Introduction
Angiogenesis is an important process during tumor
growth (1). As the new blood
vessels provide oxygen and nutrients, it is difficult for tumors to
grow beyond 1–2 mm3 in size without tumor angiogenesis
(2,3). After angiogenesis, the previously
dormant tumors start to grow rapidly and begin to invade
surrounding tissues or transfer to distant sites. The balance
between pro-angiogenic molecules (e.g. VEGF, FGF or EGF) and
anti-angiogenic molecules (e.g. angiostatin, endostatin or
thrombospondin) decides the time and site where angiogenesis occurs
(4,5). Sunitinib is a second-generation
multi-targeted receptor tyrosine kinase inhibitor, which has been
approved for the treatment of renal cell carcinoma and
gastrointestinal stromal tumors (6,7). It
inhibits angiogenesis by cutting-off the signal transduction of
VEGF that is the main growth factor during tumor angiogenesis.
Integrins are the main targets for anti-angiogenic molecules such
as endostatin. Being cell adhesion molecules, integrins are
involved in a wide range of cell-ECM and cell-cell interactions
(8,9–11). As
integrin signaling is important for tumor growth, angiogenesis and
metastasis, cutting-off integrin signaling is a promising treatment
strategy for cancer treatment (9,11).
Integrin αvβ3 is a highly expressed integrin in various tumor cells
or activated endothelial cells whereas its expression on the rest
of endothelial cells in normal organs remains low (12). Treatment of integrin αvβ3 with
monoclonal antibodies, cyclic RGD peptides or peptidemimetics
induced endothelial cell apoptosis (13) and angiogenesis inhibition (11). HM-3 is an RGD modified
endostatin-derived synthetic peptide that targets integrin αvβ3
(14). It inhibited endothelial
cell migration and angiogenesis in vitro and inhibited tumor
growth in vivo (15).
Pharmacodynamic studies confirmed that the in vitro and
in vivo antitumor activity of HM-3 was dose-dependent in the
concentration range of 0.75–3 mg/kg whereas further increase of
HM-3 dosage did not have a higher tumor growth inhibition (16), however, the reason for this has not
been investigated.
In the present study, three tumor cell lines were
used to determine their expression levels of integrin αv, β3, α5,
β1 subunits by a western blotting technique. Flow cytometric assays
were carried out for detection of the adhesion of FITC-HM-3 to the
selected tumor cell lines. Furthermore, the in vitro
dose-efficacy relationship of HM-3 was investigated by a Transwell
cell migration procedure. Moreover, in vivo imaging was
carried out for determination of the drug distribution and
tumor-targeting effects in BALB/c-nu nude mice. In addition, the
in vivo dose-efficacy relationship was investigated in
BALB/c-nu nude mice xenografted with HCT-116 cells that were highly
inhibited by HM-3 in a cell migration assay. Immunohistochemistry
assays for evaluation of expression levels of angiogenesis-related
factors, including CD31, HIF-1α and VEGF, were performed. These
studies tried to explain the relationship of tumor
microenvironments and the antitumor effect of an angiogenesis
inhibitor HM-3. These studies helped to itinerate that HM-3
inhibited angiogenesis and tumor growth by directly inhibiting
HUVEC migration and the bell-shaped dose-efficacy relationship
should be explained on a molecular level by focusing on the HM-3
special dose-efficacy relationship on HUVEC migration (16).
Materials and methods
Materials
Sunitinib (>99% purity) was purchased from
Melonepharma with a Cas no. 341031-54-7. FITC-HM-3 with 98.3%
purity was obtained from GL Biochem Ltd. (Shanghai, China) (catalog
no. 340664). HM-3 with 99.5% purity was synthesized by GL Biochem
Ltd. (catalog no. 140214-2). All laboratory chemicals were of
molecular biology grade. Tumor cell lines were obtained from
Shanghai Cell Biology Institutes (Shanghai, China).
Cell cultures
HCT-116 colorectal cancer cells were cultured in
McCoy's 5A medium containing 50 IU/ml penicillin, 100 μg/ml
streptomycin, 2,200 μg/ml NaHCO3 and 10% (v/v)
fetal bovine serum (FBS) (Gibco). SMMC-7721 and Hep G-2
hepatocellular carcinoma cells were cultured in Dulbecco's modified
Eagle's medium (DMEM) containing 50 IU/ml penicillin, 100
μg/ml streptomycin, 2,000 μg/ml NaHCO3 and
10% (v/v) FBS. All cells were incubated at 37°C in a humidified
atmosphere of 5% CO2 (17) while MDA-MB-231 were grown in the
absence of CO2. Cells were detached through incubation
with trypsin/EDTA and sub-cultured every 2–5 days (18).
Western blot assays
Equal quantities of extracted proteins were loaded
on a 10% SDS-polyacrylamide gel and were electrophoretically
separated. The separated proteins in the gel were electrically
transferred to a polyvinylidene fluoride (PVDF) membrane (Roche),
according to the procedure described by Laemmli (19). Membranes were blocked with 5%
defatted milk in Tris-buffered saline (TBS) with 0.5% Tween-20 for
30 min. Then, the membrane was incubated with a primary antibody
overnight at 4°C. The first antibodies included rabbit monoclonal
anti-αv integrin (catalog no. 4711S); rabbit monoclonal anti-β3
integrin (catalog no. 4702); mouse monoclonal anti-β1 integrin
(catalog no. 4706S) (Cell Signaling Technology); rabbit polyclonal
IgG for α5 integrin (catalog no. 130609) (Santa Cruz
Biotechnology). β-actin was used as the internal control. After
incubation with the primary antibody, membranes were washed twice
for 1 min and once for 10 min with TBS containing 0.5% Tween-20,
and were then incubated with goat anti-rabbit IgG with horseradish
peroxidase-conjugated (catalog no. 3223449; Biotech, Manufacturing
Co. Ltd.) or goat anti-mouse IgG with horseradish
peroxidase-conjugated (catalog no. 422331022; Bioss, Beijing,
China) at 25°C for 45 min. After incubation with the secondary
antibody, membranes were washed three times with TBS with 0.5%
Tween-20 for 10 min. Finally, development and detection of bands
were carried out by chemiluminescence (20,21).
Briefly, the membranes were incubated with enhanced
chemiluminescence detection reagents with SuperSignal West Pico
Chemiluminescent Substrate (Pierce) for 5 min, and quantification
was carried out using ImageJ software (22).
Flow cytometric assays
The adhesion of FITC-HM-3 to three tumor cell lines
was evaluated by the flow cytometry technique, a method described
by Janouskova et al (22)
with minor modifications. Briefly, the cultured tumor cells were
spread on a 6-well plate and were incubated at 37°C overnight.
After digestion and wash with phosphate-buffered saline (PBS), 1 ml
PBS (containing 1% BSA) was used to resuspend the cells with a
density of 1×106 cells/ml. The cells were incubated with
10 μl of 1 mg/ml FITC-HM-3 at 4°C for 30 min in the dark.
Then, the cells were collected by centrifugation at 800 rpm for 5
min. After another wash with PBS, the cells were re-suspended in
800 μl PBS for flow cytometric detection. The mean
fluorescence intensity that characterized drug affinity to
integrins αvβ3 was measured using the FlowJo software version
7.6.1.
Transwell cell migration
During this assay, cells were placed in the upper
layer of a cell permeable membrane in presence of 0.5% BSA and
HM-3. The well below the cell permeable membrane was filled with
cell medium that contained 5% FBS. Cell migration was performed for
18 h. Then, migrated cells were stained and counted. The main
advantage of this assay was the need of low levels of angiogenic
inducers. Images were captured and the migration inhibition rate
(MIR) was obtained, following the equation: MIR% = (1 −
Ntest/Ncontrol) × 100%, in which MIR is the
migration inhibition rate, Ntest the number of cells in
test samples and Ncontrol the number of cells in the control
sample.
In vivo imaging
In consideration of the above mentioned results in
pre-experiment, in vivo image was carried out to identify
drug distribution and tumor targeting effect in mice. Two female
BALB/c nude mice (6 weeks and 20 g) were subcutaneously injected
with HCT-116 (1×106 cell/ml) (22). Another two mice were tumor-free as
controls. After tumor grew to 300 mm3, detection was
performed in which mice were intravenously injected with FITC-HM-3
(6 mg/kg) and anesthetized with isoflurane inhaler. Images were
captured by Caliper IVIS Spectrum system (Caliper Life Sciences,
Waltham, MA, USA) with excitation wavelength of 490 nm and emission
wavelength of 520 nm. Images were captured at indicated time
points.
