Introduction
The receptor tyrosine kinases (RTKs) are part of the
larger family of protein tyrosine kinases that relay signals from
extracellular growth factors into the cells (1,2). They
regulate many essential cellular processes in mammalian development
and adult tissue homeostasis. However, although RTKs are critical
in maintaining normal physiology, dysregulation of certain RTKs has
been implicated during the development and progression of several
types of cancer (3).
AXL is a member of the Tyro2-Axl-Mer (TAM) RTK
subfamily (4,5), and is activated by the vitamin
K-dependent protein product of the growth arrest-specific gene 6
(Gas6) (6,7). The biological function of AXL/Gas6
signaling is associated with cell growth and survival in normal and
cancer cells (8). AXL
overexpression and signaling, in particular, has been implicated in
progression of cancers such as glioma, melanoma, and colon, breast,
gastric, and lung cancer (9–14).
Some reports have demonstrated that AXL overexpression is
correlated with poor prognosis in several cancers (15–17).
It has also been suggested that AXL regulates metastasis in various
cancer types (18–20). Our previous studies, along with
recent findings, have reported that AXL mediates cancer cell
resistance to multiple targeted agents such as ALK (21), EGFR (22,23),
or BRAF inhibitors (24). Thus, AXL
has now been recognized as a potential therapeutic target in
various tumors.
The MET proto-oncogene encodes for the RTK c-MET,
which is also known as hepatocyte growth factor receptor. Various
cells widely express MET, where it is essential for embryonic
development (15,25) and tissue repair (26,27).
MET is activated by a single ligand, termed either hepatocyte
growth factor (HGF) or scatter factor. In cancer cells, aberrant
signaling of MET may occur via various mechanisms such as gene
amplification, overexpression, mutations, increased autocrine or
paracrine ligand-mediated stimulation, and interaction with another
cell-surface receptor. In addition, a number of studies have
demonstrated that MET is overexpressed in several types of cancers,
including lung, breast, ovary, kidney, colon, thyroid, and liver
cancer. Aberrant activation of MET under any pathological condition
can confer proliferative, survival, and metastatic abilities on
cancer cells (28–34). Thus, along with AXL, MET is also a
potential biomarker and therapeutic target in cancer therapy.
Our previous studies have shown that AXL and MET are
associated with acquired resistance to EGFR tyrosine kinase
inhibitors (TKIs) (23,32). In fact, many studies have
demonstrated that each of these two receptors is associated with
chemoresistance, metastasis, cell proliferation and survival, and
poor survival in patients with non-small cell lung cancer (NSCLC)
(14,33–36).
Salian-Mehta et al first demonstrated that crosstalk between
AXL and MET promotes neuronal cell migration and survival through
gonadotropin releasing hormone (37). However, the possibility of crosstalk
between AXL and MET in cancer cells, and the role of their
interaction in cancer progression, is still unknown. The aim of
this study was to evaluate the potential role of AXL and MET in
lung metastasis, and investigate their association with patient
outcomes.
Materials and methods
Cell culture and reagents
The human NSCLC cell lines A549, H2009, and Calu-1
were obtained from the American Type Culture Collection (Rockville,
MD, USA). The cells were cultured in RPMI-1640 medium supplemented
with 10% fetal bovine serum, 100 U/ml penicillin, and 100 µg/ml
streptomycin (Life Technologies, Carlsbad, CA, USA) at 37°C in
humidified air containing 5% CO2. HGF and Gas6 were
purchased from Sigma-Aldrich (St. Louis, MO, USA). PHA665752 and
XL880 were purchased from Selleck Chemicals (Houston, TX, USA).
MTT assay
Cells were seeded in 96-well sterile plastic plates
overnight and then treated with drugs. After 72 h, 15 µl of MTT
solution (5 mg/ml) was added to each well and the plates were
incubated for 4 h. Crystalline formazan was solubilized with 100 µl
of a 10% (w/v) SDS solution for 24 h, and then absorbance at 595 nm
was read spectrophotometrically using a microplate reader. The
results are representative of at least three, independent
experiments, and the error bars signify standard deviations
(SDs).
Western blot analysis
The cells were lysed in EBC buffer containing 50 mM
Tris-HCl (pH 8.0), 120 mM NaCl, 1% Triton X-100, 1 mM EDTA, 1 mM
EGTA, 0.3 mM phenylmethylsulfonyl fluoride, 0.2 mM sodium
orthovanadate, 0.5% NP-40, and 5 U/ml aprotinin. To evaluate the
interaction of AXL and MET, lysates were immunoprecipitaed with an
anti-AXL or MET antibody. Antibodies specific for p-EGFR (Tyr1173),
EGFR, MET, and β-actin were obtained from Santa Cruz Biotechnology
(Santa Cruz, CA, USA), and antibodies specific for p-MET
(Tyr1234/1235) and p-AXL (Tyr702) were purchased from Cell
Signaling Technology (Beverly, MA, USA). Proteins were detected by
the enhanced chemiluminescence system (Thermo Scientific, Rockford,
IL, USA) according to the manufacturer's instructions.
Migration and invasion assays
Migration and invasion assays were performed
according to a previously described method (38). Results are representative of at
least three independent experiments, and migrated or invaded cells
were counted under a microscope. Results were expressed as means ±
standard deviations.
Lentivirus-mediated shRNA
infection
The RNAi Consortium clone IDs for the shRNAs used in
this study are as follows: TRCN0000194971 (shAXL) and
TRCN0000196685 (shMET). For lentiviral infection, the cells were
infected with shControl, shAXL, or shMET lentivirus. To validate
the migration and invasion ability, the cells were infected with
shControl, shAXL, or shMET for 48 h, and then treated with 2 µg/ml
puromycin for 72 h. The suppression of each protein was confirmed
by western blotting before performing the migration and invasion
assay.
