Introduction
Gastric cancer is one of the most common
gastrointestinal tumors. Although its mechanism of development is
unclear, the invasive behavior and metastasis of cancer cells are
the main cause of death in patients with gastric cancer. In recent
years, many studies have suggested that epithelial-mesenchymal
transition (EMT) in epithelial tumors such as gastric cancer is an
important step in tumor metastasis (1). During the EMT process, cell
invasiveness and migration are enhanced, and tumor cells acquire
stem cell-like characteristics, which promote tumor invasion and
metastasis (2–4).
High mobility group protein A2 (HMGA2) is a
non-histone chromosomal protein, which plays a structural role as a
transcription factor primarily by binding the AT-rich regions in
the DNA sequence of target genes; this protein thus affects
embryogenesis, tissue development and tumorigenesis (5). In recent years, HMGA2 has been
reported to be highly expressed in tissues derived from thyroid
(6), colon (7) and gastric cancer (5). It has also been reported that HMGA2
can induce tumor cells to undergo EMT via interference of the cell
cycle (8–11), that HMGA2 can help tumor cells
acquire stem cell properties, and that it can promote tumor
metastasis (12). Recent studies
have demonstrated that HMGA2 may promote EMT in gastric cancer
(13) and promote an increase in
the expression of several tumor markers involved in breast cancer
(14). HMGA2 serves as a specific
downstream target of Lin28b, which regulates the self-renewal of
mouse hematopoietic stem cells (15). HMGA2 has been suggested to mediate
breast tumor metastasis via the promotion of the expression of LOX
and syndecan-2 (16). Previous
studies, including ours, have shown that HMGA2 may be responsible
for the invasiveness and metastasis of gastric cancer and for its
poor prognosis (5,13,17),
yet the related mechanism remains unclear.
Recently, TWIST proteins have been re-established as
important transcription factors associated with EMT (18,19);
Twist-related protein 1 (TWIST1) is most closely related to tumor
metastasis and angiogenic mimicry formation. Some studies have
reported that the potential combination of TWIST1 and E-box, which
is an E-cadherin promoter, negatively regulates the transcription
of the E-cadherin gene. This in turn induces EMT and the
acquisition of tumor stem cell properties, which enhances
invasiveness, metastasis and the angiogenic capacity of breast
cancer cells (20–22). One study revealed that TWIST1
enhanced cell migration beyond EMT, as TWIST1 induced the
activation of Rac1 (23). Yang
et al (24) confirmed that
TWIST1 may be incorporated into the intron region of the BMI1 gene
and that it can generate a stem cell-like phenotype and promote
tumor initiation. TWIST1 overexpression and poor tumor prognosis
are associated with a high rate of tumor invasion and metastasis
(25). Research has shown that
TWIST1 is involved in the malignant progression of colorectal
cancer (26), but no evidence has
indicated its involvement in gastric cancer.
Our previous study found that HMGA2 induced EMT in
gastric cancer cells and that this phenomenon was related to the
activation of the Wnt/β-catenin signaling pathway. Using
ChIP-on-chip by chromatin immunoprecipitation (ChIP) technology, we
also screened TWIST genes that may be associated with EMT in
gastric cancer cells (13,27). We speculated that HMGA2 may regulate
EMT and the acquisition of tumor stem cell properties via the
regulation of the downstream target gene TWIST1. Therefore, the aim
of the present study was to investigate the relationship between
HMGA2 and TWIST1 in gastric cancer and to determine whether HMGA2
can regulate EMT in gastric cancer cells, as well as the
acquisition of tumor stem cell properties, through the regulation
of TWIST1.
Materials and methods
Ethics statement
This study was approved by the Medical Ethics Review
Committee of the First Affiliated Hospital of Chongqing Medical
University. All of the procedures that involved animals were
conducted as indicated in the Guidelines of the National Institutes
of Health (NIH) for Animal Care (Guide for the Care and Use of
Laboratory Animals, Department of Health and Human Services, NIH
Publication no. 86–23, revised 1985). We made every effort to
minimize suffering and to minimize the number of animals used.
Patient collection
In total, 172 paraffin-embedded tissue specimens
(142 gastric cancer tissues and 30 peritumoral tissues) were
collected from the archives of the Department of Pathology,
Chongqing Medical University. These tissue samples were subjected
to the avidin-biotin-peroxidase method for immunohistochemical
analysis. An additional 72 fresh surgical specimens (36 gastric
cancer tissues and corresponding peritumoral tissues) were obtained
from the Department of Gastrointestinal Surgery, First Affiliated
Hospital of Chongqing Medical University. All specimens were frozen
in liquid nitrogen immediately after surgical resection and were
maintained at −80°C until protein extraction was performed. None of
the patients had received preoperative treatment, such as radiation
or neoadjuvant chemotherapy.
Immunohistochemical analysis
Tissue sections were deparaffinized in two changes
of xylene, and antigen retrieval was performed by heating the
sections in 0.01 M sodium citrate buffer in a microwave oven at
95°C for 15 min. The slides were incubated in 3% hydrogen peroxide
for 15 min and were then incubated in 0.5% Triton-X-100 (Beyotime
Biotech, Jiangsu, China) for 15 min. The slides were blocked with
5% BSA for 30 min and were incubated with rabbit primary antibodies
against human HMGA2 (1:100; Cell Signaling Technology, Danvers, MA,
USA) and TWIST1 (1:100, ProteinTech Group, Inc., Wuhan, China)
overnight at 4°C. Negative controls were treated identically but
were not incubated with the primary antibodies. Tissue sections
were washed in PBS (3×5 min), incubated with the appropriate
secondary antibody for 30 min, washed in PBS (3×5 min) and
incubated for 30 min with streptavidin-HRP (Beijing Zhongshan
Jinqiao Biotechnology Co., Ltd., Beijing, China). Immunoreactivity
was visualized using a DAB kit (Beijing Zhongshan Jinqiao
Biotechnology Co., Ltd.); the slides were counterstained with
hematoxylin and mounted with PVP. The percentage of positive cells
was scored according to the following criteria: 0, <10%; 1,
10–30%; 2, >30–50%; and 3, >50% (28). The patients were subsequently
categorized into either a positive expression group (score of 1–3)
or a negative expression group (score of 0).
