proliferation assay
To determine the influence of GPX3 on the
proliferative ability of the melanoma cells, the trypan blue
exclusion assay was performed in GPX3-depleted SK-MEL-24
(GPX3Δ) cells and cells transfected with the control
vector (Mock). Cells were seeded in 6-well plates and the number of
cells in each group was counted at 36 and 72 h using a
hemocytometer.
Cell migration assay
To evaluate the influence of GPX3 on the migration
ability of the melanoma cells, a wound healing assay was performed
in GPX3Δ- and Mock-SK-MEL-24 cells. The cells were
seeded in 24-well culture plates in triplicate. After 24 h of
culturing, linear scrape injuries were made with a yellow pipette
tip on the cells that displayed growth to a confluent monolayer.
Wound closing in each group was photographed at the indicated time
points and analyzed with the Image J analysis software package
(National Institutes of Health, Bethesda, MD, USA).
Cell invasion assay
To evaluate the influence of GPX3 on the
invasiveness of the melanoma cells, an invasion assay was performed
using Transwell filters (pore size, 8 µm; BD Biosciences,
Bedford, MA, USA) in GPX3Δ- and Mock-SK-MEL-24 cells.
Both the lower and top sides of Transwell filters were coated with
8 µg/µl Matrigel (BD Biosciences, San Jose, CA, USA)
and the filters were placed on 24-well culture plates with a
culture medium with 15% BSA. The cells were suspended in a medium
containing 3% BSA and then seeded in the coated Transwell chambers.
After 36 h of culturing, the bottom side of the membrane with the
invaded cells was fixed with 4% paraformaldehyde and then stained
with crystal violet. The number of invaded cells was counted with a
microscope and comparatively investigated between GPX3Δ-
and Mock-SK-MEL-24 cells.
Statistical analysis
In this study, we used commercially available
software (SPSS version 15.0 for Windows; SPSS, Inc., Chicago, IL,
USA) for statistical analysis. The Mann-Whitney U test was used to
analyze differences between groups in the in vitro studies.
The χ2 test and Fisher's exact test were used to
investigate differences between groups of clinical samples.
Survival analysis for melanoma patients was performed using
Kaplan-Meier analysis, and the differences were compared using the
log-rank test. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
GPX3 is downregulated in melanoma cell
lines by promoter hypermethylation
In the current study, both mRNA and protein
expression of GPX3 were comparatively investigated in HEMs and
melanoma cell lines. Our results showed that GPX3 expression was
downregulated in melanoma cell lines to a greater extent than in
HEMs on both the mRNA and protein levels (Fig. 1B and C). The methylation status of
GPX3 was evaluated in both HEMs and melanoma cell lines
using MSP primers against the GPX3 promoter. Full
methylation (presence of only methylated CpGs) was found in the
SK-MEL-2 cell line, which was negative for GPX3 on both the RNA and
protein levels. Moreover, HEMs and SK-MEL-24 cells, with expression
of GPX3 on both the RNA and protein levels, showed unmethylation
(presence of only unmethylated CpGs) and partial methlylation
(presence of both methylated and unmethylated CpGs) for GPX3,
respectively (Fig. 1E). To
investigate whether methylation status was related to repression of
GPX3 expression, melanoma cell lines were treated with the DNA
demethylation agent 5-Aza, and our results showed that
demethylation of the GPX3 gene by 5-Aza in both SK-MEL-2 and
SK-MEL-24 cell lines restored GPX3 expression at mRNA and protein
levels (Fig. 1D and F). No change
was found in GPX3 expression of HEMs according to the
presence or absence of 5-Aza treatment (Fig. 1D).
