Introduction
Lung cancer is one of the leading causes of
cancer-related death in the world (1). Non-small cell lung cancer (NSCLC) is a
major type of lung cancer accounting for nearly 85% of all lung
cancer cases. Despite great efforts in diagnostic procedures and
therapeutic options, it is often diagnosed at an advanced stage and
the overall 5-year survival rate is only approximately 15%
(2). For treatment of lung cancer,
platinum-based combination chemotherapy is still the standard
first-line treatment for NSCLC, particularly for tumors without
EGFR mutation or ALK translocation (3). However, chemoresistance is a major
obstacle in the treatment of lung cancer. Therefore, the discovery
of new biomarkers is critical not only for early detection of the
disease but also for the prediction of chemotherapeutic
efficacy.
A Parkinson's disease-associated gene, PINK1
[phosphatase and tensin homolog deleted on chromosome 10
(PTEN)-induced kinase 1] was initially identified in HeLa cells as
a gene upregulated by overexpression of the main tumor suppressor,
PTEN (4). Loss of function in PINK1
could cause autosomal recessive forms of Parkinson's disease
(5). The PTEN-induced putative
kinase 1 (PINK1) gene encodes a 581-amino acid protein with a
mitochondrial targeting sequence, a highly conserved
serine/threonine protein kinase domain and a regulatory C-terminal
sequence (6). Research during the
past decade revealed that PINK1 plays an important role in
pro-survival, anti-apoptosis and cytoprotection (7–9),
mechanistically via proteasomal and autophagic pathways,
PI3-kinase/Akt, NF-κB pathway and calcium-dependent signaling
(10–13). Substantial studies have indicated a
potential role for PINK1 in cancer cell biology, including cell
survival, mitochondrial homeostasis, stress resistance and the cell
cycle (14–16). As a tumor oncogene, PINK1 is
critical for activation of a well-known oncogenic pathway,
insulin-like growth factor-1-dependent Akt signaling (17).
Moreover, deletion of PINK1 increases sensitivity of
cancer cells to paclitaxel (18).
Although PINK1 dysfunction has been related with the progression of
numerous cancers such as breast cancer, colorectal cancer and
endometrial carcinoma, the precise role of PINK1 in the development
of NSCLC remains unknown. However, numerous studies demonstrate
that NF-κB signaling is intimately linked with cellular adhesion,
migration, invasion, and chemoresistance of NSCLC (19,20).
Furthermore, it has been reported that PINK1 could specifically
bind to TRAF6, which results in auto-ubiquitination of TRAF6 and
activation of NF-κB pathway (21).
Therefore, we assume that PINK1 might play a role in NSCLC
development through NF-κB signaling.
In the present study, we determined the expression
of PINK1 in NSCLC tissues and NSCLC cell lines. In addition, we
explored the correlation between PINK1 expression and various
clinicopathological features as well as its prognostic value for
NSCLC patients. Furthermore, we explored the role of PINK1
expression in regulating cell cycle progression, cell
proliferation, and drug resistance in NSCLC cells. This study might
provide a better understanding of the mechanism underlying NSCLC
development.
Materials and methods
Patients and tissue samples
Paired specimens of NSCLC were obtained from 114
patients, who underwent surgery without preoperative systemic
chemotherapy at the Surgery Department of the Affiliated Hospital
of Nantong University between 2008 and 2016. Immediately after
surgical removal, NSCLC specimens were fixed in formalin and
embedded in paraffin, and 5-µm sections were prepared for
immunohistochemistry. All human tissues were collected in
accordance with protocols approved by the ethics committee of the
Affiliated Hospital of Nantong University. The main clinical and
pathological variables of patients are summarized in Table I.
 | Table I.Expression of PINK1 in 114 human lung
adenocarcinoma tissues. |
Table I.
Expression of PINK1 in 114 human lung
adenocarcinoma tissues.
|
|
| PINK1 expression |
|
|---|
|
|
|
|
|
|---|
| Clinicopathological
features | Total | Low | High | P-value |
|---|
| Age (years) |
|
|
| 0.349 |
|
<60 | 50 | 22 | 28 |
|
|
≥60 | 64 | 33 | 31 |
|
| Gender |
|
|
| 0.708 |
|
Female | 59 | 27 | 32 |
|
|
Male | 55 | 28 | 27 |
|
| Tumor size
(cm) |
|
|
| 0.008a |
|
<3 | 49 | 31 | 18 |
|
| ≥3 | 65 | 24 | 41 |
|
| Smoking status |
|
|
| 0.206 |
|
Yes | 85 | 38 | 47 |
|
| No | 29 | 17 | 12 |
|
| Lymph node
metastasis |
|
|
| 0.001a |
| 0 | 60 | 38 | 22 |
|
|
>0 | 54 | 17 | 37 |
|
| Clinical stage |
|
|
| 0.035a |
| I | 67 | 39 | 28 |
|
| II | 30 | 11 | 19 |
|
|
III | 17 | 5 | 12 |
|
| Histological
differentiation |
|
|
| 0.045a |
|
Well | 26 | 7 | 19 |
|
|
Mod | 63 | 35 | 28 |
|
|
Poor | 25 | 13 | 12 |
|
| Ki-67
expression |
|
|
| 0.000a |
|
Low | 51 | 31 | 20 |
|
|
High | 63 | 24 | 39 |
|
Western blot analysis
The tissues and cell samples were immediately
homogenized in a lysis buffer containing 50 mM Tris-Cl, pH 8.0,
0.1% NP-40, 150 mM NaCl, 1 mM EDTA, 60 mM β-glycerophosphate, 0.1
mM NaF, 0.1 mM sodium orthovanadate, and complete protease
inhibitor cocktail (Roche Diagnostics) and then centrifuged at
12,000 × g, 4°C for 15 min to collect the supernatant. Protein
concentrations were determined using a BCA protein assay kit
(Bio-Rad, Hercules, CA, USA). Subsequently, the supernatants were
added with equal volume of 2X sodium dodecyl sulfate (SDS) sample
buffer and boiled for 15 min. The protein samples were subjected to
10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) separation
and then transferred to polyvinylidene difluoride filter (PVDF)
membranes (Millipore, Bedford, MA, USA). Next, the membranes were
blocked with 5% no-fat milk in TBST (150 mM NaCl, 20 mM Tris and
0.05% Tween-20) for 2 h and then incubated with primary antibodies
overnight at 4°C.
The primary antibodies used for western blotting
were as follows: rabbit polyclonal anti-PINK1 antibody (Abgent, San
Diego, CA, USA), mouse monoclonal anti-p27 antibody (Santa Cruz
Biotechnology, Santa Cruz, CA, USA), rabbit polyclonal anti-cyclin
D1 antibody (Santa Cruz Biotechnology), mouse monoclonal anti-p65
antibody (Santa Cruz Biotechnology), rabbit polyclonal anti-GAPDH
antibody (Santa Cruz Biotechnology), rabbit polyclonal
anti-cleaved-caspase-3 antibody (Santa Cruz Biotechnology), and
rabbit polyclonal anti-cleaved poly (ADP-ribose) polymerase (PARP)
antibody (Immunoway, Newark, DE, USA). Secondary antibody
incubation was performed using horseradish peroxidase linked IgG
(Pierce Biotechnology, Rockford, IL, USA) at a dilution of 1:5000.
