Introduction
Immediate early response 2 (IER2) or ETR101, a human
homolog of mouse and rat for pip92 is a cellular immediate early
response protein that is induced by extracellular stimuli, such as
growth factors, 12-O-tetradecanoylphorbol-13-acetate, and some
infection of pathogens (1–4). Previous studies have demonstrated that
IER2 may function as a fibroblast growth factor intracellular
binding protein 1-interacting protein, or as a DNA-binding protein
acting as a transcriptional activator for myo-inositol 1-phosphate
synthase gene (3,5). Recently, evidence suggests that IER2
seems to play a pivotal role in tumor cell motility and metastasis
(6), however, little is known about
the mechanism involved. We have previously shown that IER2
regulated human umbilical vein endothelium cell motility, adhesion,
in vitro capillary tube formation, and the actin
cytoskeleton rearrangement in a FAK-dependent manner (7). Furthermore, we also found that IER2,
as a direct and functional target for miR-30c in the hepatocellular
carcinoma (HCC) cell lines (SMMC-7721 and HepG2 cells), may
function as a positive regulator in cell migration and invasion
(8), suggesting that IER2 may play
a role in the HCC cell motility.
HCC is the fifth most commonly diagnosed human
cancer worldwide, and the leading cause of death for the vast
majority of HCC patients is a result of the direct metastases
(9), which is a complex process
including cancer cells shed from the primary organization,
cell-extracellular matrix (ECM) adhesion, cell migration and
invading ECM, and each step of metastases is associated with
changes in gene expression involved in cell migration, invasion and
adhesion (10,11). Thus, understanding the underlying
mechanisms involved in these processes will likely contribute to
our insight to developing strategies for maximizing the efficacy of
HCC treatment. Previous studies have shown that IER2 is upregulated
in some metastatic cancer cells and in several human primary
tumors, such as pancreas cancer, breast cancer and HCC (6), and IER2 expression promotes tumor cell
migration and invasion and correlates with the colorectal
adenocarcinoma metastasis (6,8).
However, the biological roles and underlying mechanisms involved in
HCC of IER2 are largely unknown.
In the present study, we aimed to investigate the
effect of IER2 on HCC cell motility and cell-ECM adhesion and
spreading, and to unveil the underlying mechanisms by which IER2
regulated HCC cell motility and adhesion. We demonstrated that
overexpression of IER2 in HCC cells promoted cell adhesion to
fibronectin and motility, whereas knockdown expression of IER2
displayed the opposite effects. Furthermore, we demonstrated a
critical role for IER2 in regulation of HCC cell-ECM adhesion and
motility probably by the transcriptionally promoted integrin β1
(ITGB1) and then activated ITGB1-focal adhesion kinase
(FAK)-Src-paxillin signal pathway.
Materials and methods
Cell lines and culture conditions
Human embryonic kidney (HEK) 293T cells, HCC cell
lines HepG2 and SMMC-7721 were obtained from the Type Culture
Collection of the Chinese Academy of Sciences (Shanghai, China),
and MHCC97H and HCCLM3 cells were obtained from the KeyGen Biotech
(Nanjing, China). All cells were routinely maintained in DMEM
(Gibco-BRL, Grand Island, NY, USA) supplemented with 10% fetal
bovine serum, 100 U/ml penicillin and 100 µg/ml streptomycin, and
cultured at 37°C in a humidified atmosphere of 5%
CO2.
Plasmid construction and lentiviral
production
The full-length of human IER2 cDNA was
amplified from pEZ-Lv105-IER2 vector obtained from GeneCopoeia
(Rockville, MD, USA) by polymerase chain reaction (PCR) using the
primers: forward, 5′-ccggaattctggccaccatggaagtgcagaaagaggcac-3′,
and reverse, 5′-cgcggatcccggaaggccaccacggcccgc-3′, and followed by
cloning into EcoRI and BamHI sites of the pEGFP-N1
vector (Clontech, Mountain View, CA, USA) to generate pEGFP-N1-IER2
vector, which contained a Kozak sequence and ATG to be in frame
with GFP on the vector. Cells were transfected with pEGFP-N1-IER2
or pEGFP-N1 vector using Lipofectamine 3000 according to the
manufacturer's instruction (Invitrogen), and G418-resistant
transfectants were selected. Construct expressing shRNA specific
targeting ITGB1 (shITGB1) was produced by annealing the synthesized
primers, 5′-CCGGCAAGAGAGCTGAAGACTATCTCGAGATAGTCTTCAGCTCTCTTGTTT
TTG-3′, sense and
5′-AATTCAAAAACAAGAGAGCTGAAGACTATCTCGAGATAGTCTTCAGCTCTCTTG-3′,
antisense and cloned into the AgeI and EcoRI sites of
pGV115, and the non-silencing control shRNA vector (NC) were both
obtain from the GeneChem Corporation (Shanghai, China). Cells were
transfected with shITGB1 or with NC using Lipofectamine 3000
according to the manufacturer's instruction. ITGB1 promoter
was cloned from human genomic DNA by PCR using the primers:
forward, 5′-CGGGGTACCGATAGCAGCTTGCCAGTAGC-3′ and reverse,
5′-CCCAAGCTTGGTGGCGGCGGCTCCTCCTCCT-3′, and then inserted into the
pGL3-basic vector (Promega) at KpnI and HindIII
sites. All of the constructs were validated by DNA sequencing.
The production and transduction of the recombinant
lentiviruses encoding IER2 (LV-IER2), shRNA against human IER2
(LV-shR), and indicated control lentiviruses (LV-C and LV-shC) were
generated or acquired as previously described (7,8). After
lentiviral-infection for 48 h, the growth medium was supplemented
with puromycin and maintained at least for two weeks to select
stably transduced cell lines, and cells were screened for IER2
expression by real-time reverse transcription quantitative PCR
(RT-qPCR) and western blotting.
RT-qPCR
Total RNA was extracted and purified from the cells
using the RNA isolator total RNA extraction reagent, and subjected
to reverse transcription using the HiScript First Strand cDNA
synthesis kit according to the supplier's instruction (Vazyme,
Nanjing, China). The real-time PCR and data collection were
subsequently performed as described previously using the
AceQ® qPCR SYBR® Green master mix kit
(Vazyme) (12). Primers used for
the amplification were as follows: IER2 forward,
5′-CCAAAGTCAGCCGCAAACGA-3′ and reverse,
5′-TTTCTTCCAGACGGGCTTTCTTGC-3′; ITGB1 forward,
5′-AATGTAACCAACCGTAGC-3′ and reverse, 5′-CAGGTCCATAAGGTAGTAGA-3′;
integrin α5 (ITGA5) forward, 5′-AGGCAGCTATGGCGTCCCACTGT-3′ and
reverse, 5′-CATCAGAGGTGGCTGGAGGCTTG-3′; and
glyceraldehyde-3-phosphate dehydrogenase (GAPDH) forward,
5′-GCACCGTCAAGGCTGAGAAC-3′ and reverse, 5′-TGGTGAAGACGCCAGTGGA-3′.
The relative expression levels of the indicated mRNA normalized
against GAPDH mRNA were calculated using the 2−ΔΔCT
methods.
Western blot analysis
Western blot analysis was performed following
standard methods. Briefly, cell lysates were prepared, separated
and transferred onto polyvinylidene fluoride membranes (Millipore,
Billerica, MA, USA). The membranes were then blocked with 5% nonfat
dried milk or 3% bovine serum albumin (BSA) for tyrosine
phosphorylation blots and incubated with primary antibodies, and
followed by incubation with horseradish peroxidase (HRP)-linked
secondary antibodies. The antibodies used in the current study
were: mouse polyclonal anti-IER2 (1:1,000; Abcam); rabbit
monoclonal anti-ITGB1, rabbit polyclonal anti-ITGA5, rabbit
monoclonal anti-FAK, rabbit polyclonal anti-pY397FAK, rabbit
polyclonal anti-pY576/Y577FAK, rabbit polyclonal anti-pY925FAK,
rabbit monoclonal anti-Src, rabbit polyclonal anti-pY527Src
(1:1,000), HRP-linked anti-mouse IgG and HRP-linked anti-rabbit IgG
(1:2,000) (Cell Signaling Technology, Danvers, MA, USA); goat
polyclonal anti-pY407FAK, goat polyclonal anti-pY861FAK, goat
polyclonal anti-pS910FAK, mouse monoclonal anti-paxillin and goat
polyclonal anti-pY118paxillin (1:200; Santa Cruz Biotechnology,
Inc., Santa Cruz, CA, USA); and mouse monoclonal anti-GAPDH
(1:1,000) and HRP-linked donkey anti-goat IgG (1:3,000) (KangChen
Biotech, Shanghai, China). Immunoreactive protein bands were
visualized with the Pierce ECL Plus Western Blotting Substrate
(Thermo Fisher Scientific, Rockford, IL, USA). GAPDH was performed
as an internal loading control, and quantification of the band
intensity was performed using ImageJ software (NIH).