GPX3 promoter is methylated in primary
melanoma tissues
The methylation status of GPX3 was also
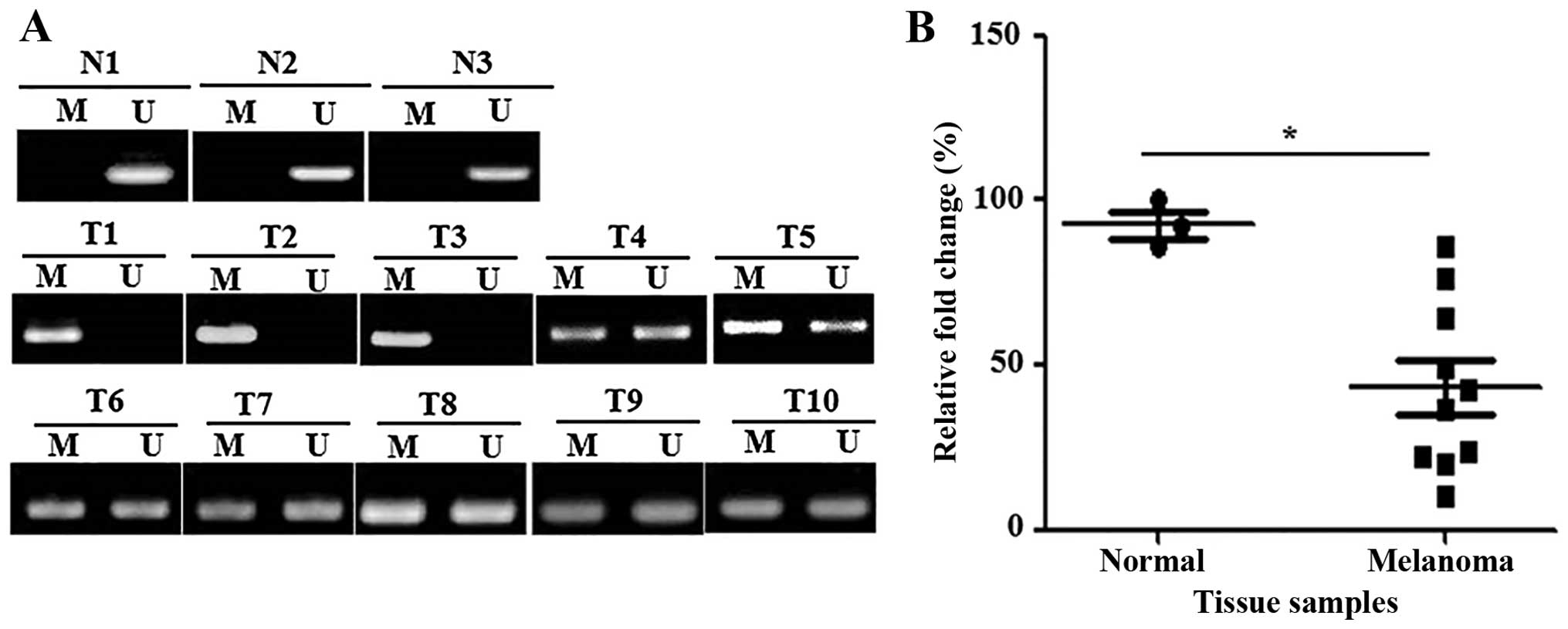
determined in 3 normal skin and 10 primary melanoma tissue samples.
Unmethylation for GPX3 was found in the 3 normal skin samples. By
contrast, full methylation and partial methylation were detected in
three and seven melanoma tissues samples, respectively (Fig. 2A). Moreover, RT-PCR analysis showed
that GPX3 mRNA expression was significantly decreased in
primary melanoma tissues compared to normal skin tissues (Fig. 2B).
The influence of GPX3 downregulation on
the biological behavior of SK-MEL-24
SK-MEL-2 showed increased invasion ability than
SK-MEL-24 cells, which was found to be negative for GPX3 on both
RNA and protein levels (data not shown). However, SK-MEL-2 and
SK-MEL-24 cells may have a different molecular basis. Therefore,
the differences in invasive ability between the cells cannot
exactly represent the role of GPX3. We also found that invasive
activities were suppressed to a greater extent in SK-MEL-2 and
SK-MEL-24 cells with 5-Aza treatment than without 5-Aza treatment
(data not shown). However, no change was found in HEMs according to
the presence or absence of 5-Aza treatment. Other tumor suppressor
genes in addition to GPX3 have been found to be methylated in
melanoma (30). Consequently, 5-Aza
treatment can restore the RNA and protein expression of various
genes that are methylated in melanoma. Therefore, we constructed
GPX3-depleted cell lines to investigate the influence of GPX3 on
the biological behavior of melanoma cells.