Cell culture
MKN-45, MKN-28 and SGC-7901 human gastric carcinoma
cell lines were obtained from the Key Laboratory of General Surgery
at the First Affiliated Hospital of Chongqing Medical University.
The GES-1 human immortalized gastric epithelial cell line was
purchased from the Chinese Type Culture Collection. The cell lines
were cultured in RPMI-1640 medium (Hyclone, Shanghai, China)
supplemented with 10% fetal bovine serum (FBS) (Hyclone) and 2%
penicillin-streptomycin (Beyotime) in a humidified atmosphere of
95% air and 5% CO2 at 37°C. The cells were cultured
until a confluence of 80% was reached, at which point they were
passaged by trypsinization.
Western blot analysis
The cells were lysed in buffer containing 1% NP40, 1
mmol/l EDTA, 50 mmol/l Tris-HCl (pH 7.5), and 150 mmol/l NaCl
supplemented with a complete protease inhibitor mixture (Sangon
Biotech, Shanghai, China). Total proteins were separated by
SDS-PAGE and transferred to PVDF membranes (Millipore, Billerica,
MA, USA). The membranes were then blocked in 5% non-fat milk in TBS
for 2 h and probed with antibodies to HMGA2 (1:1,000; Cell
Signaling Technology), TWIST1 (1:1,000; Proteintech Group),
E-cadherin (1:100; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA), N-cadherin (1:1,000) and vimentin (1:1,000; both from
eBioscience, Inc., San Diego, CA, USA), Oct4 (1:1,000; San Ying
Biotechnology, Wuhan, China), CD44 (1:1,000; San Ying
Biotechnology) and GAPDH (1:500; Millipore). After the membranes
were washed, they were incubated with the appropriate secondary
antibodies for 1 h at 37°C. The results were visualized using an
enhanced chemiluminescence kit (Beyotime Biotech). Each band was
quantified using ImageJ software and normalized to GAPDH (Beyotime
Biotech).
Cell transfection
Human HMGA2 and TWIST1 cDNAs were cloned into
pLV-UbC-IRES2-EGFP via homologous recombination [S&E (Shanghai)
Bio-pharmaceutical, Shanghai, China]. The HMGA2 shRNA sequences,
the TWIST1 shRNA sequences and the scrambled RNA were synthesized
by Ribobio Corporation (Guangzhou, China). The TWIST1 shRNA
sequences and the scrambled RNA were cloned into the hU6-Neomycin
Vector by S&E (Shanghai) Bio-pharmaceutical. The cells were
transiently transfected with TWIST1 shRNA or scrambled RNA (con)
using Lipofectamine 2000 (Invitrogen Life Technologies, Shanghai,
China) according to the manufacturers instructions. The TWIST1
overexpression vector was purchased from S&E (Shanghai)
Bio-pharmaceutical. The sequences of the HMGA2- and TWIST1-specific
shRNAs used in these experiments were as follows: control-shRNA,
5-GAC GAGCGGCACGTGCACATT-3; HMGA2-shRNA, 5-CGGC
CAAGAGGCAGACCTATT-3; TWIST1-shRNA, 5-CGGAC AAGCTGAGCAAGATTC-3. The
cells were plated in 6-well clusters and transfected for 24 h or 48
h. Transfected cells were used in further assays or for RNA/protein
extraction.
Luciferase reporter assay
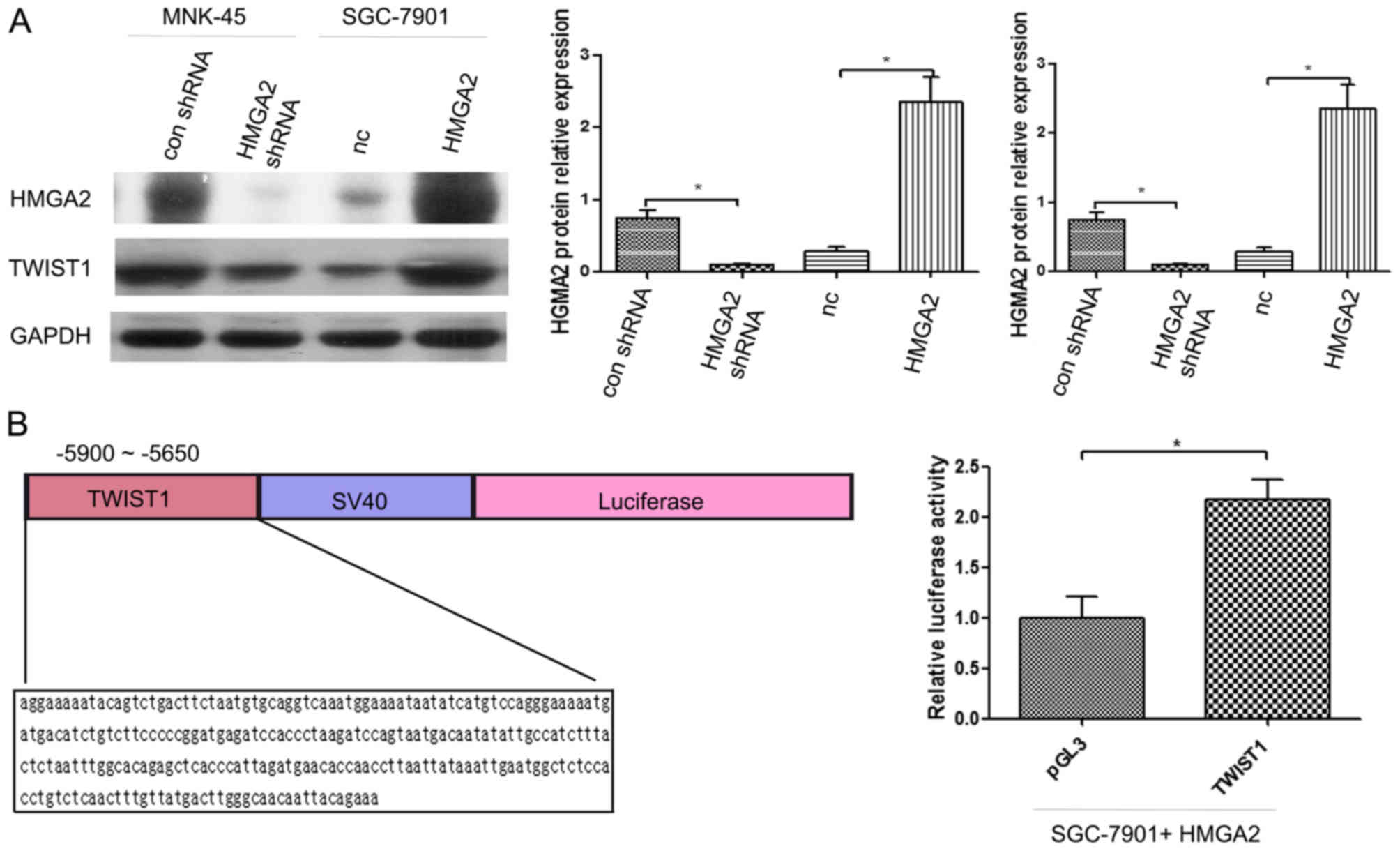
The ChIP-on-chip experiments showed that HMGA2 could
bind to the 5650–5900 bp region before the TWIST1 transcriptional
start site; therefore, this region was amplified by PCR and
inserted into the pGL3-Promoter luciferase reporter vector
(Promega, Madison, WI, USA). All constructs were verified by DNA
sequencing. SGC-7901 cells were plated in 96-well clusters and then
co-transfected with 100-ng constructs with or without the HMGA2
overexpression plasmid. At 48 h after transfection, the luciferase
activity was detected using a dual-luciferase reporter assay system
(Promega) and was normalized to Renilla luciferase activity.
