Introduction
Despite significant scientific innovations in cancer
treatments over the last few decades, melanoma remains one of the
most aggressive malignant tumors, with increasing incidence, poor
prognosis and insensitivity to chemotherapy (1–3).
Melanoma is extremely difficult to treat once it has metastasized
with a short 5-year survival rate (4). The phenomenon of
epithelial-mesenchymal transition (EMT) with characteristic changes
in the loss of epithelial markers (E-cadherin and α-catenin) and
the gain of mesenchymal markers (vimentin and N-cadherin) have been
well documented in cancer invasion and metastasis (5–9).
B-lymphoma Moloney murine leukemia virus insertion
region-1 (Bmi-1), the first member of the polycomb group (PcG) gene
family identified in mammals, plays an essential role in the
self-renewal of stem cells and functions as an oncogene in human
malignancies as well (5,6,8,10–12).
Bmi-1 was found to be highly expressed in cell lines from
metastatic melanoma compared to cell lines from primary melanoma,
and may be relevant for the metastatic behavior of melanoma cells
(13). The rapidly growing
information shows that Bmi-1 overexpression can induce EMT
properties and promote the metastasis of cancers, such as
nasopharyngeal carcinoma, endometrial and breast cancer (6,8,10). In
cultured human melanoma WM115A cells, it was reported that
silencing of Bmi-1 induced upregulation of E-cadherin, one of the
key epithelial markers of EMT (14). This evidence suggests that
overexpression of Bmi-1 induces EMT-like changes and promotes
metastasis in melanoma. However, the effects of Bmi-1 on EMT-like
changes, invasion and proliferation of melanoma have not been fully
addressed and the underlying mechanism of Bmi-1 mediating EMT-like
changes in melanoma remains currently unclear.
Nuclear factor-κB (NF-κB), as a transcriptional
factor, regulates the transcription of genes in several critical
pathways in cancer biology, such as apoptosis, angiogenesis,
proliferation and tumor cell invasion and metastasis (4,15).
Several studies show that activation of the NF-κB pathway directly
regulates EMT in melanoma (4).
Moreover, the activation of NF-κB could lead to the inhibition of
phosphatase and tensin homolog (PTEN) transcription in non-small
cell lung cancer cells (16). The
tumor-suppressor PTEN plays a significant role in cell cycle
arrest, regulation of cell adhesion, migration and proliferation
(4). It has been reported that PTEN
participates in Bmi-1-induced EMT in human nasopharyngeal
epithelial cells (8). Hereby,
whether NF-κB and PTEN participate in Bmi-1-induced EMT-like
changes in melanoma cells have not been fully addressed.
The present study focused on the effects of the
silencing of Bmi-1 on EMT-like changes in cultured melanoma cells
in vitro and on a human melanoma mouse xenograft model in
vivo. We demonstrated that Bmi-1 knockdown reduced melanoma
cell invasion detected by Matrigel invasion assay and tumor growth
evaluation in xenografts, respectively. Increased expression of
epithelial marker genes and decreased expression of mesenchymal
marker genes indicate the reversal of EMT-like changes induced by
silencing of Bmi-1 in melanoma. In addition, knockdown of Bmi-1 led
to the suppression of NF-κB and MMP-2 while significantly
upregulating PTEN. Collectively, these results provide evidence
that knockdown of Bmi-1 significantly inhibited the aggressive
behavior of melanoma cells through the reversal of EMT-like changes
via the regulation of related genes of the NF-κB and PTEN
pathways.
Materials and methods
Cell culture
The melanoma cell lines A375, C8161, SK-Mel-5 and
Mel-Juso were cultured in Dulbeccos modified Eagles medium (DMEM)
supplemented with 10% fetal bovine serum (Gibco, Grand Island, NY,
USA). All cultures were maintained in a 5% CO2 incubator
at 37°C. After detecting the Bmi-1 levels in the aforementioned
cell lines, the optimal cells were selected for further study.
Bmi-1 silencing by shRNA
The siRNA target sequence for the Bmi-1 gene
(NM_005180) was designed and the si-Bmi-1 constructs were inserted
into the GV248 green fluorescent protein-expressing lentiviral
vector (GeneChem, Shanghai, China). After a series of programmes to
transfect, concentrate and purify, Bmi-1 shRNA lentivirus and empty
lentivirus were obtained with a range of titer yields:
1–1.5×109 TU/ml and transfected into A375 cells
(MOI=20). The expression of Bmi-1 in A375 cells after transfection
was confirmed by real-time PCR and western blot analysis.
Real-time quantitative RT-PCR
Total RNA from A375, Bmi-1-knockdown
(A375-Bmi-1-shRNA) and pMSCV vector control cells (A375-vector)
were extracted using the RNeasy Midi kit (Qiagen, Inc., Valencia,
CA, USA) and reverse transcribed to cDNA with a PrimeScript RT
reagent kit (Takara, Tokyo, Japan). Real-time PCR was performed
using SYBR Premix Ex Taq (Takara) on a StepOnePlus™ Real-Time PCR
System (Applied Biosystems, Invitrogen). The cycling conditions
were as follows: 95°C for 30 sec, 95°C for 10 sec and 60°C for 30
sec. The following are the gene-specific primer pairs: Bmi-1,
5′-CTGGTTGCCCATTGACAGCG-3′ and 5′-AAATCCCGGAAAGAGCAGCC-3′;
E-cadherin, 5′-CGGTGGTCAAAGAGCCCTTA-3′ and
5′-TGAGGGTTGGTGCAACGTCGTTA-3′; α-catenin,
5′-GGCAGCCAAAAGACAACAGG-3′ and 5′-GGCCTTATAGGCTGCGACAT-3′;
vimentin, 5′-CAGGCAAAGCAGGAGTCCAC-3′ and
5′-GCAGCTTCAACGGCAAAGTTC-3′; N-cadherin,
5′-CGAATGGATGAAAGACCCATCC-3′ and 5′-GCCACTGCCTTCATAGTCAAACACT-3′;
and β-actin, 5′-TGGCACCCAGCACAATGAA-3′ and
5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′. The gene expression levels were
calculated using the 2−ΔΔCt method.