The detection of chemiluminescent signals was performed by ECL
method (Zhongshan Biotechnology Co., Ltd., Beijing, China).
Immunohistochemistry and
immunohistochemistrical evaluation
Immunohistochemistry was performed in accordance
with previous reports. In brief, paired tissue sections were
dewaxed, washed, and blocked. Afterward, tissue sections were
incubated overnight at 4°C with primary antibodies: anti-PINK1
(1:100) or mouse monoclonal anti-Ki-67 (1:100; Santa Cruz
Biotechnology), followed by horseradish peroxidase (HRP)-conjugated
secondary antibodies. After rinsing in water, the sections were
counterstained with hematoxylin, dehydrated, and cover slipped. The
slides were then mounted for observation under a fluorescence
microscope.
All immunostained sections were independently
examined by three pathologists in a blinded manner without
knowledge of the clinical and pathological variables of the
patients. At least five high-power fields in each specimen were
randomly selected, and the cytoplasm or nuclear staining was
examined under a high magnification. More than 500 cells were
examined to determine the mean percentage of signal-positive cells.
For determining PINK1 expression, the intensity of immunostaining
was assessed as 0 (negatively or poorly staining), 1 (moderately
staining), and 2 (strongly staining), and according to PINK1
expression ratio (50%, 75%), we divided patients into three groups:
low expression group (<50%) scored 1, moderate expression group
(50–75%) scored 2, and high expression group (>75%) scored 3.
Then, we multiplied the two scores and divided patients into two
groups according to the average scores (4.2): high-expression group
(>4.2) and low expression group (≤4.2). The expression of
proliferation marker Ki-67 was scored in a semi-quantitative
fashion: high expression (≥50%) and low expression (<50%)
(22).
Cell culture and transfection
The human NSCLC cell lines (A549, H1299 and Spca-1)
and normal human bronchial epithelial cell line BEAS-2B were
obtained from the Institute of Cell Biology, Academic Sinica, and
all cells were cultured in the 1640 medium (Gibco BRL, Grand
Island, NY, USA) supplemented with 10% heat-inactivated fetal
bovine serum in 5% CO2 at 37°C.
The PINK1-siRNA and control-siRNA were purchased
from Genechem (Shanghai, China). The PINK1-specific siRNA target
sequence was as follows: PINK1-siRNA#1 was
5′-TCCTCGTTATGAAGAACTA-3′, PINK1-siRNA#2 was
5′-AAGCCATCTTGAACACAAT-3′, PINK1-siRNA#3 was
5′-GCTGGAGGAGTATCTGATA-3′, and PINK1-siRNA#4 was
5′-AGCGTAGCATGTCTGATTT-3′. A549 and H1299 cells were grown in
dishes until they reached 80% confluence. The medium was replaced 6
h later with fresh medium for transfection. A549 and H1299 cells
were transfected with PINK1-siRNA or control-siRNA according to the
manufacturer's instructions. Cells were collected for western
blotting, CCK-8, and flow cytometry assays after transfection for
36 h. A549 and H1299 cells were seeded the day before transfection
using 1640 with 10% fetal bovine serum (FBS) without antibiotics.
A549 and H1299 cells were transfected with the PINK1-siRNA or the
control siRNA using Lipofectamine 2000 transfection reagent
(Invitrogen) according to the manufacturer's protocol, and the
media were replaced with 1640 supplemented with 10% FBS at 6 h
after transfection. The transfected cells were subjected to
subsequent experiments at 48 h after transfection.
Cell cycle analysis and cell
proliferation assay
Cells were harvested, washed twice with ice-cold 1
ml phosphate buffered saline (PBS), and fixed in 70% ethanol for 24
h at 4°C. Then, the cells were washed three times with ice cold 1
ml PBS and incubated with 1 mg/ml RNase A for 30 min at 37°C.
Subsequently, cells were stained with 50 µg/ml propidium iodide
(PI; Becton Dickinson, San Jose, CA, USA) in 0.5% Tween-20 in PBS
and subjected to analysis of cell cycle distribution using a BD
FACScan flow cytometer (Becton Dickinson) coupled with Cell Quest
acquisition and analysis programs.
Cell proliferation was evaluated using the Cell
Counting Kit-8 (CCK-8, Dojindo, Kumamoto, Japan) in accordance with
the manufacturer's protocol. Briefly, A549 and H1299 cells were
seeded at a cell density of 2×104 cells per well into a
96-well plate and grown overnight. For the measurement of CCK-8
absorbance, the cells were incubated with 10 µl CCK-8 reagent
coupled with 100 µl 1640 medium for another 2 h at 37°C in an
incubator. The absorbance was recorded at a test wavelength of 490
nm and a reference wavelength of 650 nm using a microplate reader
(Bio-Rad). These experiments were repeated for at least three times
with similar results.
Colony formation assay
Cells were seeded in a 6-well plate at
5×103 cells per well and cultured for 10 days. Colonies
were fixed for 5 min using 10% formaldehyde and then stained with
1.0% crystal violet for 30 sec, and cell numbers were counted.
Apoptotic analysis
The apoptosis assays were performed at 72 h after
the cells were transfected with PINK1-siRNA or negative control.
The A549 and H1299 cells transfected with PINK1-siRNA or
control-siRNA were washed three times in ice-cold PBS, resuspended
in 100 µl of 1X 19 binding buffer and incubated with Annexin V-FITC
(Bestbio, China) for 15 min at 4°C in the dark, according to the
manufacturer's instructions. After staining, the cells were
incubated with propidium iodide for 5 min at 4°C in the dark and
then analyzed using a flow cytometer (Beckman, Palo Alto, CA,
USA).
Statistical analysis
The SPSS 17.0 software package was used for all
statistical analysis. The association between PINK1 and Ki-67
expression and clinicopathological features was analyzed using the
χ2 test. For analysis of the survival data of patients,
the Kaplan-Meier curves and the log-rank test were performed.
Multivariate analysis was constructed using the Cox proportional
hazards model. The hazard ratio and its 95% confidence interval
were recorded for each variable. P<0.05 was considered
statistically significant. All values were expressed as mean ± SEM
(23).