In vivo studies
All animal studies were conducted following a
protocol approved by the Institutional Animal Care and Use
Committee of the Dongnam Institute of Radiological and Medical
Sciences. BALB/c nude mice (male, 18–20 g, 6-week old) were
purchased form SLC Co. (Shizuoka, Japan). All experiments used 5
mice per group. Cells (1×106) stably expressing control,
AXL, or MET shRNA were suspended in 100 µl serum-free RPMI-1640
medium, and injected into the tail vein or the right flank of mice.
After 21 days, lung metastasis was determined by examining serial
sections of every lung tissue block by microscopy, and tumor of the
right flank of mice were extracted and weighed.
Patient and study design
For this retrospective study, a total of 126
patients were recruited from the Asan Medical Center. All patients
underwent curative resection for NSCLC between January 2006 and
December 2010 and were diagnosed with stage II (the American Joint
Committee on Cancer seventh edition). All participants had adequate
tumor specimens for immunohistochemial staining and detailed
prognosis records. Patients with other primary cancers that could
affect survival were excluded. Patients who died of perioperative
complications within 3 months of surgery were also excluded.
Clinical, pathological, and radiological data were retrospectively
reviewed, as well as the follow-up information obtained until
December 2013. Tumor recurrences were assessed by computed
tomography, magnetic resonance imaging, or bone scans. The primary
endpoint was to assess whether the expression of AXL and MET
affected survival in terms of disease-free survival (DFS). DFS was
defined as the time from resection to locoregional or distant
recurrences or death from any cause (whichever was earlier). The
institutional review board approved this study protocol.
Immunohistochemical staining of AXL
and MET and analysis of AXL and MET expression
Paraffin-embedded tumor samples were collected from
the patients and deparaffinized. After rehydration in alcohol,
immunohistochemical staining for AXL and MET was performed using
the anti-AXL antibody (Santa Cruz Biotechnology) and the anti-MET
antibody (Ventana Medical Systems, Tucson, AZ, USA), respectively.
Immunohistochemical analyses were evaluated at the Asan Medical
Center. The expression levels of AXL and MET were scored
semiquantitatively according to standard protocols. The percentage
of positively stained tumor cells was scored as follows: 0
(<5%), 1 (5–25%), 2 (25–50%), or 3 (>50%). The staining
intensity was scored as follows: 0 (no staining), 1 (weakly
stained), 2 (moderately stained), or 3 (strongly stained). Based on
