Introduction
Lung cancer has become the world's leading cause of
cancer-related deaths; the 5-year survival rate of patients with
small cell lung cancer (SCLC), in particular, remains less than
15%. High aggressiveness and a high metastatic potential are key
features of SCLC and are the main causes of treatment failure among
patients with SCLC (1,2). Although SCLC is sensitive to
chemotherapy and radiotherapy, most patients show only a partial
response (PR), and few achieve complete remission (CR). In
addition, the SCLC patients who do reach a PR or CR are known to be
prone to relapse and metastasis within a short period (3,4).
Prevention of the invasiveness and metastasis of SCLC has become a
bottleneck in the prognosis of SCLC patients and an urgent clinical
problem (5).
Proteins or lipids on the cell surface are often
linked to diverse sugar residues, thus, forming sugar chains with a
variety of special structures that play an important role in the
regulation of cellular functions; this process is known as
glycosylation (6). Sialic acid (SA)
is a nine-carbon monosaccharide that is added to the sugar chain
ends during protein or lipid glycosylation and is covalently linked
to the subterminal galactose (Gal) or N-acetylgalactosamine
(N-GalNAc) via α-2,3, α-2,6 or α-2,8 bonds. Polysialic acid (PSA)
is a long polymer of sialic acid monosaccharide residues connected
via α-2,8 bonds (7). In mammals,
the most important role of PSA is covalent modification of certain
cell surface proteins, among which the most common is neural cell
adhesion molecule (NCAM). PSA acts as a post-translational
modification for these proteins and regulates their functions; its
attachment is mainly catalyzed and completed by one of two
polysialyltransferases: ST8SiaII or ST8SiaIV (8). Various studies showed that PSA is
highly expressed during embryogenesis, but in healthy adults, its
expression is low except in nerve tissues. Recent studies showed
(9,10) that re-expression of PSA and its
transferases are detectable in certain tumors, such as melanoma,
breast cancer, malignant Wilms' tumor, glioma, lung and colorectal
cancer, medullary thyroid-like tumors and T-cell malignant
lymphoma, among others, and that the expression of PSA-NCAM is
closely related to the tumor progression and poor prognosis of the
patients, particularly in SCLC, pancreatic cancer and
neuroblastoma. However, the specific causes and mechanisms are not
yet clear.
The aims of the present study were to modify the
expression of polysialyltransferase within SCLC cells in culture to
change the modification by PSA of cellular NCAM and to analyze its
effects on the invasive and metastatic potential of SCLC cells (and
the possible mechanisms). The ultimate aim was to lay a foundation
for further exploration of the metastatic mechanism and development
of new targeted modalities for prevention of SCLC.
Materials and methods
Culture and transfection of SCLC
cells
Cells of the human SCLC cell line H446 were
purchased from the Shanghai Institutes for Biological Sciences
(Shanghai, China), then routinely cultured in RPMI-1640 medium
(HyClone Laboratories, Inc., Logan, UT, USA) containing 10% fetal
bovine serum (Gibco Life Technologies, Grand Island, NY, USA) at
37°C and 5% CO2.
H446 cells were transfected with the
ST8SiaII-encoding lentiviral expression vector and a
recombinant plasmid (Sigma-Aldrich, St. Louis, MO, USA) that can
intracellularly express short hairpin RNA (shRNA). The
transfection-positive cells were then screened and used to
establish the lung cancer cell line H446-ST8SiaII-LENTI that
overexpress the polysialyltransferase ST8SiaII (ST8SiaII was
previously abbreviated as STX; and ST8SiaIV was previously
abbreviated as PST; these terms may cause confusion) as well as the
H446-ST8SiaII-shRNA cell line that shows inhibited expression of
ST8SiaII. The plasmid transfection reagent Lipofectamine 2000 was
purchased from Invitrogen Life Technologies (Grand Island, NY,
USA).
Immunofluorescence and confocal
detection
Immunofluorescence analysis (IF) was performed to
detect the expression of PSA-NCAM and polysialyltransferase within
the SCLC H446 cells. The procedures were as follows: i) the cells
were seeded in pretreated 12-well plates for subculturing, and when
the cells attained ~70% confluency, the coverslip was removed and
the cells were washed with phosphate-buffered saline (PBS) three
times, for 5 min each time. ii) The cells were fixed in 4%
paraformaldehyde for 15 min, and then stored in PBS at 4°C. iii)
The cells were removed from the solution and washed with PBS three
times, 5 min each; before the assay for polysialyltransferase,
Tween-20 was added, and the cells were processed for 15 min, and
then washed with PBS three times, for 5 min each time. iv)
Non-specific binding sites on the cells were blocked with 10%
normal serum for 10 min. v) A mouse anti-human PSA-NCAM monoclonal
antibody (mAb; diluted 1:300; LifeSpan BioSciences, Inc., Seattle,
WA, USA) and rabbit anti-human anti-ST8SiaII mAb (diluted 1:300;
Sigma-Aldrich) were added for the immunoassay, and the mixtures
were incubated in a humidified chamber at 4°C overnight. vi) The
cells were removed from these solutions and washed with PBS three
times, 5 min each, after which the fluorescence-labeled
immunoglobulin G (IgG) [green fluorescence staining with
fluorescein isothiocyanate (FITC), Cy3 red fluorescence staining,
diluted 1:200] was added with subsequent incubation for 1 h in the
dark at room temperature, and 4′,6-diamidino-2-phenylindole (DAPI)
was used to stain the nuclei. Then, the cells were washed with PBS
three times, for 5 min each time, followed by one rinse with
distilled water. vii) The cells were mounted with 90% glycerol; an
anti-quenching mounting liquid was added dropwise, and the cells
were then examined and photographed under an ordinary fluorescence
microscope or a laser scanning confocal microscope [LSCM, Leica
TCS-SP5; Leica Application Suite Advanced Fluorescence (LAS AF)
software].
RT-PCR
RT-PCR was utilized to analyze the mRNA expression
of polysialyltransferases ST8SiaII and ST8SiaIV in the human SCLC
H446 cells for further experimental screening. In order to detect
the expression of ST8SiaII and ST8SiaIV in H446 cells, the
following primers were designed (5→3): ST8SiaII-F,
GGATCAGCAAGCAGGAGTATG and ST8SiaII-R, AGAAAGGGTGTAACGCAACTAA;
ST8SiaIV-F, GAGATGTGTCAGTGGTCAAGAG and ST8SiaIV-R,
GAAACTTCAGGTAGGAGGCTATG.