Cell viability and proliferation
assay
Cell viability and proliferation was assessed by
using the Cell Counting kit-8 (CCK-8; Obio Technology, Shanghai,
China) assay according to the manufacturer's instruction. In brief,
aliquots (100 µl) of cell suspension of SMMC-7721 or MHCC-97H cells
(5×103 cells/well) were seeded in 96-well culture plates
and cultured in DMEM complete medium for 24, 48 and 72 h. At the
indicated time-points, 10 µl of CCK-8 solution was added to each
well and incubated for another 1 h at 37°C, and the absorbance at
450 nm was measured with a multifunctional microplate reader.
Assays were performed in six wells and repeated five times.
Cell migration and invasion assay
Cell migration and invasion assays were performed as
described previously (12). In
brief, cells were serum-starved overnight and then 200 µl of
1×105 or 2×105 cells in serum free medium
were seeded in uncoated or Matrigel-coated chambers with 8.0 µm
pore sizes (6.5 mm; Costar, Cambridge, MA, USA) of the 24-well
Transwell, respectively. The inserts were then placed into the
lower chambers containing 600 µl of the complete growth medium. The
migrated or invaded cells on the lower surface of the inserts were
fixed and stained. Five random fields at ×100 magnification were
analyzed under a light microscope. Assays were performed in
triplicate from three independent experiments.
Cell-ECM adhesion and cell spreading
assays
Cell-ECM adhesion assays were performed on the
collagen type I, fibronectin and Matrigel (BD Biosciences, Franklin
Lakes, NJ, USA), respectively. For collagen type I or fibronectin
coated, the 96-well culture plates were coated with 10 µg/ml
collagen type I or fibronectin overnight at 4°C and blocked with 1%
BSA in PBS at 37°C for at least 1 h, and then washed with PBS. For
Matrigel coated, Matrigel (200 µg/ml) was added to the 96-well
culture plates, incubated at 37°C for at least 1 h, and blocked
with 1% BSA. For cell-ECM adhesion assays, aliquots (100 µl) of
cell suspension at a density of 2×106 cells/ml were
seeded to each well of the matrix coated 96-well culture plates and
allowed to adhere to the matrix for 45 min at 37°C. Unattached
cells were gently removed and adhesive cells were quantified by
using the CCK-8 assay according to the supplier's instruction.
Assays were performed in six wells from five independent
experiments. For cell spreading assays, cells (2×105
cells/well) were seeded to each well of the fibronectin coated
coverslips in 24-well culture plates prepared as above and
incubated at 37°C for the indicated time. The cells were then fixed
immediately with 4% paraformaldehyde and the spreading cells were
captured by light microscopy equipped with a digital image
capturing system. Spreading cells were defined as cells adherent
with protrusions or lamellipodia (fried egg shape), whereas
unspreading cells were identified by their round and adherent with
less projections appearance. Spreading was quantified from 10
randomly selected fields from three independent experiments.
Chromatin immunoprecipitation (ChIP)
assay
The ChIP assay was performed in the cells stably
transfected with pEGFP-N1-IER2 or pEGFP-N1 vectors using the
SimpleChIP® Plus Enzymatic Chromatin IP kit (Cell
Signaling Technology) according to the supplier's instruction. In
brief, cells were grown to confluence in culture dishes and fixed
in formaldehyde (1%), scraped from the dishes and lysed, and the
homogenized chromatin was sheared by micrococcal nuclease for 20
min at 37°C and sonicated with several pulses to break the nuclear
membrane. The GFP antibody (Invitrogen), positive control histone
H3 antibody, or negative control normal rabbit IgG was used to
precipitate the precleared cross-linked chromatin at 4°C overnight
with rotation, and followed by adding protein G beads and
incubation for 4 h at 4°C with rotation. After washing, the
immunoprecipitated chromatin was eluted from the antibody/protein G
beads and purified by reversal of cross-links, removal of RNA with
RNase A, and treatment with proteinase K. The immunoprecipitated
DNA fragments were performed as the template for PCR using the
human ITGB1 promoter-specific primers forward,
5′-TCTTAGACACAGGCAAGCACA-3′ and reverse,
5′-CCTGAGTCCCGAGGCAAATC-3′. The PCR products were run on a 2 %
agarose gel and visualized with ethidium bromide staining.
Luciferase reporter gene assay
For luciferase reporter gene assay, the luciferase
reporter constructs (200 ng) and pTK-Renilla vectors (100
ng) were transiently transfected into the lentiviral-transduced
cells in 24-well culture plates using Lipofectamine 3000. After
incubation for 48 h, luciferase activities were determined using
the luciferase reporter assay system (Promega), and normalized to
Renilla luciferase activities.
Statistical analysis
Data are shown as the mean ± standard deviation (SD)
and analyzed by the Student's t-test. The statistical significance
was determined by p<0.05.
Results
IER2 promoted HCC cell migration and
invasion
To understand the contribution of IER2 expression to
the biological functions in HCC cells, the lentivirus-mediated IER2
overexpression or knockdown in the low metastatic potential
SMMC7721 cells and in the high metastatic potential MHCC97H cells
were initially established. Overexpression and knockdown of IER2 in
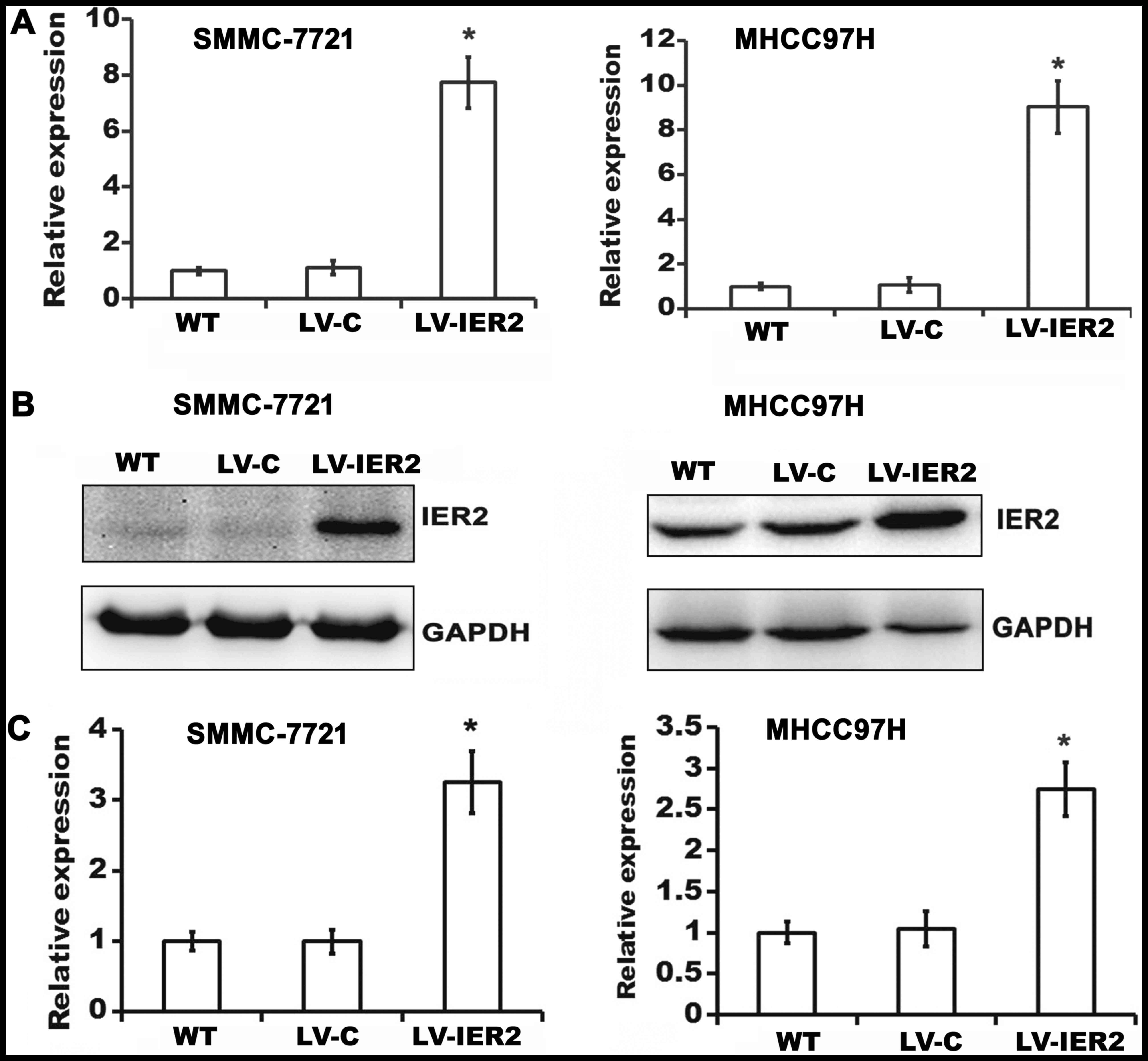
SMMC-7721 cells and in MHCC97H cells were confirmed by RT-qPCR
(Figs. 1A and 2A) and western blot analysis (Figs. 1B and C and 2B and C). The data suggested that the
indicated lentiviruses have been successfully transduced into the
SMMC-7721 and MHCC97H cells, and IER2 overexpression was shown in
LV-IER2-infected cells, and efficient knockdown of IER2 was shown
in LV-shR-infected cells, thus, the indicated lentiviruses could be
used in the following experiments.