Results
PINK1 is upregulated in human NSCLC
tissues
To detect a possible involvement of PINK1 in NSCLC,
western blot analysis was performed to examine the expression
pattern of PINK1 in eight paired NSCLC and adjacent non-tumorous
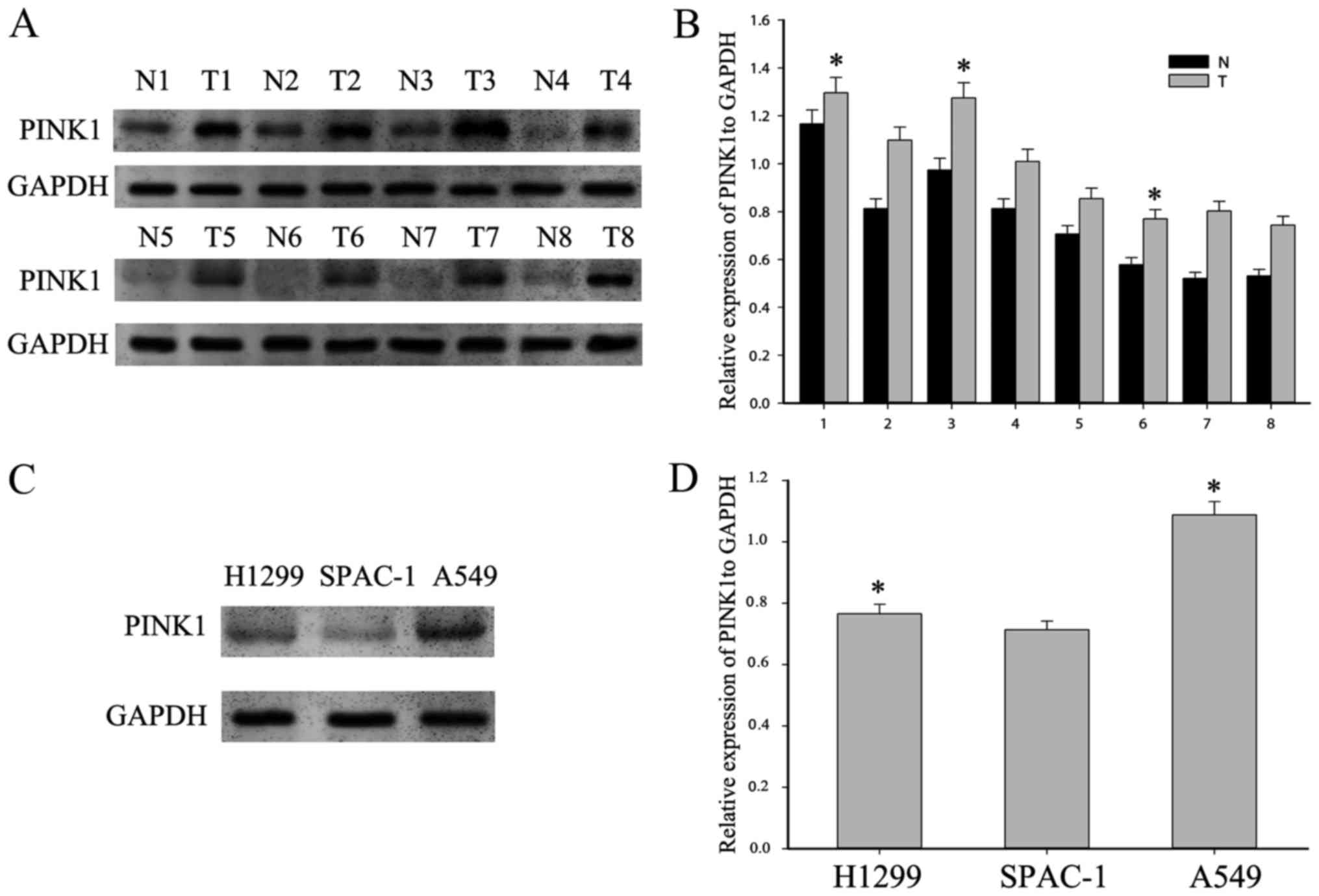
tissues. As shown in Fig. 1A and B,
PINK1 expression was remarkably higher in tumorous tissues than in
adjacent non-tumorous ones. Next, we examined the basic expression
of PINK1 in three human NSCLC cell lines, A549, H1299, and SPCA-1.
We found that PINK1 was highly expressed in NSCLC cell lines
(Fig. 1C). Moreover, we analyzed
the expression of PINK1 in 114 NSCLC tissues using
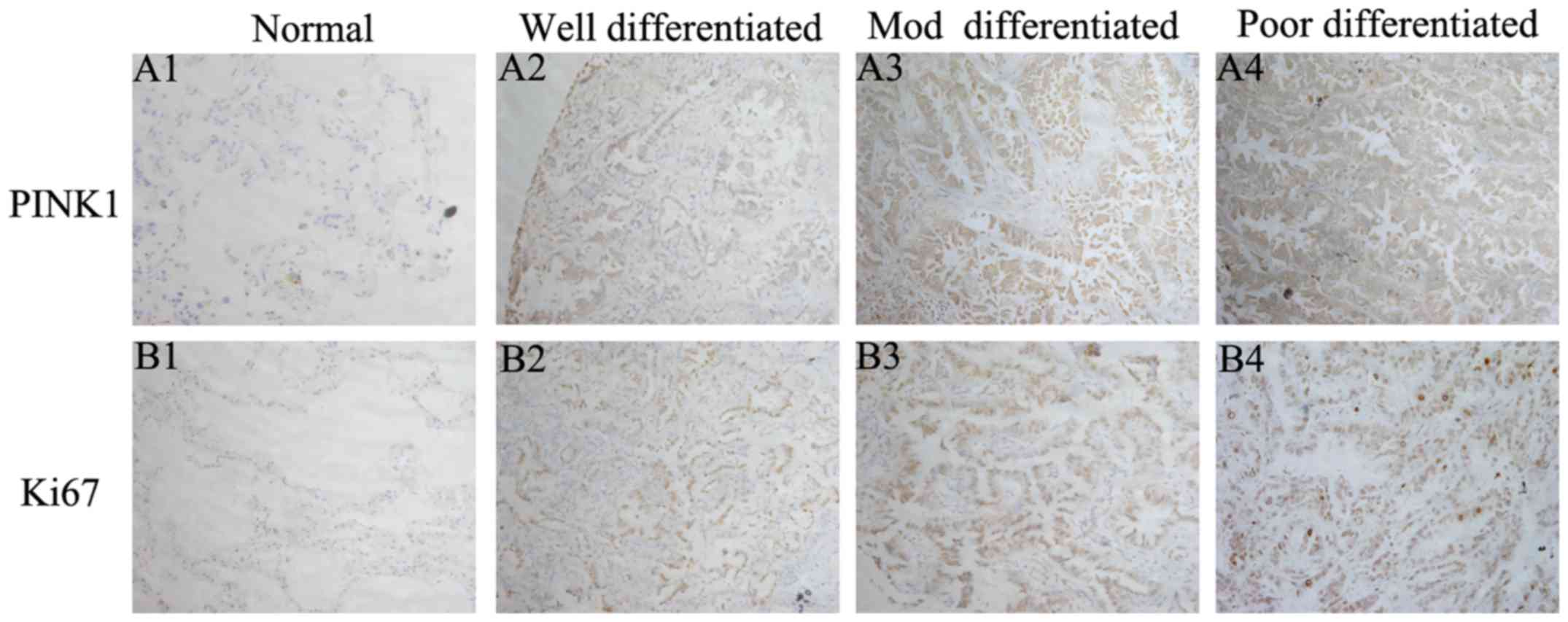
immunohistochemical assay (Fig. 2).
NSCLC tumor tissues displayed cytoplasmic diffuse staining of
PINK1, together with a prominent membrane staining. While for Ki-67
which is a marker for cell proliferation, its immunoreactivity was
found predominantly in the nucleus. As expected, PINK1 was highly
expressed in poorly differentiated specimens compared with
well-differentiated ones, which was consistent with Ki-67. There
was low or even no expression for both markers in adjacent
non-tumor tissues. Thus, PINK1 was upregulated in NSCLC specimens
and might be associated with tumor cell proliferation.