the immunohistochemical staining scores, which were obtained by
adding the positive proportion scores to the intensity scores, the
tumors were classified into negative tumors (score 0–3) and
positive tumors (score 4–6).
Statistical analysis
The Kruskal-Wallis test was performed for
statistical analysis (for continuous variables), and the Chi-square
test or Fisher's exact test was performed for categorical
variables. Survival rates were estimated by the Kaplan-Meier method
and compared by the log-rank test. All P-values reported were the
result of two-sided tests, and values <0.01 were considered
statistically significant. All statistical analyses were performed
with SAS software, version 9.4 (SAS Institute, Inc., Cary, NC,
USA).
Results
Activation of AXL and MET receptor
promotes migration and invasion of NSCLC cells
We have previously demonstrated that the expression
of AXL and MET is associated with acquired resistance to EGFR-TKIs
(23,32). In addition, these receptors have
been recently referred to as biomarkers that predict poor prognosis
of various tumors (15–17,28–30).
Thus, we investigated whether AXL and MET signaling affects the
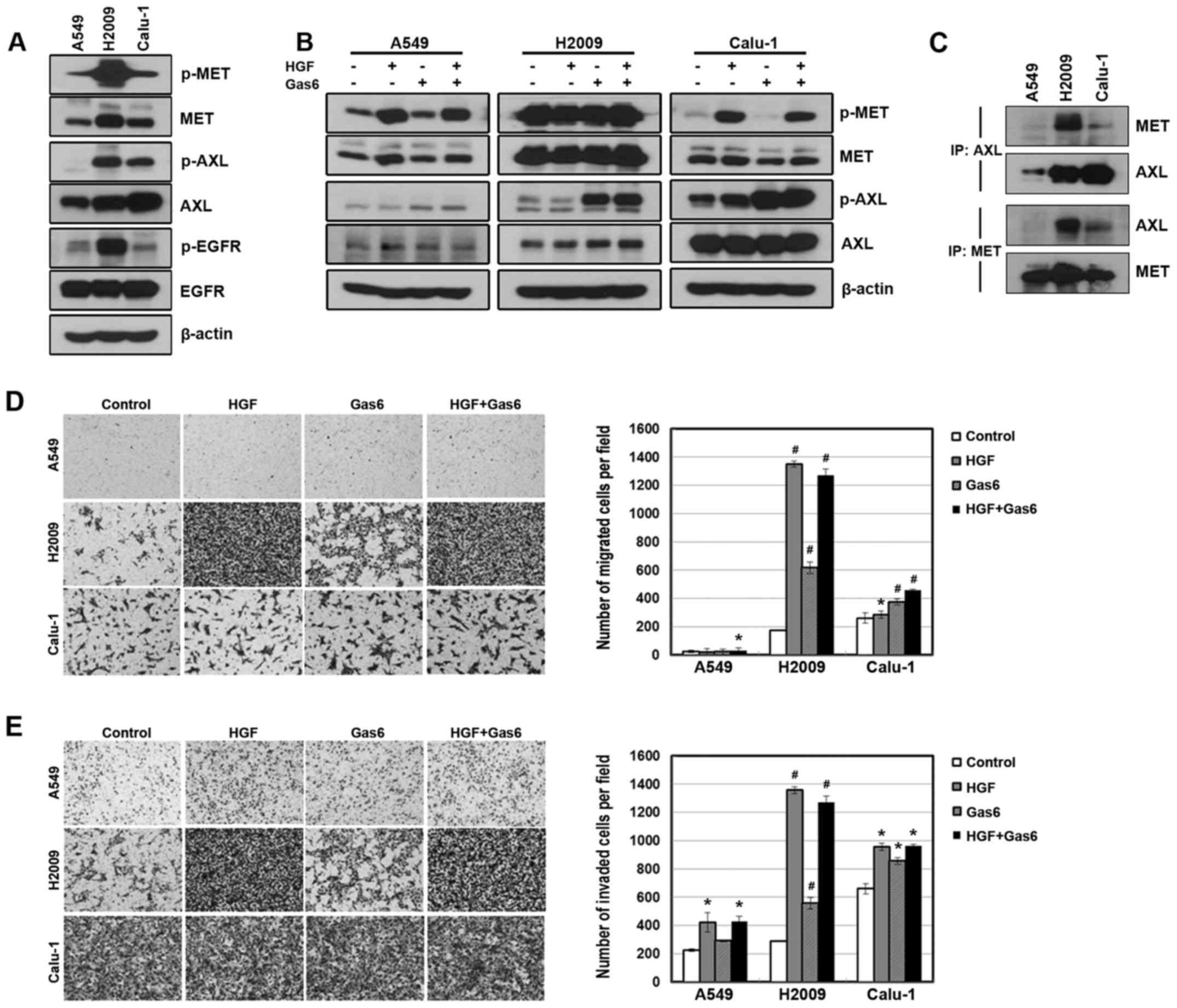
cellular mobility of NSCLC cells. We found that AXL and MET were
expressed or activated in three NSCLC cell lines, although the
expression and activation of these receptors was more or less cell
line-dependent (Fig. 1A). Treatment
of the cells with ligands to each receptor increased the activity
of the receptors (Fig. 1B). In
addition, AXL receptors interacted with MET receptors at basal
level of endogenous proteins, although the interaction of the
receptors was weak in A549 cells (Fig.
1C). Consistent with this observation, ligand treatment
significantly enhanced the migration and invasiveness of the cells
(Fig. 1D and E). However, the
combined treatment of the ligands did not show a synergistic effect
on the migratory and invasive abilities.
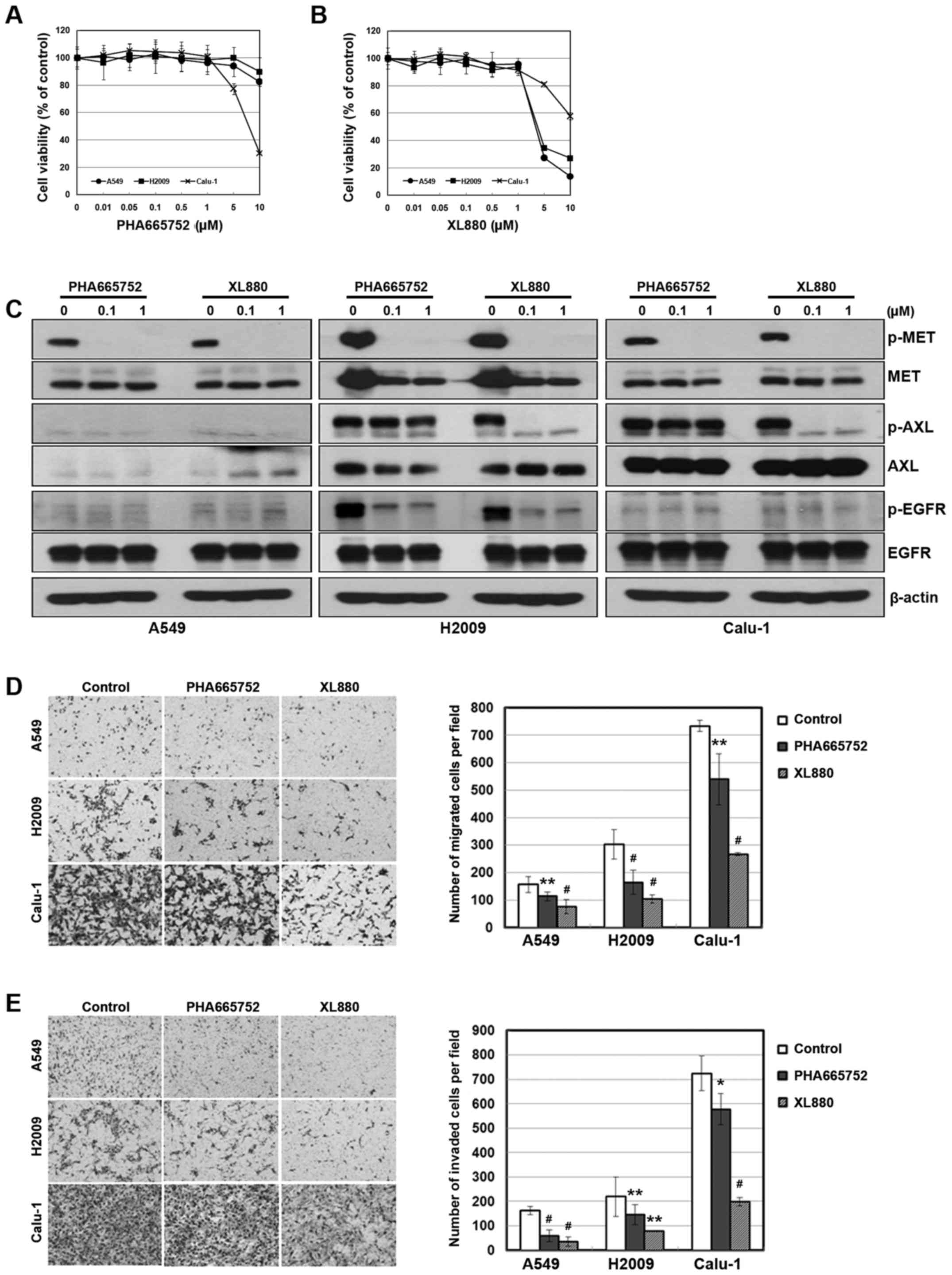
To further investigate the relationship between AXL
or MET signaling and cellular mobility, we treated the cells with
inhibitors of each receptor. We used the PHA665752 and XL880 as a
selective inhibitor of MET and AXL, respectively. Although XL880
did not show selectivity to the AXL receptor, both the inhibitors
effectively inhibited the activity of their corresponding receptors
(Fig. 2C). In addition, cellular
toxicity was not revealed until inhibitor concentrations were 1 µM
or higher (Fig. 2A and B). We found
that the inhibition of AXL or MET resulted in reduced migration and
invasiveness (Fig. 2D and E).