All primers used were purchased from Genewiz Biotechnologies, Inc.
(Suzhou, China) and are as follows: forward,
5-TCTATCGATAGGTACCAGGAAAA ATACAGTCTGACTTC-3′ and reverse,
5-GATCGCAGATC TCGAGTTTCTGTAATTGTTGCCCAAG-3.
Wound healing assay
HepG2 and SMMC7721 cells were transfected with the
following: HMGA2 shRNA or HMGA2 overexpression vectors alone; HMGA2
shRNA combined with the TWIST1 overexpression clone; or the HMGA2
overexpression clone with TWIST1 shRNA. The transfected cells were
seeded in 6-well plates and allowed to grow to 100% confluence;
afterwards, a scratch was made across the cell monolayer. The cells
were rinsed with PBS and fresh growth medium was added. The cells
were incubated for 24 h and then imaged using a phase-contrast
microscope.
Cell invasion assay
For the cell invasion assay, 1×104 cells
were seeded on Matrigel-coated Transwell cell-culture inserts
(Invitrogen Life Technologies, Carlsbad, CA, USA) with RPMI-1640
medium with 2% FBS. The bottom chamber was filled with 600 µl
RPMI-1640 medium with 10% FBS. After 48 h of incubation, cells on
the lower surface of the membrane were stained with crystal violet.
The number of cells was counted in three random fields under a
light microscope.
Animal experiment
Female BALB/c mice at 6 weeks of age were obtained
from the National Biological Industry Base, Laboratory Animal
Center of Chongqing Medical University. The mice were randomly
divided into four groups (five mice per group) and were maintained
under pathogen-free conditions. The mice were intravenously
injected with MKN-45 cells, and the mice were observed using an
imaging system at one and two weeks after inoculation. The mice
were then dissected four weeks after inoculation to observe MKN-45
cell metastasis.
Statistical analysis
All experimental data are presented as the means ±
SD, and single comparisons between two groups were evaluated by
Students t-test using SPSS 20.0. Associations between the
expression levels of HMGA2 and TWIST1 were analyzed by the Pearson
correlation coefficient. P<0.05 was considered statistically
significant.
Results
Protein expression of HMGA2 and TWIST1
is high in gastric cancer according to immunohistochemistry and
western blot analysis
To investigate the association between HMGA2 and
TWIST1 expression in gastric cancer, we performed
immunohistochemical staining for these proteins in tissue specimens
from 142 gastric cancer patients. The staining for HMGA2 and TWIST1
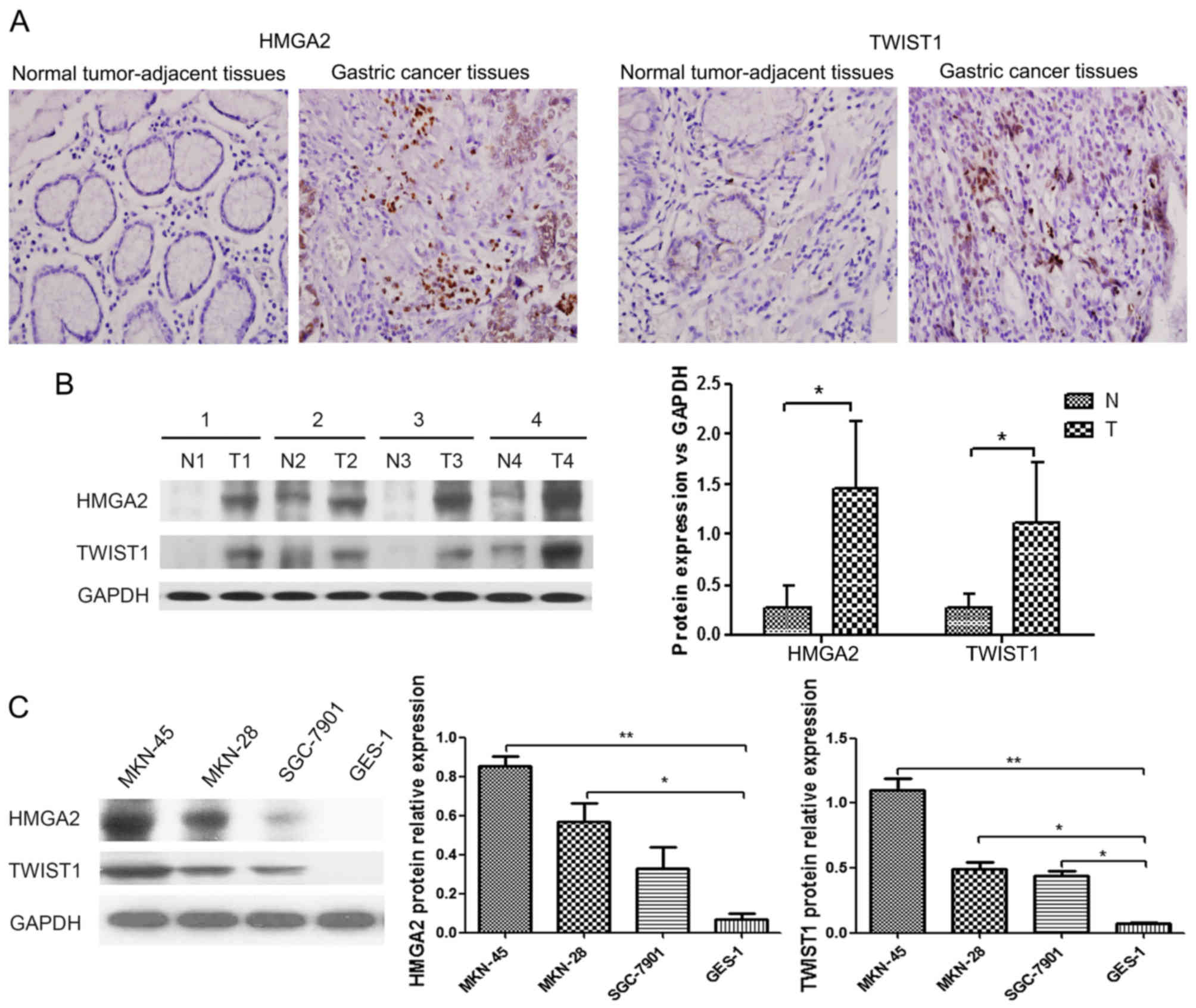
in representative clinical samples is shown in Fig. 1A. Among the 142 gastric cancer
specimens, 75 (52.8%) and 65 (45.8%) cases were positive for HMGA2
and TWIST1 expression, respectively. In addition,
immunohistochemical analysis showed that the expression of HMGA2
and that of TWIST1 were positively correlated (P<0.01; Table I). HMGA2 and TWIST1 were largely
expressed in the nucleus of gastric cancer cells. The expression
levels of HMGA2 and TWIST1 were significantly higher in gastric