Dose-efficacy relationship of HM-3 on an
HCT-116 xenograft model in nude mice and the expression levels of
angiogenesis-related factors in tumor tissues
Animal experiments were carried out in female BALB/c
nude mice (5 weeks and 18–21 g). All mice were subcutaneously
injected in the right flank with HCT-116 cells (a density of
1×106 cell/ml and a total volume of 0.2 ml). After tumor
volume reached 70–100 mm3, mice were put into different
groups, which included the negative control group (n=12) that were
intravenously injected with 0.2 ml normal saline every day for 21
days; positive controls (n=6) that were fed by oral gavage with 0.2
ml sunitinib (60 mg/kg) every day for 17 days; mice in the three
HM-3 groups (n=6) were intravenously injected with HM-3 (3, 12 or
48 mg/kg) every day for 21 days (Table
I). During the treatment period tumor volume and body weight of
mice were measured every two days for determination of drug
efficacy and signs of toxicity. Animals were sacrificed after 21
days from the first injection and the tumor mass was extracted,
dissected, blotted on filter paper and immediately weighed.
Immunohistochemistry (IHC) was performed where (CD31, HIF-1α and
VEGF) antibodies were used for detection of tumor angiogenesis.
Tumors were immediately fixed in 4% formaldehyde and embedded in
paraffin, then, sectioned for immunohistochemical staining of CD31,
HIF-1α and VEGF with rabbit anti-CD31, HIF-1α and VEGF polyclonal
antibodies (Beijing Zhongshan Golden Bridge Biotechnology Co.,
Ltd.). Briefly, staining for the mentioned angiogenesis factors was
performed on sections that were incubated with their specific
primary antibodies, biotinylated goat anti-rabbit secondary
antibodies, horseradish peroxidase-labeled streptavidin, and the
results were visualized with diaminobenzidine (DAB) chromogen.
Images were captured under a microscope with a magnification of
×200 (23).
 | Table IExperimental strategy for tumor
inhibitory effect of HM-3. |
Table I
Experimental strategy for tumor
inhibitory effect of HM-3.
| Group | No. | Drug | Dosage |
|---|
| G1 | 12 | Normal saline | 0.2 ml/day/mouse,
IV for 21 days |
| G2 | 6 | Sunitinib | 60 mg/kg, 0.2
ml/day/mouse, IG for 17 days |
| G3 | 6 | HM-3 | 3 mg/kg, 0.2
ml/day/mouse, IV for 21 days |
| G4 | 6 | HM-3 | 12 mg/kg, 0.2
ml/day/mouse, IV for 21 days |
| G5 | 6 | HM-3 | 48 mg/kg, 0.2
ml/day/mouse, IV for 21 days |
Ethics statement
The experiments involving animals conformed to the
ethical standards of China Pharmaceutical University and the care
of animals was in accordance with the guidelines of care and use of
laboratory animals of China Pharmaceutical University. In
vivo imaging was performed under isoflurane anesthesia, and all
efforts were carried out to minimize suffering.
Statistical analysis
The data were analyzed using the statistics software
SPSS statistics 17.0 (Softonic, San Francisco, CA, USA) and are
expressed as mean ± SD. Statistical significance was assessed using
the Student's t-test. P<0.05 was considered statistically
significant; P<0.01 was considered statistically very
significant.
Results
Integrin expression and FITC-HM-3 binding
to three tumor cell lines
The expression levels of integrin subunit αv, β3, α5
and β1 on three human tumor cell lines were detected by western
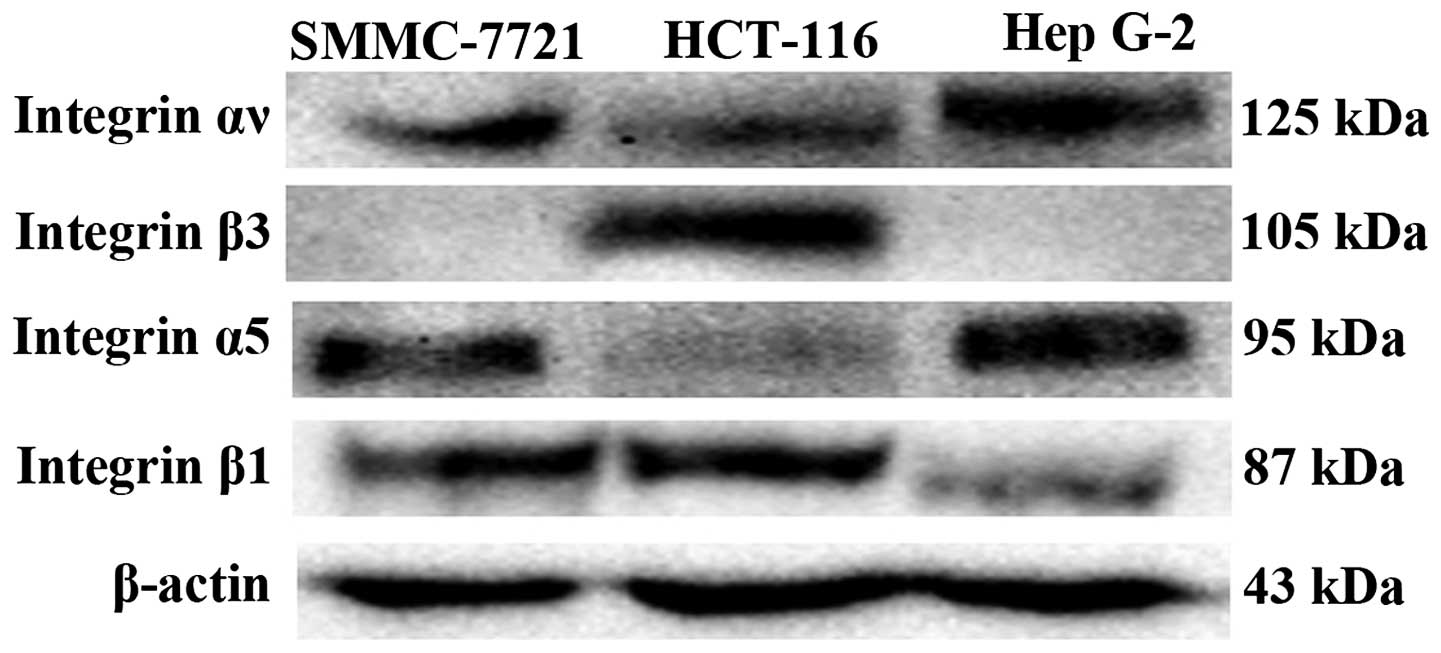
blot assays. As shown in Fig. 1,
integrin αv and β1 were expressed on HCT-116, Hep G-2 and SMMC-7721
cells and integrin α5 was expressed on Hep G-2 and SMMC-7721 cells.
Integrin β3 was expressed on HCT-116 cells (Fig. 1). Only HCT-116 expressed a
substantial amount of integrin αvβ3. Hep G-2 and SMMC-7721
expressed a substantial amount of αv, but trace mount of β3,
probably they had integrin αvβ5. All three cell lines obviously
expressed integrin subunit β1 whereas only Hep G-2 and SMMC-7721
expressed a substantial amount of α5. HCT-116 did express α5 only
slightly, probably it expressed other integrin subunits as β1
subunit can combine with various α subunits. These results were in
line with a previous study (24).
Similar study was performed to detect the expression levels of
integrin subunits on HUVECs and other human tumor cell lines (A549,
MCF-7, HeLa, BEL-7402, MGC-803, HT-29, MDA-MB-231 and U87). Various
cells (e.g. A549, MCF-7 and HeLa) expressed high levels of integrin
αvβ3 and α5β1 whereas HT-29 expressed low levels of integrin αvβ3
and α5β1 (data not shown). This difference in integrin expression
levels may influence the antitumor effect of anti-angiogenic
reagents that use integrins as targets.