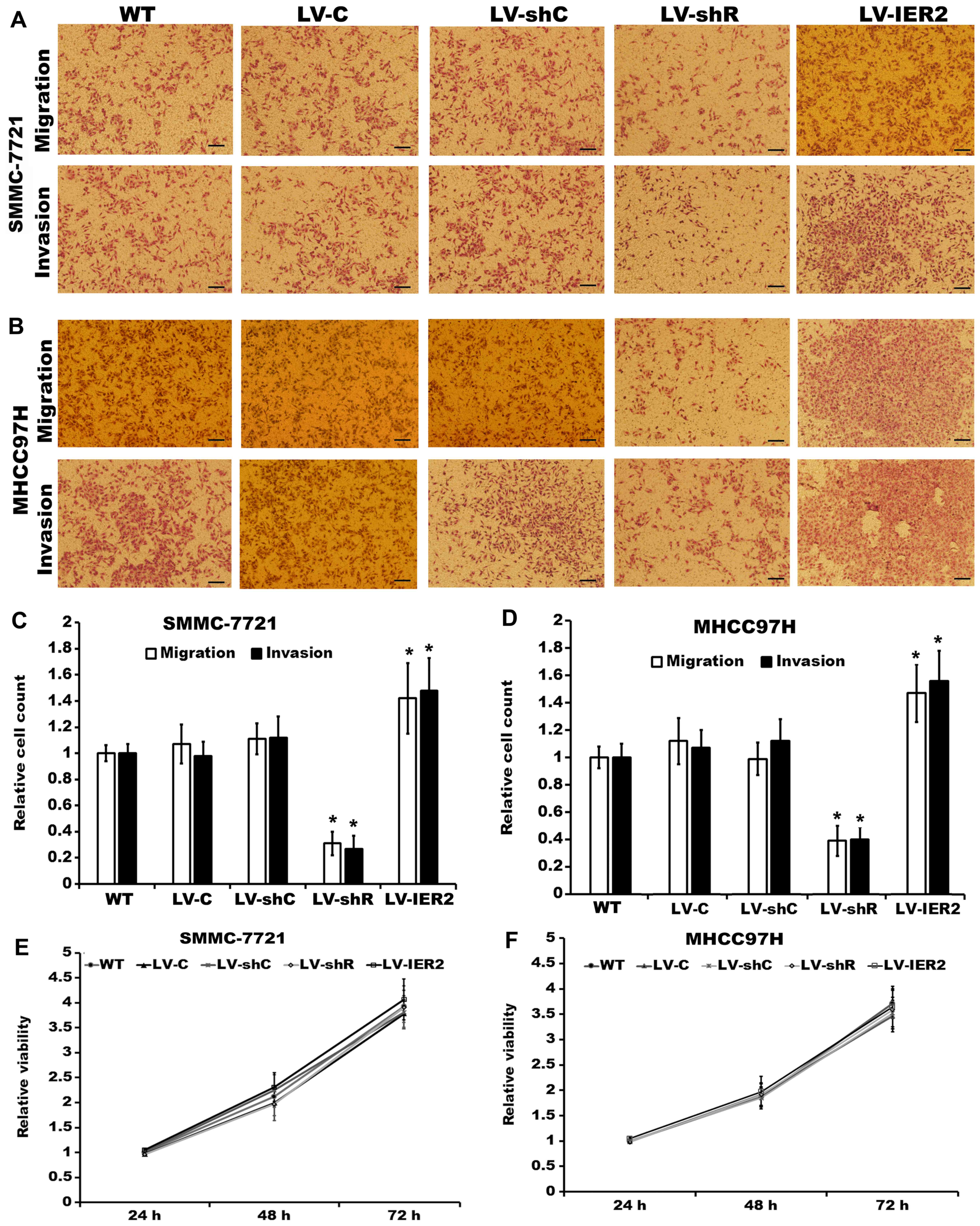
We employed lentivirus-mediated IER2 overexpression
or knockdown in the SMMC7721 cells and MHCC97H cells, respectively,
and performed Transwell cell migration and invasion assays.
Compared with the corresponding empty vector-transduced cells (LV-C
and LV-shC) and the non-transduced cells [wild-type (WT)], both
SMMC-7721 and MHCC97H cells stably transduced with LV-IER2 showed
significant increase of the cell migratory and invasiveness
capacity, whereas silencing of IER2 obviously attenuated the
motility of SMMC-7721 and MHCC97H cells (Fig. 3A-D). No significant differences in
cell motility were observed among the empty vector-infected cells
and the WT. In addition, the alteration in cell motility did not
appear to be due to affected cell viability and proliferation.
Under these conditions, knockdown and the ectopic expression of
IER2 had no obvious effects on cell viability and proliferation of
SMMC-7721 and MHCC97H cells in vitro (Fig. 3E and F). These observations
supported the notion that IER2 played an important role in HCC cell
migration and invasion.
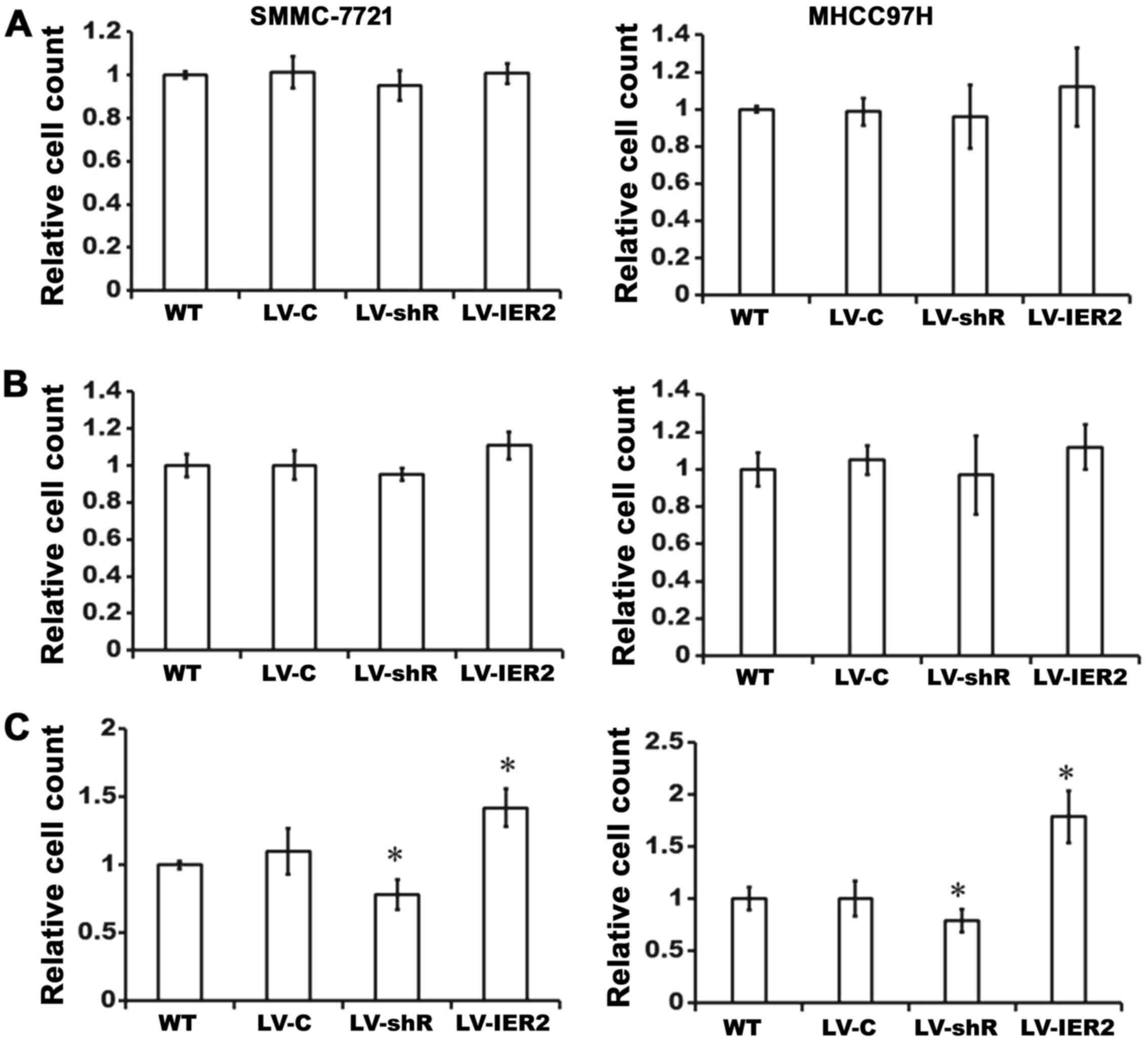
IER2 regulates cell-ECM adhesion and
spreading
Considering that cell adhesion and spreading onto
ECM are crucial steps in cellular migration and invasion (13), we then investigated whether IER2
positively regulates HCC cell-ECM adhesion and spreading.