cancer tissues compared with those in the peritumoral tissues
(P<0.01). Additionally, we tested the protein expression of
HMGA2 and TWIST1 in 36 paired tissue samples via western blot
analysis. As expected, the protein expression levels of HMGA2 and
TWIST1 were significantly higher in the gastric cancer tissues than
these levels the in peritumoral tissues (P<0.05; Fig. 1B).
 | Table I.Relationship between the protein
expression of HMGA2 and TWIST1 by immunohistochemistry. |
Table I.
Relationship between the protein
expression of HMGA2 and TWIST1 by immunohistochemistry.
|
| TWIST1
expression |
|
|
|
|---|
|
|
|
|
|
|
|---|
|
| High | Low | Total | χ2 | P-value |
|---|
| HMGA2 expression |
| High | 40 | 23 | 63 | 29.254 | <0.001 |
| Low | 15 | 64 | 79 |
|
|
| Total | 55 | 87 | 142 |
|
|
Expression of HMGA2 and TWIST1 in
gastric cancer cell lines
To investigate whether HMGA2 and TWIST1 levels are
also increased in gastric cancer cell lines compared with
immortalized gastric epithelial cells, we assessed the HMGA2 and
TWIST1 protein levels in the human gastric cancer cell lines
MKN-45, MKN-28, and SGC-7901, as well as in the human immortalized
gastric epithelial cell line GES-1, by western blot analysis. The
HMGA2 and TWIST1 protein levels were significantly higher in the
poorly differentiated gastric cancer cell line (MKN-45), the
moderately differentiated cell line (SGC-7901), and the
well-differentiated cell line (MKN-28) compared with the GES-1
cells (P<0.01; Fig. 1C).
TWIST1 is a direct target of HMGA2 in
gastric cancer cells
HMGA2 and TWIST1 protein expression levels in
various gastric epithelial cell lines such as MKN-45-con shRNA,
MKN-45-HMGA2 shRNA, SGC-7901-NC, and SGC-7901, in which HMGA2 was
originally overexpressed, were detected by western blot analysis.
It was found that after the application of HMGA2 shRNA in the
MKN-45 cells, the expression levels of HMGA2 and TWIST1 were
significantly decreased (P<0.05; Fig. 2A). After HMGA2 was overexpressed in
the SGC-7901 cells, the expression levels of TWIST1 and HMGA2 were
significantly increased (P<0.05; Fig. 2A). Simultaneously, we co-transfected
the pGL3-TWIST1-Promoter vector and the HMGA2 expression plasmid or
the negative control (NC) into SGC-7901 cells. The luciferase
activity of the HMGA2-overexpressing plasmid-transfected cells was
significantly increased compared with that of the NC-transfected
cells (Fig. 2B).
HMGA2 regulates the invasion and
migration of gastric cancer cells via TWIST1
The inhibition of HMGA2 suppressed the invasion and
migration of MKN-45 cells, and the simultaneous overexpression of
TWIST1 reversed the inhibitory effect of HMGA2 expression on the
invasion and migration of gastric cancer cells (Fig. 3A and B). The overexpression of HMGA2
significantly promoted the invasion and migration of SGC-7901
cells, whereas the simultaneous inhibition of TWIST1 expression
reversed the promotive effect of HMGA2 overexpression on the
invasion and migration of gastric cancer cells (Fig. 3A and B).