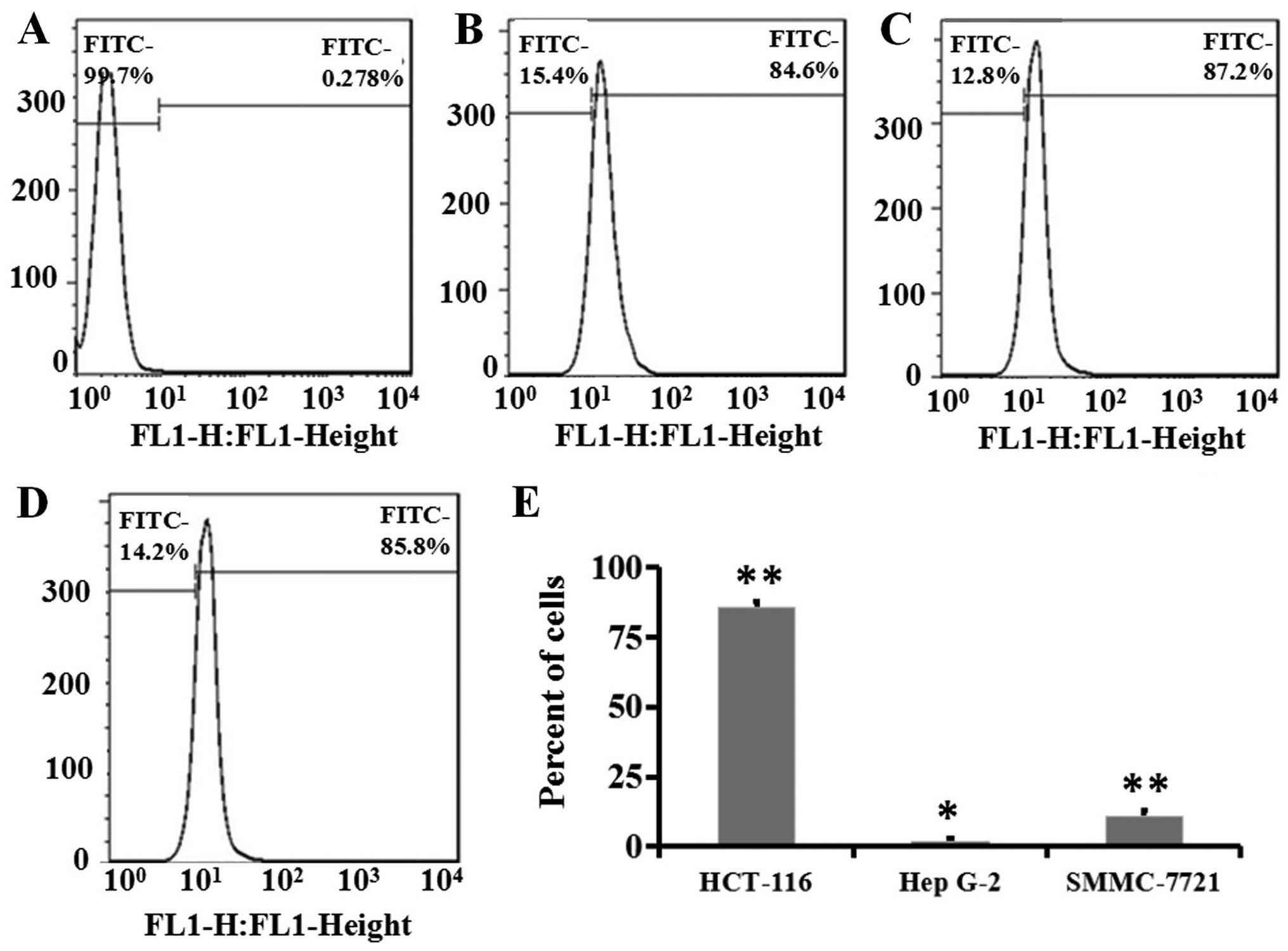
In the flow cytometric assays, the fluorescence of
cells in the control group that were treated with free FITC
molecules was used to define the 'gate' and 99.7% cells with
autofluorescence were included in this gate (Fig. 2A). After FITC-HM-3 incubation, an
average of 85.9% of HCT-116 cells bound with FITC-HM-3, the
fluorescence shifted rightward and out of the gate (three tests in
Fig. 2B–D). HCT-116 cells displayed
high adhesion with FITC-HM-3 and this correlated to its high
expression of integrins αvβ3 (Fig.
1) as integrin αvβ3 is a target for HM-3. Similar studies were
performed for Hep G-2 and SMMC-7721. SMMC-7721 appeared to possess
very low adherence with FITC-HM-3 (10.3%) which was 21 times
compared to the control (0.5%), whereas Hep G-2 displayed a 1.2%
binding with FITC-HM-3 that was approximately four times the
control cells (0.3%) (Fig. 2E). As
these two cell lines expressed high levels of integrin α5β1, this
result confirmed that HM-3 bound with a high affinity to integrin
αvβ3, but not to α5β1 (25). The
present study demonstrated that the three tumor cells varied in
their expression levels of integrin subunits and in their capacity
to adhere with FITC-HM-3.
HM-3 inhibits the migration of HCT-116
and Hep G-2 cells
Cell proliferation and migration are both important
procedures during angiogenesis. A previous study found that HM-3
did not have inhibitory effect on the proliferation of HUVECs,
HCT-116 and Hep G-2 cells (data not shown). The inhibitory effect
of HM-3 in the migration of HCT-116 and Hep G-2 cells was evaluated
in the present study. The Transwell assay is commonly used to
evaluate the migratory response of endothelial cells or cancer
cells to angiogenic inducers or inhibitors (26). The regulatory effect of HM-3 at low
or high doses on the migration of two different tumor cell lines
(HCT-116 and Hep G-2) was investigated. Cells were chosen based on
their high and low affinity with integrins in the flow cytometric
assays (Fig. 2E). The positive
control sunitinib was chosen based on its broad spectrum effect in
tumor growth, angiogenesis and metastasis and its inhibition of
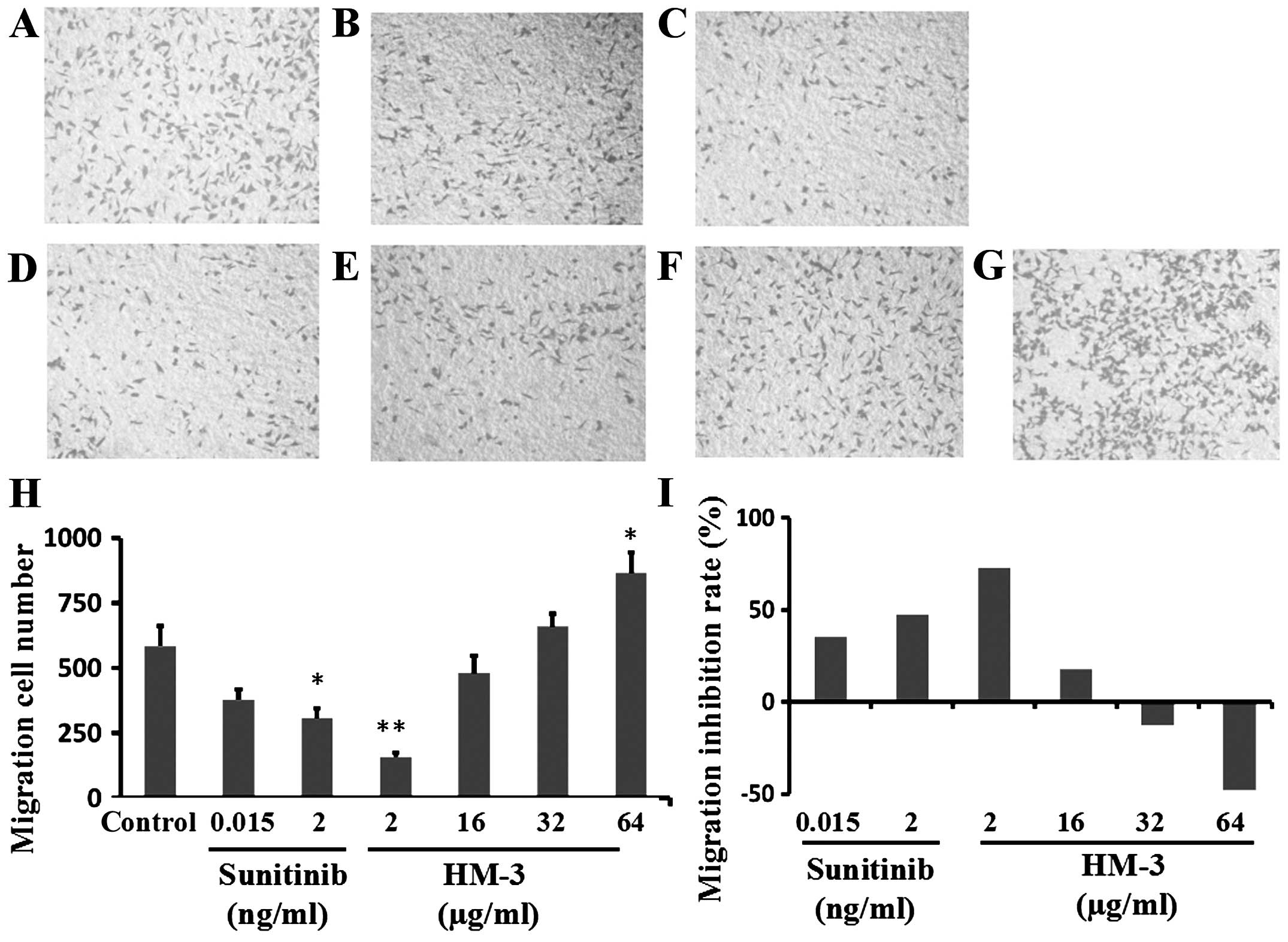
multiple receptor tyrosine kinases (RTKs) (27). Typical images of migrated HCT-116
cells under different conditions are shown in Fig. 3A–G. HM-3 at a concentration of 2
μg/ml displayed high inhibition of HCT-116 migration with a
cell migration number of 159 cells, while the cell migration number
in the negative control group was 585 cells (Fig. 3H). The inhibition rates of 2
μg/ml HM-3 in the migration of HCT-116 cells was 72.8%
(Fig. 3I). The inhibition rate
(MIR) of sunitinib at 0.015 and 2 ng/ml were 35.2 and 47.4%,
respectively (Fig. 3I).