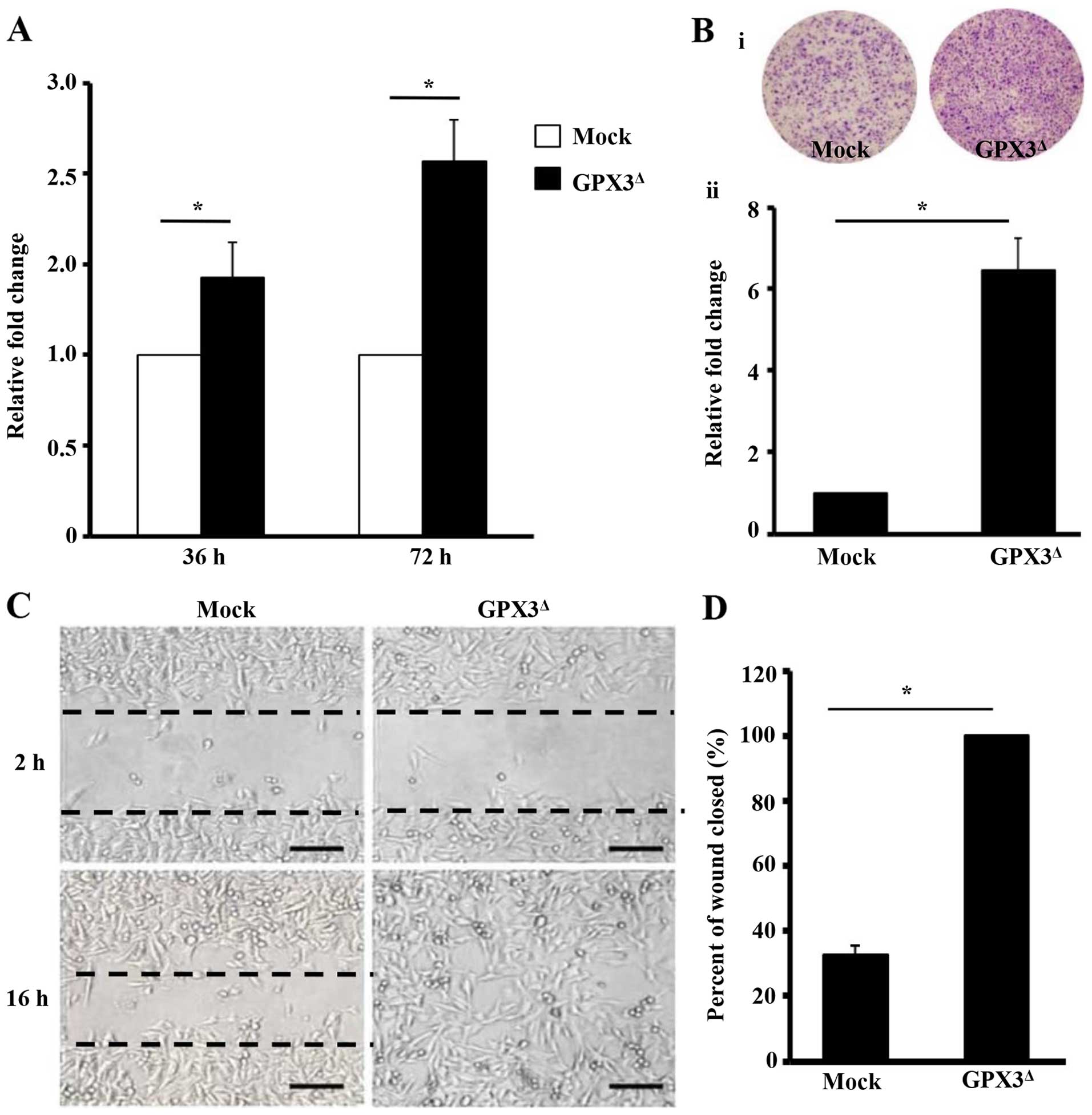
The proliferative, migration, and invasive ability
of cells were comparatively investigated in the GPX3Δ-
and Mock-SK-MEL-24cells. Compared to Mock-SK-MEL-24 cells,
GPX3Δ-SK-MEL-24 cells demonstrated 1.43- and 2.07-fold
higher proliferative ability in each time point, respectively
(P<0.001, and P<0.001, respectively). Moreover,
GPX3Δ-SK-MEL-24 cells showed 3.06-fold (P<0.001), and
6.47-fold (P<0.001) higher migration and invasive ability,
respectively (Fig. 3A–D).