Notably, XL880 treatment was more effective in the inhibition of
cellular mobility than PHA665752 treatment.
AXL signaling is more potent than MET
in inhibition of metastasis
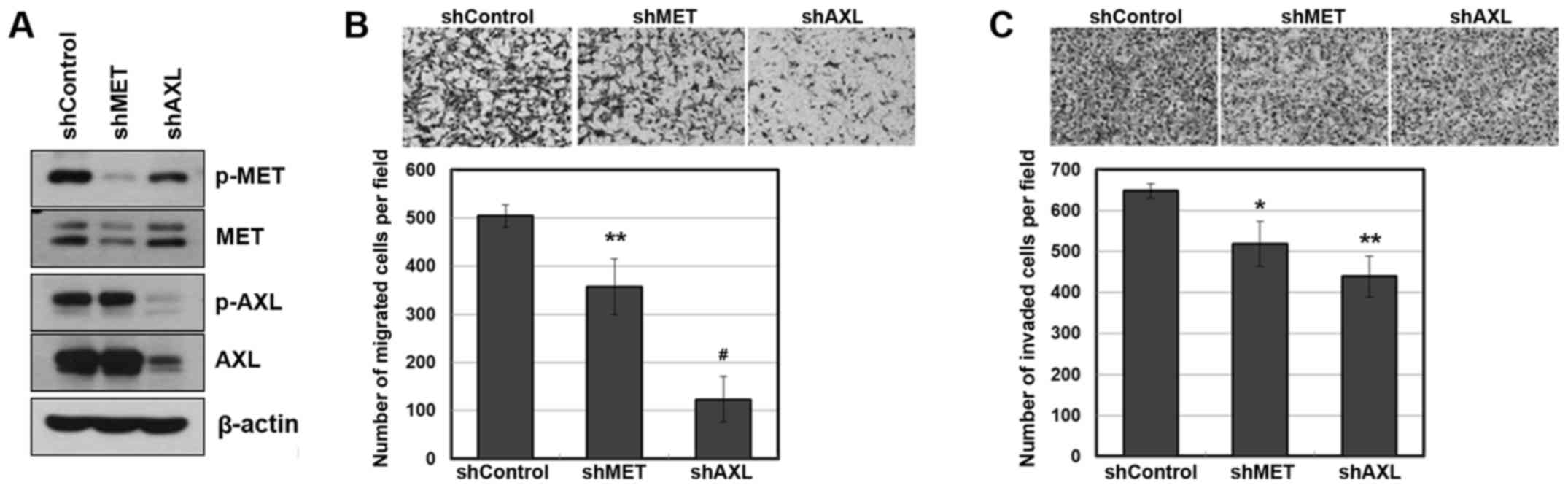
To further evaluate the role of AXL and MET in
metastasis, we used an shRNA against each receptor. Calu-1 cells
were infected with AXL or MET-specific shRNA lentiviral particles,
which substantially suppressed the amount and activity of each
receptor, as determined by western blotting (Fig. 3A). As a consequence, the suppression
of AXL or MET resulted in reduced migratory and invasive
capabilities (Fig. 3B and C).
Notably, the suppression of AXL was more effective in the
inhibition of cellular mobility than the suppression of MET
(P=0.00034 and P=0.00032 for AXL shRNA versus MET shRNA, in
migration assay and invasion assay, respectively).