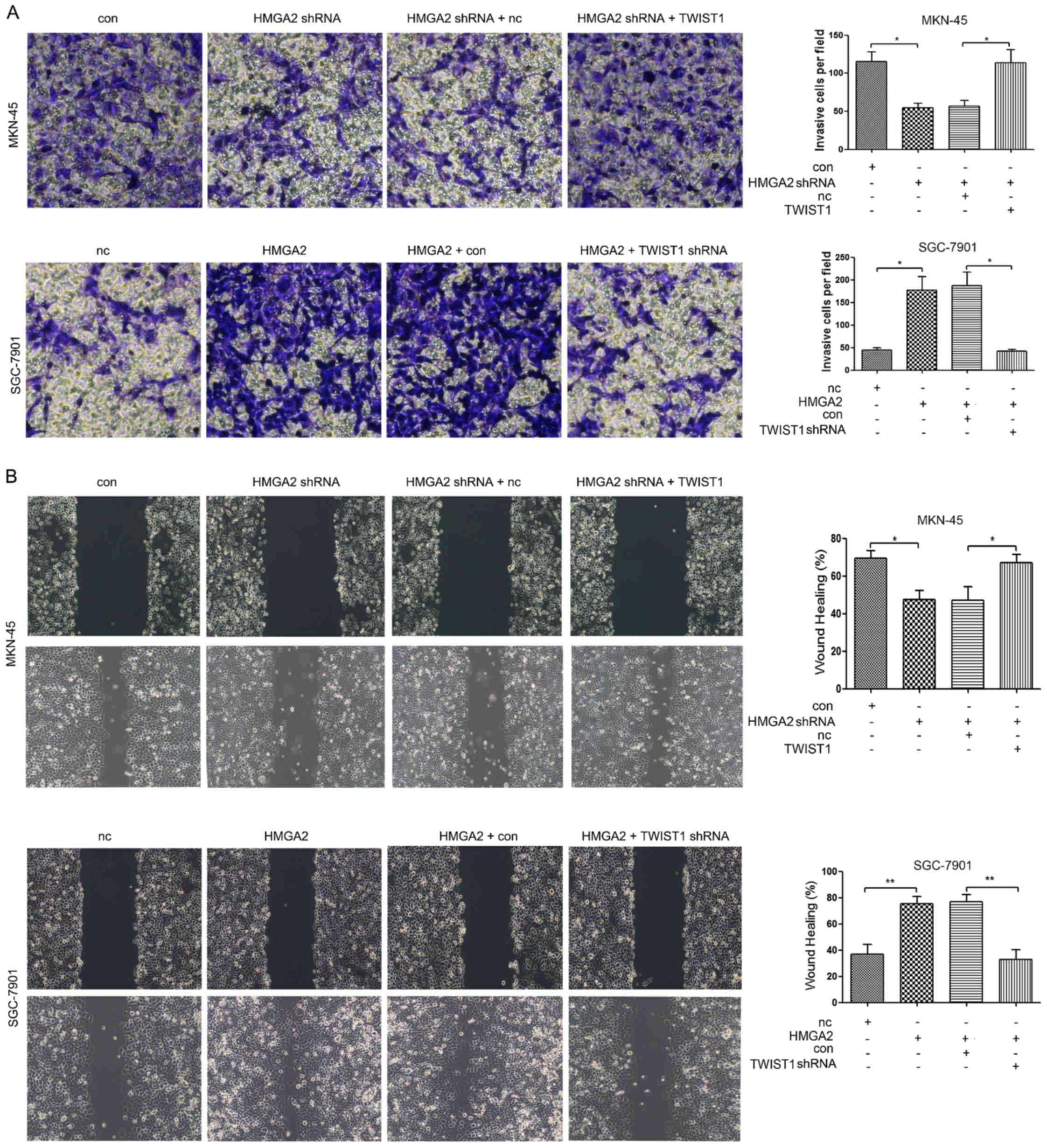 | Figure 3.HMGA2 affects the invasion and
migration of gastric cancer cells via TWIST1. (A) MKN-45 cells were
transfected with con, HMGA2 shRNA, HMGA2 shRNA + NC, or HMGA2 shRNA
+ TWIST1. Then, a Transwell assay was used to detect cell
invasiveness in each group; SGC-7901 cells were transfected with
NC, HMGA2, HMGA2 + con, or HMGA2 + TWIST1 shRNA. Then, a Transwell
invasion assay was used to detect cell invasiveness in each group.
(B) MKN-45 cells were transfected with con, HMGA2 shRNA, HMGA2
shRNA + NC, or HMGA2 shRNA + TWIST1; and then a scratch assay was
used to detect cell migration in each group. SGC-7901 cells were
transfected with NC, HMGA2, HMGA2 + con, or HMGA2 + TWIST1 shRNA;
and then a scratch assay was used to detect cell migration in each
group. *P<0.05, **P<0.01. HMGA2, high mobility group protein
A2. |
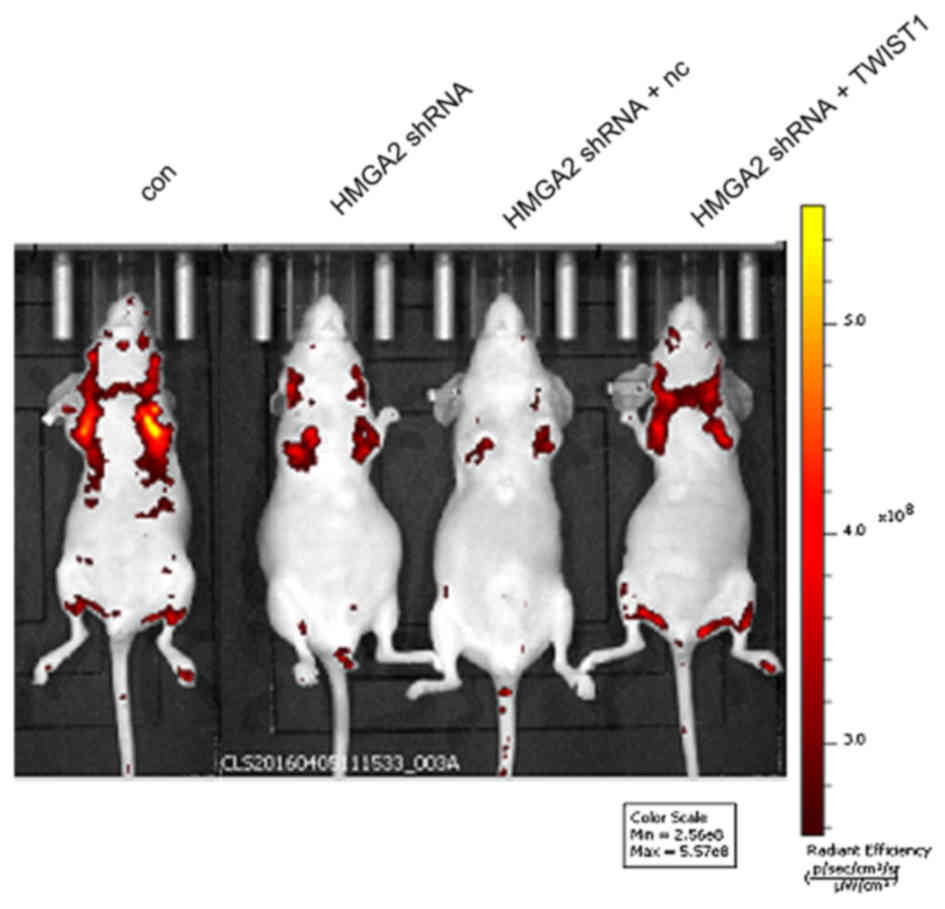
Overexpression of TWIST1 reverses the
inhibitory effect of HMGA2 abrogation on the metastasis of gastric
cancer cells in nude mice
To further investigate whether HMGA2 regulates the
invasion and migration of gastric cancer cells via TWIST1, we
injected MKN-45 cells into the tail veins of mice to detect the
in vivo metastasis of MKN-45 cells after the application of
HMGA2 shRNA. The results showed that the metastatic ability of
MKN-45 cells was significantly decreased after the abrogation of
HMGA2 expression, whereas the simultaneous overexpression of TWIST1
reversed the inhibitory effect of HMGA2 interference on the
metastasis of gastric cancer cells in nude mice (Fig. 4).