Clinicopathological significance of GPX3
expression in melanoma patients
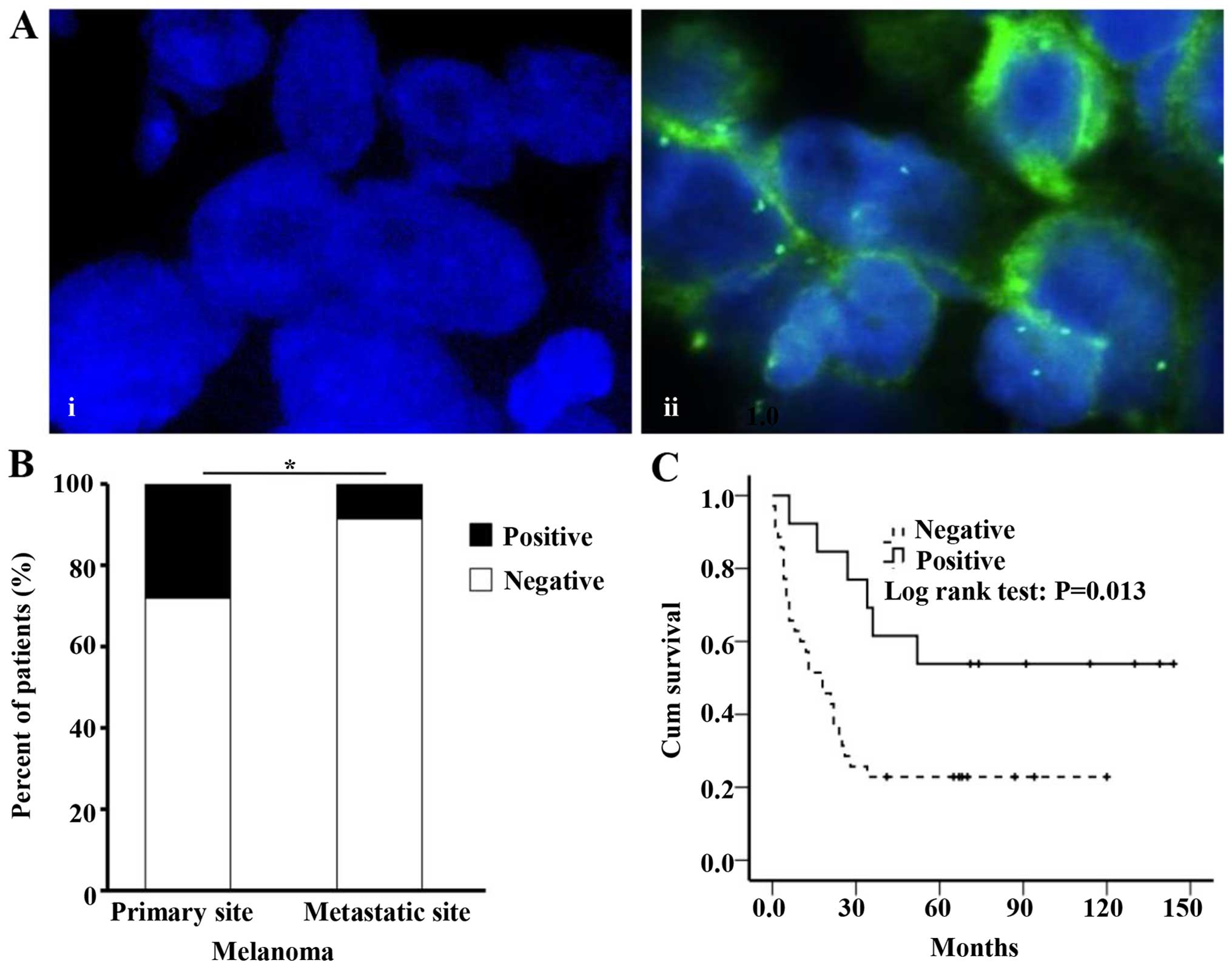
GPX3 expression was detected in the cytoplasm of
cancer cells in melanoma tissue samples (Fig. 4A). The expression frequency of GPX3
was significantly increased in primary sites (33.8%) compared to
metastatic sites (10.8%) of melanoma tissues (P=0.036) (Fig. 4B). Kaplan-Meier analysis was
performed in 46 primary malignant melanoma patients. We found that
patients with GPX3 expression showed increased survival rates
(median survival duration 71 months) compared to patients without
GPX3 expression (median survival duration 18 months) (P=0.013)
(Fig. 4C). No significant
association was found between GPX3 expression and age or gender of
melanoma patients.