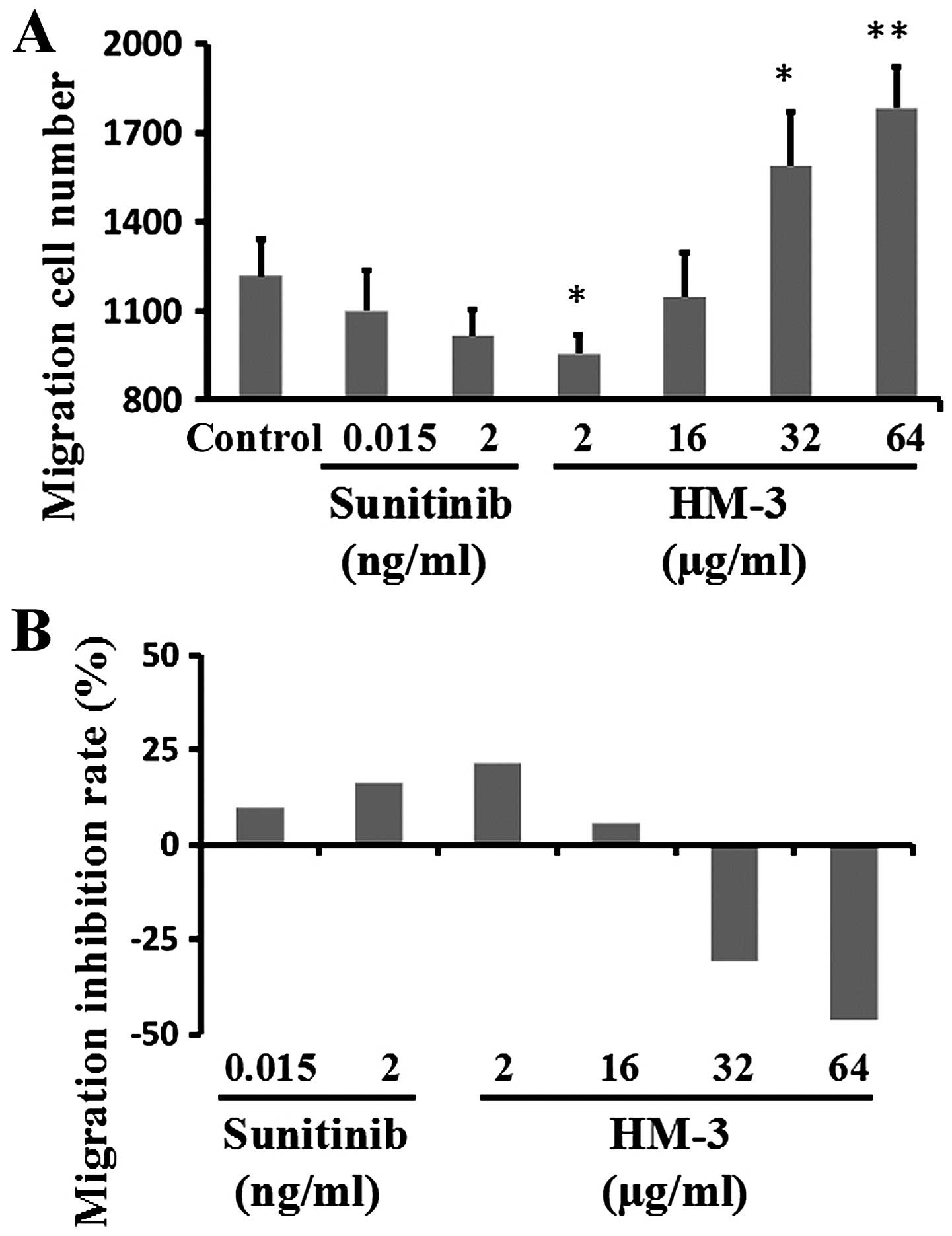
Furthermore, HM-3 showed lower inhibitory effects in Hep G-2
migration compared with HCT-116 migration at the corresponding
concentrations, e.g. the inhibition rate by HM-3 at 2 μg/ml
and sunitinib at 2 ng/ml were 21.6 and 7.9%, respectively (Fig. 4). However, both HM-3 and sunitinib
showed a similar dose-efficacy relationship in inhibition of
HCT-116 and Hep G-2 migration (Figs.
3I and 4B). These cell
migration results were in line with the above presented western
blot results (Fig. 1) and flow
cytometry results (Fig. 2E). The
inhibition of cell migration by HM-3 was also performed for HUVECs
(16) and HM-3 showed a similar
dose-efficacy relationship. At 8 μg/ml, HM-3 inhibited HUVEC
migration with an inhibition rate of 67.0% whereas at 32
μg/ml HM-3 promoted HUVEC migration with a promotion rate of
10.0%.
As HCT-116 cells showed high integrin expression,
high FITC-HM-3 binding and highly inhibitory effect in cell
migration by HM-3, this cell line was selected for further in
vivo study.
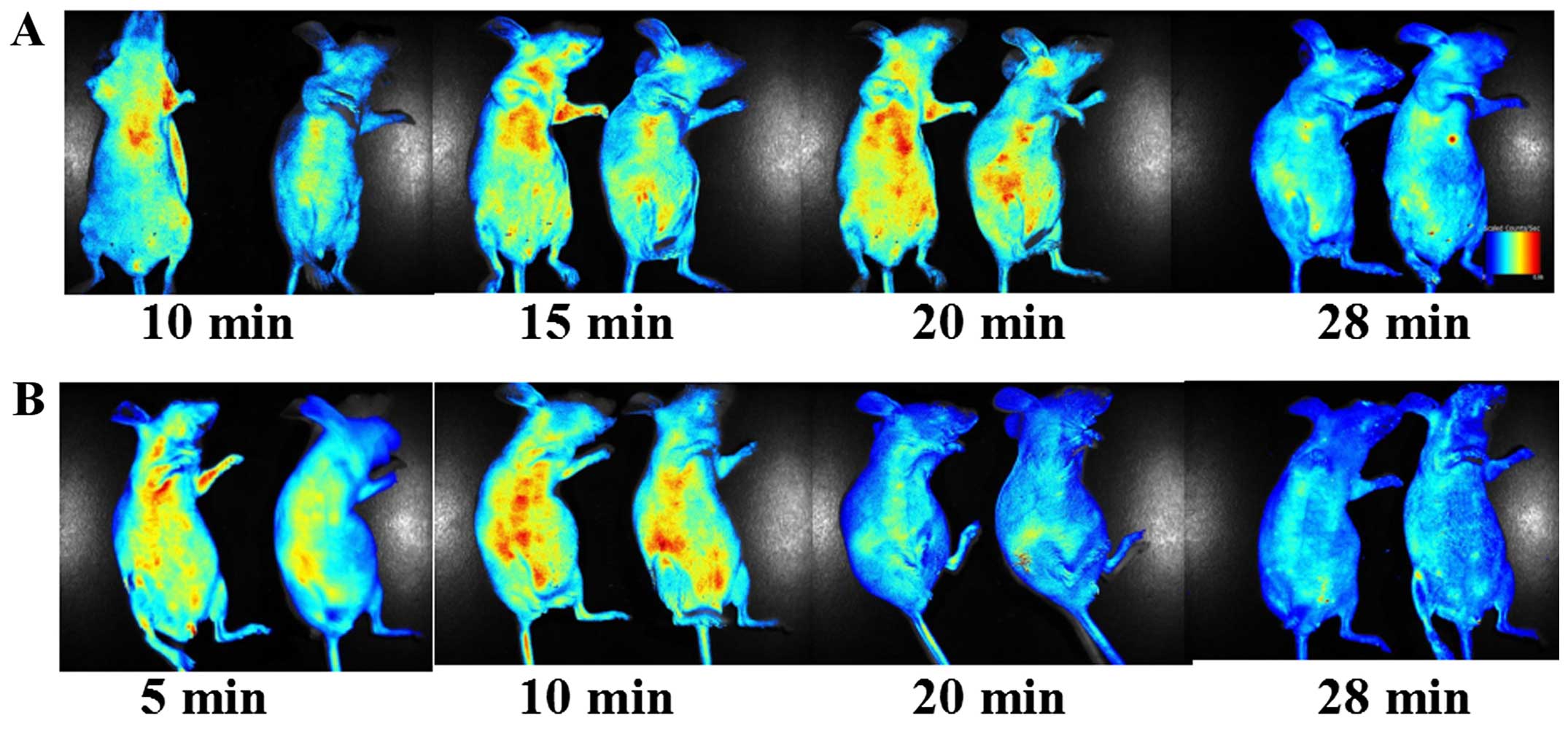
In vivo imaging
Being a peptide, HM-3 is prone to be degraded by
in vivo proteases (14,28).