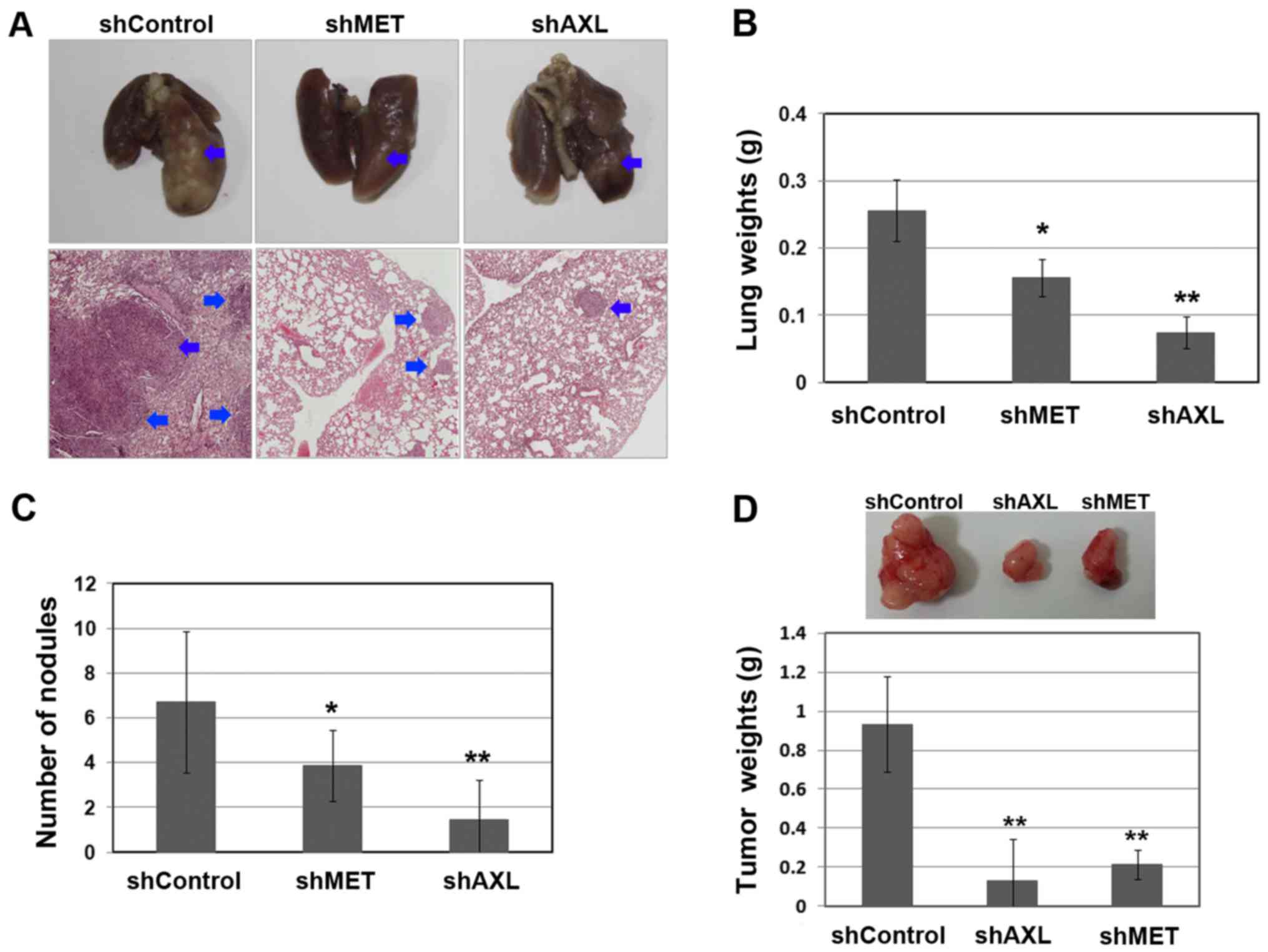
To determine whether AXL and MET play a key role in
the regulation of lung colonization and metastasis, we
downregulated the AXL and MET expression in Calu-1 cells using
shRNA before implantation in the tail vein injections, and then
examined the lung weight and number of tumor nodules, respectively.
As shown in Fig. 4, knockdown of
AXL or MET significantly reduced the lung tumor colonization and
lung weight. Consistent with in vitro studies, AXL knockdown
yielded a significantly higher relative inhibition of metastasis
than MET knockdown (P=0.012 and P=0.035 for AXL shRNA versus MET
shRNA, in lung weight and number of tumor nodules, respectively).
In addition, the suppression of AXL or MET significantly reduced
the tumor cell proliferation (Fig.
4D). Taken together, the results indicate that AXL and MET
signaling are associated with cellular mobility and metastasis as
well as proliferation in lung cancer, but AXL inhibition might
exhibit higher efficacy in blocking metastasis than MET
inhibition.
Clinical significance of AXL and MET
expression in NSCLC cells
A total of 126 patients with surgically resected
NSCLC were analyzed. The median patient age was 63 years (range,
56–69 years). Of the 126 patients, 26 (20.6%) were female. All the
patients had stage II lung cancer. Regarding histological tumor
types, 53 patients (42.1%) had adenocarcinoma and 73 (57.9%) had
squamous cell carcinoma.
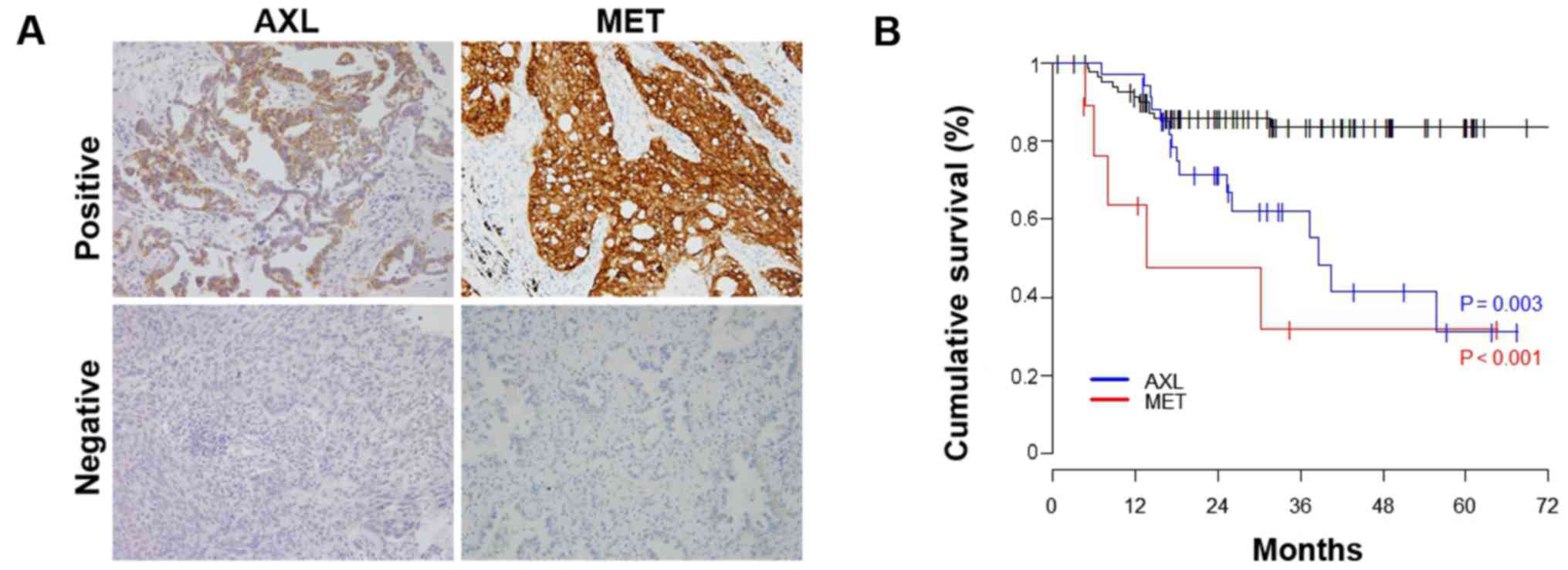
As shown in Fig. 5A,
of the 126 patients, 34 (27.0%) had AXL-positive tumors and 9
(5.6%) had MET-positive tumors. The 1- and 2-year DFS rates for
AXL-positive patients were 97.1 and 71.3%, respectively; and for
MET-positive patients, were 63.5 and 47.6%, respectively. DFS was
significantly shorter in the AXL-positive group than in the
negative group (2-year DFS rate 71.3 vs 85.8%, P=0.003, Fig. 5B). In addition, DFS was
significantly shorter in the MET-positive group than in the
negative group (2-year DFS rate 47.6 vs 85.8%, P<0.001, Fig. 5B).