The procedures were as follows: i) according to the
GenBank sequences, IDTDNA Network Software (http://www.idtdna.com) was used to design the forward
and reverse RT-PCR primers for polysialyltransferase ST8SiaII,
ST8SiaIV, NCAM1 and NCAM2. ii) Total RNA was extracted according to
the kit instructions. iii) After treatment with DNase I, 2 µg of
RNA and avian myeloblastosis virus (AMV) reverse transcriptase were
added into the reaction mixture for reverse transcription. iv)
Forward and reverse primers for different genes were added, and the
PCR reaction was run in PCR buffer. v) The PCR reaction involved 30
cycles, with the annealing temperature set to 56, 59 or 63°C. vi)
The PCR products were subjected to gel electrophoresis in 1.5%
agarose and ethidium bromide staining, with
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as the internal
reference. We used Quantity One software (Bio-Rad Laboratories,
Inc., Hercules, CA, USA) for analysis of the gel images, and the
relative densitometric data from the strips under study and GAPDH
were then compared.
Western blotting
Western blot analysis was performed to analyze the
expression of polysialyltransferase proteins in each group after
the treatment. We analyzed the expression differences in such
signaling proteins as fibroblast growth factor receptor (FGFR),
phospholipase C γ1 (PLCγ1), and mitogen-activated protein kinase
(MAPK) [extracellular signal-related kinase 1/2 (ERK1/2)], and
their phosphorylation levels as well as the differences in
expression of blood metastasis-related genes, such as matrix
metalloproteinase-9 (MMP-9) and vascular endothelial growth
factor (VEGF). A mouse anti-human FGFR1 mAb, anti-FGFR1
(phospho-Y766) mAb, anti-PLCγ1 mAb, anti-MMP-9 mAb and an anti-VEGF
mAb were all purchased from Abcam Co. (Cambridge, UK); a rabbit
anti-human phospho-PLCγ1 (Ser1248) mAb, mouse anti-human p44/42
MAPK (Erk1/2) mAb, and a rabbit anti-phospho-p44/42 MAPK (Erk1/2)
mAb were acquired from the Cell Signaling Technology, Inc.
(Danvers, MA, USA). The following are the specific procedures. i)
The cells were subdivided into the following groups: a, the H446
group; b, the ‘ST8SiaII is overexpressed’, group
H446-ST8SiaII-LENTI; and c, the ‘ST8SiaII expression is inhibited’
group, H446-ST8SiaII-shRNA. ii) Total cellular protein of each
group was extracted according to the kit instructions, and the
Bradford method was used to test the protein concentrations. iii) A
25-µg protein sample from each group (sample concentration 2 µg/µl)
was then subjected to sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) on a 12% gel, at 100 V for 1.5 h. iv)
The proteins were then transferred to a polyvinylidene fluoride
(PVDF) membrane, at 300 mA for 1 h. v) The gel was discarded, and
the membrane was washed with distilled water, after which the
western blocking solution was added, and the membrane was slowly
rocked on a shaker for blocking for 60 min at room temperature.
Then, antibodies to ST8SiaII, FGFR, PLCγ1, MAPK (ERK 1/2), MMP-9,
VEGF or to the phosphorylated forms of the related proteins
(diluted 1:1,000) were added for overnight incubation at 4°C. vi)
The membrane was removed from the solution and washed with
Tris-buffered saline (TBS) three times for 5 min each time, after
which a rabbit anti-mouse IgG secondary antibody (diluted 1:2500;
Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd., Beijing,
China) was added and allowed to react at 37°C for 2 h; then, the
membrane was removed from the solution and washed with TBS once for
5 min. vii) Enhanced chemiluminescence (ECL) staining (Pierce
Protein Biology, Grand Island, NY, USA) was then applied to the
membrane, and X-ray film was used for autoradiographic imaging,
with GAPDH as the internal reference. The target bands were
analyzed using Quantity One software.
Vector construction and transfection
of polysialyltransferase constructs into cells
We constructed a lentiviral vector carrying
ST8SiaII. The target vector was
plv.ExBip/Neo-EF1a-ST8SIAII-eGFP, the control vector was
plv.ExBip/Neo-EF1-eGFP, and the auxiliary vectors were
plv/helper-SL3, plv/helper-SL4 and plv/helper-SL5. Meanwhile, RNA
interference technology was used to select a polysialyltransferase
ST8SiaII-shRNA plasmid for silencing of the expression of ST8SiaII
in the SCLC H446 cells.
Transwell chamber invasion assay
The following procedure was used for the Transwell
chamber invasion assay. i) As prepared according to the
conventional 24-well Transwell chamber method, the upper chamber
was lined with Matrigel and incubated at 37°C for 5 h; then
serum-free medium was used to gently wash the gel. ii) The
post-treatment lung cancer cells from each group were collected and
diluted with the RPMI-1640 serum-free medium to adjust the cellular
concentration to 5×105/ml, and 200 µl of the cell
suspension was added to the upper chamber, while 600 µl of the cell
culture medium (containing 5 µg/ml fibronectin) was added to the
lower chamber. The chambers were cultured in a humidified incubator
(37°C and 5% of CO2) for 24 h. iii) Non-invasive cells
were carefully wiped from the upper chamber. iv) The upper chamber
was removed and placed upside down, fixed with 5% glutaraldehyde,
and precooled at 4°C for 30 min. v) The cells were stained with an
eosin solution and photographed; five visual fields were subjected
to counting, and the mean value was used to quantify the
transmembrane invasion of the lung cancer cells; the relative
multiples were compared with the control H446 cell group.
The wound healing (cell metastasis)
assay
After 48–72 h of culture, the cells of each group
were harvested with trypsin-EDTA, the cell count was adjusted to
5×105/ml, and the cells were seeded in 12-well plates
and cultured overnight. After a monolayer formed, a sterile pipette
tip was used to make scratches, and the cells were immediately
photographed. The cells were then cultured for 24 or 48 h, and were
then photographed. The ImageJ software was used to quantitatively
analyze the pixels in the scratched area, and the relative speed of
cell metastasis was compared with that in the H446 cell group.
Statistical analysis
The data were analyzed using the SPSS 13.0
statistical package (SPSS for Windows; SPSS, Inc., Chicago, IL,
USA), and the experiments were performed at least three times. The
intergroup differences were analyzed with one-way analysis of
variance (ANOVA) or the χ2 test; differences with
P<0.05 were considered statistically significant (very
significant at P<0.01).