The in vivo half-life of HM-3 in rat is only 27 min
(28). To assess the kinetics of
HM-3 in vivo and also the distribution of HM-3 in HCT-116
tumor-bearing and tumor-free mice, FITC-HM-3 was intravenously
injected and the fluorescence signal was detected at various time
points. Within 10 min of the injections, FITC-HM-3 appeared to be
widely distributed in different tissues, including the GIT, breast,
limbs, lung and also in the tumor mass (Fig. 5A and B). The signal for FITC-HM-3 in
tumor-free mice decreased quickly and at the time point of 20 min,
the signal substantially decreased (Fig. 5B). However, at 20 min, the signals
in HCT-116 tumor-bearing mice were still high (Fig. 5A). At 28 min, there was still
obvious signal for FITC-HM-3 and the signal existed in the position
of tumor mass (Fig. 5A). These
results confirmed that HM-3 had a short in vivo half-life
and also it targeted tumor tissue in vivo.
In vivo dose-efficacy relationship of
HM-3
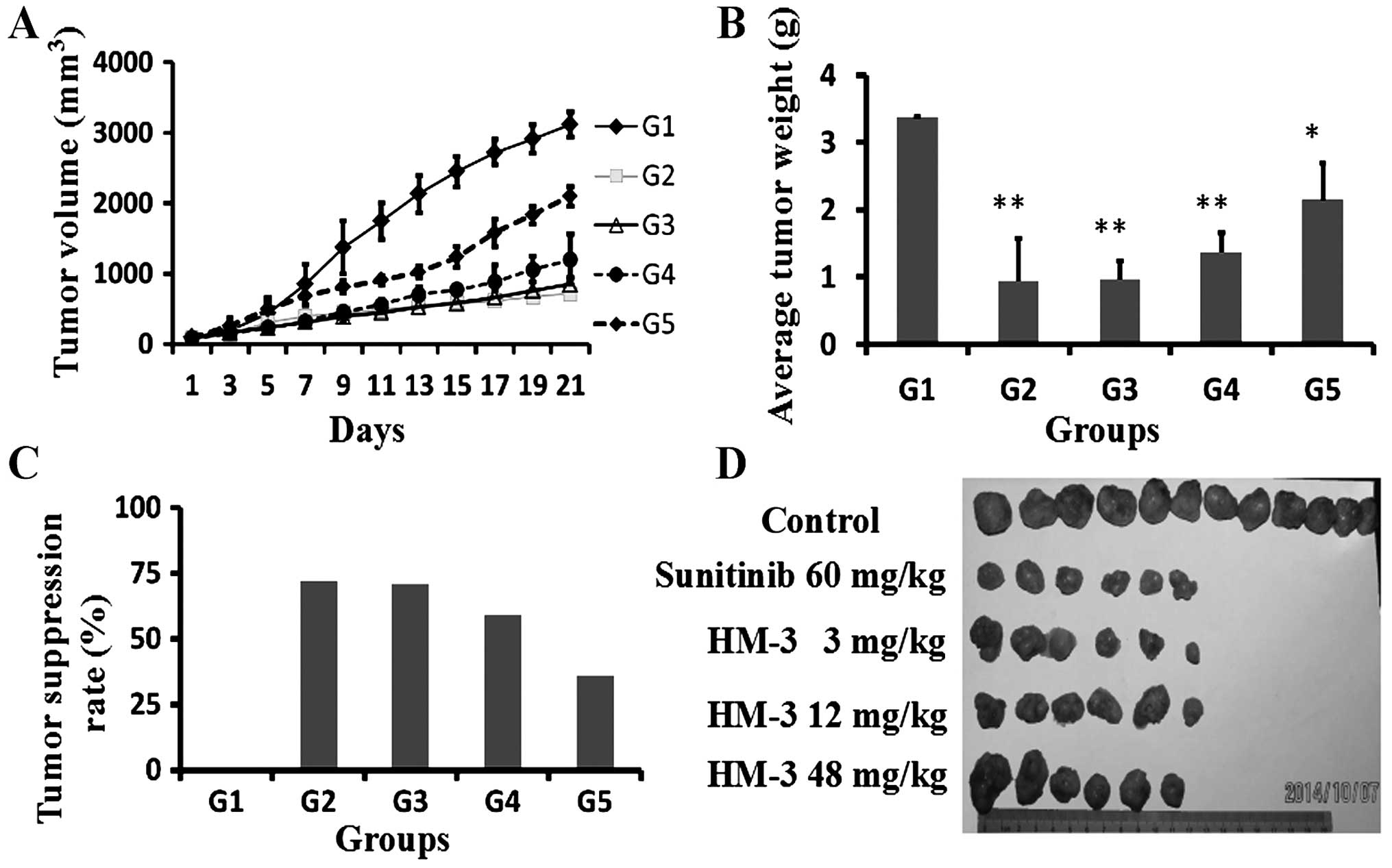
In vivo dose-efficacy relationship of HM-3
was evaluated on the HCT-116 xenograft model in nude mice. The
grouping and drug treatment strategies are shown in Table I. During the drug treatment period,
tumor volumes (Fig. 6) and mouse
weight in different groups (data not shown) were measured every two
days. At the initiation of therapy the tumor volume ranged between
75 and 100 mm3. The positive control sunitinib (60
mg/kg) was daily administered by intra oral gavage for 17 days.
Sunitinib at this dose inhibited tumor growth (Fig. 6A) with an inhibition rate of 72.0%
on day 21 (Fig. 6B and C). HM-3 at
3 mg/kg also showed inhibition with a tumor volume inhibition rate
of 74.0% on day 21 (Fig. 6A) and a
tumor mass inhibition rate of 71.5% (Fig. 6B and C). However, HM-3 did not show
a dose-dependent inhibition of tumor growth. At a dose of 12 mg/kg,
HM-3 showed a tumor volume inhibition of 61.9% and a tumor mass
inhibition of 59.2% (Fig. 6). At 48
mg/kg, HM-3 showed a tumor volume inhibition of 17.9% and tumor
mass inhibition of 36.0% (Fig. 6).
Data in the present study are in accordance with the previous
study. In a previous study, on an SMMC-7721 xenograft model in nude
mice, HM-3 at a dose of 1.5 or 3 mg/kg showed a high tumor growth
inhibition with inhibition rates of 57.4 and 53.0%. However, with
increase of HM-3 dosage, the inhibition rate decreased gradually
and at 24 mg/kg, the inhibition rate was only 9.5% (16).
HM-3 was not a cytotoxic reagent and during the
animal experiment, even the high dose of HM-3 had no sign of
toxicity as 100% of the mice survived in the tested groups until
sacrificed for tumor extraction. The present study agreed with a
previous (20) declaring that HM-3
had no apparent toxic effect on the animals.