Lentiviral-transduced SMMC-7721 or MHCC97H cells were seeded on
collagen type I-, Matrigel- or fibronectin-coated 96-well culture
plates, respectively, and incubated at 37°C for 45 min. As shown in
Fig. 4, compared with those in the
empty vector-transduced cells and WT, LV-IER2 infection
significantly promoted SMMC-7721 or MHCC97H cell adhesion onto the
fibronectin, while LV-shR infection decreased the cell adhesion
onto the fibronectin (Fig. 4C).
Interestingly, there were no significant alteration in the cell
adhesion onto the collagen type I or Matrigel either in IER2
overexpressing cells or IER2 knockdown cells (Fig. 4A and B). No significant difference
was observed either in the LV-C-infected or the LV-shC-infected
cells (data not shown). These data demonstrated that IER2 is an
important regulator of HCC cell adhesion onto the fibronectin, but
not collagen type I or Matrigel, and promotion of the cell motility
by IER2 overexpression may be the consequence of the increase
cell-fibronectin adhesion, while the reduction in the cell motility
induced by IER2 knockdown is likely the consequence of the
decreasing cell-fibronectin adhesion.
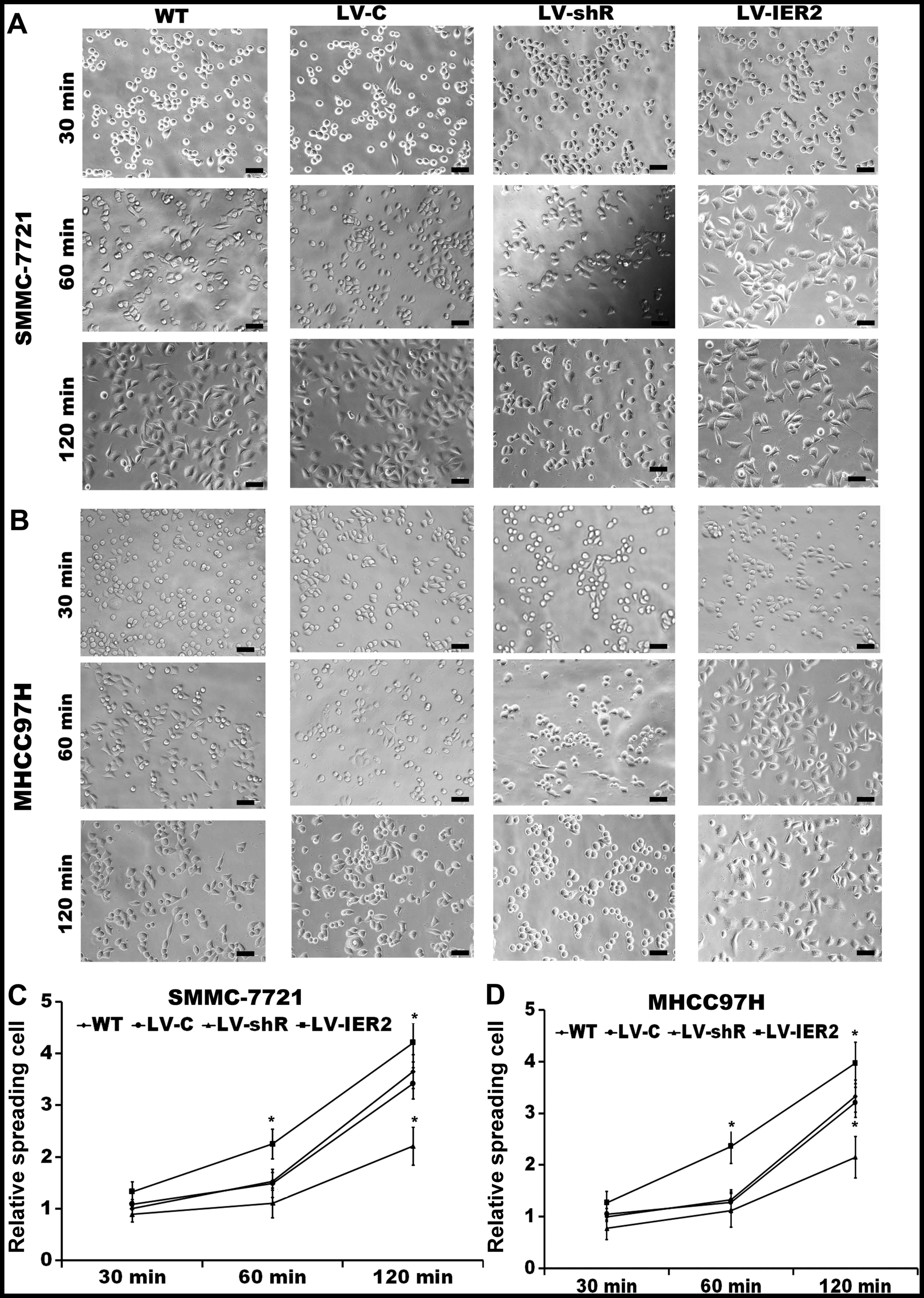
Since cell spreading is the early phase of cell
migration and invasion (13,14),
and IER2 expression promoted cell adhesion onto the fibronectin
(but not collagen type I or Matrigel) as described above, we
further examined whether IER2 positively regulated the cell
spreading on the fibronectin. With increasing the time of
incubation, both SMMC-7721 and MHCC97H cells showed significant
increase of the cell spreading (Fig.
5). Specifically, when compared to empty vector-transduced
cells or WT, overexpression of IER2 significantly increased the
ability of cells to spread on the fibronectin coated surface
(Fig. 5C and D), and the
LV-IER2-transduced cells displayed increased cell spreading surface
area and showed lamellopodia around their peripheral edges
(Fig. 5A and B). On the contrary,
IER2 depletion obviously decreased cell spreading on the
fibronectin, and these IER2 silencing cells showed poor spreading
and exhibited a non-polarized, spherical shape, and few lamellipodi
protrusions (Fig. 5A and B). No
significant difference was found either in the LV-C-transduced or
the LV-shC-transduced cells (data not shown). Taken together, these
results suggested that IER2 is required for effective spreading and
adhesion of HCC cells to fibronectin, and that the increased cell
adhesion and spreading of IER2 expressing cells, perhaps account
for the promotion of IER2 on migration and invasion.