Western blot analysis
Total protein was extracted from the aforementioned
cultured cells and quantified. Equal protein amounts were analyzed
using 10% SDS-PAGE and transferred to a polyvinylidene difluoride
membrane. After blocking with a non-specific antibody binding with
non-fat dried milk for 2 h at room temperature, the membranes were
incubated overnight with primary antibodies at 4°C and then
incubated with a horseradish peroxidase-conjugated secondary
antibody for 2 h at room temperature. The primary antibodies used
were: anti-Bmi-1 (Abcam, Cambridge, UK), anti-E-cadherin,
anti-α-catenin (both from ProteinTech, Chicago, IL, USA),
anti-vimentin, anti-N-cadherin (both from Zhongshan Co., Beijing,
China), anti-PTEN (Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA), anti-Akt, anti-phospho-Akt (both from Cell Signaling
Technology, Inc., Beverly, MA, USA), anti-NF-κB (ProteinTech),
anti-phospho-NF-κB (Cell Signaling Technology, Inc.), anti-MMP-2
(ProteinTech) and anti-β-actin (Santa Cruz Biotechnology,
Inc.).
Immunofluorescent staining
The cells were fixed in 4% paraformaldehyde,
permeabilized in a solution of 0.3% Triton X-100 and 5% bovine
serum albumin (BSA) for 30 min at room temperature and incubated
with antibodies against E-cadherin, α-catenin, vimentin or
N-cadherin overnight at 4°C. Subsequently the cells were incubated
with the appropriate Cy3-conjugated secondary antibody (Jackson
ImmunoResearch Laboratories, West Grove, PA, USA) at 37°C for 1 h.
The nuclei were counterstained with DAPI and imaged with a confocal
laser-scanning microscope (Nikon, Tokyo, Japan).
Immunohistochemistry
After dewaxing and rehydration, the sections were
subjected to antigen retrieval by boiling in 10 mM citrate buffer.
After incubation with 3% hydrogen peroxide for 20 min to block the
endogenous peroxidase activity, the sections were immersed in 5%
bovine serum for 30 min to minimize antibody non-specificity. Next,
the sections were incubated with antibodies (anti-E-cadherin,
anti-α-catenin, anti-vimentin and anti-N-cadherin) overnight at 4°C
and then washed. The sections were then treated with secondary
antibodies (horseradish peroxidase-labeled goat anti-mouse/rabbit;
Zhongshan Co.) for 30 min at 37°C. Following development by
diaminobenzidine (Zhongshan Co.), the sections were counterstained
with hematoxylin and evaluated using microscopy.
Matrigel invasion assays
The Transwell polycarbonate membranes containing
8-µm pores were coated with Matrigel (BD Biosciences, San Jose, CA,