Results
Expression of polysialylated NCAM and
polysialyltransferases in SCLC cells
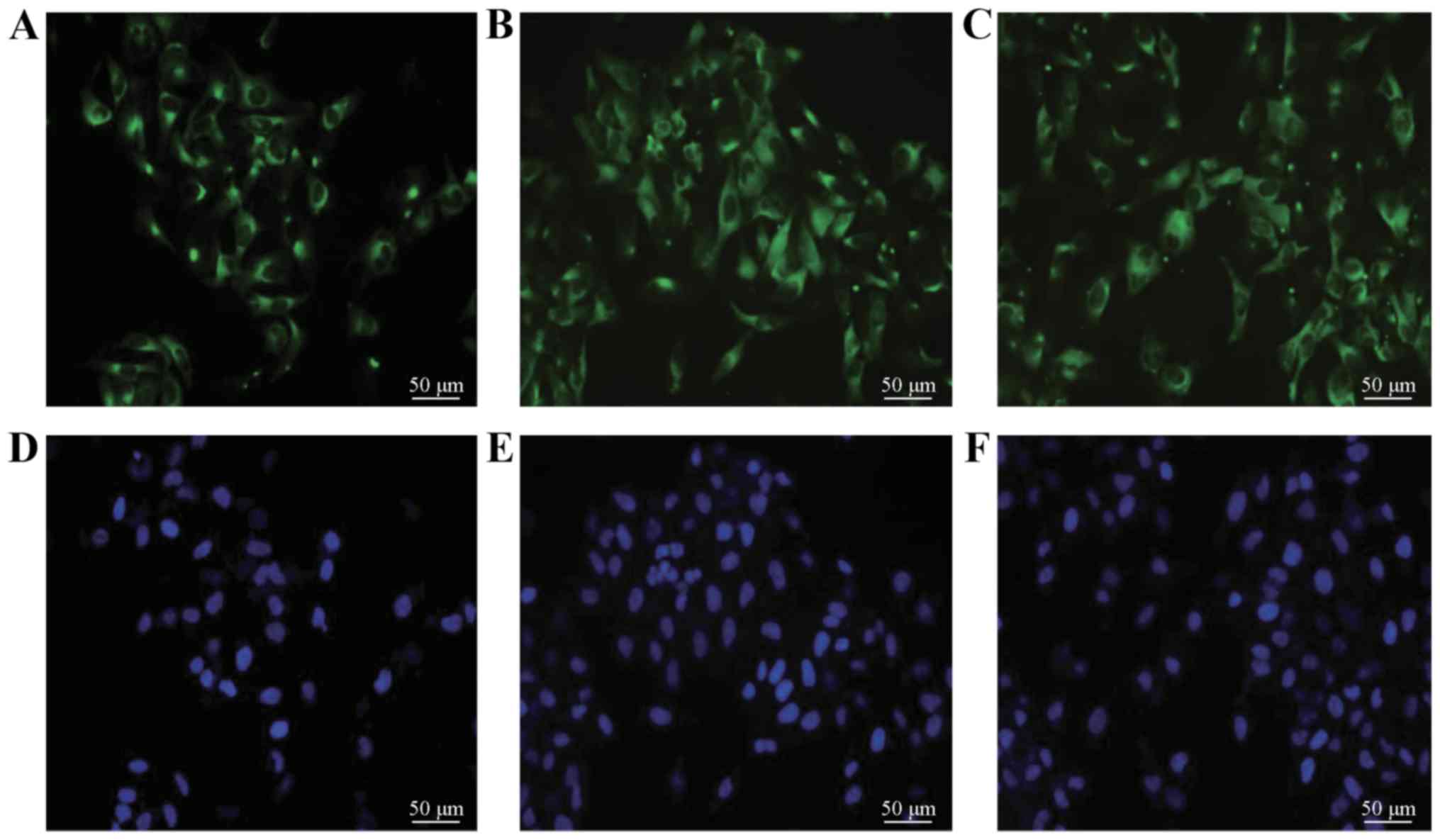
In the SCLC H446 cells, the results of IF revealed
obvious expression of PSA-NCAM, manifested as green fluorescence
(Fig. 1). The RT-PCR results at
different annealing temperatures showed that the expression of
ST8SiaII in every case was significantly stronger than that of
ST8SiaIV, and the difference was significant (P<0.05). The LSCM
results also confirmed significant expression of ST8SiaII in the
H446 cells (Fig. 2).
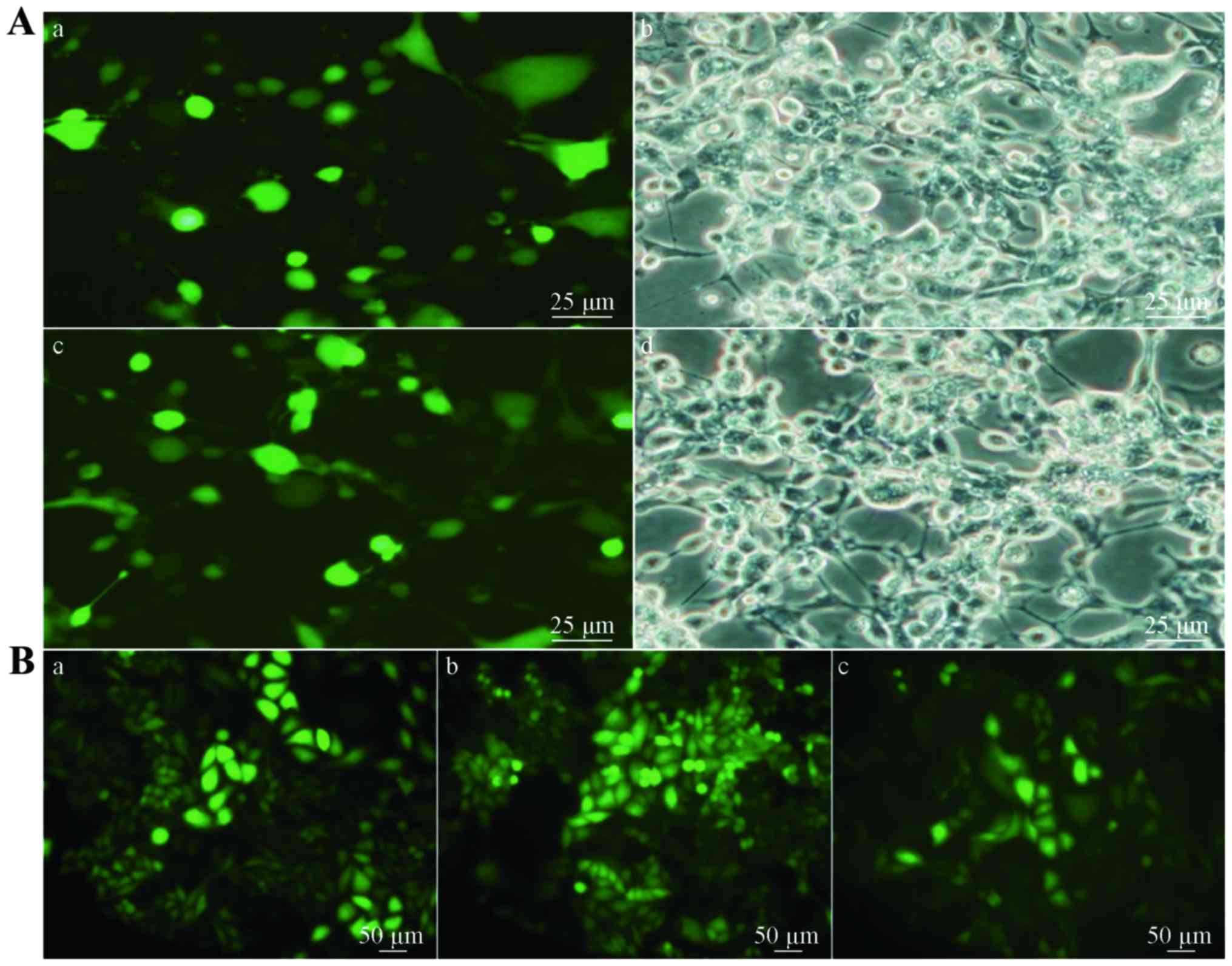
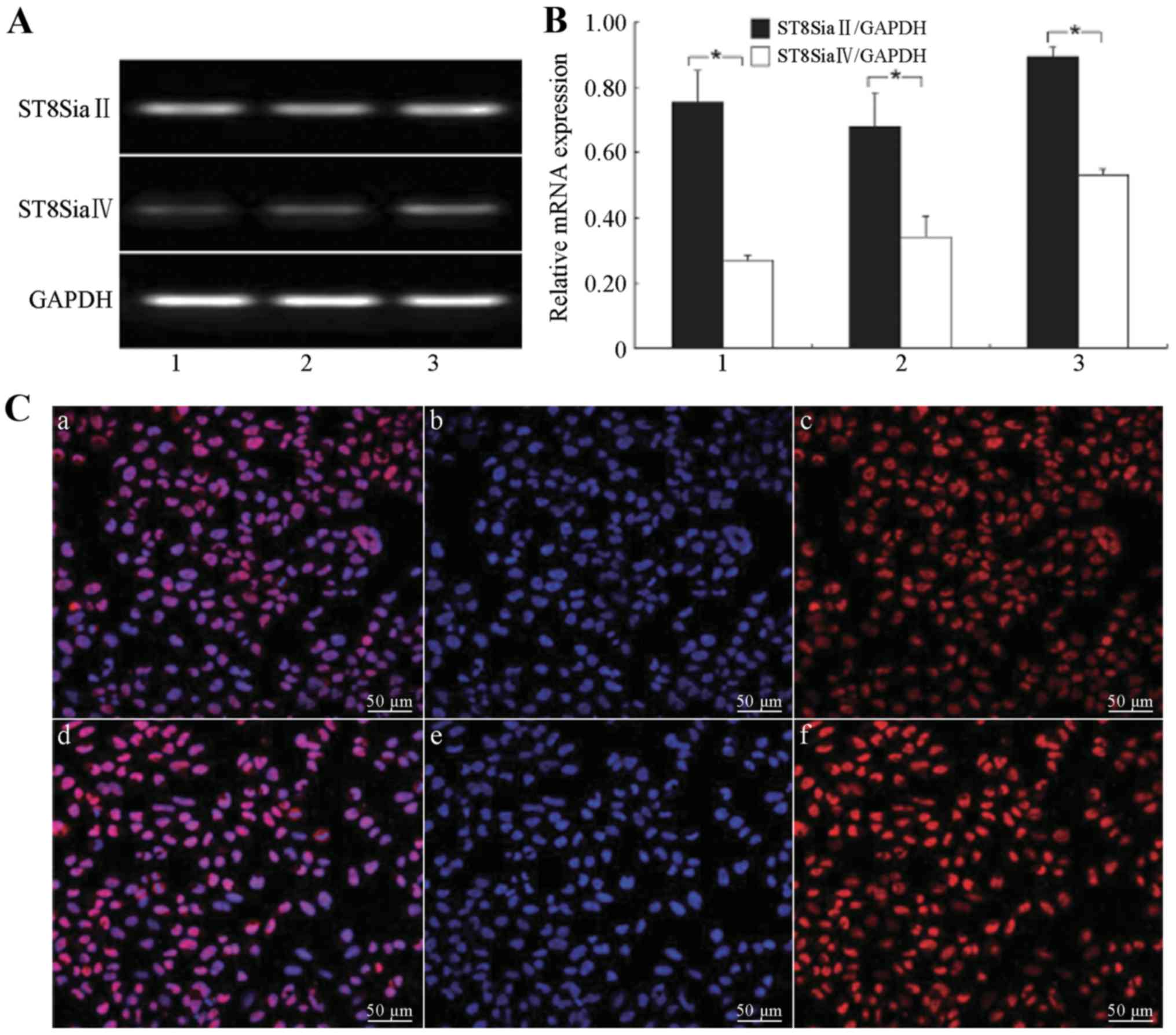 | Figure 2.Expression levels of
polysialyltransferase ST8SiaII and ST8SiaIV in the H446 cells. (A)
Expression levels of polysialyltransferase ST8SiaII and ST8SiaIV in
the H446 cells at different annealing temperatures: lane 1, 56°C;
lane 2, 59°C; and lane 3, 63°C. (B) Relative mRNA expression levels
of polysialyltransferase ST8SiaII and ST8SiaIV in the H446 cells
(*P<0.05): 1, 56°C; 2, 59°C; and 3, 63°C. (C) LSCM detection of
expression of polysialyltransferase ST8SiaII in the H446 cells: a
and d, overlay of the ST8SiaII-positive and nuclear staining
images; b and e, nuclear staining of H446 cells (DAPI, blue); c and
f, ST8SiaII-positive cells (Cy3 staining, red, exciting wavelength
561 nm). Scale bar, 50 µm. |
ST8SiaII expression in different
transfection groups
On the basis of the above results, we constructed
the lentiviral vector carrying ST8SiaII. After construction of
other recombinant vectors, the Lipofectamine 2000-cotransfected
lentivirus was used to package the cells, and 48 h later,
significant green fluorescence appeared, indicating that the
transfection efficiency was good. The packaged viral particles and
the prepared shRNA plasmid were then transfected into the target
H446 cells. After repeated screening, a cell line that could stably
and strongly express ST8SiaII was then established
(H446-ST8SiaII-LENTI), as was the cell line (H446-ST8SiaII-shRNA)
that showed stable suppression of ST8SiaII expression (Figs. 3 and 4).