Immunohistochemstry analysis of
expression of angiogenesis factors
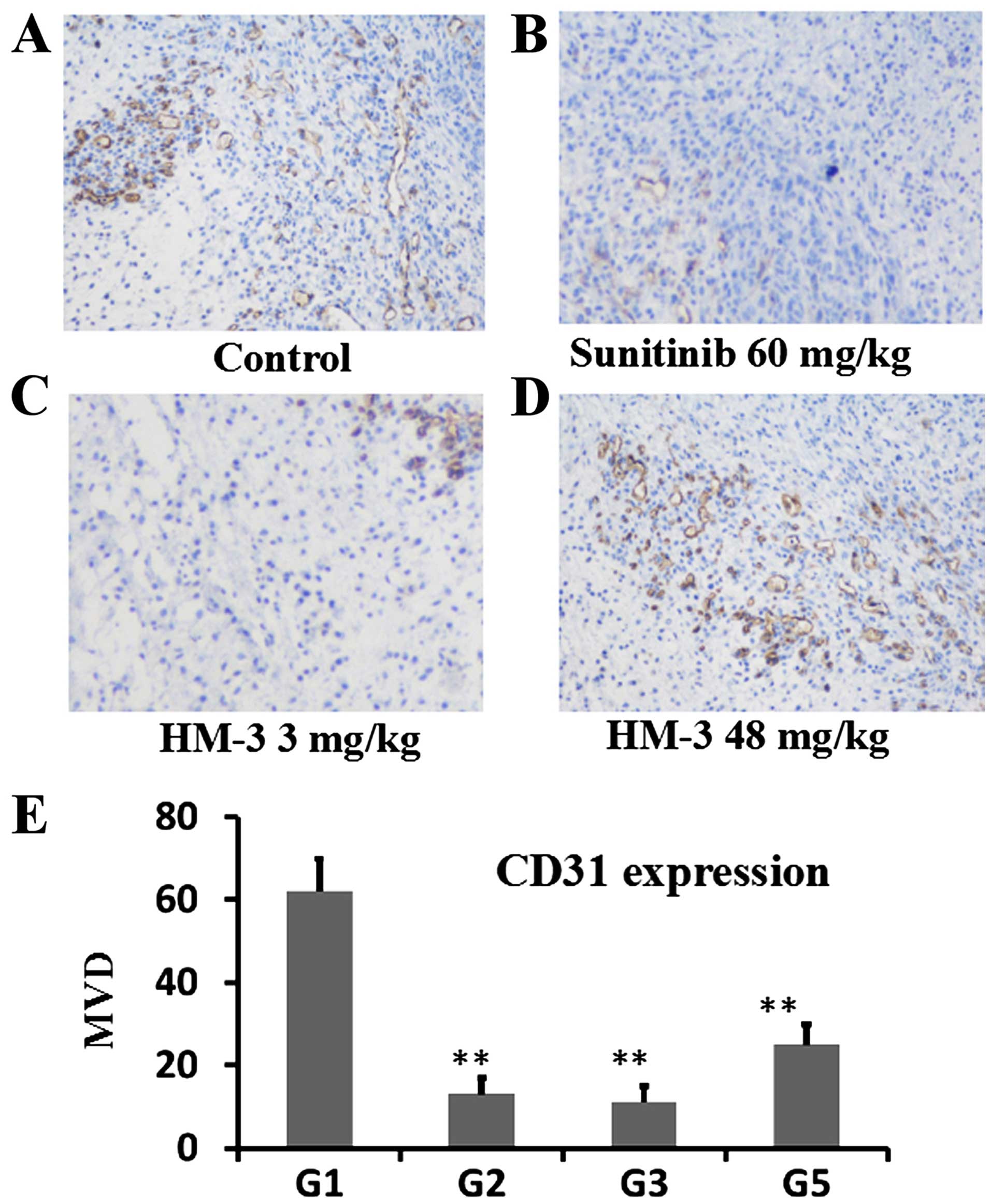
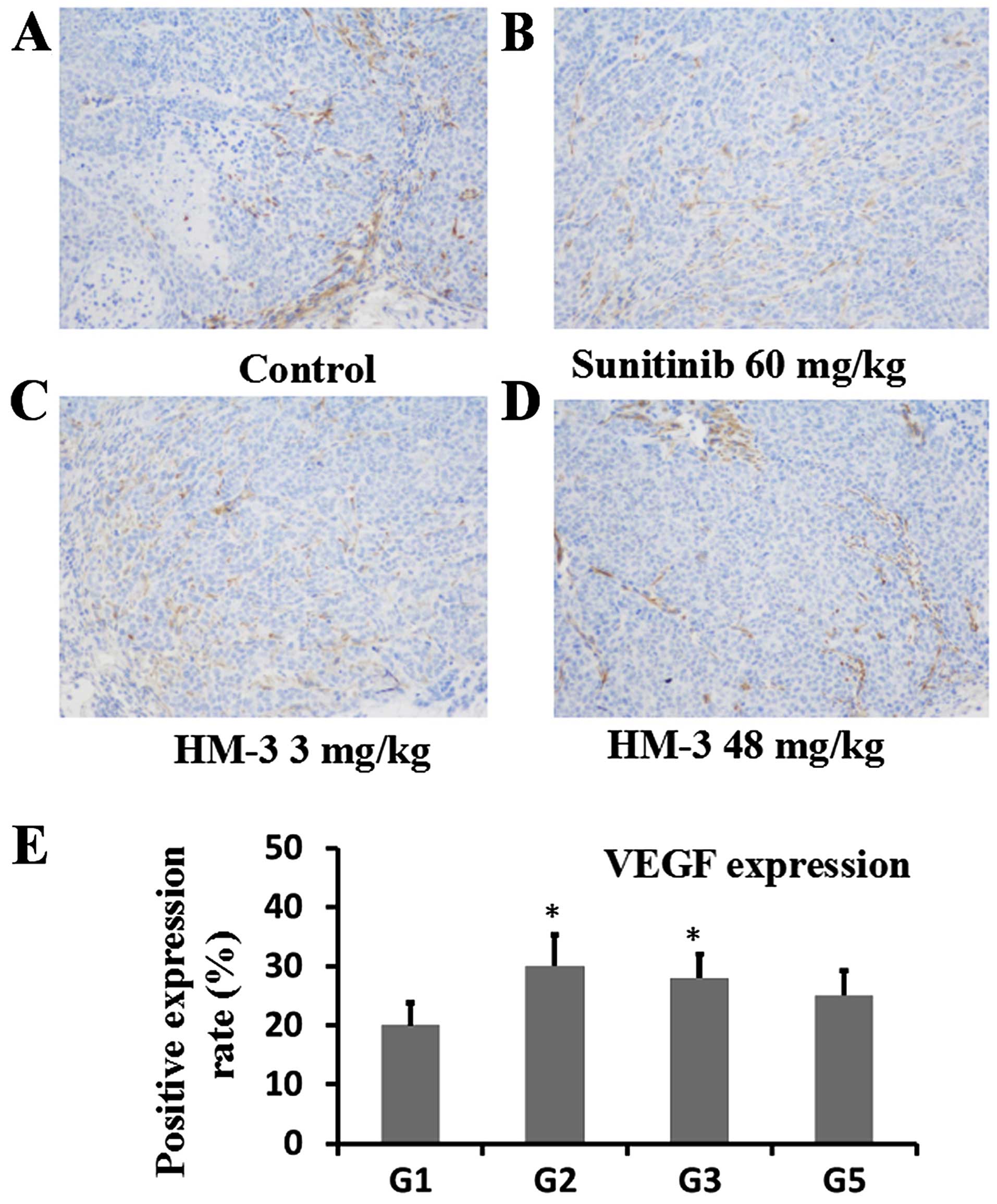
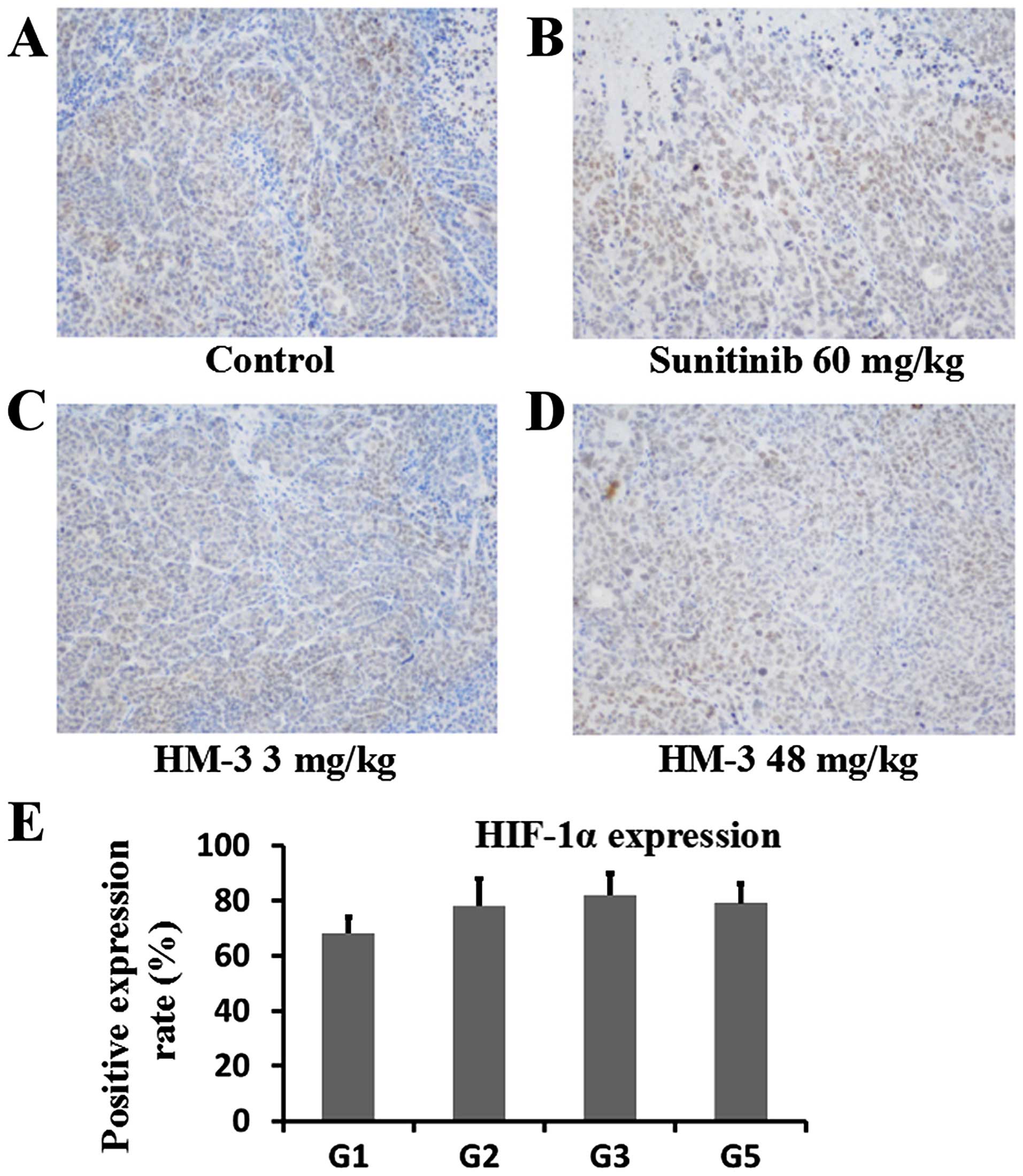
IHC analysis of CD31 (Fig. 7), VEGF (Fig. 8) and HIF-1α (Fig. 9) was performed to evaluate the
expression levels of these angiogenesis-related factors in HCT-116
tumors. Typical images for each experimental condition are
presented as panels A–D in Figs.