USA). Cells were resuspended in serum-free DMEM and seeded into the
upper wells, and DMEM supplemented with 15% bovine serum was placed
into the lower chamber (6). The
cells that migrated through the membranes were observed with
crystal violet staining after incubation for 24 h at 37°C.
Xenograft tumors
Six-week-old female BALB/c-nude mice were obtained.
Animals were housed in pathogen-free conditions with filtered air,
autoclaved food and water was available. A375-vector,
A375-Bmi-1-shRNA and 4×106 cells in serum-free DMEM were
injected subcutaneously into mice on their right flanks (six
mice/group). Tumor xenografts were measured once/week with
calipers. Mice were observed with live small-animal imaging
technology at the end of the fifth week. Tumor xenografts were
harvested for histological analysis. The volume was calculated as:
ab2/2 (a and b represent the length and width of a
tumor, respectively) and the tumor weights were measured. These
data were statistically compared by a two-sample t-test, with a
significance level of P<0.05. The histology and
immunohistochemistry of the tumors were observed under a
microscope.
Results
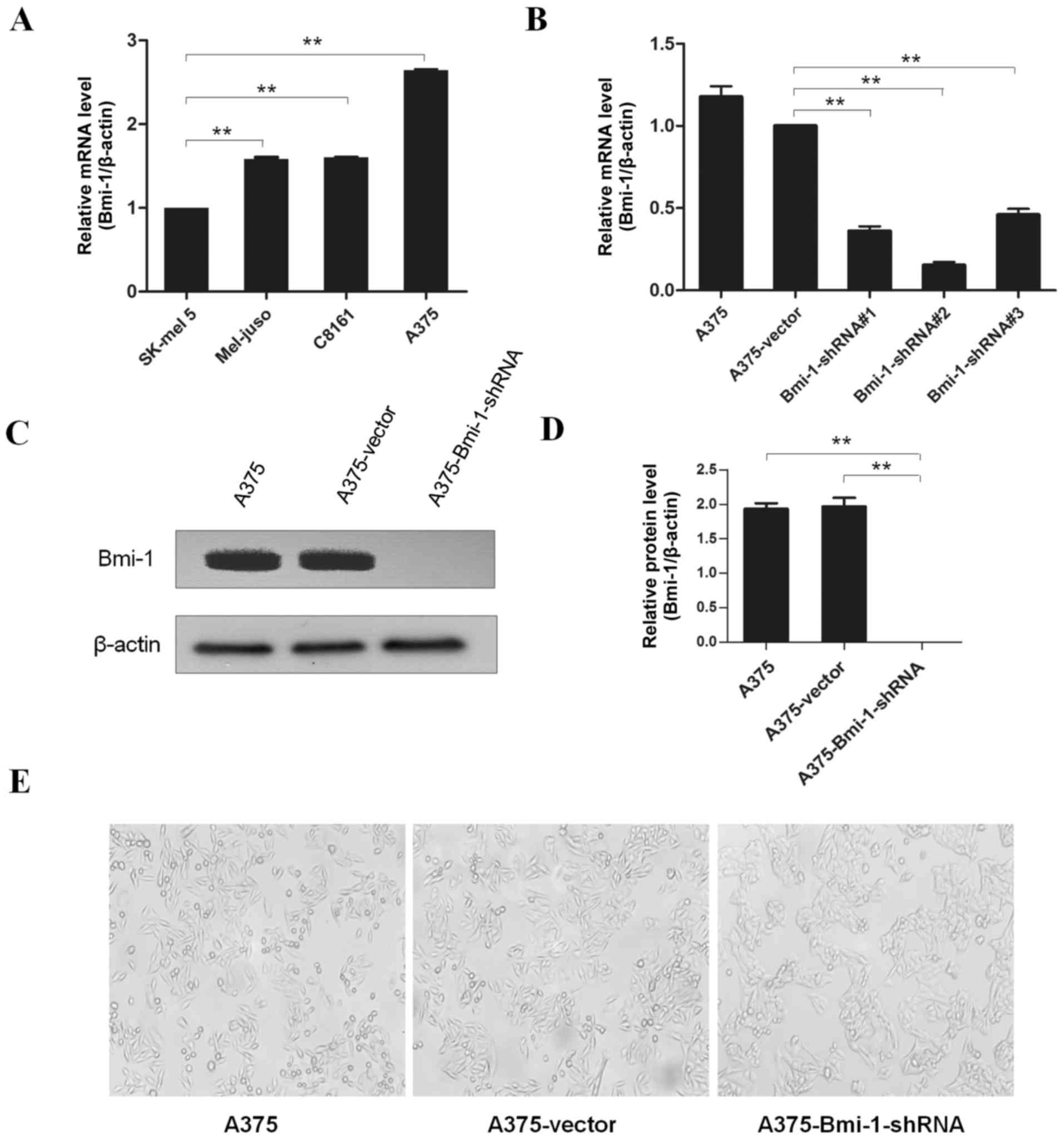
Silencing of Bmi-1 in cultured human
A375 cell line (A375-Bmi-1-shRNA)
To investigate the impact of Bmi-1 on EMT, we
compared the expression level of Bmi-1 in several melanoma cell
lines, and the A375 cell line was selected for further study
(Fig. 1A). We silenced endogenous
Bmi-1 in the A375 cells using specific shRNAs and knockdown effects
were detected by real-time PCR and western blotting. All three
shRNAs specifically knocked down the mRNA levels of Bmi-1. Due to
the best efficacy of shRNA #2, it was chosen for subsequent studies
(Fig. 1B). As shown in Fig. 1, Bmi-1 was significantly silenced
both at the mRNA and protein levels (Fig. 1B-D).
Silencing of Bmi-1 reduces A375 cell
invasion and reverses the EMT-like phenotype in vitro
Significant morphological differences were observed
in the A375-Bmi-1-shRNA cells in that there were more cohesive
cells with epithelioid changes compared to the spindle-shaped
normal A375 cells (Fig. 1E). The
A375-Bmi-1-shRNA cells gathered more closely to each other, while
the normal A375 cells appeared as spindle-like with a fibroblastic
morphology (Fig. 1E). Meanwhile,
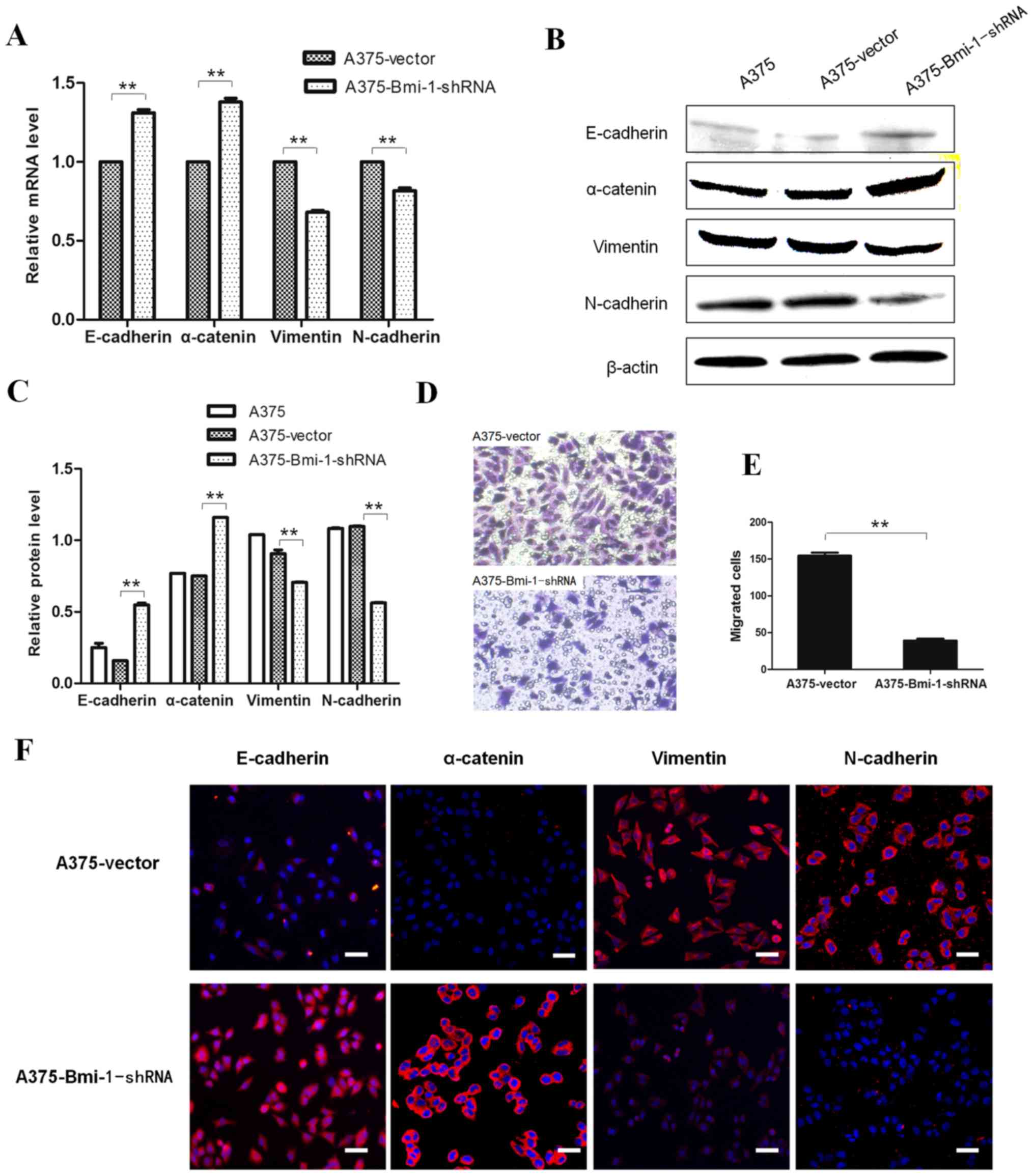
Matrigel invasion chamber assays revealed significantly reduced
invasion of the A375-Bmi-1-shRNA cells when compared to the control
cells (Fig. 2D and E).