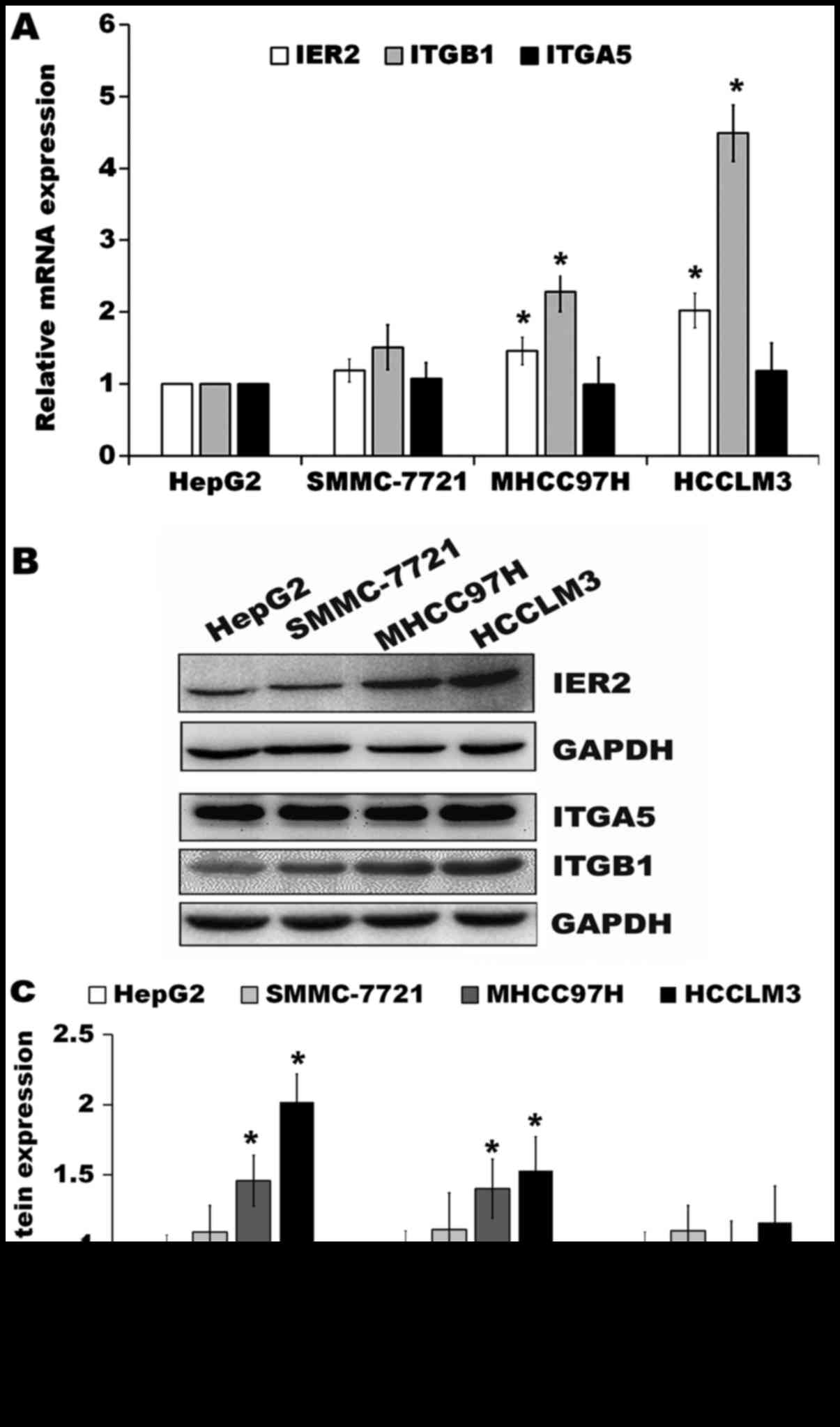
IER2 promotes ITGB1 expression in HCC
cells
ITGA5 and ITGB1 form heterodimers to mediate cell
adhesion onto the fibronectin (15)
and play very important roles in cell motility (16–18).
Since IER2 expression positively regulated cell motility, adhesion
and spreading onto the fibronectin of the SMMC-7721 and MHCC97H
cells (Figs. 3–5), we then tested whether IER2 expression
was correlated with the metastatic potential of HCC cell lines and
with ITGA5 or ITGB1 expression both in mRNA and protein levels in
HCC cells. Data from RT-qPCR and western blot analysis demonstrated
that the HCC cell lines, HepG2, SMMC-7721, MHCC97H and HCCLM3
cells, four HCC cell lines with increasing spontaneous metastatic
potential (19–21), were shown to express IER2 with
relative low expression of IER2 in HepG2 and SMMC-7721 cells, which
have low metastatic potential, and abundant expression of IER2 in
MHCC97H and HCCLM3 cells, which have high metastatic potential
(Fig. 6), indicating that IER2
expression was positively correlated with the metastatic potential
of HCC cell lines. Moreover, the results showed that IER2
expression was positively correlated with ITGB1 (but not ITGA5)
expression in these cell lines with increasing spontaneous
metastatic potential (Fig. 6).
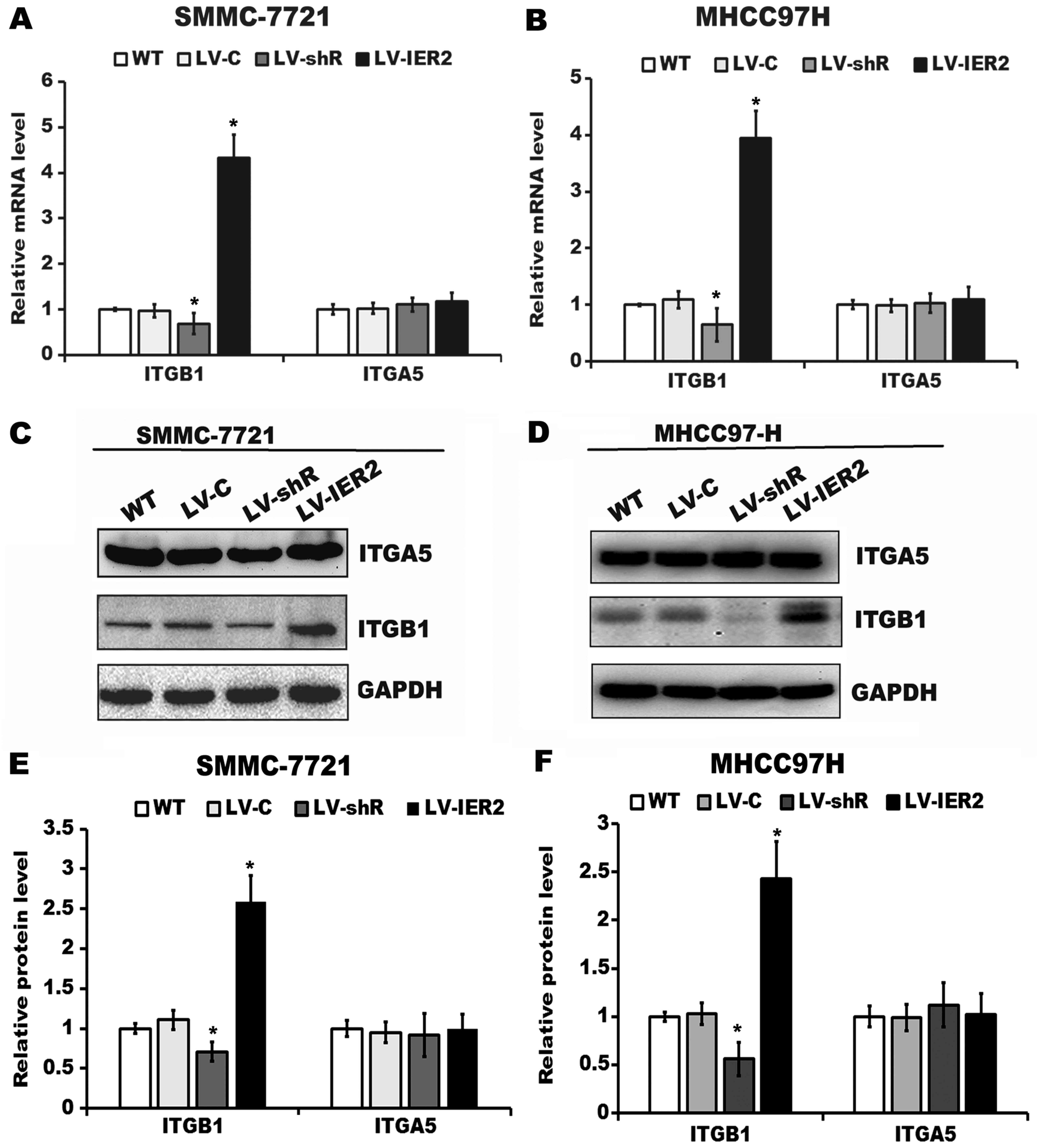
We further explored whether IER2 expression may
regulate ITGA5 or ITGB1 expression both in mRNA and protein levels.