7Figure 8–9. In Fig.
7, microvascular density (MVD) of the tumor tissue from the
control group was 62.2 in average whereas those for sunitinib (60
mg/kg) group and HM-3 (3 mg/kg) group were 12.8 and 11.0,
demonstrating that these two anti-angiogenic reagents, though with
different working mechanisms, can substantially inhibit tumor
angiogenesis during tumor growth (Fig.
7E). The MVD of the tumor tissue from HM-3 (48 mg/kg) group was
25.8, showing a decreased anti-angiogenic effect of HM-3 at this
dose (Fig. 7E). This is also in
line with the tumor growth inhibition experiments (Fig. 6). In Fig. 8, percent of cells expressing VEGF
were counted after staining of VEGF-expressing cells
immunohistochemically. On average 20.3% of total cells were
positively stained on the sections from control group tumors. On
the sections from sunitinib (60 mg/kg) and HM-3 (3 mg/kg) groups,
the percent of positively stained cells were 29.8 and 28.2.
Similarly, 25.1% of cells were positively stained on the sections
from HM-3 (48 mg/kg) group (Fig.
8E). This indicates that the anti-angiogenic effect of HM-3 was
not due to the deceased levels of growth factors such as VEGF, but
to a direct inhibitory effect of HM-3 on HUVEC migration, which is
an important procedure during angiogenesis. Similarly, sunitinib
inhibited angiogenesis since it can 'cut-off' the signal
transduction pathway of VEGFR2, which is the main VEGF receptor
during angiogenesis, and so higher VEGF levels in the sunitinib (60
mg/kg) group did not increase angiogenesis. In Fig. 9, percent of cells positively stained
with HIF-1α were counted after probing of sections from different
groups with anti-HIF-1α antibody and staining. On the sections from
control group tumors, an average of 67.9% cells were positively
stained whereas for the sections from sunitinib (60 mg/kg) group,
HM-3 (3 mg/kg) and HM-3 (48 mg/kg) groups, the percent of
positively stained cells were 78.5, 82.3 and 79.1 (Fig. 9E). This result indicated that with
the decrease of blood vessel density compared with the control
group, the tumors of the drug treatment groups all had a higher
degree of hypoxia. The higher hypoxia state made the cells in tumor
mass express higher levels of HIF-1α and thereafter higher levels
of VEGF.
Discussion
In the present study, it was found that, of the
three human tumor cell lines HCT-116, Hep G-2 and SMMC-7721, only
HCT-116 expressed a high level of integrin αvβ3. In addition, this
cell bound with FITC-HM-3 at a high level in a flow cytometric
assay. Furthermore, HM-3 efficiently inhibited HCT-116 cell
migration in a Transwell assay. In addition, this inhibitory effect
was stronger than its inhibition of Hep G-2 migration under the
same concentrations. As HM-3 did not inhibit proliferation of
cancer cells (data not shown), the cell migration assay is an
important parameter to evaluate the cellular function of HM-3.
Based on the above results, HCT-116 was selected to setup an in
vivo model to evaluate the activity and dose-efficacy
relationship of HM-3.
On an HCT-116 xenograft model in nude mice, HM-3
inhibited HCT-116 tumor growth. Three effects may account for this
inhibitory effect: inhibition of HCT-116 proliferation, affecting
the generation of growth factors, e.g. VEGF that stimulates tumor
angiogenesis, or a direct effect on vascular endothelial cells to
inhibit angiogenesis. As has been mentioned, HM-3 had no cytotoxic
effect and did not inhibit HCT-116 proliferation. In fact, acute
toxicity tests in mice proved that the maximum tolerated dose of
HM-3 was 1,920 mg/kg by intravenous injection, which was >600
times as high as the effective dose (3 mg/mg) (14). Furthermore, HM-3 seems not to
decrease the expression levels of growth factors within tumor
tissues (Fig. 8). Actually,
sunitinib (60 mg/kg) and HM-3 (3 mg/kg) significantly increased
VEGF-expressing cells within tumor tissues compared with the
control group. This result is in accordance with a previous study
(29) that decreased MVD within the
tumor tissue (Fig. 7) caused higher
levels of hypoxia in the tumor microenvironment, which resulted in
upregulation of HIF-1α expressions in tumor cells and thereafter
higher levels of VEGF expression. However, this higher level of
VEGF did not increase angiogenesis in the tumor (Fig. 7), as HM-3 directly inhibits HUVEC
migration (15). Endothelial cell
migration is an important part of tumor angiogenesis and HM-3 can
inhibit this process. This result confirmed that cell proliferation
and cell migration are two independent processes, and inhibition of
endothelial cell migration is enough to efficiently inhibit
angiogenesis. In contrast, once angiogenesis inhibitors are
removed, the higher hypoxia state and higher levels of HIF-1α and
VEGF levels will restart angiogenesis in tumor tissues and tumor
will soon grow again. This was often found in clinical practice
(30).
After VEGF engagement of VEGFR2, the intracellular
parts of VEGFR2 were tyrosine-phosphorylated that can recruit and
activate intracellular guenine-nucleotide exchange factors (GEFs)
and GTPase activating proteins (GAPs). They can also activate GEFs
and GAPs indirectly via PI3Ks. Similarly, after integrin αvβ3
activation, the intracellular parts of integrin αvβ3 recruit and
activate FAK-Src complex, which also recruit and activate GEFs and
GAPs via 'adaptor proteins'. These GEFs and GAPs regulate the
activities of RhoGTPases, which include RhoA, Rac1 and Cdc42 and
are central regulators of cell migration (31). Recent research confirmed that
angiogenic regulations by integrin αvβ3 and VEGFR2 are not two
independent events, on the contrary, they synergize with each
other. After VEGFR2 is intracellular tyrosine-phosphorylated, they
recruit and activate SFKs to activate (phosphorylated) the integrin
β3 subunit. β3 phosphorylation triggers the generation of a complex
of integrin αvβ3 and VEGFR2, the formation of which further
stimulats the phosphorylation of VEGFR2 (32–34).
Antibodies of integrin αvβ3 (35,36)
and receptor-tyrosine kinase inhibitors (e.g. sunitinib) (37) all inhibit the complex formation,
inhibit phosphorylation of intracellular parts of VEGFR2 and
integrin β3 subunit, decrease the activities of RhoGTPases and
inhibit cell migration and angiogenesis (38). This working model can be used to
further investigate the molecular mechanisms of the special
dose-efficacy relationship of integrin antagonists and other
anti-angiogenic reagents such as those targeting VEGF (e.g.
avastin) or its intracellular signaling (e.g. sunitinib). The above
molecular mechanistic aspects are worthy of further
investigation.
Acknowledgments
The present study was supported by the Priority
Academic Program Development of Jiangsu Higher Education
Institutions (PAPD), the Project Program of State Key Laboratory of
Natural Medicines (no. SKLNMBZ201403), and the National Science and
Technology Major Projects of New Drugs (nos. 2016ZX09101121 and
2014ZX09508007) in China.