EMT marker expression following the silencing of
Bmi-1 in A375 cells was detected and showed increased expression of
epithelial markers (E-cadherin and α-catenin) with concomitantly
decreased expression of mesenchymal markers (vimentin and
N-cadherin) compared to the A375 control cells (Fig. 2A-C and F). Thus, our results
suggested that Bmi-1 knockdown was crucial to reverse EMT-like
changes and suppress invasiveness of cultured A375 cells in
vitro.
Silencing Bmi-1 supresses melanoma
growth and reverses EMT-like changes in a human melanoma mouse
xenograft model in vivo
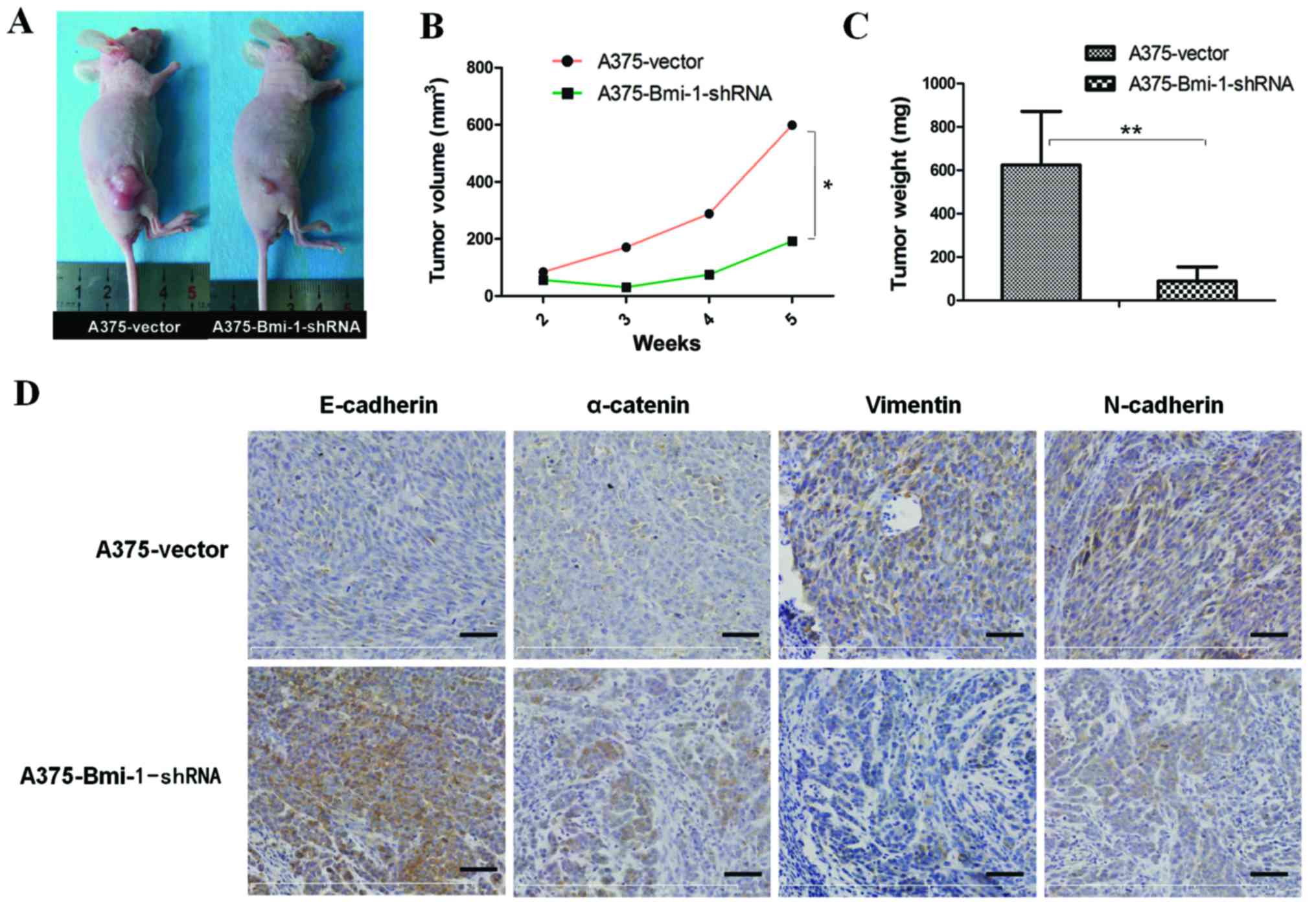
To further evaluate the effect of Bmi-1 knockdown on
tumor growth and EMT properties in vivo, six-week-old female
BALB/c-nude mice were injected with the A375-vector and the
A375-Bmi-1-shRNA cells in their flanks (six mice for each group).