HMGA2 regulates EMT and the
acquisition of stem cell properties in gastric cancer cells via
TWIST1
The interference of HMGA2 expression notably
inhibited EMT of MKN-45 cells and the expression of the stemness
markers CD44 and OCT4. However, at the same time, the
overexpression of TWIST1 reversed the inhibitory effect of HMGA2
overexpression on EMT of gastric cancer cells and the expression of
the stemness markers CD44 and OCT4 (Fig. 5A and B). The overexpression of HMGA2
significantly promoted EMT in SGC-7901 cells and the expression of
the stemness markers CD44 and OCT4. However, the interference of
TWIST1 expression also reversed the promoter effect of HMGA2
overexpression on EMT in gastric cancer cells and the expression of
the stemness markers CD44 and OCT4 (Fig. 5A and B).
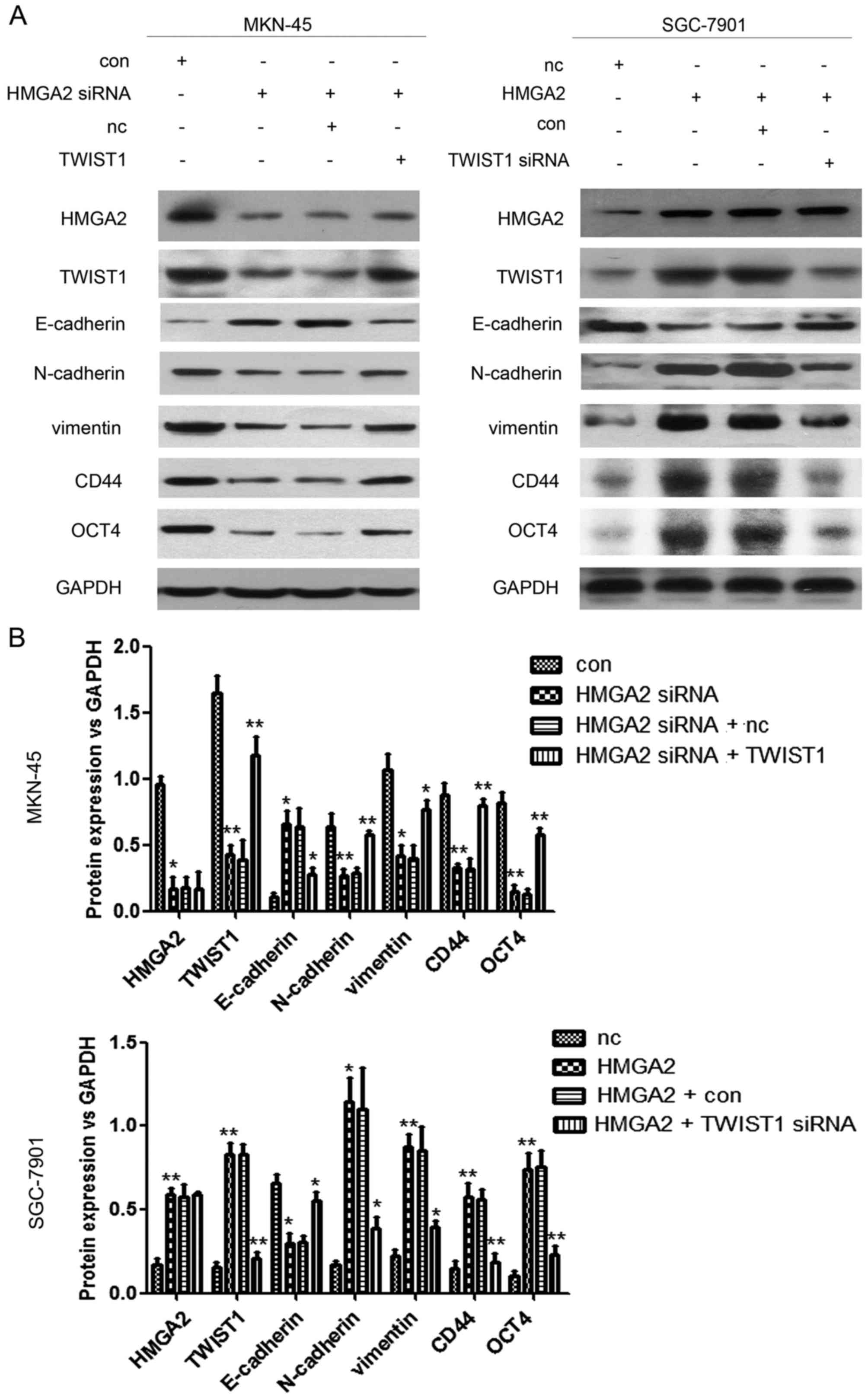 | Figure 5.HMGA2 affects EMT of gastric cancer
cells and the expression of stemness indicators via TWIST1. (A)
MKN-45 cells were transfected with con, HMGA2 shRNA, HMGA2 shRNA +
NC, or HMGA2 shRNA + TWIST1; and then EMT (E-cadherin, N-cadherin
and vimentin expression) and the expression of stemness markers
(CD44 and OCT4) in the cells were detected by western blot analysis
assay in each group. SGC-7901 cells were transfected with NC,
HMGA2, HMGA2 + con, or HMGA2 + TWIST1 shRNA; and then EMT
(E-cadherin, N-cadherin and vimentin expression) and the expression
of stemness markers (CD44 and OCT4) in the cells were detected by
western blot analysis assay in each group. The experiment was
repeated three times. (B) Quantification assays of each index of
the western blot analysis results. *P<0.05, **P<0.01. HMGA2,
high mobility group protein A2. |
Discussion
HMGA2 plays an important role in EMT and in cancer
stem cells (CSCs), but the mechanism by which HMGA2 stimulates
downstream target genes to induce EMT and the acquisition of tumor
stem cell properties remain unclear. The analysis of downstream
regulatory networks of HMGA2 may be an important clue in the
exploration of the correlation between EMT and CSCs.