Correlation of PINK1 expression with
clinicopathological variables in NSCLC patients
To further confirm the correlation of PINK1 and
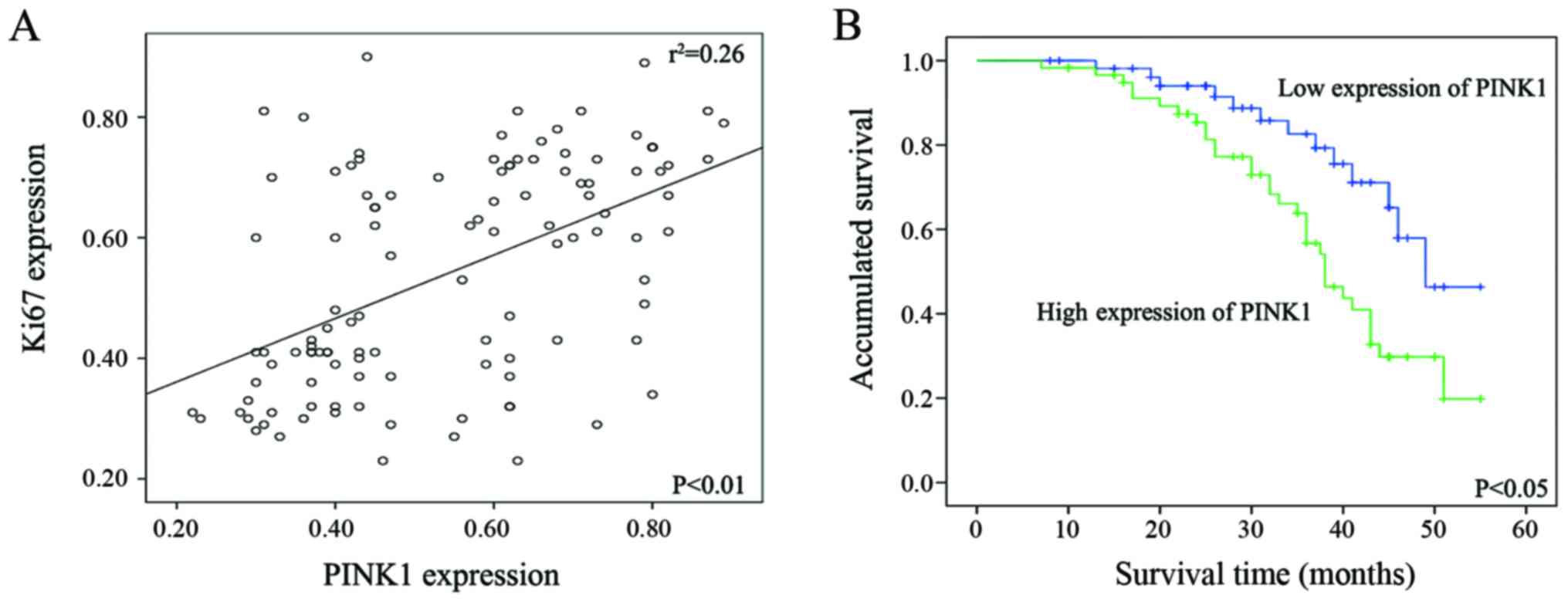
Ki-67 expression by twos, Spearman's correlation test was next
performed by percentage of positive malignant cells. A significant
positive correlation was found between the expression status of
PINK1 and that of Ki-67. Spearman's correlation coefficient
(γ2) for PINK1-Ki-67 equals to 0.037 (P<0.01)
(Fig. 3A).
In addition, we further evaluated the association of
PINK1 expression with clinicopathological variables including Ki67
by Pearson's χ2 test. The level of PINK1 and Ki-67
expression was divided into high group and low group according to
the cutoff value stated in the afore-mentioned methods. The data
are summarized in Table I.
According to Table I, PINK1
expression was correlated with tumor size (P=0.008), lymph node
metastasis (P=0.001), histological differentiation (P=0.045),
clinical stage (P=0.035) and Ki-67 expression (P=0.000), while
there was no correlation with other variables such as age
(P=0.349), gender (P=0.708), and smoking status (P=0.206).
High expression of PINK1 predicted
poor prognosis of NSCLC patients
Next, we used Kaplan-Meier analysis to determine the
effect of PINK1 expression level on patient survival. The result
revealed that NSCLC patients with high PINK1 expression was
significantly associated with poor overall survival rate, compared
with those with low PINK1 expression (Fig. 3B). In addition, Cox proportional
survival analysis showed that both PINK1 (P=0.043), and Ki67
expression (P=0.035) were independent prognostic factors in
patients with NSCLC (Table
II).
| Table II.Contribution of various potential
prognostic factors to survival by Cox regression analysis in 114
NSCLC specimens. |
Table II.
Contribution of various potential
prognostic factors to survival by Cox regression analysis in 114
NSCLC specimens.
|
Characteristics | Hazard ratio | 95.0% confidence
interval | P-value |
|---|
| Age | 0.982 | 0.948–1.017 | 0.317 |
| Gender | 0.846 | 0.538–1.861 | 0.998 |
| Clinical stage | 1.227 | 0.746–2.019 | 0.420 |
| Tumor size | 1.759 | 0.731–4.234 | 0.207 |
| Histological
differentiation | 0.833 | 0.479–1.450 | 0.518 |
| Lymph node
metastasis | 1.307 | 0.632–2.704 | 0.470 |
| Ki67
expression | 0.422 | 0.189–0.943 | 0.035a |
| PINK1
expression | 0.436 | 0.195–0.976 | 0.043a |
PINK1 overexpression promotes
proliferation of NSCLC cells
In a recent study, a novel function for PINK1 was
discovered as a positive regulator of cell cycle progression that
can promote cancer-associated phenotypes (24). Given the fact that PINK1 expression
was tightly associated with the expression of Ki-67 (Fig. 3), which is a cell proliferation
marker, we presume that PINK1 expression would be related with
proliferation and play a role in the regulation of cell cycle
progression in NSCLC cells. Thus, we decided to overexpress PINK1