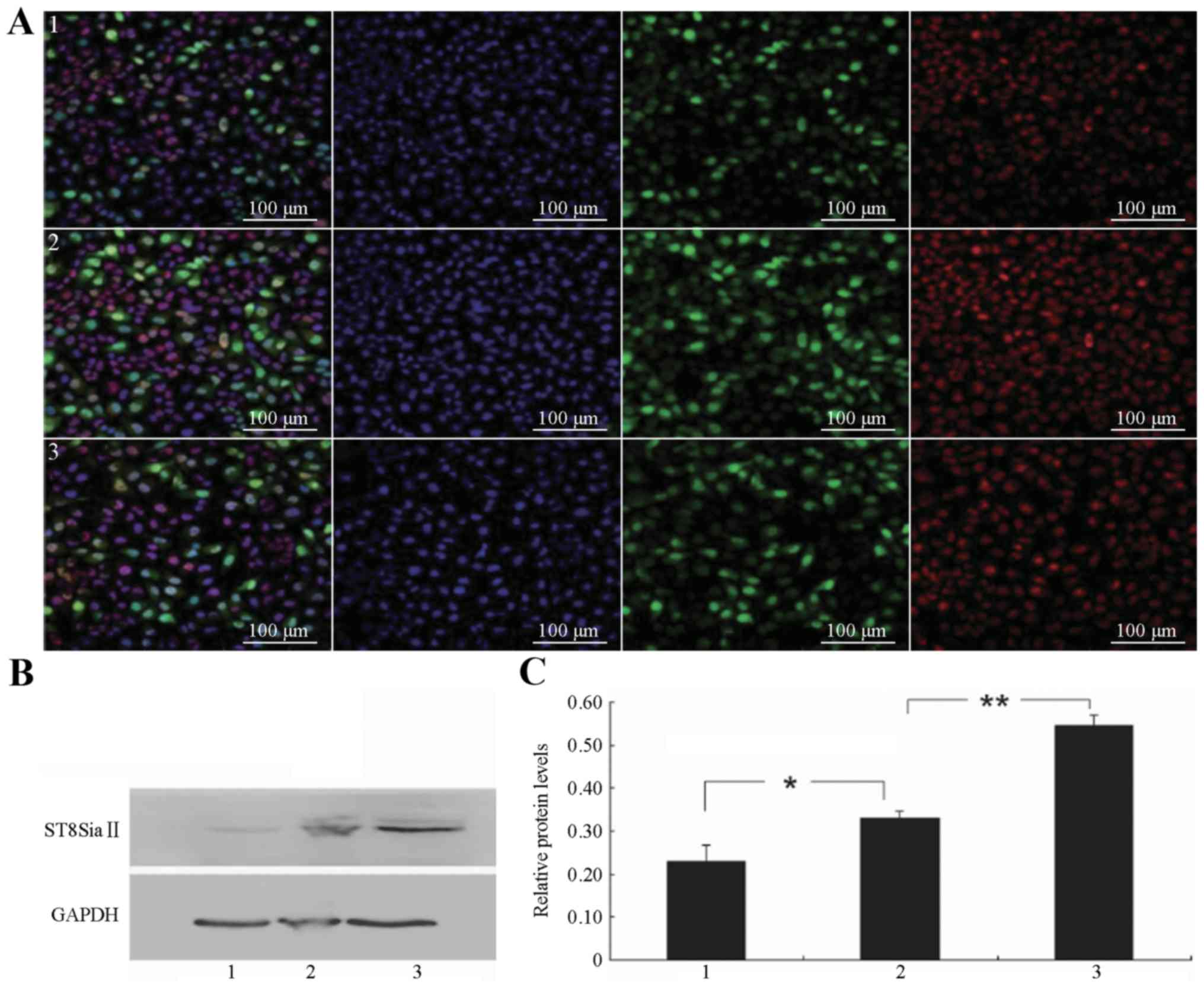 | Figure 4.Expression of ST8SiaII in the
different SCLC groups. (A) LSCM detection of ST8SiaII expression in
the different transfection groups; 1–3 (left) overlay of the
ST8SiaII-positive and nuclear staining images; (center left)
nuclear staining of SCLC cells (DAPI, blue); (center right) the
ST8SiaII-positive cells in the different groups (green, FITC
staining, exciting wavelength 488 nm); (right) the
ST8SiaII-positive cells in the different groups (red, Cy3 staining,
exciting wavelength 561 nm). Scale bar, 100 µm. (B) Western blot
detection of ST8SiaII protein expression in the different
transfection groups. (C) Relative levels of ST8SiaII protein
expression in the different transfection groups: 1,
H446-ST8SiaII-shRNA; 2, H446 blank control; and 3,
H446-ST8SiaII-LENTI; *P<0.05, **P<0.01. |
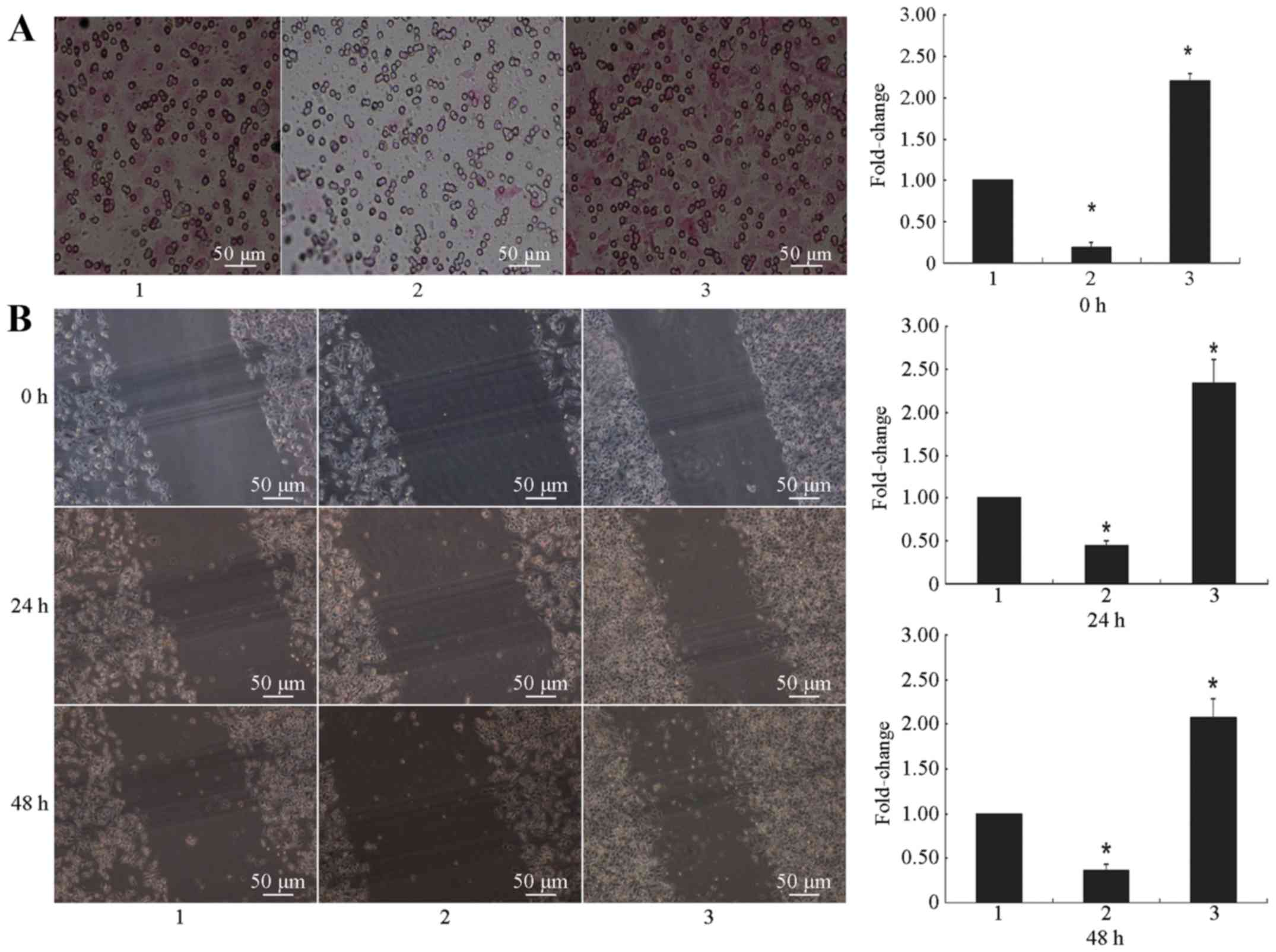
Invasion and metastasis
The in vitro invasion and metastasis
abilities of the three groups were tested. The Transwell assay
showed that the transmembrane invasion ability of the
H446-ST8SiaII-LENTI cell group was significantly increased as
compared to the control group (P<0.01), while the in
vitro transmembrane invasion ability of the H446-ST8SiaII-shRNA
group was significantly inhibited compared to the control group
(P<0.01; Fig. 5A). These results
suggested that alterations of the expression of
polysialyltransferase ST8SiaII can affect the post-transcriptional
PSA modification of NCAM, thus having significant effects on the
in vitro invasion ability of SCLC cells.