Our results indicated that abundant expression of ITGA5 was found
in HCC cell lines (Figs. 6B and
7C and D), whereas IER2
overexpression or knockdown resulted in significant upregulation or
downregulation of ITGB1, but not ITGA5 either in mRNA or protein
levels in SMMC-7721 and MHCC97H cells (Fig. 7A-F), suggesting that IER2 expression
levels positively correlate with ITGB1 expression in SMMC-7721 and
MHCC97H cells. To verify whether the observed positive correlation
of IER2 with the ITGB1 expression is the consequence of the
interaction of IER2 with the ITGB1 promoter, the luciferase
reporter gene assay and ChIP assay were performed. After transient
transfection of the luciferase reporter constructs and the internal
control vectors into the indicated lentiviral-transduced SMMC-7721
and MHCC97H cells, overexpression of IER2 markedly upregulated the
luciferase activity of the ITGB1 promoter, while knockdown of IER2
significantly decreased the luciferase activity of the ITGB1
promoter in comparison with those in the WT and empty
vector-transduced cells (Fig. 7G and
H). Moreover, results from ChIP assay, which was performed in
SMMC-7721 and MHCC97H cells stably transfected with pEGFP-N1-IER2
or pEGFP-N1 vectors, showed that the amplified product of 146 bp
was enriched in the pEGFP-N1-IER2 expressing cells relative to that
in the negative control IgG and the input (Fig. 7I and J), indicating that IER2 does
interact directly with the ITGB1 promoter. Collectively, these data
demonstrated that IER2 may regulate expression of the ITGB1 in HCC
cells by direct interaction with the ITGB1 promoter, and indicated
that IER2 promoted the cell-fibronectin adhesion and motility of
the HCC cells presumably by activating ITGB1 signaling.
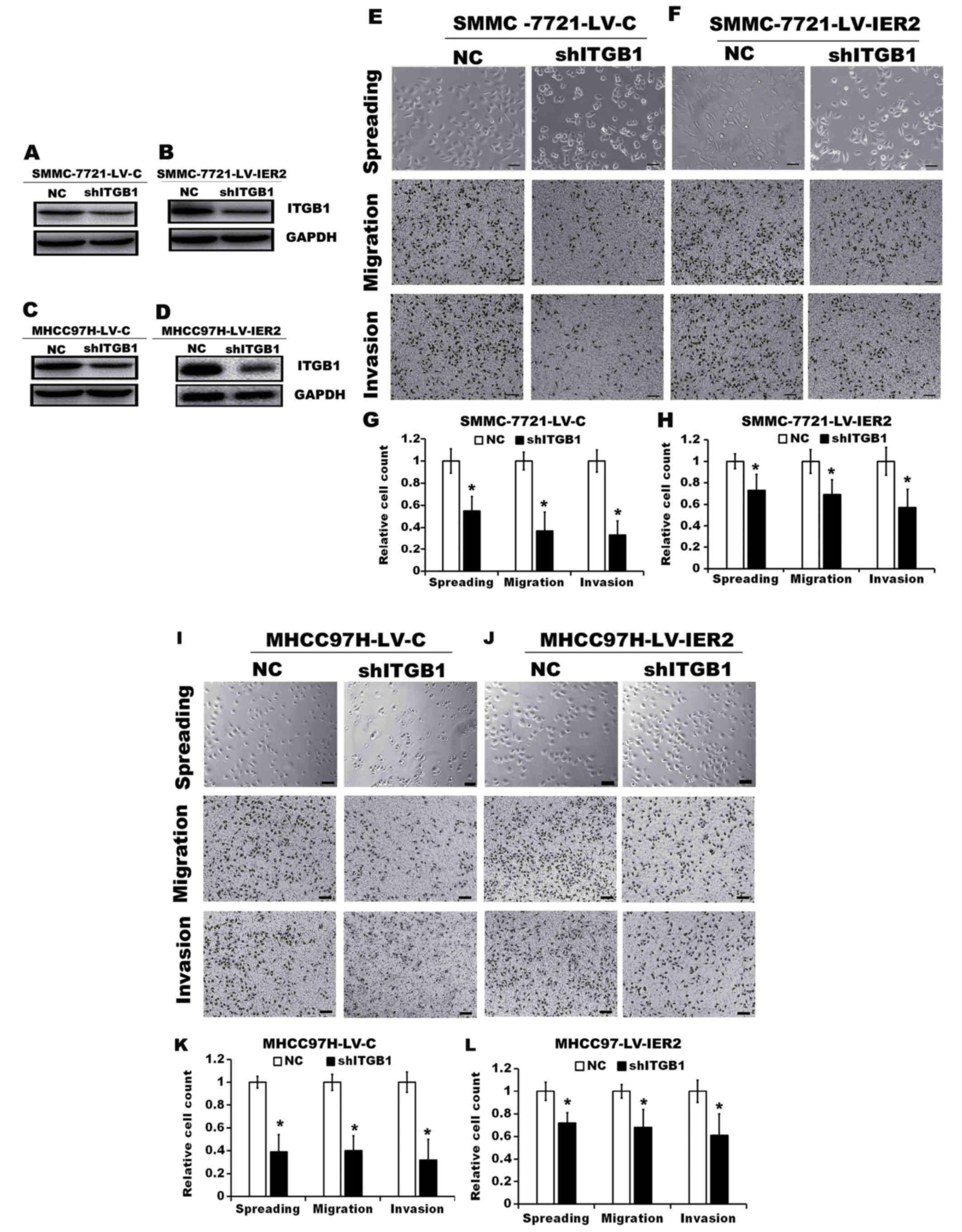
Effect of IER2 on the ITGB1-mediated
signaling pathway
In present study we observed that IER2 promoted cell
adhesion and spreading on the fibronectin (Figs. 4 and 5) and regulated expression of the ITGB1
(Fig. 7) in HCC cell lines,
suggesting that IER2-mediated ITGB1 expression may account for the
promotion of cell motility by activating ITGB1-mediated signaling
pathway. To assess the requirement of ITGB1 in IER2-promoted cell
spreading and motility, ITGB1 was knocked down in both SMMC-7721
and MHCC97H cells stably transduced with LV-C or LV-IER2, and these
cells were subjected to cell spreading, Transwell migration and
invasion assays. As shown in Fig.
8, compared with those in the cells transfected with
non-targeting shRNA (NC), ITGB1 knockdown in stably LV-C-transduced
SMMC-7721 (SMMC-7721-LV-C) and MHCC97H (MHCC97H-LV-C) cells or the
LV-IER2- transduced SMMC-7721 (SMMC-7721-LV-IER2) and MHCC97H
(MHCC97H-LV-IER2) cells significantly reduced cell spreading,
migration and invasion, suggesting that the elevated ITGB1 resulted
from IER2 overexpression in the SMMC-7721 or MHCC97H cells plays an
important role in promoting cell motility.
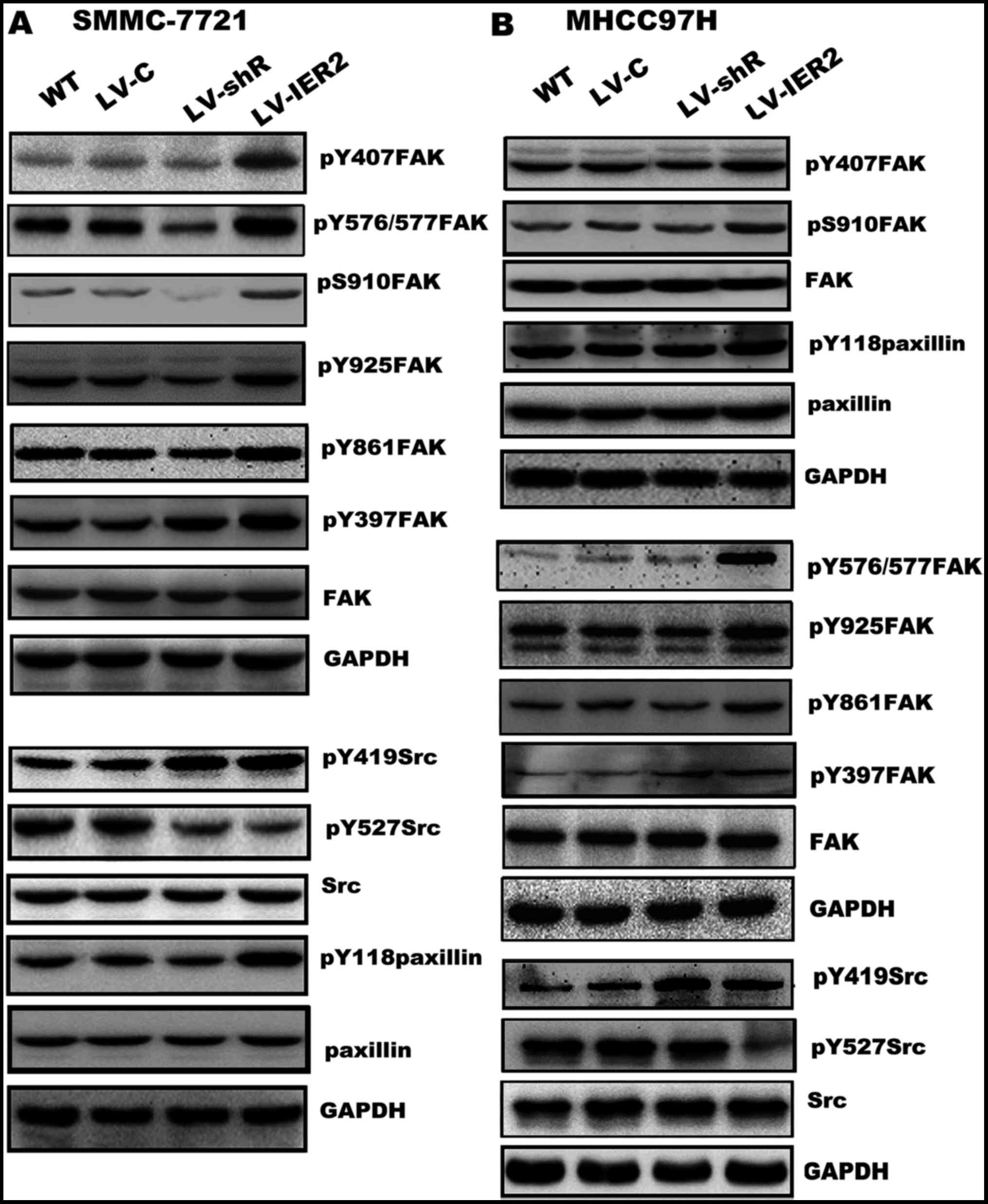
It has been well characterized for most cell types
that integrins link ECM to actin filaments activating the
integrin-mediated downstream signaling molecules, including FAK,
Src and paxillin, thereby regulating the cell-ECM adhesion,
spreading and cell motility (16–18,22–24).