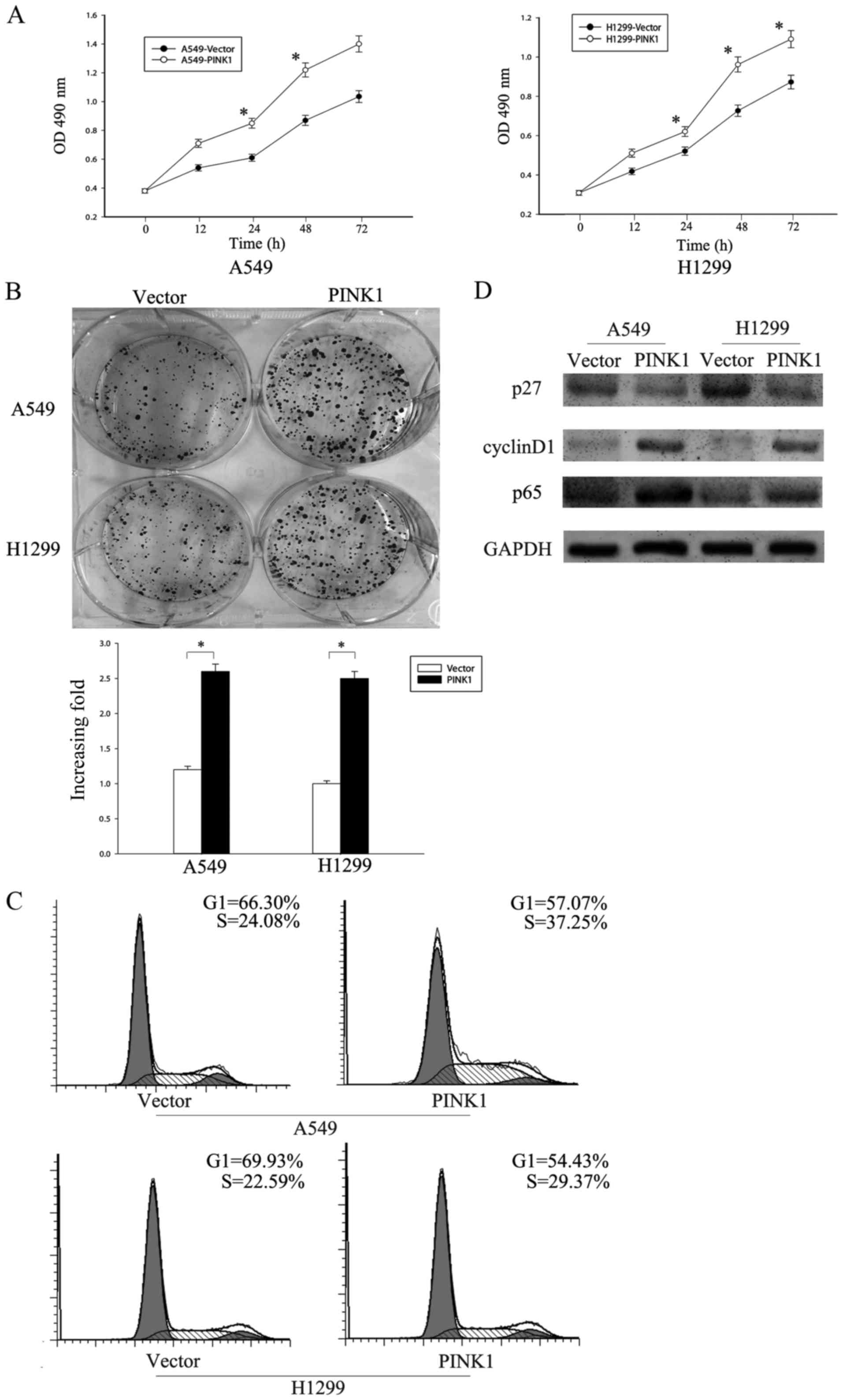
in A549 and H1299 to investigate its effect on cell proliferation.
CCK8 assay showed that overexpression of PINK1 increased the
proliferation rate in both cell lines (Fig. 4A). Colony formation assay also
confirmed that overexpression of PINK1 upregulated the colony
numbers of indicated cells (Fig.
4B).
To further explore the role of PINK1 in promoting
cell proliferation, we investigated the cell cycle progression of
A549 and H1299 following PINK1 overexpression. Flow cytometry
analysis revealed that overexpression of PINK1 increased the
S-phase cell population from 24.08 to 37.25% in A549 and from 22.59
to 29.37% in H1299 with a concomitant decrease in G1 phase
(Fig. 4C). As a CDK inhibitor p27
and CDK regulator cyclin D1 are extremely important in regulating
G1/S transition, we analyzed the expression levels of p27 and
cyclin D1 using western blotting. Our results showed that p27 was
downregulated, while cyclin D1 was upregulated in cells with PINK1
overexpression compared with negative control (Fig. 4D). Since it has been reported that
PINK1 could bind to TRAF6 and TAK1, and facilitate the
autodimerization and autoubiquitination of TRAF6, which leads to
the activation of the NF-κB pathway, we presume that NF-κB pathway
might be a downstream signaling of PINK1 in NSCLC. We analyzed the
expression of p65, which is one of the members in the NF-κB family
since its activation, and found that the level of p65 was
correlated positively with the expression of PINK1 (Fig. 4D). Therefore, these data indicated
that PINK1 expression might have an effect on the proliferation of
NSCLC cells and alter cell cycle progression through the NF-κB
pathway.
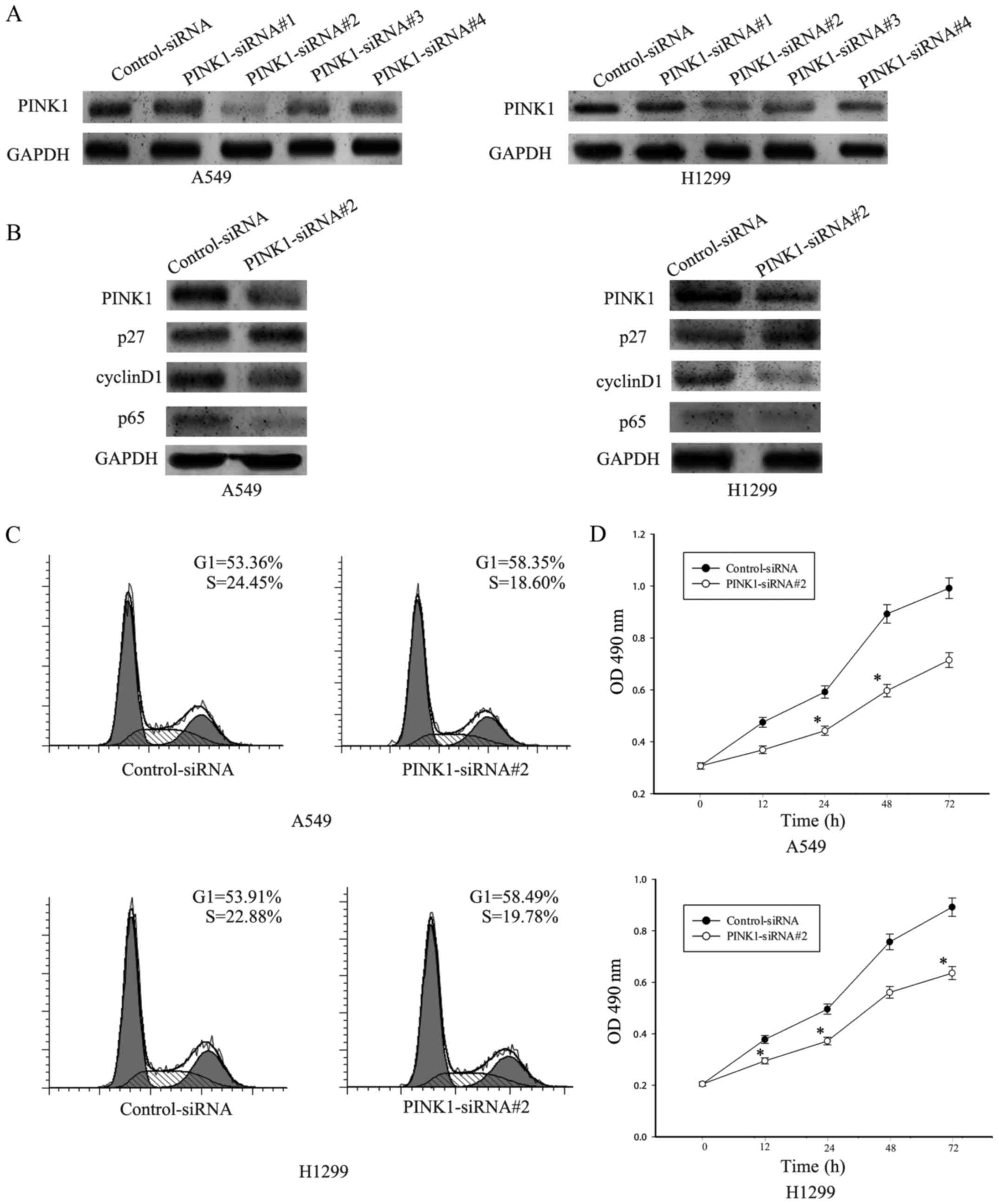
Knockdown of PINK1 inhibits NSCLC
proliferation
To further verify the role of PINK1 in NSCLC cell
proliferation and explore the possible downstream signaling, we
knocked down PINK1 expression in A549 cells using a
lentivirus-mediated RNA interference approach. A549 and H1299 cells
were transiently transfected with PINK1-siRNA#1, PINK1-siRNA#2,
PINK1-siRNA#3, PINK1-siRNA#4 and control siRNA for 36 h. To
determine the efficiency of transfection, western blotting was
used. As shown in Fig. 5A, PINK1
protein levels markedly reduced in both A549 and H1299 cells
infected with PINK1-siRNA, especially in PINK1-siRNA#2, compared
with cells treated with control-siRNA cells. Thus, we used
PINK1-siRNA#2 to carry out the following experiments. Since PINK1
was reported to activate the NF-κB signaling, western blot assay
was performed to detect the expression of p65 which is an important
member in NF-κB signaling family and cell cycle-related protein
including p27 and cyclin D1. As shown in Fig. 5B, knockdown of PINK resulted in
decrease of p65 and cyclin D1, with concomitant increase of p27
which is a CDK inhibitor. Thus these results indicated that PINK1
expression might promote cell cycle progression through NF-κB
signaling.