The in vitro metastasis potential of the
three cellular groups was also examined to study the effects of
alterations of the post-transcriptional PSA modification on the
metastatic potential of SCLC. The wound healing assay revealed that
the metastasis of the H446-ST8SiaII-LENTI cell group was
significantly enhanced, while that of the H446-ST8SiaII-shRNA group
was significantly inhibited (P<0.01 for both; Fig. 5B). These results suggested that
alterations in the expression of polysialyltransferase ST8SiaII
affected its capacity for the post-transcriptional PSA modification
of NCAM, thus influencing the in vitro metastasis of
SCLC.
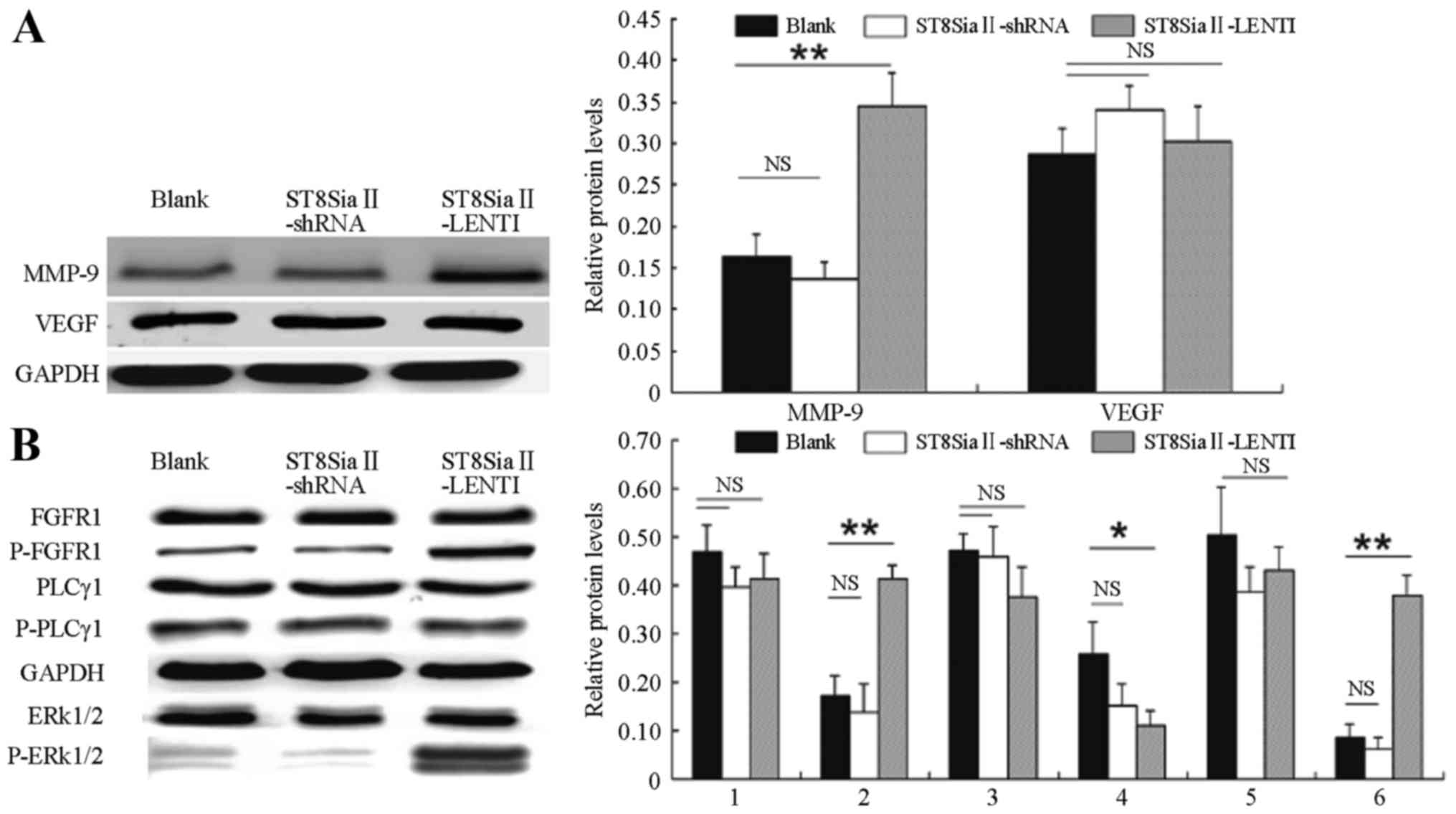
Changes in metastasis-related genes
and signaling molecules
Western blot analysis was performed to detect the
effects of the different treatments on SCLC metastasis-related
genes. We found (Fig. 6A) that when
the lentiviral transfection enhanced the expression of the
ST8SiaII gene in the SCLC cells and affected the
post-transcriptional PSA modification of NCAM
(H446-ST8SiaII-LENTI), the protein expression levels of an
invasion- and metastasis-related gene (MMP-9) was
significantly increased (P<0.01), while the expression of
VEGF showed no statistically significant change when
compared with the control group (P>0.05). In addition, in the
cells in which ST8SiaII was inhibited in a sustained fashion
(H446-ST8SiaII-shRNA), the protein expression of MMP-9 and VEGF
showed no significant differences with the control group
(P>0.05).