We next performed western blot assay to explore whether IER2
expression regulated cell motility by activating FAK-Src-paxillin
signaling pathway. As shown in Fig.
9, compared with those in the WT and empty vector-transduced
cells, overexpression of IER2 significantly increased
phosphorylation of FAK at Y397 (pY397FAK), Y407 (pY407FAK),
Y576/Y577 (pY576/Y577FAK), Y861 (pY861 FAK), Y925 (pY925FAK) and
S910 (pS910FAK), and phosphorylation of Src at Y419 (pY419Src) and
paxillin at Y118 (pY118paxillin), and decreased phosphorylation of
Src at Y527 (pY527Src). Interestingly, knockdown of IER2 showed a
slight, but not significant alteration of phosphorylation of FAK
and paxillin, whereas, obvious decrease of pY527Src and increase of
pY419Src were shown in IER2 silencing cells, suggesting that IER2
activated ITGB1-mediated signal pathway is likely a part of a
mechanism that participates in modulation of cell motility and
adhesion of HCC cells. No significant differences were found either
in the LV-C-transduced or the LV-shC-transduced cells (data not
shown). Taken together, these results indicated that IER2-mediated
ITGB1 expression is partially responsible for IER2-promoted
cell-fibronectin adhesion and motility of the HCC cells, providing
a novel molecular mechanism for IER2 promoting cell adhesion and
motility.
Discussion
Previously, a study showed that IER2 plays a role in
tumor cell motility and metastasis (6), and we also found that IER2 promotes
the motility of SMMC-7721 and HepG2 cells (8). However, the detailed roles and
underlying molecular mechanisms in HCC remained unknown. In this
study, we investigated the effects of IER2 on the migration,
invasion, cell-ECM adhesion and spreading of two HCC cell lines
SMMC-7721 with low metastatic potential and MHCC97H with high
metastatic potential, and identified IER2 as a regulator of
cell-fibronectin adhesion, spreading and motility of these two cell
lines. We further defined a new pathway that involves the cell-ECM
adhesion molecule ITGB1, and demonstrated the novel IER2-ITGB1
signaling critically contributed to the HCC cell-ECM adhesion and
motility.
In this study, we first found that IER2
overexpression obviously induced cell migration and invasion, while
silencing of IER2 decreased the motility of both SMMC-7721 and
MHCC97H cells, suggesting that IER2 may function as an important
regulator in HCC cell migration and invasion. Since cell motility
requires cell-ECM adhesion, spreading and then invading the ECM
(13–15,25) we
next evaluated whether IER2 induced HCC cell migration and invasion
via its regulation for the cell-ECM adhesion and spreading. We
demonstrated that IER2 overexpression promoted SMMC-7721 or MHCC97H
cell adhesion and spreading onto the fibronectin (but not collagen
type I or Matrigel), whereas IER2 knockdown repressed cell adhesion
and spreading onto the fibronectin. Taken together, these findings
indicated that IER2 is an important regulator of cell spreading and
adhesion onto the fibronectin, and promotion of the cell motility
by IER2 overexpression may be due to the increase in
cell-fibronectin adhesion and spreading, whereas the decrease in
the cell motility caused by IER2 depression is probably attributed
to the decreasing cell-fibronectin adhesion and spreading, and the
endogenous IER2 expression is required for cell motility and
adhesion.
Integrins are the major ECM receptors that consist
of α and β subunits and play very important roles in cell motility
and tumor progression, especially in tumor invasion and metastasis
(16–18,26).
Among integrins, the heterodimers of ITGA5 and ITGB1, receptors for
fibronectin, are thought to mediate tumor cell adhesion to
fibronectin and closely correlate with cancer progression and
metastasis (18). In the present
study, we demonstrated that IER2 expression was positively
correlated with the metastatic potential and associated with ITGB1
expression levels in HCC cell lines. These results prompted us to
examine whether IER2 may regulate the expression of ITGA5 and
ITGB1. We showed that IER2 expression enhanced ITGB1 expression
both in mRNA and protein levels in SMMC-7721 and MHCC97H cells,
while knockdown of IER2 reduced ITGB1 expression. Abundant
expression of ITGA5 was observed in HCC cell lines, but neither
IER2 overexpression nor knockdown altered the expression of ITGA5
either in mRNA or protein levels in SMMC-7721 and MHCC97H cells. We
also demonstrated that IER2 may regulate expression of ITGB1 in HCC
cell lines by functioning as a potential transcriptional factor or
transcriptional co-activator for ITGB1. Furthermore, we silenced
ITGB1 in both SMMC-7721 and MHCC97H cells stably transduced with
LV-C or LV-IER2, and found that the increase of cell migration,
invasion, and spreading on fibronectin in IER2 overexpressing cells
was prevented by knockdown of ITGB1. Collectively, these findings
suggested that IER2-induced cell-fibronectin adhesion, spreading
and motility of the HCC cells may be mediated, at least in part, by
promotion of ITGB1.
It is well documented that ITGB1 is the classic
fibronectin receptor and mediates cell adhesion onto the ECM and
confers higher metastatic capacity to a number of cancer cells
(27–29). Upon integrin-mediated signaling
activation, the heterodimers of ITGA5 and ITGB1 link fibronectin to
induce phosphorylation of the FAK at Y397 and formation of the
active FAK/Src complex, followed by further phosphorylation at
other sites on the FAK and other downstream signaling molecules
including paxillin, and subsequently modulating cell motility and
cell-fibronectin adhesion (24,30,31).
Therefore, we further examined the role of IER2 in the
integrin-mediated signaling pathway. We found that the FAK
phosphorylation (at Y397, Y407, Y576/Y577, Y861, Y925 and S910),
Src phosphorylation (at Y419), and paxillin phosphorylation were
significantly upregulated, and the Src phosphorylation at Y527 [the
inhibitory site for Src activation (32)] was downregulated in IER2
overexpressing HCC cells, suggesting that IER2 overexpression
promoted HCC cell-fibronectin adhesion, spreading and motility by
activating FAK, Src and paxillin via ITGB1. Unexpectedly, we did
not find obvious opposite effects in IER2 knockdown cells. We found
that silencing of IER2 resulted in only a slight, but no
significant alteration of phosphorylation of FAK and paxillin, but
also significant decrease of pY527Src and increase of pY419Src,
suggesting that IER2 may have some uncharacterized functions or
potential partners to regulate HCC cell motility and adhesion, and
that IER2 activated ITGB1-mediated signal pathway is likely a part
of a mechanism that participates in modulation of cell motility and
adhesion of HCC cells. Collectively, these findings indicated that
IER2-mediated ITGB1 expression and activation of
ITGB1-FAK-Src-paxillin signal pathway was likely, at least in part,
responsible for IER2-promoted cell-ECM adhesion and motility of the
HCC cells, and studies to identify potential partners and the
detail mechanisms of IER2 in regulating HCC cell-ECM adhesion and
motility are under way.
In conclusion, we demonstrated a novel pathway
modulating HCC cell motility and cell-ECM adhesion by IER2, and
identified that IER2 expression is positively correlated with the
metastatic potential of HCC cell lines, and that IER2 promoted HCC
cell motility and the cell-fibronectin adhesion as the result of
the activation of the ITGB1-FAK-Src-paxillin signal pathway.