Both the A375-vector cells and the A375-Bmi-1-shRNA cells formed
palpable tumors after 10 days, however, there was no marked
difference between the two groups until the third week (Fig. 3B). Tumors in the control group grew
quickly in the fourth and fifth week, while tumors in the
A375-Bmi-1-shRNA group grew significantly slower with no visible
substantial changes (Fig. 3B). The
mice were observed for five weeks and the xenograft tumors were
harvested. The tumor nodules formed by the A375-Bmi-1-shRNA cells
were significantly smaller than those formed by the A375 control
cells at the end of the experiment (day 35) (Fig. 3A and B). As the data showed
(Fig. 3B and C), the average volume
(A375-vector = 598.95±361.11 mm3, A375-Bmi-1-shRNA =
192.17±164.26 mm3, n=6, P=0.031) and the average of the
weight (A375-vector = 624.2±246.8 mg, A375-Bmi-1-shRNA =
89.67±65.28 mg, n=6, P=0.000) had significant differences between
the two groups.
To confirm that the silencing of Bmi-1 suppresses
melanoma proliferative ability by reversing EMT tumorigenicity in
mice, we evaluated the expression of EMT markers in tumor xenograft
tissues. Compared with tumors formed by the A375-vector cells, the
expression levels of E-cadherin and α-catenin were upregulated,
while vimentin and N-cadherin were downregulated in the tumors
formed by the A375-Bmi-1-shRNA cells (Fig. 3D). The findings were consistent with
the gene expression of EMT markers in the Bmi-1-silenced A375
cells.
Silencing of Bmi-1 reverses EMT-like
changes via the upregulation of PTEN and downregulation of
p-Akt/p-NF-κB/MMP-2 in A375 cells
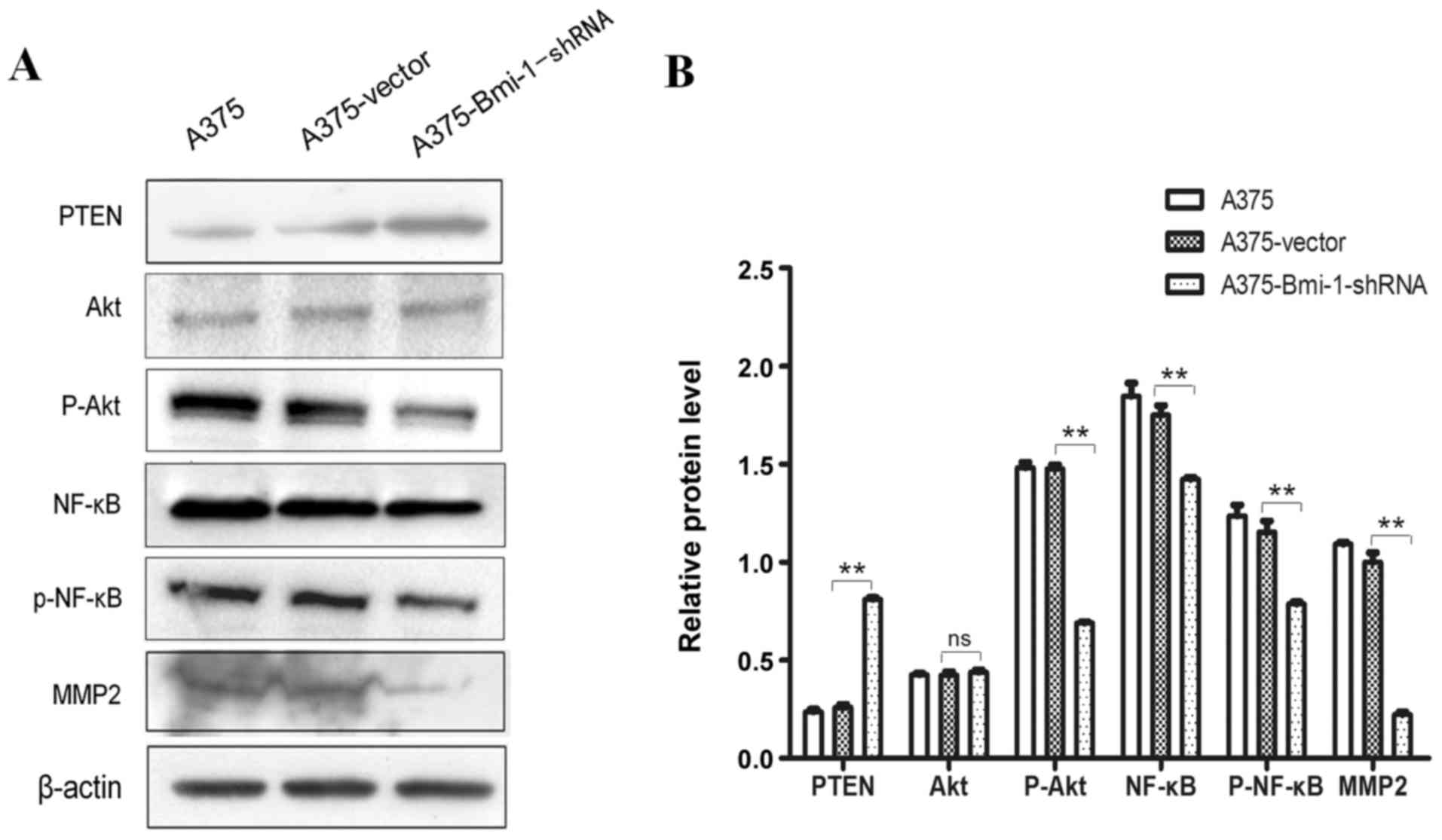
The tumor-suppressor PTEN, a downstream target of
Bmi-1, was increased in the A375-Bmi-1-shRNA cells, while its
targeted downstream molecules p-Akt, p-NF-κB and matrix
metalloproteases-2 (MMP-2) were reduced when Bmi-1 was knocked down
(Fig. 4). On the one hand,
activation of Akt activated the NF-κB pathway, which promoted EMT
in melanoma. On the other hand, the hyperactivated NF-κB pathway
contributed to EMT properties by enhancing MMP-2 syntheses in
melanoma. These results demonstrate that the silencing of Bmi-1
suppressed melanoma invasion via the regulation of
PTEN/Akt/NF-κB/MMP-2 in A375 cells.