Furthermore, flow cytometry analysis of cell cycle
showed that A549 and H1299 cells transfected with PINK1-siRNA#2 had
an increase of cell number in the G0/G1 phase from 53.36% to 58.35%
and the number in the S phase decreased from 24.45% to 18.60% in
A549, and from 53.91% to 58.49% in the G0/G1 phase in H1299 and
from 22.88% to 19.78% in the S phase in H1299 (Fig. 5C). CCK-8 assay was used to test the
effect of PINK1 on NSCLC cell growth rate. Knockdown of PINK1 can
attenuate the cell proliferation compared with cells treated with
control-siRNA (Fig. 5C). Taken
these results together, knockdown of PINK1 could inhibit
proliferation of NSCLC cells.
Depletion of PINK1 sensitizes NSCLC
cells to cisplatin (CDDP)
A cytoprotective and chemoresistant function for
PINK1 has been highlighted by some studies, supporting PINK1 as a
target in cancer therapeutics (25). Therefore, we investigated whether
PINK1 could confer chemoresistance to
cis-diamminedichloroplatinum (CDDP), a first-line drug for
treating NSCLC patients. Firstly, CCK8 assay was performed to
determine the sensitivity of NSCLC cells to CDDP. The cell
proliferation rate was decreased in a dose-dependent manner, and
drug sensitivity reached the highest level at the concentration of
25 µmol/l (Fig. 6A). In addition,
to further confirm the contribution of PINK1 to CDDP sensitivity in
NSCLC cells, CCK8 assay and flow cytometry assay were performed to
demonstrate the cell viability level and apoptosis rate following
PINK1 knocking down and CDDP addition. It showed that CDDP addition
led to obvious decrease of cell viability and increase of cell
apoptosis (Fig. 6B and C, comparing
the lower panels to the upper panels).
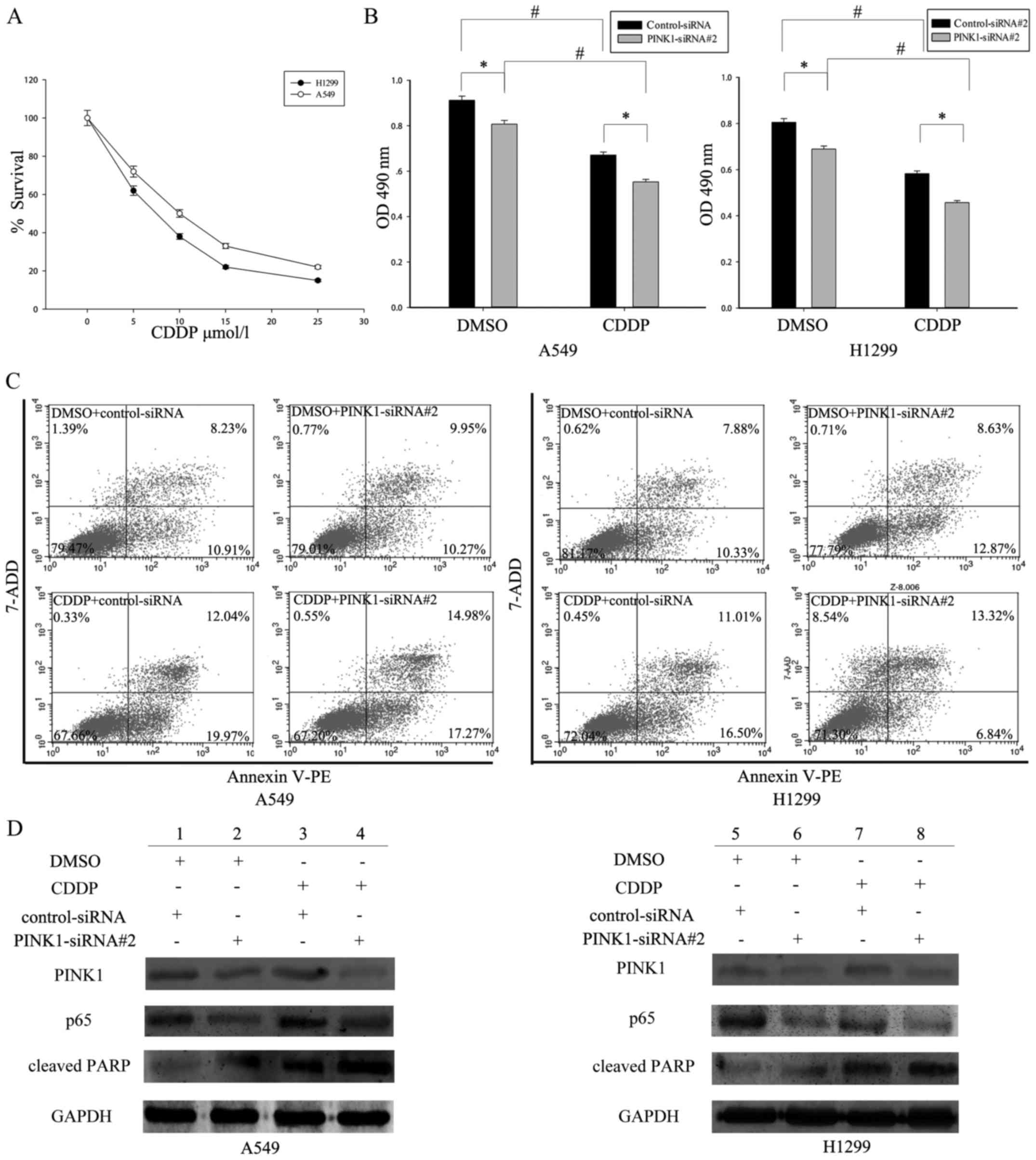 | Figure 6.PINK1-mediated activation of the NF-κB
pathway enhanced cell survival and drug resistance in NSCLC. (A)
A549 and H1299 cells were treated with different concentrations of
cisplatin (CDDP) (0, 5, 10, 15, 20, and 25 µM/l) for 48 h, and cell
viability was measured using the CCK-8 assay. (B) After
PINK1-SiRNA#2 transfection for 48 h, A549 and H1299 cells were
treated with CDDP (25 µM/l) stimulation or not. After 24 h, the
cell proliferation was determined by CCK-8 assay.