Discussion
The antioxidant system is the first line of defense
against ROS-induced damage. GPX3, a major antioxidative enzyme, is
known as a critical scavenger of ROS. Silencing of GPX3 promotes
ROS production in muscle stem cells and colon cancer cell lines
(31,32). GPX3 is selectively expressed in
several healthy tissues, such as the esophagus, stomach, and colon
(26). In the current study, GPX3
expression was detected in the cytoplasm of HEMs, but decreased or
negative expression was found in melanoma cell lines. Moreover,
unmethylation was found in HEMs, while full or partial methylation
was observed in melanoma cell lines. Methylation of GPX3 was also
frequently detected in the human melanoma tissue samples, and mRNA
expression was concurrently repressed in melanoma tissue samples,
in the present study. Supportively, 5-Aza treatment can restore the
expression of GPX3 in melanoma cell lines. 5-Aza inhibits
DNA-methyltransferase activity and thereby restores the expression
of silenced genes (33).
Downregulation and silencing of GPX3 in melanoma may result from
promoter hypermethylation.
Many studies have shown that GPX3 is frequently
methylated and downregulated in cancers compared to related normal
tissues, and it has been hypothesized that GPX3 may act as a tumor
suppressor in cancers. However, the effect of GPX3 on the biologic
behavior of cancer cells has not been fully investigated (11). Some investigators showed that GPX3
overexpression can promote cancer cell motility but not the growth
of cancer cells in gastric cancer (34). In prostate cancer cell lines, GPX3
overexpression inhibits the invasiveness of cells in vitro,
and can reduce tumor volume and prevent metastasis in vivo
(27). In the current study, we
found that GPX3 downregulation can promote the proliferation,
motility, and invasion of melanoma cells in vitro. In
support of these observations, negative expression of GPX3 was more
frequently detected in metastatic site than primary site of
melanoma and was also found to be a negative prognostic indicator
for melanoma patients in the present study. The clinical
implications of GPX3 have also been mentioned by other
investigators. In ovarian clear cell adenocarcinoma, GPX3
expression is related to cisplatin sensitivity in patients.
Moreover, downregulation and promoter methylation of GPX3 were
detected more often in gastric cancer patients with lymph node
metastasis. Downregulation of GPX3 was also related to poor
prognosis in patients with various cancers such as multiple
myeloma, gastric cancer, and gallbladder cancer (35–37).
Loss of expression of Nkx3.1, a well known tumor suppressor gene in
prostate cancer, can induce increased oxidative damage in prostate
carcinogenesis. In Nkx3.1 knockout mice, GPX3 expression strongly
increased during conversion from normal prostate epithelium to
prostatic intraepithelial neoplasia (PIN), and this expression is
mainly found in tumor cells of PIN lesions. However, the loss of
PTEN that is induced by increased oxidative damage can decrease
GPX3 expression during the transformation from PIN to cancer in
Nkx3.1 knockout mice (38). As in
other cancers, loss of GPX3 expression may also have tumor
suppressor functions in melanoma carcinogenesis and progression via
failures in the cellular antioxidant system.
The epigenetic inactivation of GPX3 may be a
crucial mechanism in the pathogenesis of melanoma, and GPX3 may
serve as a possible predictive or prognostic biomarker and
therapeutic target for melanoma patients.
Acknowledgments
This work was supported by National Natural Science
Foundation of China (NSFC, no. 81560503) and Basic Science Research
Program through the National Research Foundation of Korea (NRF)
funded by the Ministry of Education, Science and Technology
(2011-0022376).
References
|
1
|
Dreher D and Junod AF: Role of oxygen free
radicals in cancer development. Eur J Cancer. 32A:30–38. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cooke MS, Evans MD, Dizdaroglu M and Lunec
J: Oxidative DNA damage: Mechanisms, mutation, and disease. FASEB
J. 17:1195–1214. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Malins DC, Polissar NL and Gunselman SJ:
Progression of human breast cancers to the metastatic state is
linked to hydroxyl radical-induced DNA damage. Proc Natl Acad Sci
USA. 93:2557–2563. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cerutti PA: Prooxidant states and tumor
promotion. Science. 227:375–381. 1985. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Sies H: Oxidative stress: Oxidants and
antioxidants. Exp Physiol. 82:291–295. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dröge W: Free radicals in the
physiological control of cell function. Physiol Rev. 82:47–95.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tham DM, Whitin JC, Kim KK, Zhu SX and
Cohen HJ: Expression of extracellular glutathione peroxidase in
human and mouse gastrointestinal tract. Am J Physiol.