Discussion
Evidence has demonstrated that EMT plays a crucial
role in the invasion and metastasis in cancers (17,18).
The EMT process occurs at the invasive front and produces single
migratory cells with increased vimentin and loss of E-cadherin in
colon and breast carcinoma, and melanoma (17–19).
Moreover, there are other events that are highly relevant to
EMT-promoting tumor progression, such as acceleration of
proliferation and resistance to cell death, senescence,
chemotherapy and immunotherapy, as well as conferring cancer cells
with stem cell-like properties (17,20–22).
Bmi-1 plays an essential role in the self-renewal of
stem cells and its overexpression is involved in the proliferation,
metastatic behavior and drug resistance of melanoma cells (5,6,8,10,11,13,14).
Overexpression of Bmi-1 induces EMT and promotes cell proliferation
in oral epithelial cells (23).
Previous studies have demonstrated that suppressing the EMT process
could inhibit cell proliferation and tumor growth in breast cancer
and squamous cell carcinoma (22,24).
Wound-healing assays have shown an obvious decrease in cell
migration ability after the silencing of Bmi-1 in WM115A melanoma
cells (14).
In the present study, the silencing of Bmi-1
significantly reduced the invasiveness of A375 cells in
vitro as evidenced by observations of cell morphology and
Matrigel invasion assays. Further studies focused on the change in
the EMT process in the Bmi-1-silenced A375 melanoma cells. Compared
with the A375-vector cells, EMT-related epithelial markers
(E-cadherin and α-catenin) were increased and EMT-related
mesenchymal markers (vimentin and N-cadherin) were decreased in the
A375-Bmi-1-shRNA cells. Our results confirmed that Bmi-1 was
crucial to inducing melanoma EMT and enhancing invasiveness in
vitro.
Moreover, compared with controls, the average volume
and weight of xenograft tumors formed by the A375-Bmi-1-shRNA cells
were reduced in the late stages, which suggested that tumorigenesis
ability decreased when Bmi-1 was silenced in the A375 cells. To
analyze tumorigenesis differences between the two groups, we
detected the expression of Ki67 (data not shown) and found that it
was decreased in tumors formed by the A375-Bmi-1-shRNA cells
compared to the control group. This confirmed that silencing of
Bmi-1 actually inhibited the self-renewing process of melanoma
cells, however, there was no marked difference in visible tumor
mass between the two groups until the third week. Moreover,
necrosis was much more obvious in tumors formed by the A375-vector
cells compared to those formed by the A375-Bmi-1-shRNA cells.
Necrosis may have resulted from the rapid growth of tumors without
abundant blood vessel supply. We speculated that the reversal of
EMT induced by silencing Bmi-1 in the A375 cells contributed more
significantly to inhibiting tumor expansion.
In accordance with the detection in vitro,
the expression of epithelial markers (E-cadherin and α-catenin) was
increased and mesenchymal markers (vimentin and N-cadherin)
significantly decreased in xenografts tissues formed by the
A375-Bmi-1-shRNA cells. Based on these results, we proposed that
Bmi-1 silencing weakened the invasive potential by suppressing EMT
in A375 cells, which led to the delay of tumor graft growth and
expansion. These data implied that the tumor mass expansion could
be induced not only by cell proliferation, but also by EMT.
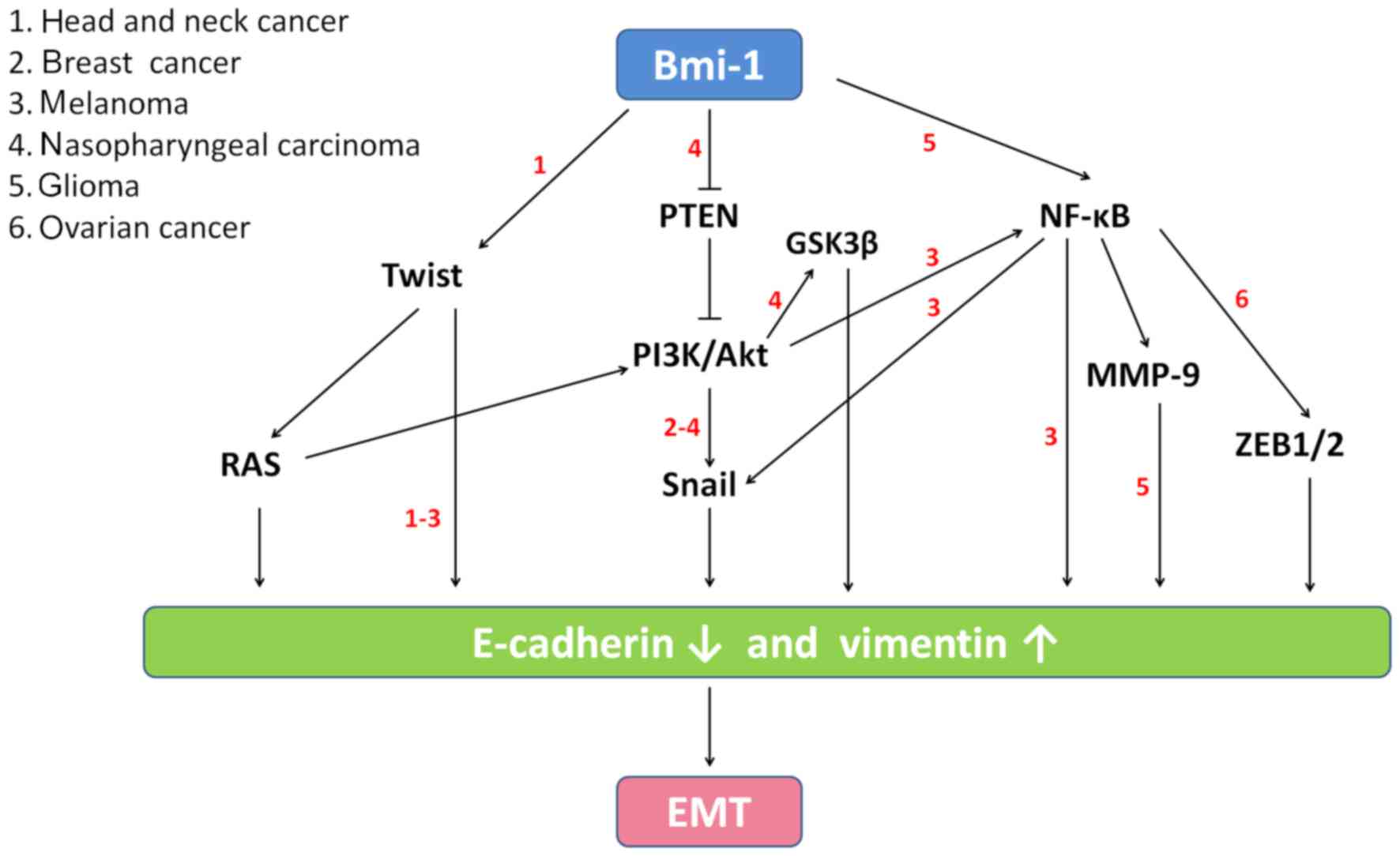
Different pathways (Fig.