Discussion
Although AXL and MET have recently been associated
with acquired resistance to anticancer drugs, they are well known
for their roles in tumor progression, such as those in development,
survival and metastasis (5,39–41).
However, crosstalk between AXL and MET in tumor progression remains
unclear. In general, the interaction between RTKs and non-RTKs, or
self-assembly of homodimers or heterodimers, plays an important
role in cellular signaling. This interaction is mediated in a
ligand-dependent or -independent manner. In this study, we found
that HGF-dependent MET activation enhanced the AXL activity in
Calu-1 cells, but not in the other cells. However, all cells showed
interaction of both AXL and MET at endogenous levels. Previous
studies have demonstrated the physical interaction of AXL and MET
by co-immunoprecipitation under conditions of ectopic expression
(42). Notably, we found that AXL
knockdown decreased the MET activity without reduction of MET
expression. AXL and MET signaling share similar downstream
molecules such as PI3K and Akt. Thus, a better understanding of
crosstalk between AXL and MET is needed to evaluate their roles in
tumor progression.
Many reports have suggested that both AXL and MET
contribute to metastasis in many types of cancer (3,5,10,20,
36,39–41).
We showed that Gas6 or HGF treatment enhanced the migration and
invasiveness of the cells, whereas the inhibitors of each receptor
decreased these properties. We used the PHA66572 and XL880 to
inhibit the activity of MET and AXL, respectively. However, XL880
treatment was more potent than PHA665752 treatment in the
inhibition of the migratory and invasive abilities. The difference
in inhibitory rates may result from that fact that XL880 is a
multi-kinase inhibitor, and inhibits AXL as well as MET. In
addition, AXL knockdown was found to be more effective than MET
knockdown in the inhibition of cellular mobility in vitro
and lung metastasis in vivo. This might have resulted from
crosstalk between AXL and MET, wherein AXL knockdown leads to
inhibition of MET activity.
The essential steps required for tumor metastasis
include penetration of tumor cells into the blood or lymphatic
vessels, circulation through the intravascular stream, and
proliferation at another site (43). AXL and MET signaling promote
cellular proliferation in normal as well as tumor cells. Thus, they
can play an important role when tumor cells proliferate at another
site. Our results showed that the suppression of AXL and MET
reduced tumor cell proliferation. Consistent with the results of
metastasis, tumor cell proliferation was remarkably decreased upon
suppression of AXL as compared with that upon suppression of MET.
These results suggested that AXL is more effective than MET in
regulating the inhibition of lung metastasis.
Consistent with our results, previous data have
shown that high AXL or MET expression is an adverse prognostic
factor (44–48). In our data, DFS was remarkably
shorter in MET-positive patients than in AXL-positive patients
during the early periods, but there was no significant difference
between the two groups after 4 years. Owing to the small number of
double-positive patients, the DFS of both AXL- and MET-positive
patients could not be evaluated. Thus, more clinical samples are
needed to investigate crosstalk between AXL and MET.
In conclusion, AXL and MET are associated with cell
proliferation and metastasis in lung cancer, and the crosstalk
between these receptors affects tumor progression.
Acknowledgements
This study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF, grant 2014R1A2A2A01003200438 to J.C.L.).
References
|
1
|
Robinson DR, Wu YM and Lin SF: The protein
tyrosine kinase family of the human genome. Oncogene. 19:5548–5557.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Schlessinger J: Cell signaling by receptor
tyrosine kinases. Cell. 103:211–225. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Krause DS and Van Etten RA: Tyrosine
kinases as targets for cancer therapy. N Engl J Med. 353:172–187.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hafizi S and Dahlbäck B: Signalling and
functional diversity within the Axl subfamily of receptor tyrosine
kinases. Cytokine Growth Factor Rev. 17:295–304. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Linger RM, Keating AK, Earp HS and Graham
DK: TAM receptor tyrosine kinases: Biologic functions, signaling,
and potential therapeutic targeting in human cancer. Adv Cancer
Res. 100:35–83. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Manfioletti G, Brancolini C, Avanzi G and
Schneider C: The protein encoded by a growth arrest-specific gene
(gas6) is a new member of the vitamin K-dependent proteins related
to protein S, a negative coregulator in the blood coagulation
cascade. Mol Cell Biol. 13:4976–4985. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Schneider C, King RM and Philipson L:
Genes specifically expressed at growth arrest of mammalian cells.