We previously reported that HMGA2 can bind to the
TWIST1 promoter region in gastric cancer (13). Therefore, an important regulatory
relationship exists between HMGA2 and TWIST1. First, we detected
the expression of HMGA2 and TWIST1 in pathological specimens and
found that the immunohistochemical and protein expression of HMGA2
and TWIST1 in gastric cancer cells were high and showed a positive
correlation compared with that of matched normal adjacent tissues.
Furthermore, we detected the correlation between the protein
expression of HMGA2 and TWIST1 in gastric epithelial cell lines and
found that HMGA2 and TWIST1 were highly expressed in MKN-45 cells
and were lowly expressed in SGC-7901 and MKN-28 cells, which was
consistent with what has been observed in gastric cancer. To
investigate the changes in TWIST1 after its expression was blocked
and after the HMGA2 gene was overexpressed, we abrogated HMGA2
expression in MKN-45 cells and overexpressed HMGA2 in SGC-7901
cells. We found that the TWIST1 protein expression level was
correspondingly increased and decreased, respectively, in MKN-45
and SGC-7901 cells. To further confirm the regulatory effect of
HMGA2 on TWIST1, we constructed a reporter vector of TWIST1 and
found that HMGA2 significantly improved the promoter activity of
TWIST1. Therefore, both our preliminary results and this
experimental result indicate that TWIST1 is a direct downstream
target gene of HMGA2.
Previous studies have shown that the overexpression
of HMGA2 can significantly promote the invasion and migration of
gastric cancer cells (27), whereas
TWIST1 overexpression was also found to promote migration and
invasion in breast, esophageal and colorectal cancer (26,29–32).
However, whether HMGA2 regulates migration and invasion in gastric
cancer through TWIST1 has not been confirmed. First, we inhibited
the expression of HMGA2 in MKN-45 cells and simultaneously
overexpressed TWIST1; we also overexpressed HMGA2 in SGC-7901 cells
and simultaneously inhibited the expression of TWIST1. We then
observed the role of TWIST1 and HMGA2 on invasion and migration,
initiation of EMT and acquisition of tumor stem cell properties in
gastric cancer cells. The results showed that TWIST1 overexpression
reversed the decrease in the invasion and migration abilities and
EMT and the expression of stemness markers induced by HMGA2
interference in MKN-45 gastric cancer cells; the results also
showed that TWIST1 interference inhibited the increase in
HMGA2-mediated invasion and migration, EMT and the expression of
stemness markers induced by HMGA2 overexpression in SGC-7901
gastric cancer cells. To further validate the effect of TWIST1 on
the metastasis of gastric cancer cells, we performed in vivo
tumor metastasis experiments in nude mice. In this model, the
expression of HMGA2 was inhibited and TWIST1 was overexpressed in
MKN-45 cells. We found that TWIST1 overexpression could reverse the
inhibition of HMGA2 interference on the metastasis of MKN-45
cells.
In summary, HMGA2 can directly regulate the
expression of TWIST1, which induces migration and invasion, EMT and
the acquisition of tumor stem cell properties in gastric cancer
cells.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (no. 81272753). The authors
gratefully thank the Molecular Oncology and Epigenetics Laboratory
of the First Affiliated Hospital of Chongqing Medical University
for providing the clinical tissue specimens and the corresponding
data.
References
|
1
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial- mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sethi S, Macoska J, Chen W and Sarkar FH:
Molecular signature of epithelial-mesenchymal transition (EMT) in
human prostate cancer bone metastasis. Am J Transl Res. 3:90–99.
2010.PubMed/NCBI
|
|
3
|
Biddle A, Liang X, Gammon L, Fazil B,
Harper LJ, Emich H, Costea DE and Mackenzie IC: Cancer stem cells
in squamous cell carcinoma switch between two distinct phenotypes
that are preferentially migratory or proliferative. Cancer Res.
71:5317–5326. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen C, Wei Y, Hummel M, Hoffmann TK,
Gross M, Kaufmann AM and Albers AE: Evidence for
epithelial-mesenchymal transition in cancer stem cells of head and
neck squamous cell carcinoma. PLoS One. 6:e164662011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Motoyama K, Inoue H, Nakamura Y, Uetake H,
Sugihara K and Mori M: Clinical significance of high mobility group
A2 in human gastric cancer and its relationship to let-7 microRNA
family. Clin Cancer Res. 14:2334–2340. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Belge G, Meyer A, Klemke M, Burchardt K,
Stern C, Wosniok W, Loeschke S and Bullerdiek J: Upregulation of
HMGA2 in thyroid carcinomas: A novel molecular marker to
distinguish between benign and malignant follicular neoplasias.
Genes Chromosomes Cancer. 47:56–63. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Huang ML, Chen CC and Chang LC: Gene
expressions of HMGI-C and HMGI(Y) are associated with stage and
metastasis in colorectal cancer. Int J Colorectal Dis.
24:1281–1286. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Fusco A and Fedele M: Roles of HMGA
proteins in cancer. Nat Rev Cancer. 7:899–910. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hock R, Furusawa T, Ueda T and Bustin M:
HMG chromosomal proteins in development and disease. Trends Cell
Biol. 17:72–79. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Wu J, Liu Z, Shao C, Gong Y, Hernando E,
Lee P, Narita M, Muller W, Liu J and Wei JJ: HMGA2
overexpression-induced ovarian surface epithelial transformation is
mediated through regulation of EMT genes. Cancer Res. 71:349–359.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Chaw SY, Majeed AA, Dalley AJ, Chan A,
Stein S and Farah CS: Epithelial to mesenchymal transition (EMT)
biomarkers - E-cadherin, beta-catenin, APC and vimentin - in oral
squamous cell carcinogenesis and transformation. Oral Oncol.