5) are involved in the Bmi-1 regulation of EMT in various types
of cancer (4,8,25–27).
However, the mechanism by which Bmi-1 induces EMT in melanoma is
not clear. In the present study, we also addressed how Bmi-1
mediated EMT in melanoma. The tumor suppressor PTEN plays a
significant role in cell cycle arrest, regulation of cell adhesion,
migration and proliferation (4).
PTEN regulates the switch from E- to N-cadherin and activates the
PTEN/PI3K/Akt pathways in melanoma (4,8,28).
Furthermore, activation of Akt has been shown to activate the NF-κB
pathway, which could directly regulate EMT in melanoma (4).
NF-κB upregulates MMPs leading to EMT and malignant
characteristics have been reported in breast cancers, osteosarcoma
and glioma (27,29,30).
MMPs degrade structural components of the extracellular matrix,
contribute to EMT, and promote tumor invasion and metastasis
(7,31). MMP-2 plays an important role in
melanoma invasion and correlates with progression and survival
(7).
Whether the PTEN/PI3K/Akt/NF-κB/MMP pathway
participates in Bmi-1-induced EMT in melanoma cells has not been
previously elucidated. In the present study, Bmi-1 knockdown
increased PTEN and reduced the levels of p-Akt, p-NF-κB and MMP-2,
suggesting that the PTEN/PI3K/Akt/NF-κB/MMP pathway participated in
Bmi-1 regulation of EMT.
In summary, the present study demonstrates that
Bmi-1 plays a critical role in the proliferation and metastasis of
melanoma. Silencing of Bmi-1 significantly reversed EMT in melanoma
in vitro and in vivo via the suppression of the the
PTEN/Akt/NF-κB pathway, accompanied by MMP-2 reduction. Hence, the
targeting of Bmi-1 plays a potential role in the treatment of
melanoma.
Acknowledgements
The present study was supported by the National
Natural Science Foundation of China (no. 81372912) and the
Northwest Hospital Research Fund.
References
|
1
|
Bedogni B, Warneke JA, Nickoloff BJ,
Giaccia AJ and Powell MB: Notch1 is an effector of Akt and hypoxia
in melanoma development. J Clin Invest. 118:3660–3670. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sinnberg T, Lasithiotakis K, Niessner H,
Schittek B, Flaherty KT, Kulms D, Maczey E, Campos M, Gogel J,
Garbe C, et al: Inhibition of PI3K-AKT-mTOR signaling sensitizes
melanoma cells to cisplatin and temozolomide. J Invest Dermatol.
129:1500–1515. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hu Z, Fan H, Lv G, Zhou Q, Yang B, Zheng J
and Cao W: 5-Aminolevulinic acid-mediated sonodynamic therapy
induces anti-tumor effects in malignant melanoma via
p53-miR-34a-Sirt1 axis. J Dermatol Sci. 79:155–162. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lin K, Baritaki S, Militello L, Malaponte
G, Bevelacqua Y and Bonavida B: The role of B-RAF mutations in
melanoma and the induction of EMT via dysregulation of the
NF-κB/Snail/RKIP/PTEN circuit. Genes Cancer. 1:409–420. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Siddique HR and Saleem M: Role of BMI1, a
stem cell factor, in cancer recurrence and chemoresistance:
Preclinical and clinical evidences. Stem Cells. 30:372–378. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Dong P, Kaneuchi M, Watari H, Hamada J,
Sudo S, Ju J and Sakuragi N: MicroRNA-194 inhibits epithelial to
mesenchymal transition of endometrial cancer cells by targeting
oncogene BMI-1. Mol Cancer. 10:992011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Liu F, Garcia AM Gomez and Meyskens FL Jr:
NADPH oxidase 1 overexpression enhances invasion via matrix
metalloproteinase-2 and epithelial-mesenchymal transition in
melanoma cells. J Invest Dermatol. 132:2033–2041. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Song LB, Li J, Liao WT, Feng Y, Yu CP, Hu
LJ, Kong QL, Xu LH, Zhang X, Liu WL, et al: The polycomb group
protein Bmi-1 represses the tumor suppressor PTEN and induces
epithelial-mesenchymal transition in human nasopharyngeal
epithelial cells. J Clin Invest. 119:3626–3636. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ota I, Masui T, Kurihara M, Yook JI,
Mikami S, Kimura T, Shimada K, Konishi N, Yane K, Yamanaka T, et
al: Snail-induced EMT promotes cancer stem cell-like properties in
head and neck cancer cells. Oncol Rep. 35:261–266. 2016.PubMed/NCBI
|
|
10
|
Guo BH, Feng Y, Zhang R, Xu LH, Li MZ,
Kung HF, Song LB and Zeng MS: Bmi-1 promotes invasion and
metastasis, and its elevated expression is correlated with an
advanced stage of breast cancer. Mol Cancer. 10:102011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Song LB, Zeng MS, Liao WT, Zhang L, Mo HY,
Liu WL, Shao JY, Wu QL, Li MZ, Xia YF, et al: Bmi-1 is a novel
molecular marker of nasopharyngeal carcinoma progression and
immortalizes primary human nasopharyngeal epithelial cells. Cancer
Res. 66:6225–6232. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Xu X, Liu Y, Su J, Li D, Hu J, Huang Q, Lu
M, Liu X, Ren J, Chen W, et al: Downregulation of Bmi-1 is
associated with suppressed tumorigenesis and induced apoptosis in
CD44+ nasopharyngeal carcinoma cancer stem-like cells.