The protein expression and phosphorylation levels of
intracellular signaling molecules were then further tested after
treatment with various factors. The results (Fig. 6B) showed that in the
H446-ST8SiaII-LENTI group, the protein expression levels of FGFR1,
PLCγ1 and ERK1/2 did not significantly differ from levels in the
control group (P>0.05). In contrast, the expression levels of
phosphorylated FGFR1 and ERK1/2 were significantly increased
(P<0.01 for both). The expression level of phosphorylated PLCγ1
was decreased when compared with the level in the control group
(P<0.05). In the H446-ST8SiaII-shRNA group, the expression and
phosphorylation levels of the above proteins showed no significant
differences with the control group (P>0.05).
Discussion
Invasiveness and metastasis of SCLC seriously affect
patient prognosis, but since the specific mechanisms have not been
fully elucidated yet, the treatment effectiveness remains
unsatisfactory. Active research into (and improved understanding
of) the mechanisms of metastasis of SCLC and identification of new
therapeutic targets will have a positive influence on the
prevention and treatment of SCLC.
In the present study, we found that glycosylation of
proteins or lipids on the tumor cell surface had significant
effects on the metastatic potential of the tumor cells. The glycans
are a class of biological molecules within cells, in addition to
nucleic acids, proteins and lipids, and mainly include N-glycans
that are covalently linked to asparagine, O-glycans that are linked
to serine or threonine, GAGs that are linked to serine, and
glycolipids that are linked to lipid structures. Many of the body's
physiological and pathological processes including inflammation,
cancer, metabolic regulation, development, aging and immune defense
are closely related to cellular glycans (11). PSA is a long linear chain formed by
sialic acid monosaccharide residues (N-acetylneuraminic acid,
Neu5AC; or N-hydroxy-acetylneuraminic acid, Neu5GC) via α-2,8 bonds
(7). The PSA chain in mammalian
tissues generally consists of 50–150 residues, while the longest
chain detected is >370 sialic acid residues long. PSA first
caught the attention of researchers in the context of its roles in
post-transcriptional modification of NCAM (12).
NCAM is a transmembrane protein on the cell surface
and performs important functions in the development of nerve
tissues as well as adhesion and metastasis of neural cells in the
brain. The extramembranous structure of NCAM is composed of five Ig
homeodomains (IgI, IgII, IgIII, IgIV and IgV) and two type III
fibronectins (FN1 and FN2); the two adjacent domains are linked via
disulfide bonds (13). The
polysialylation of NCAM is mainly performed by ST8SiaII and
ST8SiaIV, which have their catalytic regions in the Golgi lumen,
and only ST8SiaII and ST8SiaIV can attach PSA to NCAM. The above
two enzymes can catalyze the reaction alone or have synergistic
effects, but the exact mechanisms have not been fully elucidated
(7–9). The binding sites for
polysialyltransferase on NCAM are mainly located in IgV and FN1
(14,15). NCAM2 is also called olfactory cell
adhesion molecule (OCAM) or Rb-8 neural cell adhesion molecule
(RNCAM). NCAM1 and NCAM2 share overall sequence identity of ~44%,
but their functions are not exactly the same, and NCAM-2 cannot be
covalently linked to PSA (16).
The present study showed for the first time that
when the SCLC H446 cells were transfected with a lentivirus and the
expression of polysialyltransferase ST8SiaII was enhanced (followed
by enhancement of the PSA modification of NCAM) the in vitro
invasion and metastasis abilities of the H446 cells were
significantly increase. Expression of the invasion- and
metastasis-related MMP-9 gene was also significantly
increased (P<0.01), while VEGF showed no significant change.
After further examining the changes in signaling molecules, we
found that when ST8SiaII expression was enhanced, the expression
levels of FGFR1, PLCγ1 and ERK1/2 were not significantly changes,
while the levels of phosphorylated FGFR1 and ERK1/2 were
significantly increased (P<0.01). When shRNA was used to knock
down the expression of polysialyltransferase ST8SiaII, the in
vitro invasion and metastasis abilities of the H446 cells were
also suppressed. In a similar fashion, Al-Saraire et al
found that when cytidine monophosphate was used to inhibit the
expression of ST8SiaII in neuroblastoma (SH-SY5Y) and glioma cells
(C6-ST8SiaII type) (this treatment secondarily inhibits the
ST8SiaII-mediated polysialylation of NCAM) the migration ability of
these two tumor cell types was significantly decreased (10). In contrast, the migration of
ST8SiaII itself [as well as the colon cancer cells (DLD-1) and
wild-type glioma cells (C6 wild-type) that showed no expression of
PSA] was not affected (10).
Recent studies (17–19)
showed that the post-transcriptional modification of NCAM by PSA
does not only affect its adhesion function but also has a ‘switch’
effect on the NCAM signal transduction functions. Before
modification by PSA, it is easy for NCAMs to form stable
homological adhesion. In contrast, when NCAM is highly modified by
PSA, the steric hindrance effects of the long-chain PSA (as well as
the mutual repulsion arising within negatively charged PSAs)
inhibit the homological adhesion among NCAMs. This change results
not only in separation of cancer cells but also in activation of
the NCAM-mediated intracellular signaling pathways.
Various studies have confirmed that activated NCAM
can activate intracellular signaling pathways mainly in two ways.
One is the receptor tyrosine kinase-dependent signaling pathway;
specifically, NCAM can act on FGFR or tyrosine kinase receptor B
(TrkB) on the adjacent cell membrane, thus activating the receptor
tyrosine kinase and promoting the downstream signaling cascades
(20–22). Li et al (23) reported that PSA-modified NCAM can
bind to the CAM homology domain (CHD) on the adjacent film FGFR,
thereby modulating the activities of FGFR and promoting the
metastasis of NIH-3T3 cells and African green monkey kidney cells
COS-7. This finding is related to the phosphorylation of PLCγ and
ERK1/2 in the FGFR downstream signaling pathways.
The other pathway that NCAM activates using
intracellular signals is the non-receptor-dependent pathway, which
is not mediated by FGF receptors (24–26).
It causes focal adhesion kinase (FAK) to bind to the NCAM-Fyn
complex directly via phosphorylation of intracellular tyrosine
protein kinase-related Fyn and FAK, then continues to activate the
downstream Ras/mitogen-activated protein kinase (MEK)/ERK or
phosphoinositide 3-kinase (PI-3K) signaling pathways (24–26).
The specific mechanisms of how the activated NCAM can activate the
intracellular signal transduction inside SCLC cells are still
unclear. Expression of NCAM is present in almost all tissues of
SCLC, and the diagnosis of SCLC can be confirmed by the results of
immunohistochemical analysis (27).