275:G1463–G1471. 1998.PubMed/NCBI
|
|
8
|
Björnstedt M, Xue J, Huang W, Akesson B
and Holmgren A: The thioredoxin and glutaredoxin systems are
efficient electron donors to human plasma glutathione peroxidase. J
Biol Chem. 269:29382–29384. 1994.PubMed/NCBI
|
|
9
|
Yamamoto Y and Takahashi K: Glutathione
peroxidase isolated from plasma reduces phospholipid
hydroperoxides. Arch Biochem Biophys. 305:541–545. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Esworthy RS, Chu FF, Paxton RJ, Akman S
and Doroshow JH: Characterization and partial amino acid sequence
of human plasma glutathione peroxidase. Arch Biochem Biophys.
286:330–336. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Brigelius-Flohé R and Kipp A: Glutathione
peroxidases in different stages of carcinogenesis. Biochim Biophys
Acta. 1790:1555–1568. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yoshimura S, Suemizu H, Taniguchi Y,
Watanabe K, Nomoto Y, Katsuoka Y, Arimori S and Moriuchi T:
Molecular cloning of human plasma glutathione peroxidase gene and
its expression in the kidney. Nucleic Acids Symp Ser. (25):
163–164. 1991.PubMed/NCBI
|
|
13
|
Yoshimura S, Suemizu H, Taniguchi Y,
Arimori K, Kawabe N and Moriuchi T: The human plasma glutathione
peroxidase-encoding gene: Organization, sequence and localization
to chromosome 5q32. Gene. 145:293–297. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Takebe G, Yarimizu J, Saito Y, Hayashi T,
Nakamura H, Yodoi J, Nagasawa S and Takahashi K: A comparative
study on the hydroperoxide and thiol specificity of the glutathione
peroxidase family and selenoprotein P. J Biol Chem.
277:41254–41258. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Comhair SA and Erzurum SC: The regulation
and role of extracellular glutathione peroxidase. Antioxid Redox
Signal. 7:72–79. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baker RD, Baker SS, LaRosa K, Whitney C
and Newburger PE: Selenium regulation of glutathione peroxidase in
human hepatoma cell line Hep3B. Arch Biochem Biophys. 304:53–57.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Clark LC, Combs GF Jr, Turnbull BW, Slate
EH, Chalker DK, Chow J, Davis LS, Glover RA, Graham GF, Gross EG,
et al Nutritional Prevention of Cancer Study Group: Effects of
selenium supplementation for cancer prevention in patients with
carcinoma of the skin. A randomized controlled trial. JAMA.
276:1957–1963. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Avissar N, Ornt DB, Yagil Y, Horowitz S,
Watkins RH, Kerl EA, Takahashi K, Palmer IS and Cohen HJ: Human
kidney proximal tubules are the main source of plasma glutathione
peroxidase. Am J Physiol. 266:C367–C375. 1994.PubMed/NCBI
|
|
19
|
Schomburg L and Köhrle J: On the
importance of selenium and iodine metabolism for thyroid hormone
biosynthesis and human health. Mol Nutr Food Res. 52:1235–1246.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Köhrle J: Selenium and the control of
thyroid hormone metabolism. Thyroid. 15:841–853. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lapointe J, Kimmins S, Maclaren LA and
Bilodeau JF: Estrogen selectively up-regulates the phospholipid
hydroperoxide glutathione peroxidase in the oviducts.
Endocrinology. 146:2583–2592. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hoffenberg EJ, Deutsch J, Smith S and
Sokol RJ: Circulating antioxidant concentrations in children with
inflammatory bowel disease. Am J Clin Nutr. 65:1482–1488.