*P<0.05,statistically different with the control group;
#P<0.05, statistically different with the
doxorubicin-treated group, the data are mean ± SEM. (C)
Interference of PINK1 promoted CDDP-triggered cell apoptosis in
A549 and H1299 cells as analyzed by Annexin V-FITC/7-ADD double
staining. (D) Western blot analysis of the expression of
apoptosis-related proteins in control siRNA or PINK1-siRNA#2
transfected A549 and H1299 cells treated with or without CDDP. |
In addition, depletion of PINK1 augmented the
cytotoxic effect of CDDP in NSCLC cells (Fig. 6B and C comparing the right panels to
the left panels). Moreover, to test whether PINK1-induced CDDP
resistance depends on NF-κB activity, A549 and H1299 cells were
treated with CDDP for 24 h following PINK1 addition. Western blot
assay was then used to determine the expression of p65, as well as
cleaved caspase-3 and PARP1, which are apoptosis markers. As shown
in Fig. 6D, PINK1 knockdown
resulted in decreased expression of p65 (compare lane 2 to 1, lane
4 to 3 in A549 cells; compare lane 6 to 5, lane 8 to 7 in H1299
cells). Consequently, PINK1-depleted NSCLC cells had much higher
level of apoptosis markers compared with the control silenced
cells, especially following CDDP treatment (compare lane 3 to 1,
lane 4 to 2 in A549 cells; compare lane 7 to 5, lane 8 to 6 in
H1299 cells). Taken together, PINK1 might enhance cell survival and
drug resistance by NF-κB signaling.
Discussion
Despite great progress in diagnostic and therapeutic
strategies, non-small cell lung cancer is still one of the most
common causes of cancer death due to late diagnosis, high
metastasis and chemoresistance. As molecular characterization of
lung cancer has significantly affected treatment strategies, it is
necessary to discover novel therapeutic targets that can complement
present chemotherapy. Although increasing attention has been drawn
toward PINK1 in a number of processes including cancer (13,14,26),
whether PINK1 was involved in NSCLC carcinogenesis has yet not been
clarified. In this study, we investigated the potential role of
PINK1 in NSCLC development. We found that PINK1 was significantly
overexpressed in NSCLC tissues and NSCLC cell lines, and correlated
with clinical pathologic variables of NSCLC. Univariate and
multivariate analysis indicated that PINK1 was an independent
prognostic indicator for the survival of NSCLC patients. Moreover,
we showed that PINK1 knockdown resulted in decreased NSCLC cell
proliferation rate, colony formation ability, and increased cell
cycle arrest. In addition, our results also indicated that PINK1
could induce lung cancer cell survival and chemoresistance through
the nuclear factor-κB (NF-κB) pathway. Taken together, these
results revealed that PINK1 might be a novel therapeutic target for
NSCLC.
PINK1 [phosphatase and tensin homolog deleted on
chromosome 10 (PTEN)-induced kinase 1], a serine/threonine kinase,
is widely expressed, and localizes in mitochondria and cytosol
(27). PINK1 plays a role in
mitochondrial homeostasis and dynamics, including bioenergetics,
mitophagy, fission and fusion. The role of PINK1 in cancer biology
is controversial. However, increasing attention to this kinase in
regulating cell survival systems indicated that PINK1 has a
potential role in tumorigenesis. Early studies indicated that PINK1
protein is highly expressed in breast, colorectal and endometrial
cancer tissues. Moreover, knockdown of PINK1 inhibits
proliferation, colony formation and migration (16), and increases the sensitivity of
cancer cells to numerous stressors (28,29).
Besides, PINK1 was tightly associated with the major oncogenic
PI3-kinase/Akt axis. Recent studies showed that PINK1 can activate
Akt via the mTORC2/mitochondrial control axis to enhance
invasiveness in cancer cells (14),
and accelerated cancer stem cell renewal through Notch signaling
(30). Furthermore, loss of PINK1
may sensitize breast cancer cells to paclitaxel (18), while overexpression of PINK1 can
over-ride the sensitization and lead to chemoresistance (31,32).
This highlighted PINK1 could be a novel target for chemoresistance
in cancer.
NF-κB pathway plays an important role in cell
proliferation, differentiation and chemoresistance in many solid
cancers. The NF-κB family of transcription factors has five
members, Rel, p65, RelB, p50 and p52. Once activated, the NF-κB was
activated and transferred into the nucleus to maintain homeostasis.
It was reported that PINK1 can bind to TRAF6 and TAK1, and finally
activate NF-κB pathway, which indicated PINK1 may induce lung
cancer cell survival and drug resistance by activating the NF-κB
pathway. In this study, we found that the PINK1 knockdown could
inhibit activation of NF-κB signaling, which consequently might
promote proliferation, cell cycle progression and chemoresistance
of NSCLC cells.
In summary, this study showed that PINK1 may
contribute to proliferation and chemoresistance of NSCLC through
NF-κB pathway. As a result, PINK1 may be a novel target for NSCLC.
However, further studies are needed to clarify the precise role of
PINK1 in NSCLC pathogenesis.
Acknowledgements
This study was supported by the National Natural
Science Foundation of China (no. 81501975), University Science
Research Project of Jiangsu Province (15KJB310013), and Nantong
Science and Technology Project (MS12015103).
References
|
1
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Howlader N, Noone AM, Krapcho M, Garshell
J, Miller D, Altekruse SF, Kosary CL, Yu M, Ruhl J, Tatalovich Z,
et al: SEER Cancer Statistics Review. 1975–2012. National Cancer
Institute; 2015 http://seer.cancer.gov/csr/1975_2012/Accessed.
November 18–2015
|
|
3
|
Schiller JH, Harrington D, Belani CP,
Langer C, Sandler A, Krook J, Zhu J and Johnson DH: Eastern
Cooperative Oncology Group: Comparison of four chemotherapy
regimens for advanced non-small-cell lung cancer. N Engl J Med.