Further study of the underlying mechanism as to how IER2 depletion
coordinates the ITGB1-FAK-Src-paxillin signaling is ongoing,
however, the findings presented here may reinforce a role for IER2
as a novel molecular target for HCC.
Acknowledgements
This study was supported by the National Nature
Science Foundation of China (no. 81172278).
References
|
1
|
Shimizu N, Ohta M, Fujiwara C, Sagara J,
Mochizuki N, Oda T and Utiyama H: Expression of a novel immediate
early gene during 12-O-tetradecanoylphorbol-13-acetate-induced
macrophagic differentiation of HL-60 cells. J Biol Chem.
266:12157–12161. 1991.PubMed/NCBI
|
|
2
|
Zeng F, Hon CC, Sit WH, Chow KY, Hui RK,
Law IK, Ng VW, Yang XT, Leung FC and Wan JM: Molecular
characterization of Coriolus versicolor PSP-induced apoptosis in
human promyelotic leukemic HL-60 cells using cDNA microarray. Int J
Oncol. 27:513–523. 2005.PubMed/NCBI
|
|
3
|
Takaya T, Kasatani K, Noguchi S and Nikawa
J: Functional analyses of immediate early gene ETR101 expressed in
yeast. Biosci Biotechnol Biochem. 73:1653–1660. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Chen L, Ma S, Li B, Fink T, Zachar V,
Takahashi M, Cuttichia J, Tsui LC, Ebbesen P and Liu X:
Transcriptional activation of immediate-early gene ETR101 by human
T-cell leukaemia virus type I Tax. J Gen Virol. 84:3203–3214. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hong SK and Dawid IB: FGF-dependent
left-right asymmetry patterning in zebrafish is mediated by Ier2
and Fibp1. Proc Natl Acad Sci USA. 106:2230–2235. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Neeb A, Wallbaum S, Novac N,
Dukovic-Schulze S, Scholl I, Schreiber C, Schlag P, Moll J, Stein U
and Sleeman JP: The immediate early gene Ier2 promotes tumor cell
motility and metastasis, and predicts poor survival of colorectal
cancer patients. Oncogene. 31:3796–3806. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wu W, Zhang X, Lv H, Liao Y, Zhang W,
Cheng H, Deng Z, Shen J, Yuan Q, Zhang Y, et al: Identification of
immediate early response protein 2 as a regulator of angiogenesis
through the modulation of endothelial cell motility and adhesion.
Int J Mol Med. 36:1104–1110. 2015.PubMed/NCBI
|
|
8
|
Wu W, Zhang X, Liao Y, Zhang W, Cheng H,
Deng Z, Shen J, Yuan Q, Zhang Y and Shen W: miR-30c negatively
regulates the migration and invasion by targeting the immediate
early response protein 2 in SMMC-7721 and HepG2 cells. Am J Cancer
Res. 5:1435–1446. 2015.PubMed/NCBI
|
|
9
|
Yan M, Li H, Zhu M, Zhao F, Zhang L, Chen
T, Jiang G, Xie H, Cui Y, Yao M, et al: G protein-coupled receptor
87 (GPR87) promotes the growth and metastasis of CD133+
cancer stem-like cells in hepatocellular carcinoma. PLoS One.
8:e610562013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Woodhouse EC, Chuaqui RF and Liotta LA:
General mechanisms of metastasis. Cancer 80 (Suppl). 1529–1537.
1997. View Article : Google Scholar
|
|
11
|
Geiger TR and Peeper DS: Metastasis
mechanisms. Biochim Biophys Acta. 1796:293–308. 2009.PubMed/NCBI
|
|
12
|
Wu W, Zhang X, Qin H, Peng W, Xue Q, Lv H,
Zhang H, Qiu Y, Cheng H, Zhang Y, et al: Modulation of tumor cell
migration, invasion and cell-matrix adhesion by human monopolar
spindle-one-binder 2. Oncol Rep. 33:2495–2503. 2015.PubMed/NCBI
|
|
13
|
Khalili AA and Ahmad MR: A review of cell
adhesion studies for biomedical and biological applications. Int J
Mol Sci. 16:18149–18184. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ishikawa T and Kramer RH: Sdc1 negatively
modulates carcinoma cell motility and invasion. Exp Cell Res.
316:951–965. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Desgrosellier JS and Cheresh DA: Integrins
in cancer: Biological implications and therapeutic opportunities.
Nat Rev Cancer. 10:9–22. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
16
|
Margadant C, Monsuur HN, Norman JC and
Sonnenberg A: Mechanisms of integrin activation and trafficking.
Curr Opin Cell Biol. 23:607–614. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Keely S, Glover LE, MacManus CF, Campbell
EL, Scully MM, Furuta GT and Colgan SP: Selective induction of
integrin beta1 by hypoxia-inducible factor: Implications for wound
healing. FASEB J. 23:1338–1346. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Qin L, Chen X, Wu Y, Feng Z, He T, Wang L,
Liao L and Xu J: Steroid receptor coactivator-1 upregulates
integrin α5 expression to promote breast cancer cell
adhesion and migration. Cancer Res. 71:1742–1751. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhou L, Wang DS, Li QJ, Sun W, Zhang Y and
Dou KF: The downregulation of Notch1 inhibits the invasion and
migration of hepatocellular carcinoma cells by inactivating the
cyclooxygenase-2/Snail/E-cadherin pathway in vitro. Dig Dis Sci.
58:1016–1025. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang C, Jin G, Jin H, Wang N, Luo Q, Zhang
Y, Gao D, Jiang K, Gu D, Shen Q, et al: Clusterin facilitates
metastasis by EIF3I/Akt/MMP13 signaling in hepatocellular
carcinoma. Oncotarget. 6:2903–2916. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang C, Jiang K, Kang X, Gao D, Sun C, Li
Y, Sun L, Zhang S, Liu X, Wu W, et al: Tumor-derived secretory
clusterin induces epithelial-mesenchymal transition and facilitates
hepatocellular carcinoma metastasis. Int J Biochem Cell Biol.
44:2308–2320. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Fraley SI, Feng Y, Krishnamurthy R, Kim
DH, Celedon A, Longmore GD and Wirtz D: A distinctive role for
focal adhesion proteins in three-dimensional cell motility. Nat
Cell Biol. 12:598–604. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
23
|
van Nimwegen MJ and van de Water B: Focal
adhesion kinase: A potential target in cancer therapy. Biochem
Pharmacol. 73:597–609. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Mitra SK and Schlaepfer DD:
Integrin-regulated FAK-Src signaling in normal and cancer cells.
Curr Opin Cell Biol. 18:516–523. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Webb DJ, Donais K, Whitmore LA, Thomas SM,
Turner CE, Parsons JT and Horwitz AF: FAK-Src signalling through
paxillin, ERK and MLCK regulates adhesion disassembly. Nat Cell
Biol. 6:154–161. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hood JD and Cheresh DA: Role of integrins
in cell invasion and migration. Nat Rev Cancer. 2:91–100. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Xu JK, Chen HJ, Li XD, Huang ZL, Xu H,
Yang HL and Hu J: Optimal intensity shock wave promotes the
adhesion and migration of rat osteoblasts via integrin β1-mediated
expression of phosphorylated focal adhesion kinase. J Biol Chem.
287:26200–26212. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Moursi AM, Globus RK and Damsky CH:
Interactions between integrin receptors and fibronectin are
required for calvarial osteoblast differentiation in vitro. J Cell
Sci. 110:2187–2196. 1997.PubMed/NCBI
|
|
29
|
Broustas CG, Zhu A and Lieberman HB: Rad9
protein contributes to prostate tumor progression by promoting cell
migration and anoikis resistance. J Biol Chem. 287:41324–41333.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Carvajal-Gonzalez JM, Mulero-Navarro S,
Roman AC, Sauzeau V, Merino JM, Bustelo XR and Fernandez-Salguero
PM: The dioxin receptor regulates the constitutive expression of
the vav3 proto-oncogene and modulates cell shape and adhesion. Mol
Biol Cell. 20:1715–1727. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mulero-Navarro S, Pozo-Guisado E,
Pérez-Mancera PA, Alvarez-Barrientos A, Catalina-Fernández I,
Hernández-Nieto E, Sáenz-Santamaria J, Martínez N, Rojas JM,
Sánchez-García I, et al: Immortalized mouse mammary fibroblasts
lacking dioxin receptor have impaired tumorigenicity in a
subcutaneous mouse xenograft model. J Biol Chem. 280:28731–28741.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Crosara-Alberto DP, Inoue RY and Costa CR:
FAK signalling mediates NF-kappaB activation by mechanical stress
in cardiac myocytes. Clin Chim Acta. 403:81–86. 2009. View Article : Google Scholar : PubMed/NCBI
|