The present study showed that, in SCLC, the
post-transcriptional modification of NCAM with PSA glycans is
closely associated with the metastatic potential of SCLC cells. We
also found that activation of intracellular P-FGFR/P-ERK 1/2/MMP-9
signaling by PSA-NCAM may be a new mechanism that promotes the
invasiveness and metastasis of SCLC. This approach requires the FGF
receptors to be phosphorylated, but not via PLCγ; thus, there may
be additional mechanisms connecting phosphorylation of the FGF
receptor to the activation of ERK1/2 phosphorylation. Future
studies should further examine how the saccharification of cell
surface molecules can activate specific intracellular signal
transduction pathways and how the intracellular signaling pathways
driving PSA-NCAM-mediated invasion and metastasis are regulated in
SCLC. These lines of inquiry may lead to new strategies for the
prevention or treatment of SCLC.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China under grant no. 81272596.
References
|
1
|
Inamura K and Ishikawa Y: Lung cancer
progression and metastasis from the prognostic point of view. Clin
Exp Metastasis. 27:389–397. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Neal JW, Gubens MA and Wakelee HA: Current
management of small cell lung cancer. Clin Chest Med. 32:853–863.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Manapov F, Klöcking S, Niyazi M, Levitskiy
V, Belka C, Hildebrandt G, Fietkau R and Klautke G: Primary tumor
response to chemoradiotherapy in limited-disease small-cell lung
cancer correlates with duration of brain-metastasis free survival.
J Neurooncol. 109:309–314. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kalemkerian GP: Advances in
pharmacotherapy of small cell lung cancer. Expert Opin
Pharmacother. 15:2385–2396. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Morabito A, Carillio G, Daniele G,
Piccirillo MC, Montanino A, Costanzo R, Sandomenico C, Giordano P,
Normanno N, Perrone F, et al: Treatment of small cell lung cancer.
Crit Rev Oncol Hematol. 91:257–270. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Paulson JC and Rademacher C: Glycan
terminator. Nat Struct Mol Biol. 16:1121–1122. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Janas T and Janas T: Membrane oligo- and
polysialic acids. Biochim Biophys Acta. 1808:2923–2932. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Foley DA, Swartzentruber KG and Colley KJ:
Identification of sequences in the polysialyltransferases ST8Sia II
and ST8Sia IV that are required for the protein-specific
polysialylation of the neural cell adhesion molecule, NCAM. J Biol
Chem. 284:15505–15516. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Falconer RA, Errington RJ, Shnyder SD,
Smith PJ and Patterson LH: Polysialyltransferase: A new target in
metastatic cancer. Curr Cancer Drug Targets. 12:925–939. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Al-Saraireh YM, Sutherland M, Springett
BR, Freiberger F, Morais G Ribeiro, Loadman PM, Errington RJ, Smith
PJ, Fukuda M, Gerardy-Schahn R, et al: Pharmacological inhibition
of polysialyltransferase ST8SiaII modulates tumour cell migration.
PLoS One. 8:e733662013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Dube DH and Bertozzi CR: Glycans in cancer
and inflammation - potential for therapeutics and diagnostics. Nat
Rev Drug Discov. 4:477–488. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Durbec P and Cremer H: Revisiting the
function of PSA-NCAM in the nervous system. Mol Neurobiol.
24:53–64. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Soroka V, Kasper C and Poulsen FM:
Structural biology of NCAM. Adv Exp Med Biol. 663:3–22. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sandi C: Stress, cognitive impairment and
cell adhesion molecules. Nat Rev Neurosci. 5:917–930. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Thompson MG, Foley DA, Swartzentruber KG
and Colley KJ: Sequences at the interface of the fifth
immunoglobulin domain and first fibronectin type III repeat of the
neural cell adhesion molecule are critical for its polysialylation.
J Biol Chem. 286:4525–4534. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kulahin N and Walmod PS: The neural cell
adhesion molecule NCAM2/OCAM/RNCAM, a close relative to NCAM. Adv
Exp Med Biol. 663:403–420. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Mühlenhoff M, Rollenhagen M, Werneburg S,
Gerardy-Schahn R and Hildebrandt H: Polysialic acid: Versatile
modification of NCAM, SynCAM 1 and neuropilin-2. Neurochem Res.
38:1134–1143. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Berois N and Osinaga E: Glycobiology of
neuroblastoma: Impact on tumor behavior, prognosis, and therapeutic
strategies. Front Oncol. 4:1142014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Colley KJ, Kitajima K and Sato C:
Polysialic acid: Biosynthesis, novel functions and applications.
Crit Rev Biochem Mol Biol. 49:498–532. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ćirović S, Vještica J, Mueller CA, Tatić
S, Vasiljević J, Milenković S, Mueller GA and Marković-Lipkovski J:
NCAM and FGFR1 coexpression and colocalization in renal tumors. Int
J Clin Exp Pathol. 7:1402–1414. 2014.PubMed/NCBI
|
|
21
|
Cassens C, Kleene R, Xiao MF, Friedrich C,
Dityateva G, Schafer-Nielsen C and Schachner M: Binding of the
receptor tyrosine kinase TrkB to the neural cell adhesion molecule
(NCAM) regulates phosphorylation of NCAM and NCAM-dependent neurite
outgrowth. J Biol Chem. 285:28959–28967. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nong L, Yin G, Ren K, Tang J and Fan W:
Periodic mechanical stress enhances rat chondrocyte area expansion
and migration through Src-PLCgamma1-ERK1/2 signaling. Eur J Cell
Biol. 89:705–711. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li J, Dai G, Cheng YB, Qi X and Geng MY:
Polysialylation promotes neural cell adhesion molecule-mediated
cell migration in a fibroblast growth factor receptor-dependent
manner, but independent of adhesion capability. Glycobiology.
21:1010–1018. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Dalva MB, McClelland AC and Kayser MS:
Cell adhesion molecules: Signalling functions at the synapse. Nat
Rev Neurosci. 8:206–220. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Chattopadhyaya B, Baho E, Huang ZJ,
Schachner M and Di Cristo G: Neural cell adhesion molecule-mediated
Fyn activation promotes GABAergic synapse maturation in postnatal
mouse cortex. J Neurosci. 33:5957–5968. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Eggers K, Werneburg S, Schertzinger A,
Abeln M, Schiff M, Scharenberg MA, Burkhardt H, Mühlenhoff M and
Hildebrandt H: Polysialic acid controls NCAM signals at cell-cell
contacts to regulate focal adhesion independent from FGF receptor
activity. J Cell Sci. 124:3279–3291. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Teicher BA: Targets in small cell lung
cancer. Biochem Pharmacol. 87:211–219. 2014. View Article : Google Scholar : PubMed/NCBI
|