1997.PubMed/NCBI
|
|
23
|
Comhair SA, Bhathena PR, Farver C,
Thunnissen FB and Erzurum SC: Extracellular glutathione peroxidase
induction in asthmatic lungs: Evidence for redox regulation of
expression in human airway epithelial cells. FASEB J. 15:70–78.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tham DM, Whitin JC and Cohen HJ: Increased
expression of extracellular glutathione peroxidase in mice with
dextran sodium sulfate-induced experimental colitis. Pediatr Res.
51:641–646. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Iwata K, Nishinaka T, Matsuno K and
Yabe-Nishimura C: Increased gene expression of glutathione
peroxidase-3 in diabetic mouse heart. Biol Pharm Bull.
29:1042–1045. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mörk H, Lex B, Scheurlen M, Dreher I,
Schütze N, Köhrle J and Jakob F: Expression pattern of
gastrointestinal selenoproteins - targets for selenium
supplementation. Nutr Cancer. 32:64–70. 1998. View Article : Google Scholar
|
|
27
|
Yu YP, Yu G, Tseng G, Cieply K, Nelson J,
Defrances M, Zarnegar R, Michalopoulos G and Luo JH: Glutathione
peroxidase 3, deleted or methylated in prostate cancer, suppresses
prostate cancer growth and metastasis. Cancer Res. 67:8043–8050.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Jemal A, Thomas A, Murray T and Thun M:
Cancer statistics, 2002. CA Cancer J Clin. 52:23–47. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang X, Zheng Z, Yingji S, Kim H, Jin R,
Renshu L, Lee DY, Roh MR and Yang S: Downregulation of glutathione
peroxidase 3 is associated with lymph node metastasis and prognosis
in cervical cancer. Oncol Rep. 31:2587–2592. 2014.PubMed/NCBI
|
|
30
|
Luo X, Wei B, Chen A, Zhao H, Huang K and
Chen J: Methylation-mediated loss of SFRP2 enhances melanoma cell
invasion via Wnt signaling. Am J Transl Res. 8:1502–1509.
2016.PubMed/NCBI
|
|
31
|
El Haddad M, Jean E, Turki A, Hugon G,
Vernus B, Bonnieu A, Passerieux E, Hamade A, Mercier J,
Laoudj-Chenivesse D, et al: Glutathione peroxidase 3, a new
retinoid target gene, is crucial for human skeletal muscle
precursor cell survival. J Cell Sci. 125:6147–6156. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Barrett CW, Ning W, Chen X, Smith JJ,
Washington MK, Hill KE, Coburn LA, Peek RM, Chaturvedi R, Wilson
KT, et al: Tumor suppressor function of the plasma glutathione
peroxidase gpx3 in colitis-associated carcinoma. Cancer Res.
73:1245–1255. 2013. View Article : Google Scholar :
|
|
33
|
Cameron EE, Bachman KE, Myöhänen S, Herman
JG and Baylin SB: Synergy of demethylation and histone deacetylase
inhibition in the re-expression of genes silenced in cancer. Nat
Genet. 21:103–107. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Peng DF, Hu TL, Schneider BG, Chen Z, Xu
ZK and El-Rifai W: Silencing of glutathione peroxidase 3 through
DNA hyper-methylation is associated with lymph node metastasis in
gastric carcinomas. PLoS One. 7:e462142012. View Article : Google Scholar
|
|
35
|
Zhang X, Yang JJ, Kim YS, Kim KY, Ahn WS
and Yang S: An 8-gene signature, including methylated and
down-regulated glutathione peroxidase 3, of gastric cancer. Int J
Oncol. 36:405–414. 2010.PubMed/NCBI
|
|
36
|
Yang ZL, Yang L, Zou Q, Yuan Y, Li J,
Liang L, Zeng G and Chen S: Positive ALDH1A3 and negative GPX3
expressions are biomarkers for poor prognosis of gallbladder
cancer. Dis Markers. 35:163–172. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Kaiser MF, Johnson DC, Wu P, Walker BA,
Brioli A, Mirabella F, Wardell CP, Melchor L, Davies FE and Morgan
GJ: Global methylation analysis identifies prognostically important
epigenetically inactivated tumor suppressor genes in multiple
myeloma. Blood. 122:219–226. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ouyang X, DeWeese TL, Nelson WG and
Abate-Shen C: Loss-of-function of Nkx3.1 promotes increased
oxidative damage in prostate carcinogenesis. Cancer Res.
65:6773–6779. 2005. View Article : Google Scholar : PubMed/NCBI
|