References
|
1
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: The next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Folkman J: Seminars in Medicine of the
Beth Israel Hospital, Boston. Clinical applications of research on
angiogenesis. N Engl J Med. 333:1757–1763. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sharma RA, Harris AL, Dalgleish AG,
Steward WP and O'Byrne KJ: Angiogenesis as a biomarker and target
in cancer chemoprevention. Lancet Oncol. 2:726–732. 2001.
View Article : Google Scholar
|
|
4
|
Carmeliet P and Jain RK: Angiogenesis in
cancer and other diseases. Nature. 407:249–257. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Cai W and Chen X: Anti-angiogenic cancer
therapy based on integrin alphavbeta3 antagonism. Anticancer Agents
Med Chem. 6:407–428. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Demetri GD, van Oosterom AT, Garrett CR,
Blackstein ME, Shah MH, Verweij J, McArthur G, Judson IR, Heinrich
MC, Morgan JA, et al: Efficacy and safety of sunitinib in patients
with advanced gastrointestinal stromal tumour after failure of
imatinib: A randomised controlled trial. Lancet. 368:1329–1338.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Motzer RJ, Michaelson MD, Redman BG, Hudes
GR, Wilding G, Figlin RA, Ginsberg MS, Kim ST, Baum CM, DePrimo SE,
et al: Activity of SU11248, a multitargeted inhibitor of vascular
endothelial growth factor receptor and platelet-derived growth
factor receptor, in patients with metastatic renal cell carcinoma.
J Clin Oncol. 24:16–24. 2006. View Article : Google Scholar
|
|
8
|
Folkman J: Role of angiogenesis in tumor
growth and metastasis. Semin Oncol. 29(Suppl 16): S15–S18. 2002.
View Article : Google Scholar
|
|
9
|
Brooks PC, Clark RA and Cheresh DA:
Requirement of vascular integrin alpha v beta 3 for angiogenesis.
Science. 264:569–571. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jin H and Varner J: Integrins: Roles in
cancer development and as treatment targets. Br J Cancer.
90:561–565. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kumar CC: Integrin alpha v beta 3 as a
therapeutic target for blocking tumor-induced angiogenesis. Curr
Drug Targets. 4:123–131. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu Z, Wang F and Chen X: Integrin
alpha(v)beta(3)-targeted cancer therapy. Drug Dev Res. 69:329–339.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Meerovitch K, Bergeron F, Leblond L,
Grouix B, Poirier C, Bubenik M, Chan L, Gourdeau H, Bowlin T and
Attardo G: A novel RGD antagonist that targets both alphavbeta3 and
alpha-5beta1 induces apoptosis of angiogenic endothelial cells on
type I collagen. Vascul Pharmacol. 40:77–89. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhou K, Zheng X, Xu HM, Zhang J, Chen Y,
Xi T and Feng T: Studies of poly(ethylene glycol) modification of
HM-3 polypeptides. Bioconjug Chem. 20:932–936. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Xu HM, Yin R, Chen L, Siraj S, Huang X,
Wang M, Fang H and Wang Y: An RGD-modified endostatin-derived
synthetic peptide shows antitumor activity in vivo. Bioconjug Chem.
19:1980–1986. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xu H, Pan L, Ren Y, Yang Y, Huang X and
Liu Z: RGD-modified angiogenesis inhibitor HM-3 dose: Dual function
during cancer treatment. Bioconjug Chem. 22:1386–1393. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reddy AB, Srivastava SK and Ramana KV:
Aldose reductase inhibition prevents lipopolysaccharide-induced
glucose uptake and glucose transporter 3 expression in RAW264.7
macrophages. Int J Biochem Cell Biol. 42:1039–1045. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Liew K, Yong PV, Lim YM, Navaratnam V and
Ho AS: 2-Methoxy-1,4-Naphthoquinone (MNQ) suppresses the invasion
and migration of a human metastatic breast cancer cell line
(MDA-MB-231). Toxicol In Vitro. 28:335–339. 2014. View Article : Google Scholar
|
|
19
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhao C, Wang X, Zhao Y, Li Z, Lin S, Wei Y
and Yang H: A novel xenograft model in zebrafish for
high-resolution investigating dynamics of neovascularization in
tumors. PLoS One. 6:e217682011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Plate KH, Breier G, Millauer B, Ullrich A
and Risau W: Up-regulation of vascular endothelial growth factor
and its cognate receptors in a rat glioma model of tumor
angiogenesis. Cancer Res. 53:5822–5827. 1993.PubMed/NCBI
|
|
22
|
Janouskova H, Ray AM, Noulet F,
Lelong-Rebel I, Choulier L, Schaffner F, Lehmann M, Martin S,
Teisinger J and Dontenwill M: Activation of p53 pathway by
Nutlin-3a inhibits the expression of the therapeutic target α5
integrin in colon cancer cells. Cancer Lett. 336:307–318. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Cheong SJ, Lee CM, Kim EM, Uhm TB, Jeong
HJ, Kim DW, Lim ST and Sohn MH: Evaluation of the therapeutic
efficacy of a VEGFR2-blocking antibody using sodium-iodide
symporter molecular imaging in a tumor xenograft model. Nucl Med
Biol. 38:93–101. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Zhang W, Ran S, Sambade M, Huang X and
Thorpe PE: A monoclonal antibody that blocks VEGF binding to VEGFR2
(KDR/Flk-1) inhibits vascular expression of Flk-1 and tumor growth
in an orthotopic human breast cancer model. Angiogenesis. 5:35–44.
2002. View Article : Google Scholar
|
|
25
|
Taherian A, Li X, Liu Y and Haas TA:
Differences in integrin expression and signaling within human
breast cancer cells. BMC Cancer. 11:2932011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Senger DR, Perruzzi CA, Streit M,
Koteliansky VE, de Fougerolles AR and Detmar M: The alpha(1)beta(1)
and alpha(2)beta(1) integrins provide critical support for vascular
endothelial growth factor signaling, endothelial cell migration,
and tumor angiogenesis. Am J Pathol. 160:195–204. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Szałek E, Karbownik A, Sobańska K, Płotek
W, Grabowski T, Nowak M and Grześkowiak E: The penetration of
sunitinib through the blood-brain barrier after the administration
of ciprofloxacin. Acta Pol Pharm. 71:691–697. 2014.
|
|
28
|
Zhu B, Xu HM, Zhao L, Huang X and Zhang F:
Site-specific modification of anti-angiogenesis peptide HM-3 by
polyethylene glycol molecular weight of 20 kDa. J Biochem.
148:341–347. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pàez-Ribes M, Allen E, Hudock J, Takeda T,
Okuyama H, Viñals F, Inoue M, Bergers G, Hanahan D and Casanovas O:
Antiangiogenic therapy elicits malignant progression of tumors to
increased local invasion and distant metastasis. Cancer Cell.
15:220–231. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Loges S, Mazzone M, Hohensinner P and
Carmeliet P: Silencing or fueling metastasis with VEGF inhibitors:
Antiangiogenesis revisited. Cancer Cell. 15:167–170. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ridley AJ, Schwartz MA, Burridge K, Firtel
RA, Ginsberg MH, Borisy G, Parsons JT and Horwitz AR: Cell
migration: Integrating signals from front to back. Science.
302:1704–1709. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Mahabeleshwar GH, Feng W, Reddy K, Plow EF
and Byzova TV: Mechanisms of integrin-vascular endothelial growth
factor receptor cross-activation in angiogenesis. Circ Res.
101:570–580. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Mahabeleshwar GH, Feng W, Phillips DR and
Byzova TV: Integrin signaling is critical for pathological
angiogenesis. J Exp Med. 203:2495–2507. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Mahabeleshwar GH, Chen J, Feng W, Somanath
PR, Razorenova OV and Byzova TV: Integrin affinity modulation in
angiogenesis. Cell Cycle. 7:335–347. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Byzova TV, Kim W, Midura RJ and Plow EF:
Activation of integrin alpha(V)beta(3) regulates cell adhesion and
migration to bone sialoprotein. Exp Cell Res. 254:299–308. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Byzova TV, Goldman CK, Pampori N, Thomas
KA, Bett A, Shattil SJ and Plow EF: A mechanism for modulation of
cellular responses to VEGF: Activation of the integrins. Mol Cell.
6:851–860. 2000.PubMed/NCBI
|
|
37
|
Garrett TA, Van Buul JD and Burridge K:
VEGF-induced Rac1 activation in endothelial cells is regulated by
the guanine nucleotide exchange factor Vav2. Exp Cell Res.
313:3285–3297. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Wu J, Strawn TL, Luo M, Wang L, Li R, Ren
M, Xia J, Zhang Z, Ma W, Luo T, et al: Plasminogen activator
inhibitor-1 inhibits angiogenic signaling by uncoupling vascular
endothelial growth factor receptor-2-αVβ3 integrin cross talk.
Arterioscler Thromb Vasc Biol. 35:111–120. 2015. View Article : Google Scholar
|