48:997–1006. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Nishino J, Kim I, Chada K and Morrison SJ:
Hmga2 promotes neural stem cell self-renewal in young but not old
mice by reducing p16Ink4a and p19Arf
expression. Cell. 135:227–239. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zha L, Wang Z, Tang W, Zhang N, Liao G and
Huang Z: Genome-wide analysis of HMGA2 transcription factor binding
sites by ChIP on chip in gastric carcinoma cells. Mol Cell Biochem.
364:243–251. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Dave B, Mittal V, Tan NM and Chang JC:
Epithelial-mesenchymal transition, cancer stem cells and treatment
resistance. Breast Cancer Res. 14:2022012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Copley MR, Babovic S, Benz C, Knapp DJ,
Beer PA, Kent DG, Wohrer S, Treloar DQ, Day C, Rowe K, et al: The
Lin28b-let-7-Hmga2 axis determines the higher self-renewal
potential of fetal haematopoietic stem cells. Nat Cell Biol.
15:916–925. 2013. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Sun M, Gomes S, Chen P, Frankenberger CA,
Sankarasharma D, Chung CH, Chada KK and Rosner MR: RKIP and HMGA2
regulate breast tumor survival and metastasis through lysyl oxidase
and syndecan-2. Oncogene. 33:3528–3537. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kong D, Su G, Zha L, Zhang H, Xiang J, Xu
W, Tang Y and Wang Z: Coexpression of HMGA2 and Oct4 predicts an
unfavorable prognosis in human gastric cancer. Med Oncol.
31:1302014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang J, Mani SA, Donaher JL, Ramaswamy S,
Itzykson RA, Come C, Savagner P, Gitelman I, Richardson A and
Weinberg RA: Twist, a master regulator of morphogenesis, plays an
essential role in tumor metastasis. Cell. 117:927–939. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Eckert MA, Lwin TM, Chang AT, Kim J, Danis
E, Ohno- Machado L and Yang J: Twist1-induced invadopodia formation
promotes tumor metastasis. Cancer Cell. 19:372–386. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Pozharskaya V, Torres-González E, Rojas M,
Gal A, Amin M, Dollard S, Roman J, Stecenko AA and Mora AL: Twist:
A regulator of epithelial-mesenchymal transition in lung fibrosis.
PLoS One. 4:e75592009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Yu L, Li HZ, Lu SM, Tian JJ, Ma JK, Wang
HB and Xu W: Down-regulation of TWIST decreases migration and
invasion of laryngeal carcinoma Hep-2 cells by regulating the
E-cadherin, N-cadherin expression. J Cancer Res Clin Oncol.
137:1487–1493. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mani SA, Guo W, Liao MJ, Eaton EN, Ayyanan
A, Zhou AY, Brooks M, Reinhard F, Zhang CC, Shipitsin M, et al: The
epithelial-mesenchymal transition generates cells with properties
of stem cells. Cell. 133:704–715. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Yang WH, Lan HY, Huang CH, Tai SK, Tzeng
CH, Kao SY, Wu KJ, Hung MC and Yang MH: RAC1 activation mediates
Twist1-induced cancer cell migration. Nat Cell Biol. 14:366–374.
2012. View
Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yang MH, Hsu DS, Wang HW, Wang HJ, Lan HY,
Yang WH, Huang CH, Kao SY, Tzeng CH, Tai SK, et al: Bmi1 is
essential in Twist1-induced epithelial-mesenchymal transition. Nat
Cell Biol. 12:982–992. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
25
|
Sung CO, Lee KW, Han S and Kim SH: Twist1
is up-regulated in gastric cancer-associated fibroblasts with poor
clinical outcomes. Am J Pathol. 179:1827–1838. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Valdés-Mora F, Gómez del Pulgar T, Bandrés
E, Cejas P, Ramírez de Molina A, Pérez-Palacios R, Gallego-Ortega
D, García-Cabezas MA, Casado E, Larrauri J, et al: TWIST1
overexpression is associated with nodal invasion and male sex in
primary colorectal cancer. Ann Surg Oncol. 16:78–87. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zha L, Zhang J, Tang W, Zhang N, He M, Guo
Y and Wang Z: HMGA2 elicits EMT by activating the Wnt/β-catenin
pathway in gastric cancer. Dig Dis Sci. 58:724–733. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Xu MZ, Yao TJ, Lee NP, Ng IO, Chan YT,
Zender L, Lowe SW, Poon RT and Luk JM: Yes-associated protein is an
independent prognostic marker in hepatocellular carcinoma. Cancer.
115:4576–4585. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Mani SA, Yang J, Brooks M, Schwaninger G,
Zhou A, Miura N, Kutok JL, Hartwell K, Richardson AL and Weinberg
RA: Mesenchyme Forkhead 1 (FOXC2) plays a key role in metastasis
and is associated with aggressive basal-like breast cancers. Proc
Natl Acad Sci USA. 104:10069–10074. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cheng GZ, Chan J, Wang Q, Zhang W, Sun CD
and Wang LH: Twist transcriptionally up-regulates AKT2 in breast
cancer cells leading to increased migration, invasion, and
resistance to paclitaxel. Cancer Res. 67:1979–1987. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ai L, Kim WJ, Alpay M, Tang M, Pardo CE,
Hatakeyama S, May WS, Kladde MP, Heldermon CD, Siegel EM, et al:
TRIM29 suppresses TWIST1 and invasive breast cancer behavior.
Cancer Res. 74:4875–4887. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Lee KW, Sung CO, Kim JH, Kang M, Yoo HY,
Kim HH, Um SH and Kim SH: CD10 expression is enhanced by Twist1 and
associated with poor prognosis in esophageal squamous cell
carcinoma with facilitating tumorigenicity in vitro and in vivo.
Int J Cancer. 136:310–321. 2015. View Article : Google Scholar : PubMed/NCBI
|