Cell. 54:787–793. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Goruppi S, Ruaro E and Schneider C: Gas6,
the ligand of Axl tyrosine kinase receptor, has mitogenic and
survival activities for serum starved NIH3T3 fibroblasts. Oncogene.
12:471–480. 1996.PubMed/NCBI
|
|
9
|
Berclaz G, Altermatt HJ, Rohrbach V,
Kieffer I, Dreher E and Andres AC: Estrogen dependent expression of
the receptor tyrosine kinase axl in normal and malignant human
breast. Ann Oncol. 12:819–824. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Craven RJ, Xu LH, Weiner TM, Fridell YW,
Dent GA, Srivastava S, Varnum B, Liu ET and Cance WG: Receptor
tyrosine kinases expressed in metastatic colon cancer. Int J
Cancer. 60:791–797. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hutterer M, Knyazev P, Abate A, Reschke M,
Maier H, Stefanova N, Knyazeva T, Barbieri V, Reindl M, Muigg A, et
al: Axl and growth arrest-specific gene 6 are frequently
overexpressed in human gliomas and predict poor prognosis in
patients with glioblastoma multiforme. Clin Cancer Res. 14:130–138.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Quong RY, Bickford ST, Ing YL, Terman B,
Herlyn M and Lassam NJ: Protein kinases in normal and transformed
melanocytes. Melanoma Res. 4:313–319. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sawabu T, Seno H, Kawashima T, Fukuda A,
Uenoyama Y, Kawada M, Kanda N, Sekikawa A, Fukui H, Yanagita M, et
al: Growth arrest-specific gene 6 and Axl signaling enhances
gastric cancer cell survival via Akt pathway. Mol Carcinog.
46:155–164. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Shieh YS, Lai CY, Kao YR, Shiah SG, Chu
YW, Lee HS and Wu CW: Expression of axl in lung adenocarcinoma and
correlation with tumor progression. Neoplasia. 7:1058–1064. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Brand-Saberi B, Müller TS, Wilting J,
Christ B and Birchmeier C: Scatter factor/hepatocyte growth factor
(SF/HGF) induces emigration of myogenic cells at interlimb level in
vivo. Dev Biol. 179:303–308. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Hattori S, Kikuchi E, Kosaka T, Miyazaki
Y, Tanaka N, Miyajima A, Mikami S and Oya M: Relationship between
increased expression of the Axl/Gas6 signal cascade and prognosis
of patients with upper tract urothelial carcinoma. Ann Surg Oncol.
23:663–670. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Reichl P, Dengler M, van Zijl F, Huber H,
Führlinger G, Reichel C, Sieghart W, Peck-Radosavljevic M,
Grubinger M and Mikulits W: Axl activates autocrine transforming
growth factor-β signaling in hepatocellular carcinoma. Hepatology.
61:930–941. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Brand TM, Iida M, Stein AP, Corrigan KL,
Braverman CM, Coan JP, Pearson HE, Bahrar H, Fowler TL, Bednarz BP,
et al: AXL is a logical molecular target in head and neck squamous
cell carcinoma. Clin Cancer Res. 21:2601–2612. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Dunne PD, McArt DG, Blayney JK, Kalimutho
M, Greer S, Wang T, Srivastava S, Ong CW, Arthur K, Loughrey M, et
al: AXL is a key regulator of inherent and chemotherapy-induced
invasion and predicts a poor clinical outcome in early-stage colon
cancer. Clin Cancer Res. 20:164–175. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee HJ, Jeng YM, Chen YL, Chung L and Yuan
RH: Gas6/Axl pathway promotes tumor invasion through the
transcriptional activation of Slug in hepatocellular carcinoma.
Carcinogenesis. 35:769–775. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Debruyne DN, Bhatnagar N, Sharma B, Luther
W, Moore NF, Cheung NK, Gray NS and George RE: ALK inhibitor
resistance in ALK(F1174L)-driven neuroblastoma is associated with
AXL activation and induction of EMT. Oncogene. 35:3681–3691. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Giles KM, Kalinowski FC, Candy PA, Epis
MR, Zhang PM, Redfern AD, Stuart LM, Goodall GJ and Leedman PJ: Axl
mediates acquired resistance of head and neck cancer cells to the
epidermal growth factor receptor inhibitor erlotinib. Mol Cancer
Ther. 12:2541–2558. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Zhang Z, Lee JC, Lin L, Olivas V, Au V,
LaFramboise T, Abdel-Rahman M, Wang X, Levine AD, Rho JK, et al:
Activation of the AXL kinase causes resistance to EGFR-targeted
therapy in lung cancer. Nat Genet. 44:852–860. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Müller J, Krijgsman O, Tsoi J, Robert L,
Hugo W, Song C, Kong X, Possik PA, Cornelissen-Steijger PD, Foppen
Geukes MH, et al: Low MITF/AXL ratio predicts early resistance to
multiple targeted drugs in melanoma. Nat Commun. 5:57122014.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Heymann S, Koudrova M, Arnold H, Köster M
and Braun T: Regulation and function of SF/HGF during migration of
limb muscle precursor cells in chicken. Dev Biol. 180:566–578.
1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Borowiak M, Garratt AN, Wüstefeld T,
Strehle M, Trautwein C and Birchmeier C: Met provides essential
signals for liver regeneration. Proc Natl Acad Sci USA.