Oncol Rep. 35:923–931. 2016.PubMed/NCBI
|
|
13
|
Mihic-Probst D, Kuster A, Kilgus S,
Bode-Lesniewska B, Ingold-Heppner B, Leung C, Storz M, Seifert B,
Marino S, Schraml P, et al: Consistent expression of the stem cell
renewal factor BMI-1 in primary and metastatic melanoma. Int J
Cancer. 121:1764–1770. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Liu S, Tetzlaff MT, Cui R and Xu X:
miR-200c inhibits melanoma progression and drug resistance through
down-regulation of BMI-1. Am J Pathol. 181:1823–1835. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Madonna G, Ullman CD, Gentilcore G,
Palmieri G and Ascierto PA: NF-κB as potential target in the
treatment of melanoma. J Transl Med. 10:532012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xia D, Srinivas H, Ahn YH, Sethi G, Sheng
X, Yung WK, Xia Q, Chiao PJ, Kim H, Brown PH, et al:
Mitogen-activated protein kinase kinase-4 promotes cell survival by
decreasing PTEN expression through an NF kappa B-dependent pathway.
J Biol Chem. 282:3507–3519. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial- mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gheorgheosu D, Jung M, Ören B, Schmid T,
Dehelean C, Muntean D and Brüne B: Betulinic acid suppresses
NGAL-induced epithelial-to-mesenchymal transition in melanoma. Biol
Chem. 394:773–781. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ronca R, Di Salle E, Giacomini A, Leali D,
Alessi P, Coltrini D, Ravelli C, Matarazzo S, Ribatti D, Vermi W,
et al: Long pentraxin-3 inhibits epithelial-mesenchymal transition
in melanoma cells. Mol Cancer Ther. 12:2760–2771. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: Parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Xie G, Ji A, Yuan Q, Jin Z, Yuan Y, Ren C,
Guo Z, Yao Q, Yang K, Lin X, et al: Tumour-initiating capacity is
independent of epithelial-mesenchymal transition status in breast
cancer cell lines. Br J Cancer. 110:2514–2523. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Geng S, Guo Y, Wang Q, Li L and Wang J:
Cancer stem-like cells enriched with CD29 and CD44 markers exhibit
molecular characteristics with epithelial-mesenchymal transition in
squamous cell carcinoma. Arch Dermatol Res. 305:35–47. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Qiao B, Chen Z, Hu F, Tao Q and Lam AK:
BMI-1 activation is crucial in hTERT-induced epithelial-mesenchymal
transition of oral epithelial cells. Exp Mol Pathol. 95:57–61.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Fang X, Cai Y, Liu J, Wang Z, Wu Q, Zhang
Z, Yang CJ, Yuan L and Ouyang G: Twist2 contributes to breast
cancer progression by promoting an epithelial-mesenchymal
transition and cancer stem-like cell self-renewal. Oncogene.
30:4707–4720. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang MH, Hsu DS, Wang HW, Wang HJ, Lan HY,
Yang WH, Huang CH, Kao SY, Tzeng CH, Tai SK, et al: Bmi1 is
essential in Twist1-induced epithelial-mesenchymal transition. Nat
Cell Biol. 12:982–992. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ansieau S, Bastid J, Doreau A, Morel AP,
Bouchet BP, Thomas C, Fauvet F, Puisieux I, Doglioni C, Piccinin S,
et al: Induction of EMT by twist proteins as a collateral effect of
tumor-promoting inactivation of premature senescence. Cancer Cell.
14:79–89. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jiang L, Wu J, Yang Y, Liu L, Song L, Li J
and Li M: Bmi-1 promotes the aggressiveness of glioma via
activating the NF-kappaB/MMP-9 signaling pathway. BMC Cancer.
12:4062012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hao L, Ha JR, Kuzel P, Garcia E and Persad
S: Cadherin switch from E- to N-cadherin in melanoma progression is
regulated by the PI3K/PTEN pathway through Twist and Snail. Br J
Dermatol. 166:1184–1197. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Felx M, Guyot MC, Isler M, Turcotte RE,
Doyon J, Khatib AM, Leclerc S, Moreau A and Moldovan F:
Endothelin-1 (ET-1) promotes MMP-2 and MMP-9 induction involving
the transcription factor NF-kappaB in human osteosarcoma. Clin Sci.
110:645–654. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cichon MA and Radisky DC: ROS-induced
epithelial-mesenchymal transition in mammary epithelial cells is
mediated by NF-kB-dependent activation of Snail. Oncotarget.
5:2827–2838. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Radisky DC, Levy DD, Littlepage LE, Liu H,
Nelson CM, Fata JE, Leake D, Godden EL, Albertson DG, Nieto MA, et
al: Rac1b and reactive oxygen species mediate MMP-3-induced EMT and
genomic instability. Nature. 436:123–127. 2005. View Article : Google Scholar : PubMed/NCBI
|