346:92–98. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Unoki M and Nakamura Y: Growth-suppressive
effects of BPOZ and EGR2, two genes involved in the PTEN signaling
pathway. Oncogene. 20:4457–4465. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Valente EM, Abou-Sleiman PM, Caputo V,
Muqit MM, Harvey K, Gispert S, Ali Z, Del Turco D, Bentivoglio AR,
Healy DG, et al: Hereditary early-onset Parkinson's disease caused
by mutations in PINK1. Science. 304:1158–1160. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Mills RD, Sim CH, Mok SS, Mulhern TD,
Culvenor JG and Cheng HC: Biochemical aspects of the
neuroprotective mechanism of PTEN-induced kinase-1 (PINK1). J
Neurochem. 105:18–33. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Arena G, Gelmetti V, Torosantucci L,
Vignone D, Lamorte G, De Rosa P, Cilia E, Jonas EA and Valente EM:
PINK1 protects against cell death induced by mitochondrial
depolarization, by phosphorylating Bcl-xL and impairing its
pro-apoptotic cleavage. Cell Death Differ. 20:920–930. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Pridgeon JW, Olzmann JA, Chin LS and Li L:
PINK1 protects against oxidative stress by phosphorylating
mitochondrial chaperone TRAP1. PLoS Biol. 5:e1722007. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wood-Kaczmar A, Gandhi S, Yao Z, Abramov
AY, Miljan EA, Keen G, Stanyer L, Hargreaves I, Klupsch K, Deas E,
et al: PINK1 is necessary for long term survival and mitochondrial
function in human dopaminergic neurons. PLoS One. 3:e24552008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Klinkenberg M, Thurow N, Gispert S,
Ricciardi F, Eich F, Prehn JH, Auburger G and Kögel D: Enhanced
vulnerability of PARK6 patient skin fibroblasts to apoptosis
induced by proteasomal stress. Neuroscience. 166:422–434. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wu Z, Sawada T, Shiba K, Liu S, Kanao T,
Takahashi R, Hattori N, Imai Y and Lu B: Tricornered/NDR kinase
signaling mediates PINK1-directed mitochondrial quality control and
tissue maintenance. Genes Dev. 27:157–162. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lee HJ and Chung KC: PINK1 positively
regulates IL-1β-mediated signaling through Tollip and IRAK1
modulation. J Neuroinflammation. 9:2712012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Akundi RS, Huang Z, Eason J, Pandya JD,
Zhi L, Cass WA, Sullivan PG and Büeler H: Increased mitochondrial
calcium sensitivity and abnormal expression of innate immunity
genes precede dopaminergic defects in Pink1-deficient mice. PLoS
One. 6:e160382011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Berthier A, Navarro S, Jiménez-Sáinz J,
Roglá I, Ripoll F, Cervera J and Pulido R: PINK1 displays
tissue-specific subcellular location and regulates apoptosis and
cell growth in breast cancer cells. Hum Pathol. 42:75–87. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Murata H, Sakaguchi M, Jin Y, Sakaguchi Y,
Futami J, Yamada H, Kataoka K and Huh NH: A new cytosolic pathway
from a Parkinson disease-associated kinase, BRPK/PINK1: Activation
of AKT via mTORC2. J Biol Chem. 286:7182–7189. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
O'Flanagan CH, Morais VA, Wurst W, De
Strooper B and O'Neill C: The Parkinson's gene PINK1 regulates cell
cycle progression and promotes cancer-associated phenotypes.
Oncogene. 34:1363–1374. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Akundi RS, Zhi L and Büeler H: PINK1
enhances insulin-like growth factor-1-dependent Akt signaling and
protection against apoptosis. Neurobiol Dis. 45:469–478. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
MacKeigan JP, Murphy LO and Blenis J:
Sensitized RNAi screen of human kinases and phosphatases identifies
new regulators of apoptosis and chemoresistance. Nat Cell Biol.
7:591–600. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Rathos MJ, Khanwalkar H, Joshi K, Manohar
SM and Joshi KS: Potentiation of in vitro and in vivo antitumor
efficacy of doxorubicin by cyclin-dependent kinase inhibitor
P276-00 in human non-small cell lung cancer cells. BMC Cancer.
13:292013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Yang L, Zhou Y, Li Y, Zhou J, Wu Y, Cui Y,
Yang G and Hong Y: Mutations of p53 and KRAS activate NF-κB to
promote chemoresistance and tumorigenesis via dysregulation of cell
cycle and suppression of apoptosis in lung cancer cells. Cancer
Lett. 357:520–526. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee HJ, Jang SH, Kim H, Yoon JH and Chung
KC: PINK1 stimulates interleukin-1β-mediated inflammatory signaling
via the positive regulation of TRAF6 and TAK1. Cell Mol Life Sci.
69:3301–3315. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ni T, Mao G, Xue Q, Liu Y, Chen B, Cui X,
Lv L, Jia L, Wang Y and Ji L: Upregulated expression of ILF2 in
non-small cell lung cancer is associated with tumor cell
proliferation and poor prognosis. J Mol Histol. 46:325–335. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Xue Q, Zhou Y, Wan C, Lv L, Chen B, Cao X,
Ju G, Huang Y, Ni R and Mao G: Epithelial membrane protein 3 is
frequently shown as promoter methylation and functions as a tumor
suppressor gene in non-small cell lung cancer. Exp Mol Pathol.
95:313–318. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
O'Flanagan CH, Morais VA and O'Neill C:
PINK1, cancer and neurodegeneration. Oncoscience. 3:1–2.
2016.PubMed/NCBI
|
|
25
|
O'Flanagan CH and O'Neill C: Pink1
signalling in cancer biology. Biochim Biophys Acta. 1846:590–598.
2014.PubMed/NCBI
|
|
26
|
Martin SA, Hewish M, Sims D, Lord CJ and
Ashworth A: Parallel high-throughput RNA interference screens
identify PINK1 as a potential therapeutic target for the treatment
of DNA mismatch repair-deficient cancers. Cancer Res. 71:1836–1848.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lin W and Kang UJ: Structural determinants
of PINK1 topology and dual subcellular distribution. BMC Cell Biol.
11:902010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Deng H, Jankovic J, Guo Y, Xie W and Le W:
Small interfering RNA targeting the PINK1 induces apoptosis in
dopaminergic cells SH-SY5Y. Biochem Biophys Res Commun.
337:1133–1138. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cui T, Fan C, Gu L, Gao H, Liu Q, Zhang T,
Qi Z, Zhao C, Zhao H, Cai Q, et al: Silencing of PINK1 induces
mitophagy via mitochondrial permeability transition in dopaminergic
MN9D cells. Brain Res. 1394:1–13. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Lee KS, Wu Z, Song Y, Mitra SS, Feroze AH,
Cheshier SH and Lu B: Roles of PINK1, mTORC2, and mitochondria in
preserving brain tumor-forming stem cells in a noncanonical Notch
signaling pathway. Genes Dev. 27:2642–2647. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Su JL, Cheng X, Yamaguchi H, Chang YW, Hou
CF, Lee DF, Ko HW, Hua KT, Wang YN, Hsiao M, et al:
Foxo3a-dependent mechanism of e1a-induced chemosensitization.
Cancer Res. 71:6878–6887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Sunters A, Madureira PA, Pomeranz KM,
Aubert M, Brosens JJ, Cook SJ, Burgering BM, Coombes RC and Lam EW:
Paclitaxel-induced nuclear translocation of FOXO3a in breast cancer
cells is mediated by c-Jun NH2-terminal kinase and Akt. Cancer Res.
66:212–220. 2006. View Article : Google Scholar : PubMed/NCBI
|