101:10608–10613. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Huh CG, Factor VM, Sánchez A, Uchida K,
Conner EA and Thorgeirsson SS: Hepatocyte growth factor/c-met
signaling pathway is required for efficient liver regeneration and
repair. Proc Natl Acad Sci USA. 101:4477–4482. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Di Renzo MF, Poulsom R, Olivero M,
Comoglio PM and Lemoine NR: Expression of the Met/hepatocyte growth
factor receptor in human pancreatic cancer. Cancer Res.
55:1129–1138. 1995.PubMed/NCBI
|
|
29
|
Lengyel E, Prechtel D, Resau JH, Gauger K,
Welk A, Lindemann K, Salanti G, Richter T, Knudsen B, Woude GF
Vande, et al: C-Met overexpression in node-positive breast cancer
identifies patients with poor clinical outcome independent of
Her2/neu. Int J Cancer. 113:678–682. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liu C, Park M and Tsao MS: Overexpression
of c-met proto-oncogene but not epidermal growth factor receptor or
c-erbB-2 in primary human colorectal carcinomas. Oncogene.
7:181–185. 1992.PubMed/NCBI
|
|
31
|
Ramirez R, Hsu D, Patel A, Fenton C,
Dinauer C, Tuttle RM and Francis GL: Over-expression of hepatocyte
growth factor/scatter factor (HGF/SF) and the HGF/SF receptor
(cMET) are associated with a high risk of metastasis and recurrence
for children and young adults with papillary thyroid carcinoma.
Clin Endocrinol (Oxf). 53:635–644. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Rho JK, Choi YJ, Kim SY, Kim TW, Choi EK,
Yoon SJ, Park BM, Park E, Bae JH, Choi CM, et al: MET and AXL
inhibitor NPS-1034 exerts efficacy against lung cancer cells
resistant to EGFR kinase inhibitors because of MET or AXL
activation. Cancer Res. 74:253–262. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tokunou M, Niki T, Eguchi K, Iba S, Tsuda
H, Yamada T, Matsuno Y, Kondo H, Saitoh Y, Imamura H, et al: c-MET
expression in myofibroblasts: Role in autocrine activation and
prognostic significance in lung adenocarcinoma. Am J Pathol.
158:1451–1463. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Tsao MS, Liu N, Chen JR, Pappas J, Ho J,
To C, Viallet J, Park M and Zhu H: Differential expression of
Met/hepatocyte growth factor receptor in subtypes of non-small cell
lung cancers. Lung Cancer. 20:1–16. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ma PC, Jagadeeswaran R, Jagadeesh S,
Tretiakova MS, Nallasura V, Fox EA, Hansen M, Schaefer E, Naoki K,
Lader A, et al: Functional expression and mutations of c-Met and
its therapeutic inhibition with SU11274 and small interfering RNA
in non-small cell lung cancer. Cancer Res. 65:1479–1488. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Wimmel A, Glitz D, Kraus A, Roeder J and
Schuermann M: Axl receptor tyrosine kinase expression in human lung
cancer cell lines correlates with cellular adhesion. Eur J Cancer.
37:2264–2274. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Salian-Mehta S, Xu M and Wierman ME: AXL
and MET crosstalk to promote gonadotropin releasing hormone (GnRH)
neuronal cell migration and survival. Mol Cell Endocrinol.
374:92–100. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Kim HR, Kim WS, Choi YJ, Choi CM, Rho JK
and Lee JC: Epithelial-mesenchymal transition leads to crizotinib
resistance in H2228 lung cancer cells with EML4-ALK translocation.
Mol Oncol. 7:1093–1102. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li Y, Ye X, Tan C, Hongo JA, Zha J, Liu J,
Kallop D, Ludlam MJ and Pei L: Axl as a potential therapeutic
target in cancer: Role of Axl in tumor growth, metastasis and
angiogenesis. Oncogene. 28:3442–3455. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Organ SL and Tsao MS: An overview of the
c-MET signaling pathway. Ther Adv Med Oncol. 3:(Suppl). S7–S19.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sierra JR and Tsao MS: c-MET as a
potential therapeutic target and biomarker in cancer. Ther Adv Med
Oncol. 3:(Suppl). S21–S35. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Gujral TS, Karp RL, Finski A, Chan M,
Schwartz PE, MacBeath G and Sorger P: Profiling phospho-signaling
networks in breast cancer using reverse-phase protein arrays.
Oncogene. 32:3470–3476. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Folkman J: Tumor angiogenesis: Therapeutic
implications. N Engl J Med. 285:1182–1186. 1971. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Freudlsperger C, Alexander D, Reinert S
and Hoffmann J: Prognostic value of c-Met expression in oral
squamous cell carcinoma. Exp Ther Med. 1:69–72. 2010.PubMed/NCBI
|
|
45
|
Gisterek I, Lata E, Halon A, Matkowski R,
Szelachowska J, Biecek P and Kornafel J: Prognostic role of c-met
expression in breast cancer patients. Rep Pract Oncol Radiother.
16:173–177. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Ishikawa M, Sonobe M, Nakayama E,
Kobayashi M, Kikuchi R, Kitamura J, Imamura N and Date H: Higher
expression of receptor tyrosine kinase Axl, and differential
expression of its ligand, Gas6, predict poor survival in lung
adenocarcinoma patients. Ann Surg Oncol. 20:(Suppl 3). S467–S476.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Lee CH, Yen CY, Liu SY, Chen CK, Chiang
CF, Shiah SG, Chen PH and Shieh YS: Axl is a prognostic marker in
oral squamous cell carcinoma. Ann Surg Oncol. 19:(Suppl 3).
S500–S508. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Pinato DJ, Mauri FA, Lloyd T, Vaira V,
Casadio C, Boldorini RL and Sharma R: The expression of Axl
receptor tyrosine kinase influences the tumour phenotype and
clinical outcome of patients with malignant pleural mesothelioma.
Br J Cancer. 108:621–628. 2013. View Article : Google Scholar : PubMed/NCBI
|